Terpenoids from Weedy Ricefield Flatsedge (Cyperus iria L.) Are Developmentally Regulated and Stress-Induced, and have Antifungal Properties

Abstract
:1. Introduction
2. Results and Discussion
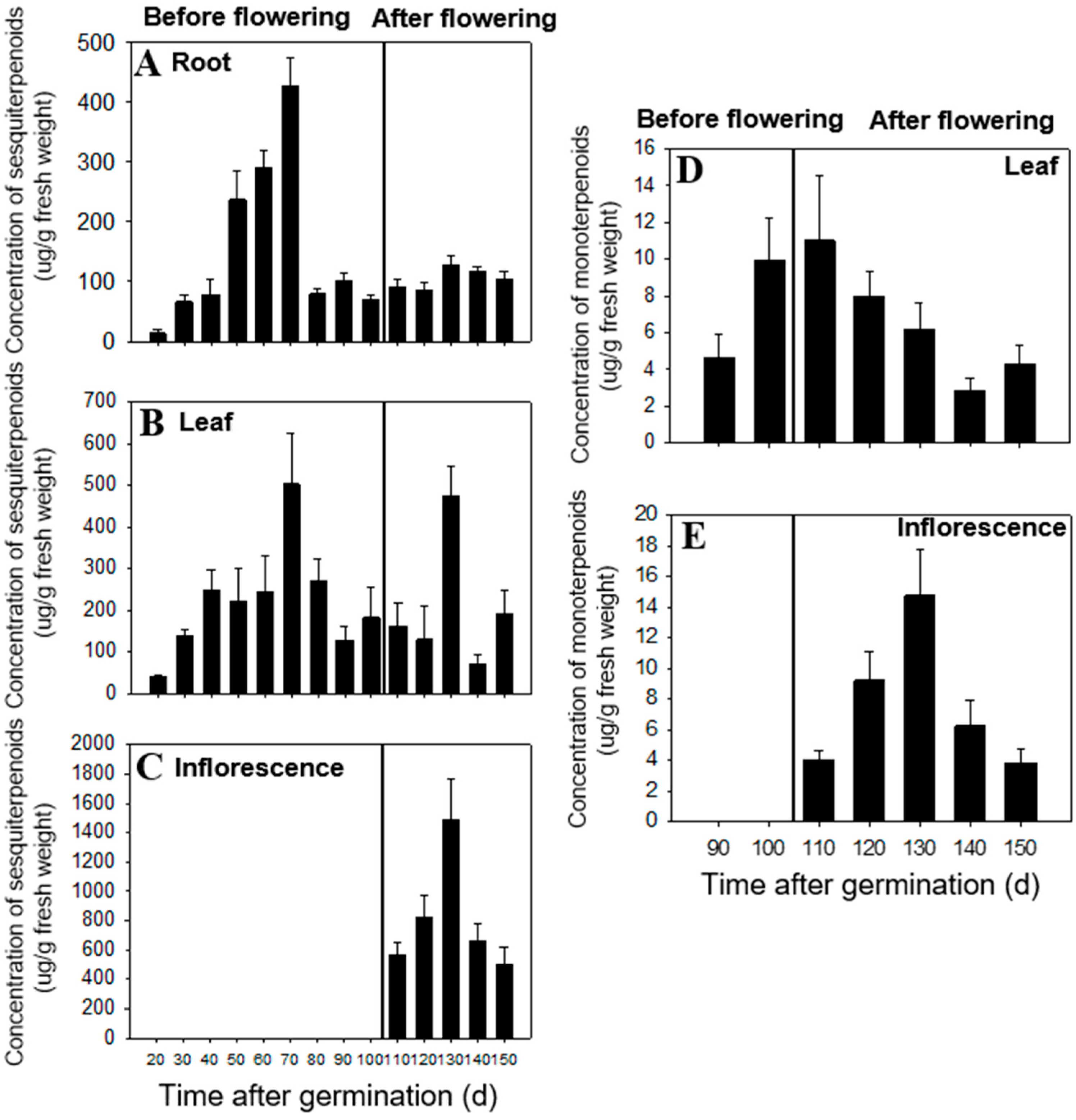
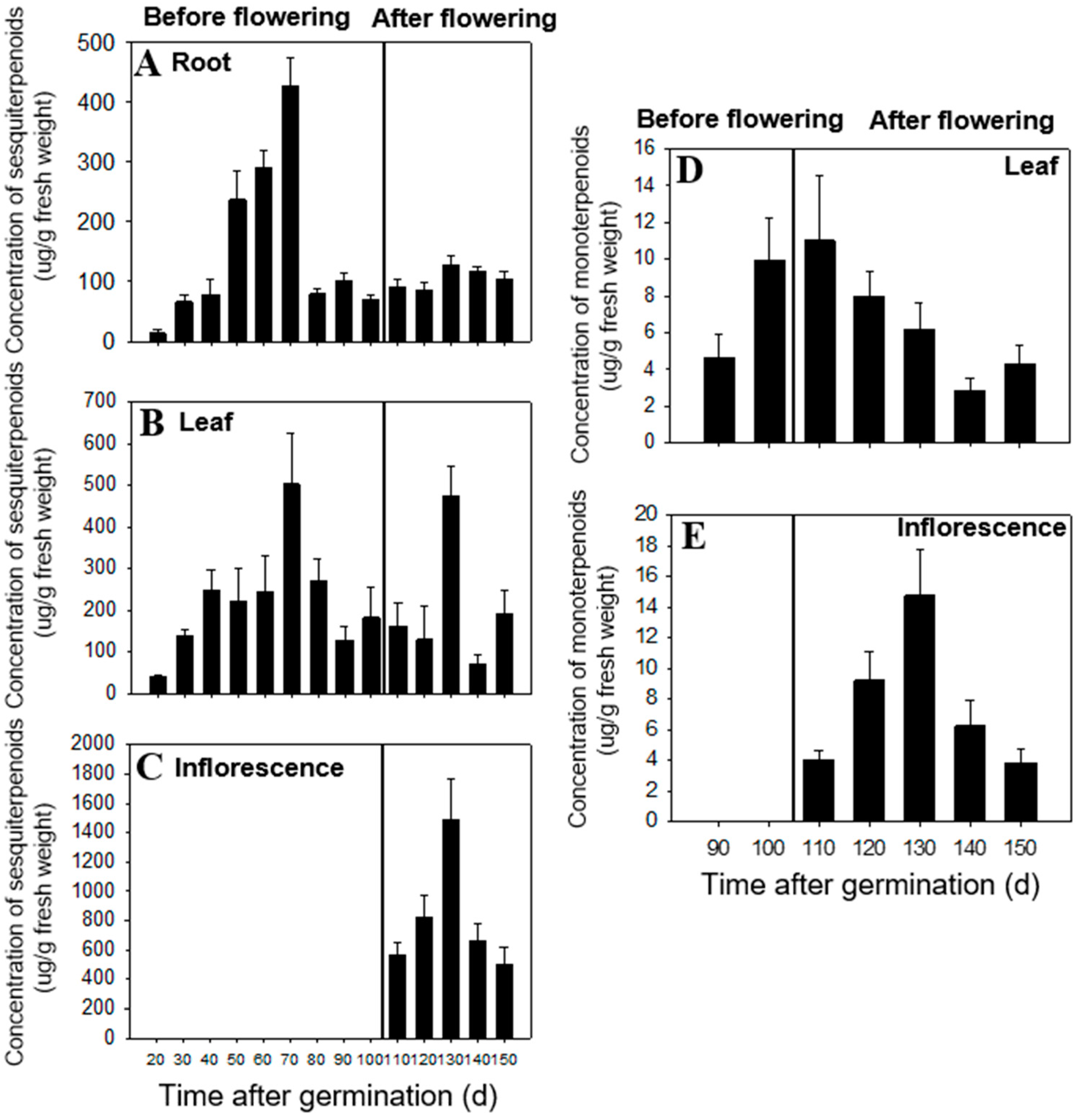
2.1. Temporal and Spatial Differences in Terpenoid Content
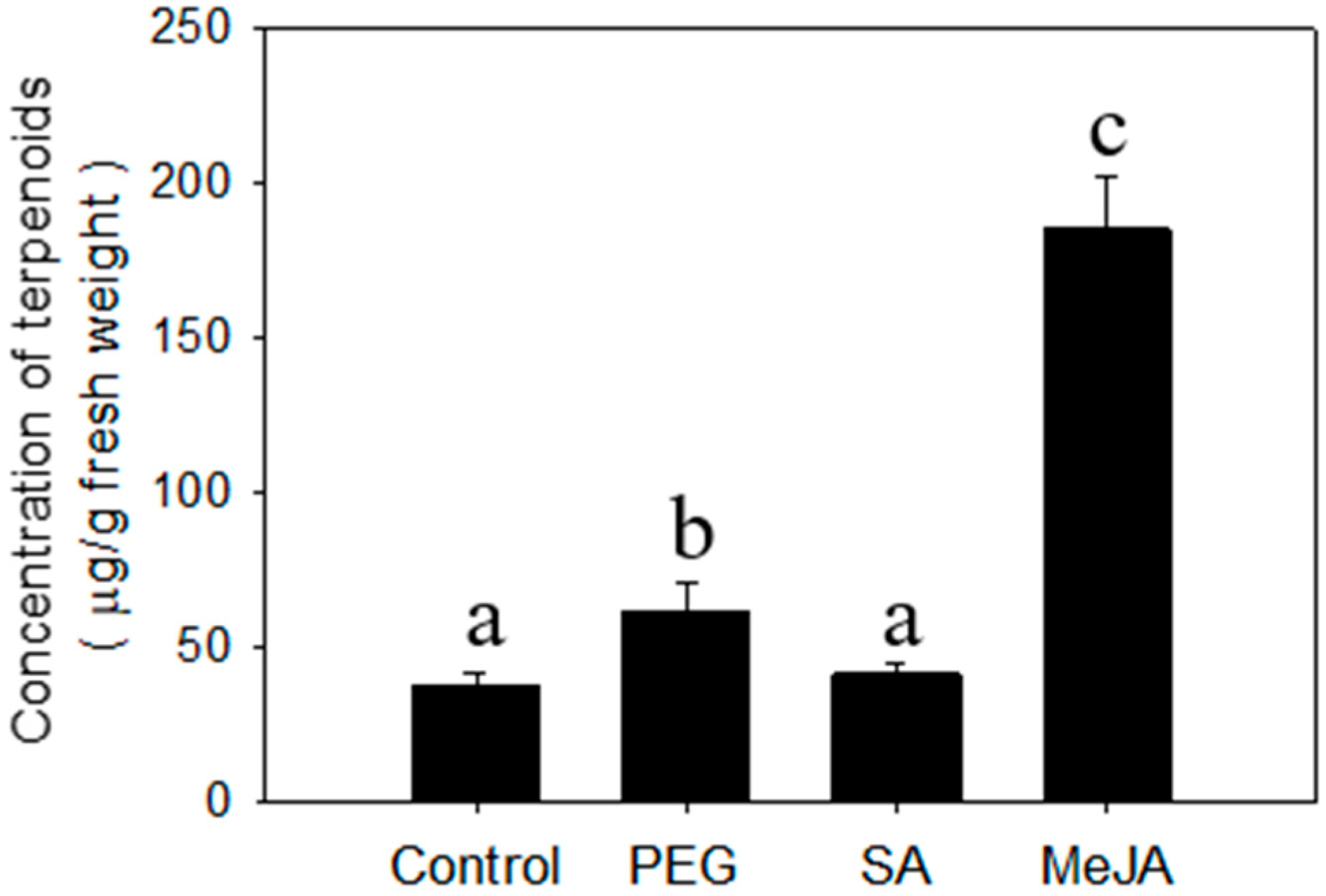
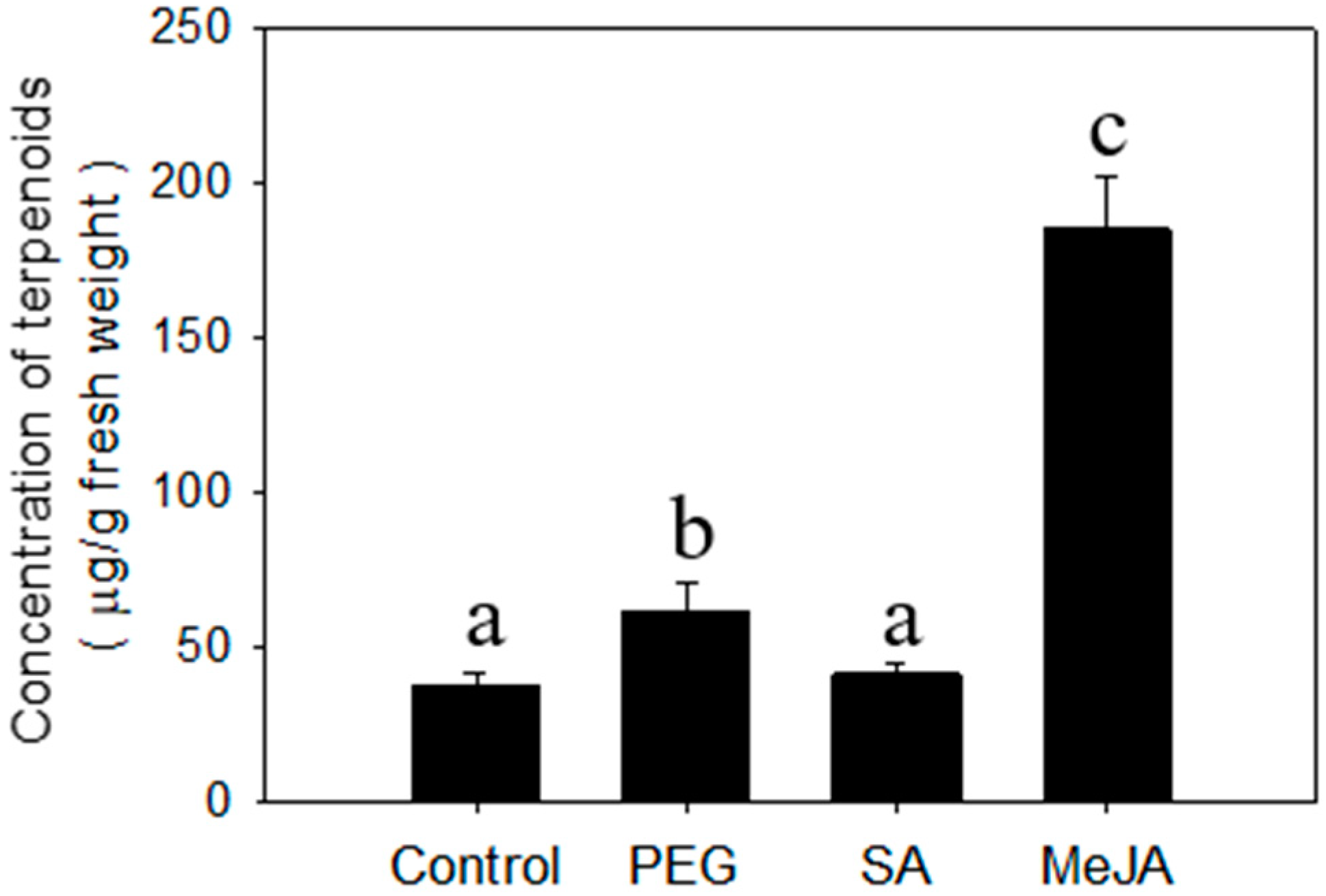
2.2. Regulation of Terpenoid Accumulation by Abiotic Stress
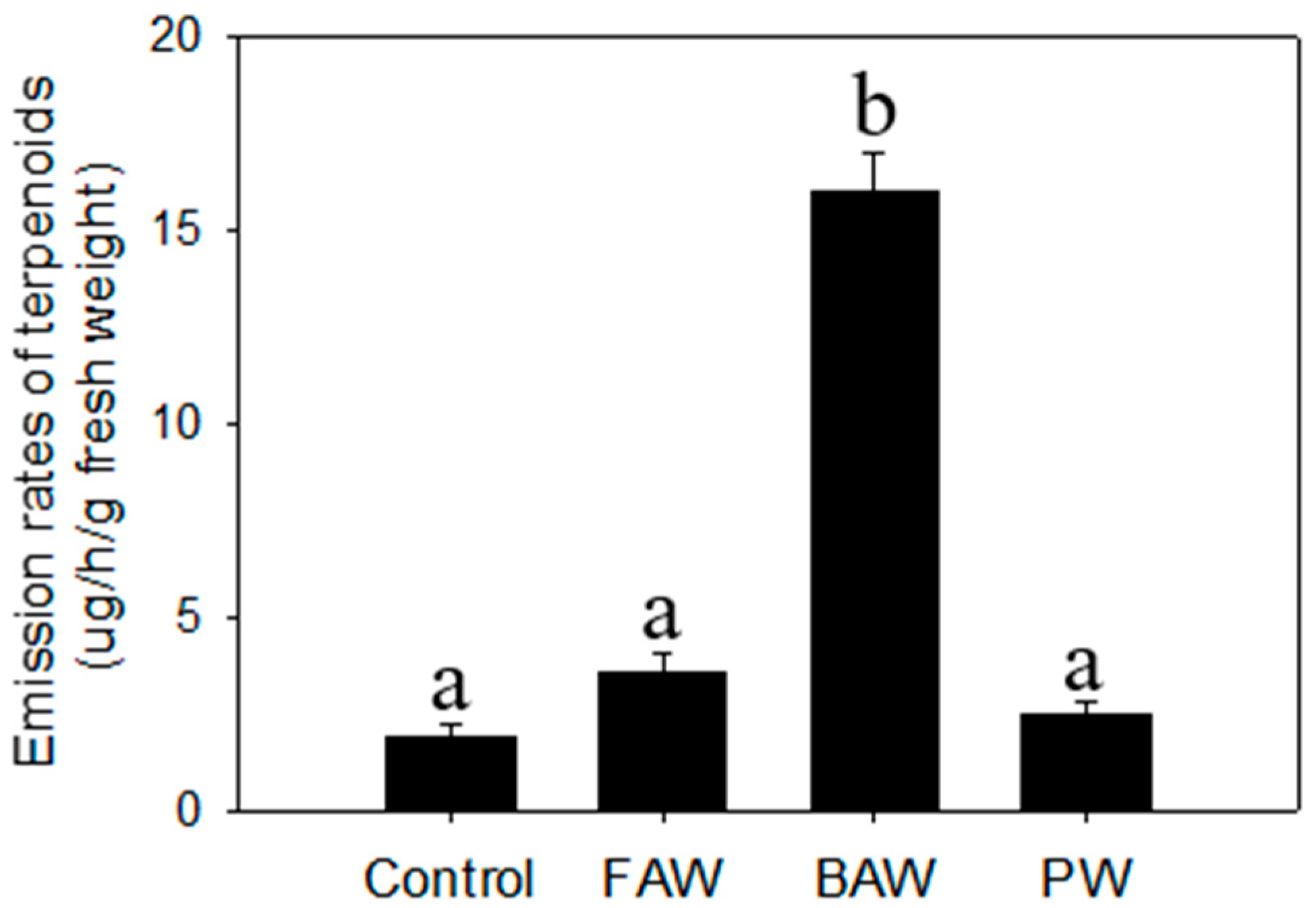
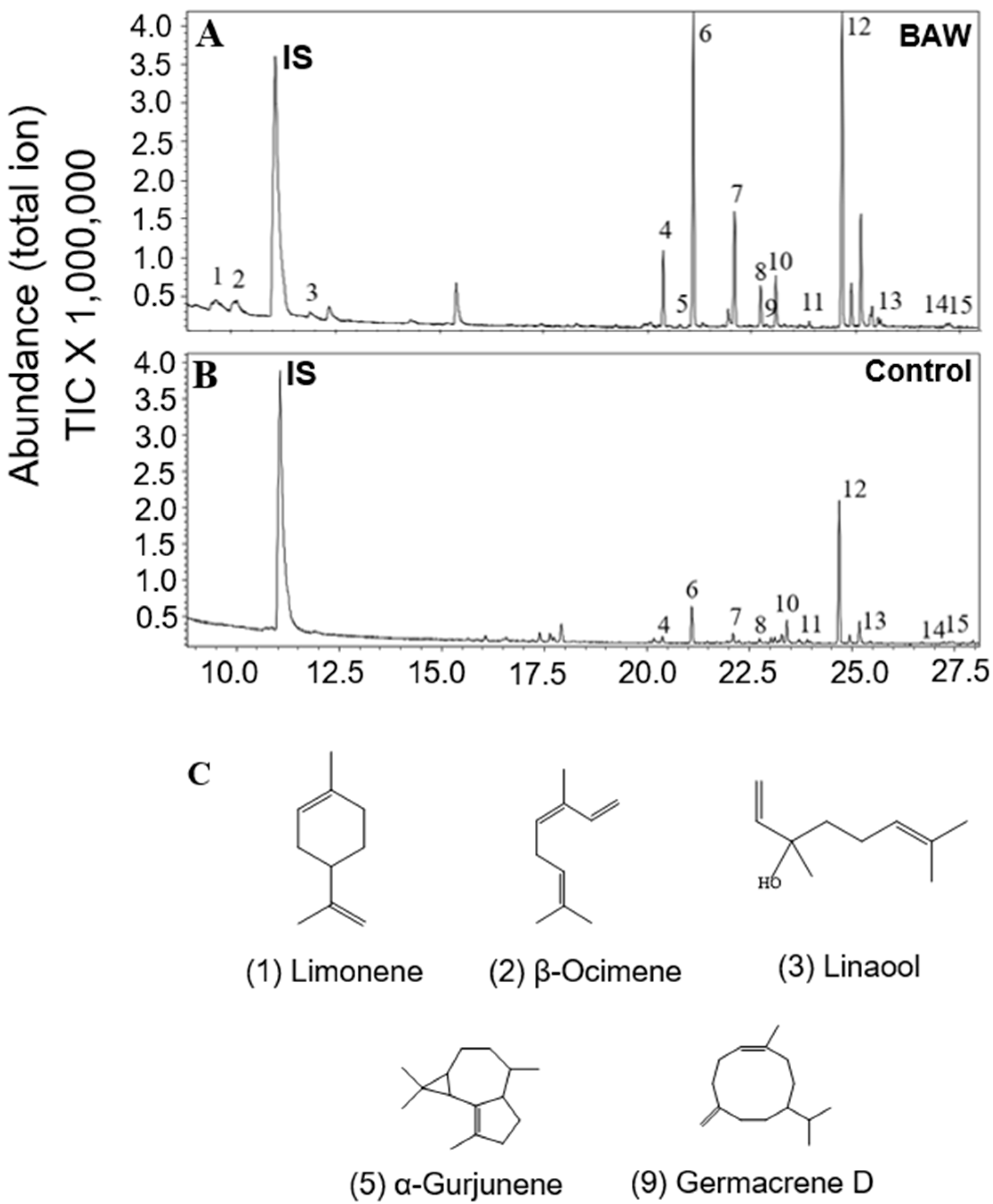
2.3. Regulation of Terpenoid Accumulation by Biotic Stress
2.4. Cluster Analysis of Sesquiterpenoids after Different Treatments
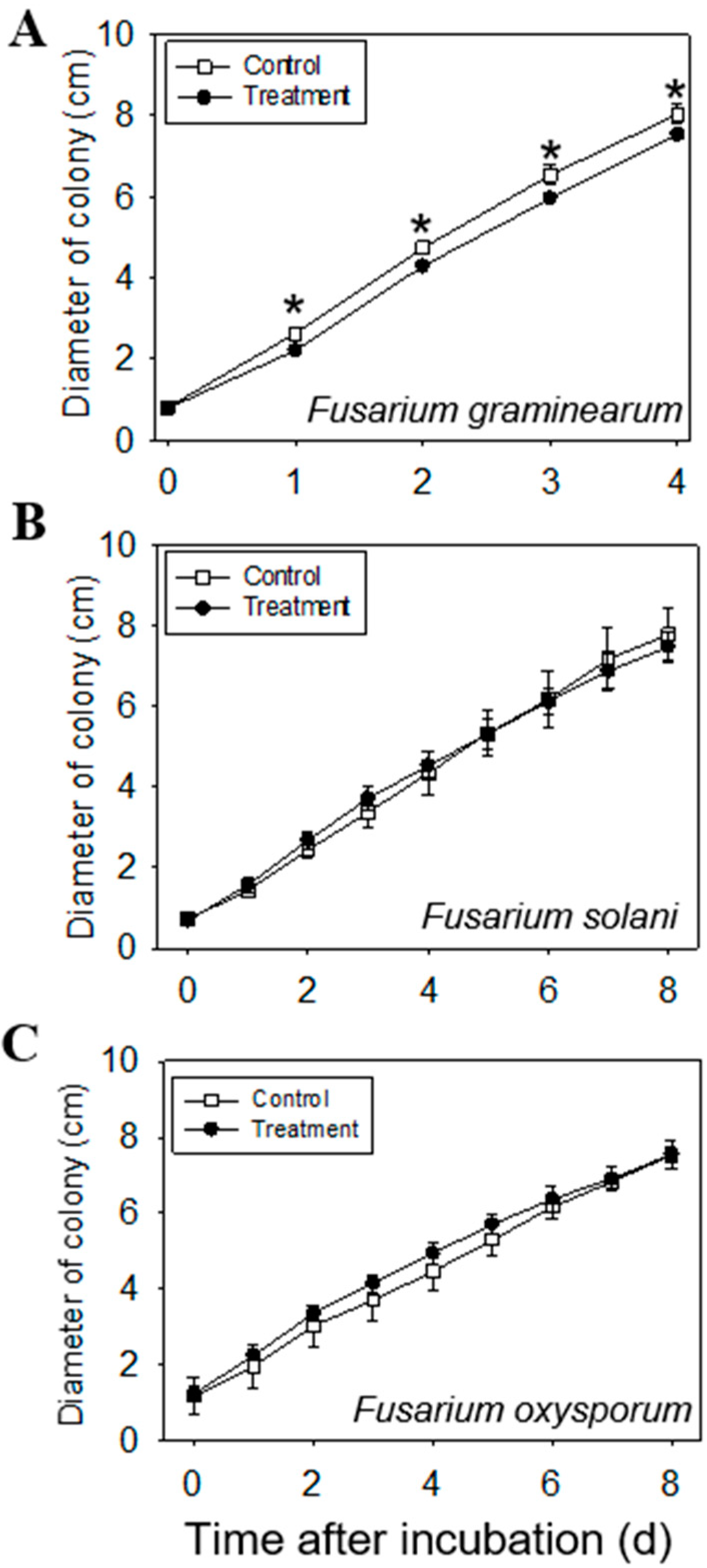
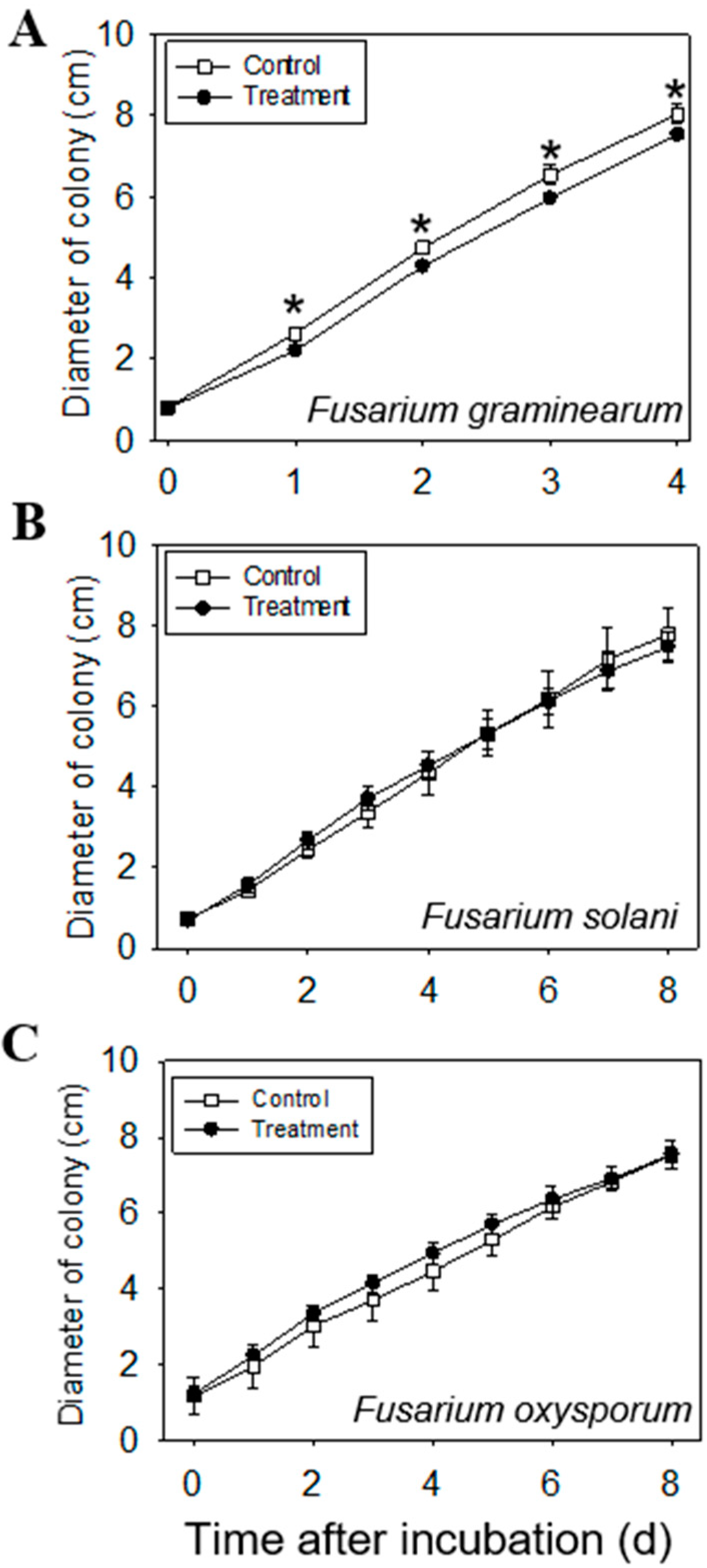
2.5. Anti-Fungal Effect of Volatiles Emitted from Leaves
3. Materials and Methods
3.1. Plant Material
3.2. Characterization of Terpenoids with Organic Extraction
3.3. GC-MS Analysis
3.4. Temporal and Spatial Distribution of Terpenoids
3.5. Chemical Treatments
3.6. Insect Infestation and Physical Wounding
3.7. Headspace-Collection of Volatiles by Insect Infestation and Physical Wounding
3.8. Anti-Fungal Effect of Volatiles Emitted from Leaves
3.9. Hierarchical Clustering and Data Visualization
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Manandhar, S.; Shrestha, B.B.; Lekhak, H.D. Weeds of paddy field at Kirtipur, Kathmandu. Sci. World J. 2007, 5, 100–106. [Google Scholar] [CrossRef]
- Kelton, J.; Price, A.J.; Mosjidis, J. Allelopathic Weed Suppression through the Use of Cover Crops. In Weed Control; Price, A.J., Ed.; Intech Press: Rijeka, Croatia, 2012; pp. 115–130. [Google Scholar]
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of specialized metabolites in plants: An evolutionary perspective. Trends Plant Sci. 2000, 5, 439–445. [Google Scholar] [CrossRef]
- Pichersky, E.; Sharkey, T.D.; Gershenzon, J. Plant volatiles: A lack of function or a lack of knowledge? Trends Plant Sci. 2006, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- McCaskill, D.; Croteau, R. Prospects for the bioengineering of isoprenoid biosynthesis. Adv. Biochem. Eng. Biotechnol. 1997, 55, 107–146. [Google Scholar] [PubMed]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and antifungal activity of aromatic constituents of essential oils. Microbios 1997, 89, 39–46. [Google Scholar] [PubMed]
- Scortichini, M.; Rossi, M.P. Preliminary in vitro evaluation of the antimicrobial activity of terpenes and terpenoids towards Erwinia amylovora (Burrill). J. Appl. Bacteriol. 2010, 71, 109–112. [Google Scholar] [CrossRef]
- Griffin, S.G.; Wyllie, S.G.; Markham, J.L.; Leach, D.N. The role of structure and molecular properties of terpenoids in determining their antimicrobial activity. Flavour Fragr. J. 2015, 14, 322–332. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Johnson, D.E. Ecological studies on Cyperus difformis, Cyperus iria and Fimbristylis miliacea: Three troublesome annual sedge weeds of rice. Ann. Appl. Biol. 2009, 155, 103–112. [Google Scholar] [CrossRef]
- Lawal, O.A.; Oyedeji, A.O. Chemical composition of the essential oils of Cyperus rotundus L. from South Africa. Molecules 2009, 14, 2909–2917. [Google Scholar] [CrossRef] [PubMed]
- Komai, K.; Tang, C.-S. Chemical constituents and inhibitory activities of essential oils from Cyperus brevifolius and C. kyllingia. J. Chem. Ecol. 1989, 15, 2171–2176. [Google Scholar] [CrossRef] [PubMed]
- Kubmarawa, D.; Ogunwande, I.A.; Okorie, D.A.; Olawore, N.O.; Kasali, A.A. Chemical constituents of the volatile oil of Cyperus esculentus L. from Nigeria. Flavour Fragr. J. 2005, 20, 640–641. [Google Scholar] [CrossRef]
- Gilbert, L.I.; Rybczynski, R.; Tobe, S.S. Endocrine Cascade in Insect Metamorphosis. In Metamorphosis; Gilbert, L.I., Tata, J.R., Atkinson, B.G., Eds.; Academic Press: New York, NY, USA, 1996; pp. 59–107. [Google Scholar]
- Chopra, N.; Tewari, G.; Tewari, L.M.; Upreti, B.; Pandey, N. Allelopathic effect of Echinochloa colona L. and Cyperus iria L. weed extracts on the seed germination and seedling growth of rice and soyabean. Adv. Agric. 2017, 4, 1–5. [Google Scholar] [CrossRef]
- Toong, Y.C.; Schooley, D.A.; Baker, F.C. Isolation of insect juvenile hormone III from a plant. Nature 1988, 333, 170–171. [Google Scholar] [CrossRef]
- Bede, J.C.; Teal, P.E.; Goodman, W.G.; Tobe, S.S. Biosynthetic pathway of insect juvenile hormone III in cell suspension cultures of the sedge Cyperus iria. Plant Physiol. 2001, 127, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Bede, J.C.; Goodman, W.G.; Tobe, S.S. Developmental distribution of insect juvenile hormone III in the sedge, Cyperus iria L. Phytochemistry 1999, 52, 1269–1274. [Google Scholar] [CrossRef]
- Yuan, J.S.; Köllner, T.G.; Wiggins, G.; Grant, J.; Degenhardt, J.; Chen, F. Molecular and genomic basis of volatile-mediated indirect defense against insects in rice. Plant J. 2008, 55, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Köllner, T.G.; Zhao, N.; Li, G.; Jiang, Y.; Zhu, L.; Ma, J.; Degenhardt, J.; Chen, F. Dynamic evolution of herbivore-induced sesquiterpene biosynthesis in sorghum and related grass crops. Plant J. 2012, 69, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Köllner, T.G.; Gershenzon, J.; Degenhardt, J. Molecular and biochemical evolution of maize terpene synthase 10, an enzyme of indirect defense. Phytochemistry. 2009, 70, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Köllner, T.G.; Held, M.; Lenk, C.; Hiltpold, I.; Turlings, T.C.; Gershenzon, J.; Degenhardta, J. A maize (E)-beta-caryophyllene synthase implicated in indirect defense responses against herbivores is not expressed in most American maize varieties. Plant Cell 2008, 20, 482–494. [Google Scholar]
- Jiang, Y.; Ye, J.; Li, S.; Niinemets, Ü. Methyl jasmonate-induced emission of biogenic volatiles is biphasic in cucumber: A high-resolution analysis of dose dependence. J. Exp. Bot. 2017, 68, 4679–4694. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Al-Ahmad, H.; Joyce, B.; Zhao, N.; Köllner, T.G.; Degenhardt, J.; Stewart, C.N. Within-plant distribution and emission of sesquiterpenes from Copaifera officinalis. Plant Physiol. Biochem. 2009, 47, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ye, J.; Li, S.; Niinemets, Ü. Regulation of floral terpenoid emission and biosynthesis in sweet basil (Ocimum basilicum). J. Plant Growth Regul. 2016, 35, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Núñez, Y.O.; Salabarria, I.S.; Collado, I.G.; Hernández-Galán, R. Antifungal activity of extracts and terpene constituents of aerial parts of Juniperus lucayana. Revista Latinoamericana de Quimica 2010, 38, 145–152. [Google Scholar]
- Wiemann, P.; Sieber, C.M.; von Bargen, K.W.; Studt, L.; Niehaus, E.M.; Espino, J.J.; Huß, K.; Michielse, C.B.; Albermann, S.; Wagner, D.; et al. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathog. 2013, 9, 6–27. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in planta mycotoxin accumulation among members of the Fusarium graminearum species complex on wheat and rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Razdan, V.K.; John, D.; Sharma, B.C. First report of leaf blight of Cyperus iria caused by Fusarium equiseti in India. Plant Dis. 2013, 97, 838. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; D’Auria, J.C.; Farooq, A.; Pichersky, E.; Gershenzon, J. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. Plant Cell 2003, 15, 481–494. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Control L(O) a | PEG L(O) | SA L(O) | MeJA L(O) | Control L(H) b | FAW L(H) | BAW L(H) | PW L(H) |
|---|---|---|---|---|---|---|---|---|
| Concentration (µg/g Fresh Weight) | Emission Rates (μg/h/g Fresh Weight) | |||||||
| Monoterpene | ||||||||
| Limonene | ND c | ND | ND | ND | ND | ND | 0.15 ± 0.02 | ND |
| β-Ocimene | ND | ND | ND | ND | ND | ND | 0.31 ± 0.06 | ND |
| Linalool | ND | ND | ND | 0.94 ± 0.17 | ND | ND | 0.12 ± 0.01 | ND |
| Sesquiterpene | ||||||||
| β-Elemene * | 1.2 d ± 0.2a e | 2.0 ± 0.2a | 1.5 ± 0.2a | 6.8 ± 0.8b | 0.051 ± 0.009a | 0.21 ± 0.02a | 1.1 ± 0.1b | 0.11 ± 0.03a |
| α-Gurjunene | ND | ND | ND | ND | ND | ND | 0.37 ± 0.05 | ND |
| E-β-Caryophyllene * | 6.7 ± 0.8a | 13 ± 1b | 8.9± 1.1a | 39 ± 3c | 0.45 ± 0.08a | 1.2 ± 0.1b | 6.7 ± 0.5c | 0.88 ± 0.13b |
| α-Humulene * | 1.7 ± 0.2a | 3.6 ± 0.6b | 2.4 ± 0.4a | 10± 1c | 0.084 ± 0.016a | 0.33 ± 0.05a | 1.7 ± 0.1b | 0.13 ± 0.03a |
| α-Bergamotene | 0.75 ± 0.12a | 1.7 ± 0.2b | 0.94 ± 0.08a | 4.2 ± 0.7c | 0.031 ± 0.007a | 0.15 ± 0.02a | 0.57 ± 0.04c | 0.22 ± 0.04b |
| Germacrene D | ND | ND | ND | ND | ND | ND | 0.038 ± 0.006 | ND |
| α-Farnesene * | 1.3 ± 0.2a | 2.4 ± 0.4b | 1.4 ± 0.3a | 5.5 ± 0.4c | 0.14 ± 0.02a | 0.12 ± 0.02a | 0.69 ± 0.09b | 0.52 ± 0.06b |
| Elemol | 0.59 ± 0.04a | 0.91 ± 0.14b | 0.56 ± 0.04a | 2.4 ± 0.3c | 0.032 ± 0.008a | 0.042 ± 0.002a | 0.075 ± 0.007a | 0.091 ± 0.019a |
| Nerolidol | ND | ND | ND | 2.73 ± 0.43 | ND | ND | ND | ND |
| Hedycaryol | 24 ± 4a | 34 ± 3b | 25 ± 4a | 107 ± 11c | 1.1 ± 0.1b | 1.4 ± 0.2b | 4.8 ± 0.6c | 0.11 ± 0.02a |
| γ-Eudesmol | 0.14 ± 0.02a | 0.72 ± 0.08b | 0.17 ± 0.04a | 1.1 ± 0.2c | 0.011 ± 0.002a | 0.074 ± 0.010b | 0.045 ± 0.004b | 0.015 ± 0.003a |
| β-Eudesmol | 0.25 ± 0.03a | 1.2 ± 0.3a | 0.34 ± 0.04a | 2.4 ± 0.2b | 0.013 ± 0.001a | 0.13 ± 0.02c | 0.045 ± 0.007b | 0.030 ± 0.002b |
| α-Eudesmol | 0.27 ± 0.04a | 1.9 ± 0.3b | 0.41 ± 0.05a | 2.2 ± 0.2b | 0.015 ± 0.004a | 0.073 ± 0.010b | 0.071 ± 0.006b | 0.083 ± 0.013b |
| Total | 37 ± 4a | 61± 9b | 41 ± 3a | 185 ± 17c | 1.9 ± 0.3a | 3.6 ± 0.5a | 16 ± 1b | 2.5 ± 0.3a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Ownley, B.H.; Chen, F. Terpenoids from Weedy Ricefield Flatsedge (Cyperus iria L.) Are Developmentally Regulated and Stress-Induced, and have Antifungal Properties. Molecules 2018, 23, 3149. https://doi.org/10.3390/molecules23123149
Jiang Y, Ownley BH, Chen F. Terpenoids from Weedy Ricefield Flatsedge (Cyperus iria L.) Are Developmentally Regulated and Stress-Induced, and have Antifungal Properties. Molecules. 2018; 23(12):3149. https://doi.org/10.3390/molecules23123149
Chicago/Turabian StyleJiang, Yifan, Bonnie H. Ownley, and Feng Chen. 2018. "Terpenoids from Weedy Ricefield Flatsedge (Cyperus iria L.) Are Developmentally Regulated and Stress-Induced, and have Antifungal Properties" Molecules 23, no. 12: 3149. https://doi.org/10.3390/molecules23123149
APA StyleJiang, Y., Ownley, B. H., & Chen, F. (2018). Terpenoids from Weedy Ricefield Flatsedge (Cyperus iria L.) Are Developmentally Regulated and Stress-Induced, and have Antifungal Properties. Molecules, 23(12), 3149. https://doi.org/10.3390/molecules23123149