A Metabolomic and HPLC-MS/MS Analysis of the Foliar Phenolics, Flavonoids and Coumarins of the Fraxinus Species Resistant and Susceptible to Emerald Ash Borer

Abstract
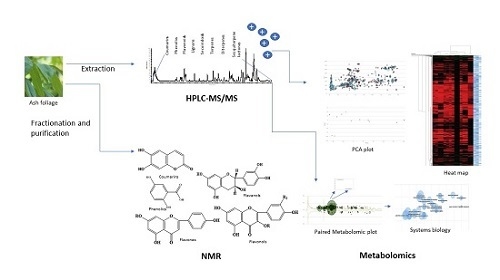
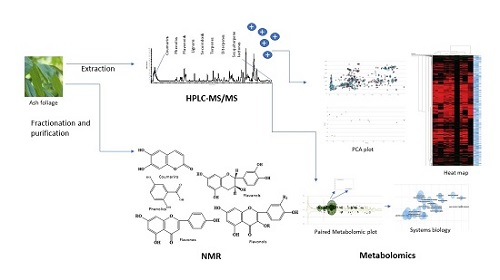
1. Introduction
2. Results and Discussion
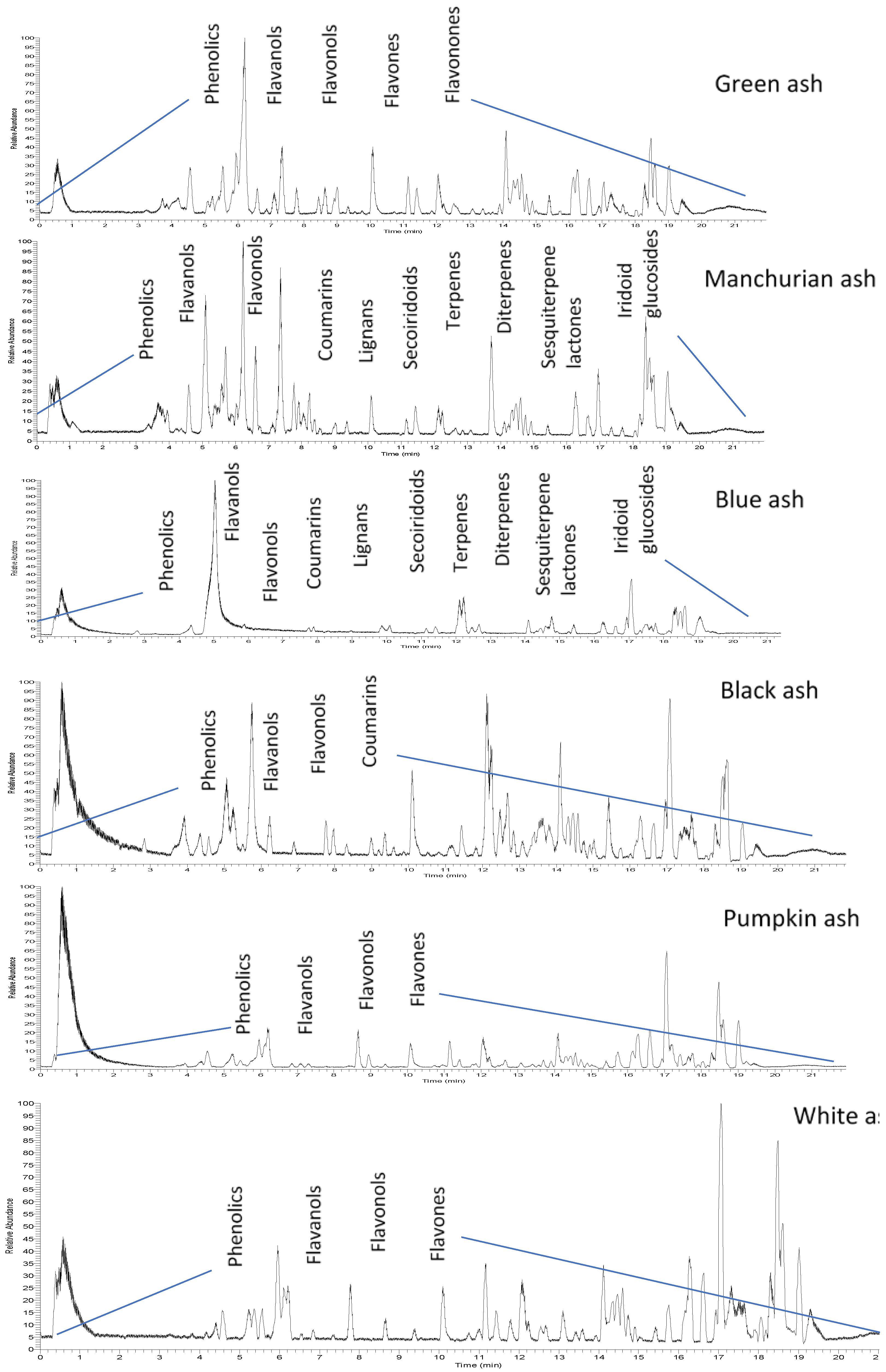
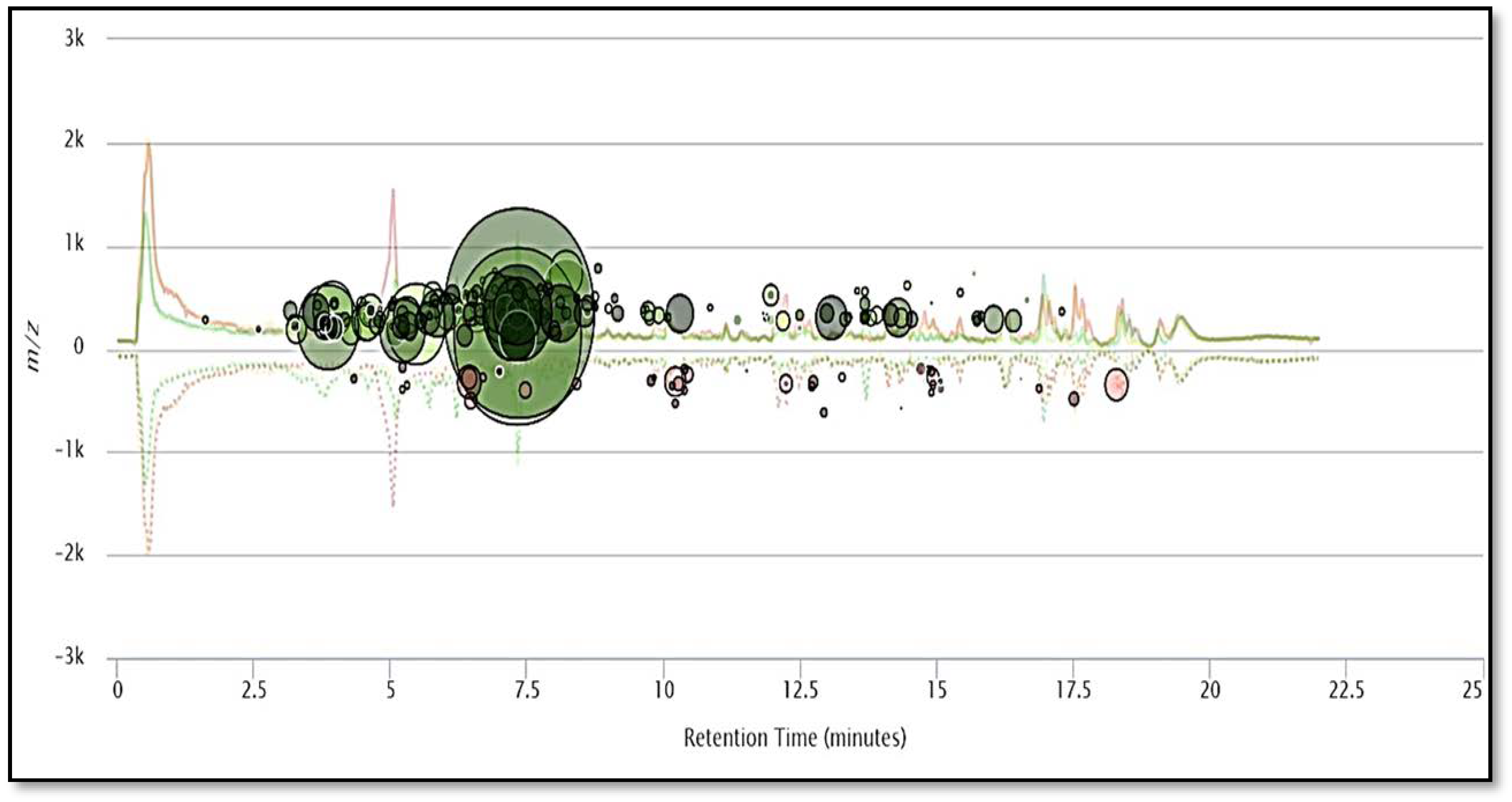
2.1. HPLC-MS/MS Orbitrap Analysis
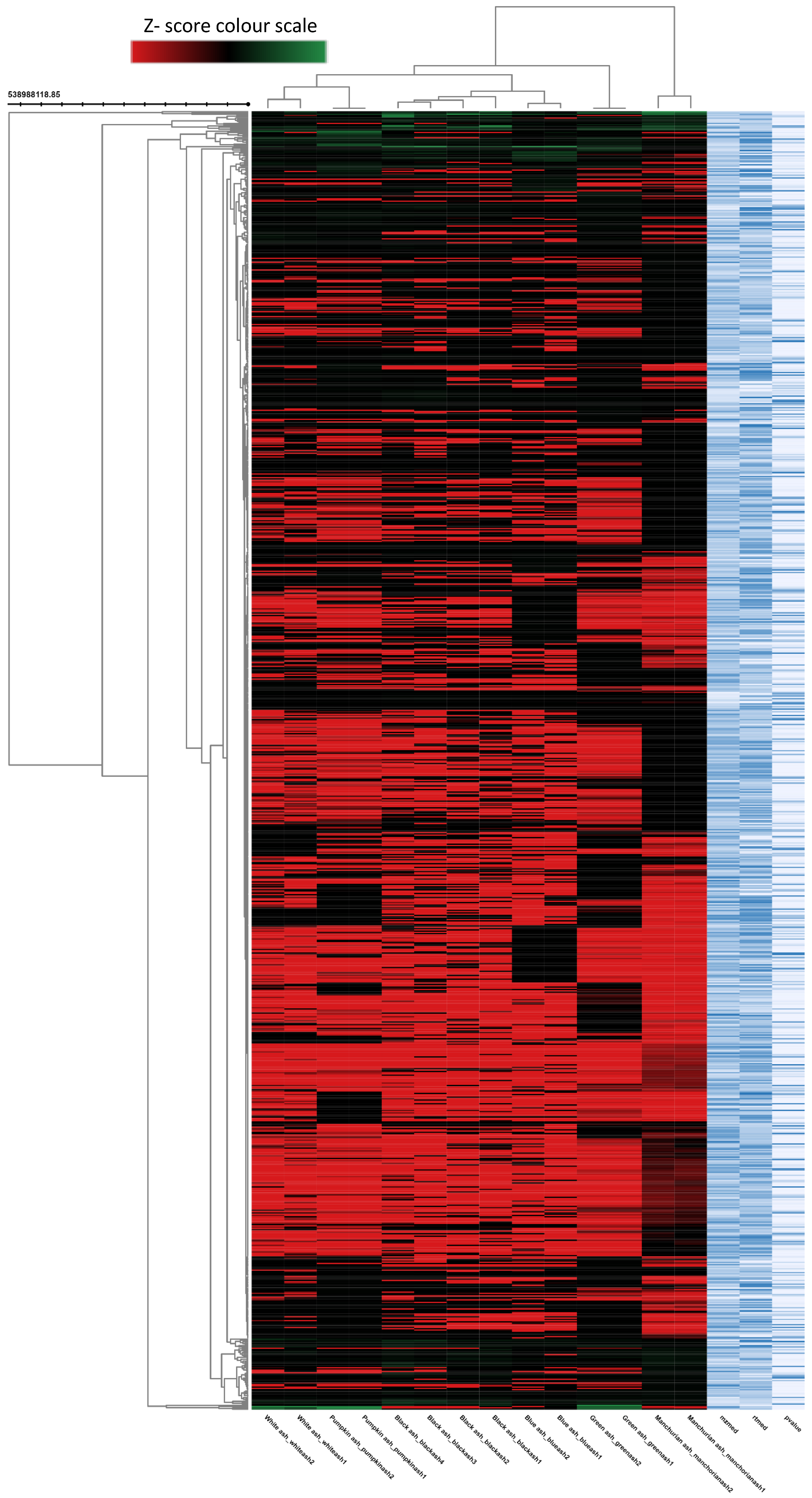
2.2. Metabolomic Analysis of Fraxinus spp.
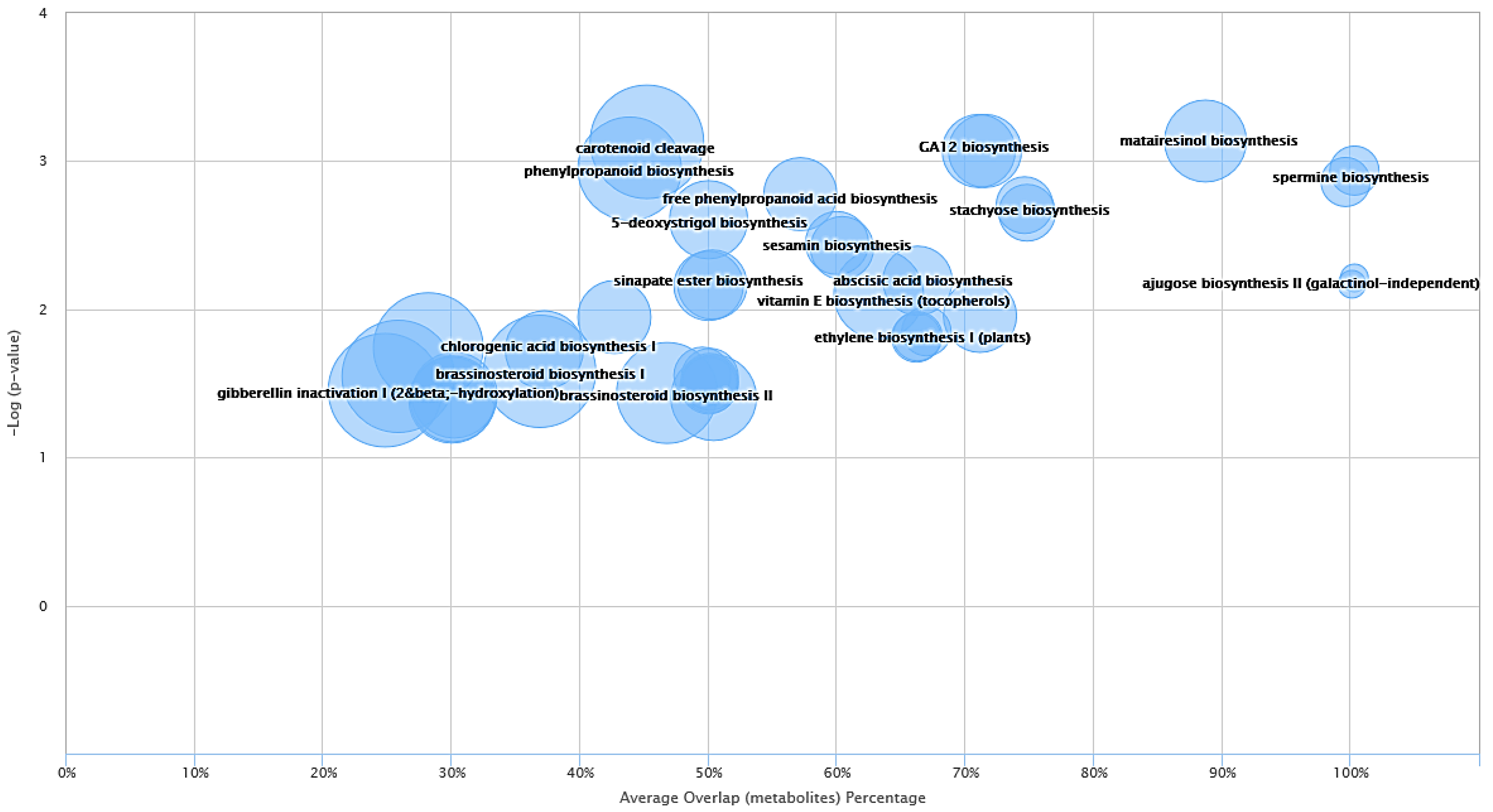
2.3. System Biology
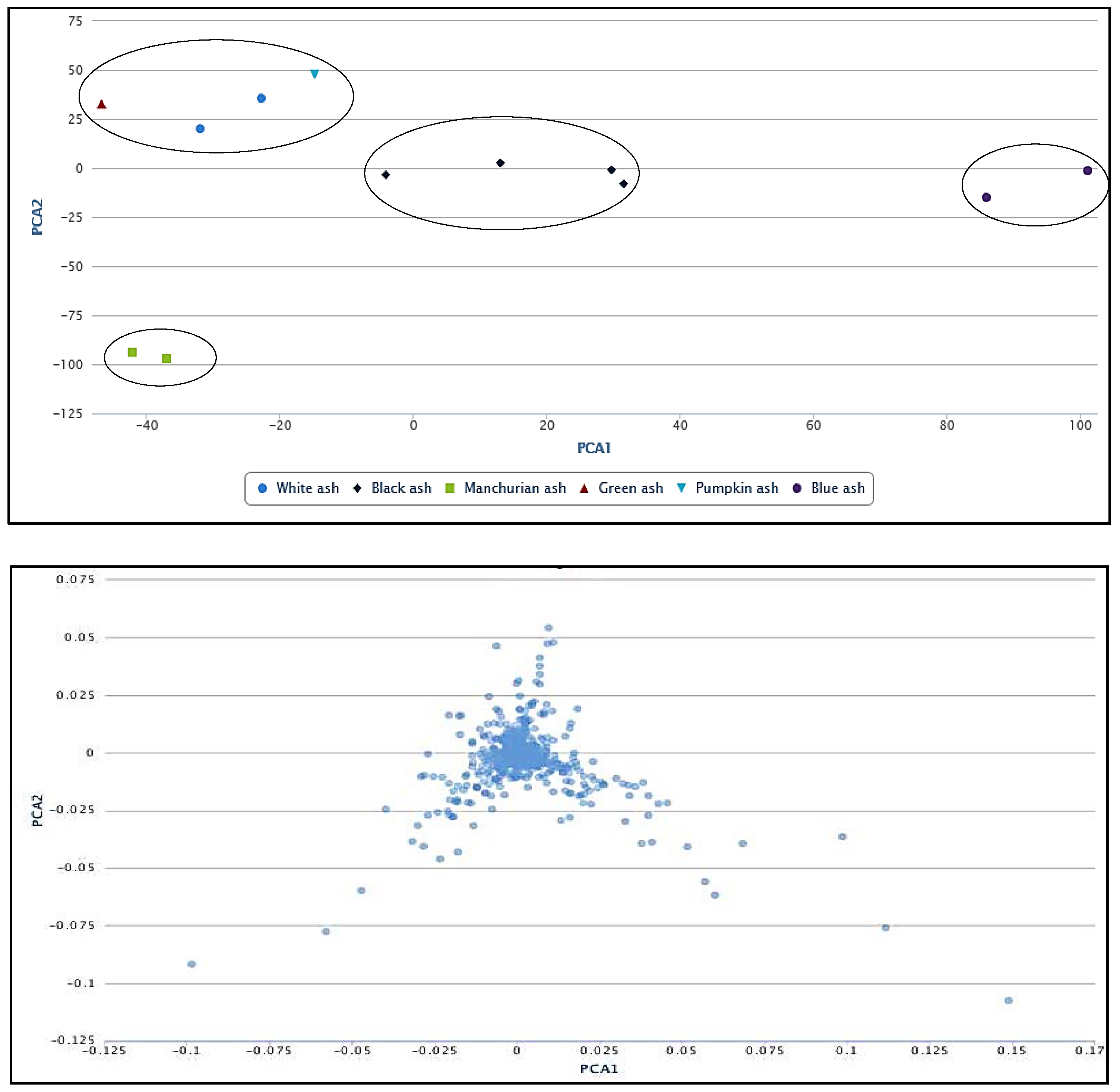
2.4. Principal Component Analysis (PCA)
3. Methods and Materials
3.1. Plants
3.2. Extractions
3.3. Fractionation
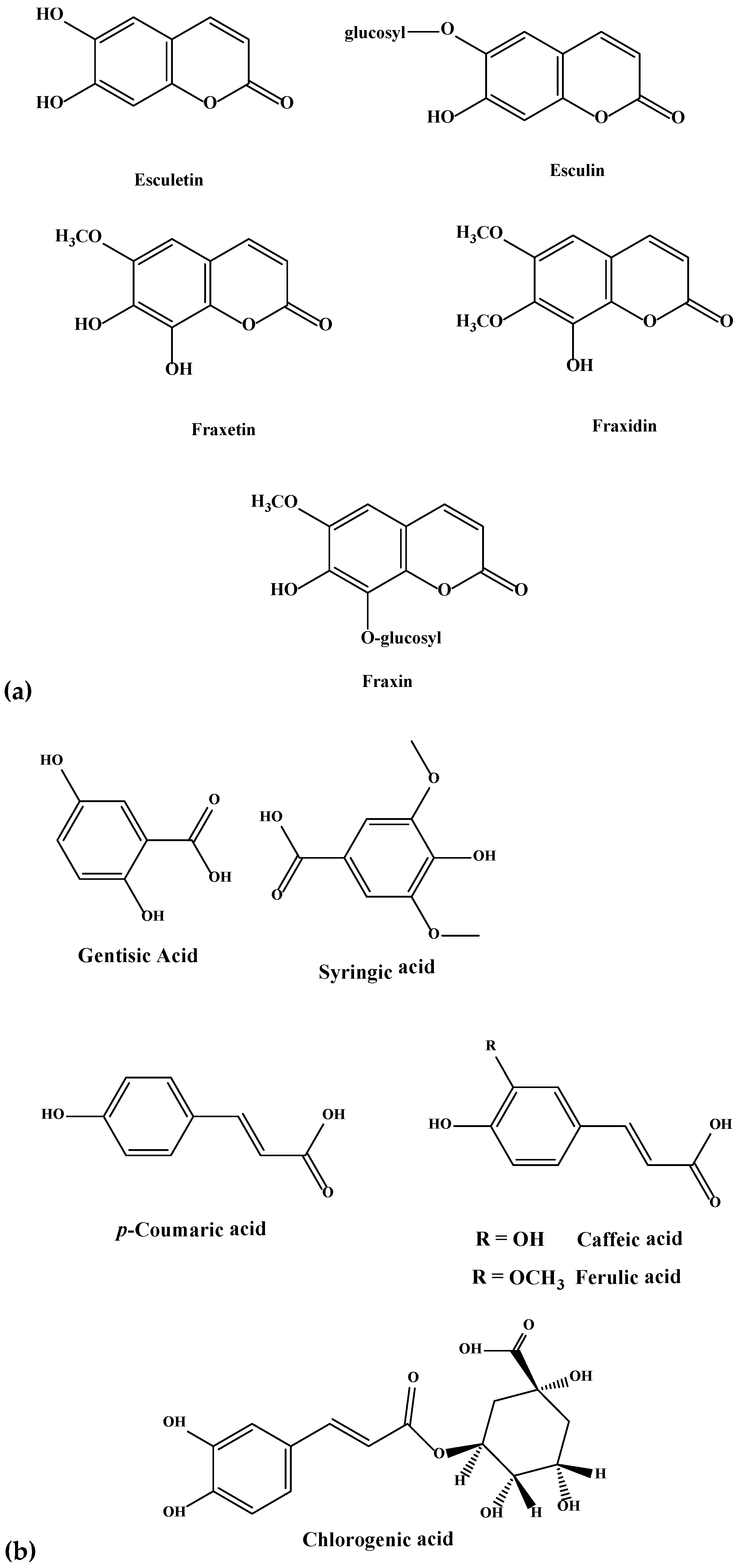
3.4. Identification of Isolated Compounds
3.5. Preparation of Extracts for HPLC-MS/MS
3.6. HPLC-MS/MS (ORBITRAP)
3.7. Untargeted Metabolomic Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Poland, T.M.; McCullough, D.G. Emerald ash borer: Invasion of the urban forest and the threat to North America’s ash resource. J. For. 2006, 104, 118–124. [Google Scholar]
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.M.; Telewski, F.D. Dendrochronological reconstruction of the epicentre and early spread of Emerald Ash Borer in North America. Divers. Distrib. 2014, 20, 847–858. [Google Scholar] [CrossRef]
- Farrar, J.L. Trees in Canada; Natural Resources Canada, Canadian Forest Service: Ottawa, ON, Canada; Fitzhenry and Whiteside Limited: Markham, ON, Canada, 1995. [Google Scholar]
- Cappaert, D.; McCullough, D.G.; Poland, T.M.; Siegert, N.W. Emerald ash borer in North America: A research and regulatory challenge. Am. Entomol. 2005, 51, 152–165. [Google Scholar] [CrossRef]
- Rebek, E.J.; Herms, D.A.; Smitley, D.R. Interspecific variation in resistance to Emerald Ash Borer (Coleoptera: Buprestidae) among North American and Asian ash (Fraxinus spp.). Environ. Entomol. 2008, 37, 242–246. [Google Scholar] [CrossRef]
- Klooster, W.S.; Herms, D.A.; Knight, K.S.; Herms, C.P.; McCullough, D.G.; Smith, A.; Gandhi, K.J.K.; Cardina, J. Ash (Fraxinus spp.) mortality, regeneration, and seed bank dynamics in mixed hardwood forests following invasion by Emerald Ash Borer (Agrilus planipennis). Biol. Invasions 2014, 16, 859–873. [Google Scholar] [CrossRef]
- Anulewicz, A.C.; Mccullough, D.G.; Cappaert, D.L.; Poland, T.M. Host range of the Emerald Ash Borer (Agrilus planipennis Fairmaire) (Coleoptera: Buprestidae) in North America: Results of multiple-choice field experiments. Environ. Entomol. 2008, 37, 230–241. [Google Scholar] [CrossRef]
- Tanis, S.R.; McCullough, D.G. Differential persistence of blue ash and white ash following Emerald Ash Borer invasion. Can. J. For. Res. 2012, 42, 1542–1550. [Google Scholar] [CrossRef]
- Westwood, M.; Oldfield, S.; Jerome, D.; Romero-Severson, J. Fraxinus Quadrangulate. The IUCN Red List of Threatened Species, 2017: E. Available online: https://www.iucnredlist.org/species/61919112/61919114 (accessed on 24 April 2018).
- Kovacs, K.F.; Haight, R.G.; McCullough, D.G.; Mercader, R.J.; Siegert, N.W.; Liebhold, A.M. Cost of potential Emerald Ash Borer damage in U.S. communities, 2009–2019. Ecol. Econom. 2010, 69, 569–578. [Google Scholar] [CrossRef]
- Kovacs, K.F.; Mercader, R.J.; Haight, R.G.; Siegert, N.W.; McCullough, D.G.; Liebhold, A.M. The influence of satellite populations of Emerald Ash Borer on projected economic costs in U.S. communities, 2010–2020. J. Environ. Manag. 2011, 92, 2170–2181. [Google Scholar] [CrossRef] [PubMed]
- Kostova, I.; Iossifova, T. Chemical components of Fraxinus species. Fitoterapia 2007, 78, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.P.; Potter, D.A.; Kimmerer, T.W. Carbohydrate reserves, radial growth, and mechanisms of resistance of oak trees to phloem-boring insects. Oecologia 1990, 83, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Eyles, A.; Jones, W.; Riedl, K.; Cipollini, D.; Schwartz, S.; Chan, K.; Herms, D.A.; Bonello, P. Comparative phloem chemistry of Manchurian (Fraxinus mandshurica) and two North American ash species (Fraxinus americana and Fraxinus pennsylvanica). J. Chem. Ecol. 2007, 33, 1430–1448. [Google Scholar] [CrossRef] [PubMed]
- Cipollini, D.; Wang, Q.; Whitehill, J.G.A.; Powell, J.R.; Bonello, P.; Herms, D.A. Distinguishing defensive characteristics in the phloem of ash species resistant and susceptible to Emerald Ash Borer. J. Chem. Ecol. 2011, 37, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Greger, H.; Hofer, O. New unsymmetrically substituted tetrahydrofurofuran lignans from Artemisia absinthium-Assignment of the relative stereochemistry by lanthanide chemical shifts. Tetrahedron 1980, 36, 3551–3558. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Hisada, S.; Nishide, S. Lignans from bark of Fraxinus mandshurica var. japonica and F. japonica. Chem. Pharm. Bull. 1984, 32, 4482–4489. [Google Scholar] [CrossRef]
- Whitehill, J.G.A.; Opiyo, S.O.; Koch, J.L.; Herms, D.A.; Cipollini, D.F.; Bonello, P. Interspecific comparison of constitutive ash phloem phenolic chemistry reveals compounds unique to Manchurian ash, a species resistant to Emerald Ash borer. J. Chem. Ecol. 2012, 38, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Whitehill, J.G.A.; Hill, A.L.; Opiyo, S.O.; Cipollini, D.; Herms, D.A.; Bonello, P. Effects of water availability on Emerald Ash Borer larval performance and phloem phenolics of Manchurian and black ash. Plant Cell Environ. 2014, 37, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Adfa, M.; Yoshimura, T.; Komura, K.; Koketsu, M. Antitermite activities of coumarin derivatives and scopoletin from Protium javanicum Burm. f. J. Chem. Ecol. 2010, 36, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. Recent advances in chemical ecology. Nat. Prod. Rep. 1999, 16, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, B.; Olechnowicz-Stepien, W. Study of Fraxinus excelsior L. leaves. I. Phenolic acids and flavonoids. Herba Pol. 1988, 34, 7–13. [Google Scholar]
- Grujic-Vaciæ, J.; Ramic, S.; Bašiæ, F.; Bosnic, T. Phenolic compounds in the bark and leaves of Fraxinus I sorts. Acta Biol. Med. Exp. 1989, 14, 17–30. [Google Scholar]
- Hosny, M. Secoiridoid glucosides from Fraxinus oxycarba. Phytochemistry 1998, 47, 1569–1576. [Google Scholar] [CrossRef]
- Ayouni, K.; Berboucha-Rahmani, M.; Kim, H.K.; Atmani, D.; Verpoorte, R.; Choi, Y.H. Metabolomic tool to identify antioxidant compounds of Fraxinus angustifolia leaf and stem bark extracts. Ind. Crops Prod. 2016, 88, 65–77. [Google Scholar] [CrossRef]
- Takenaka, Y.; Tanahashi, T.; Shintaku, M.; Sakai, T.; Nagakura, N. Secoiridoid glucosides from Fraxinus americana. Phytochemistry 2000, 55, 275–284. [Google Scholar] [CrossRef]
- Cleary, M.R.; Andersson, P.F.; Broberg, A.; Elfstrand, M.; Daniel, G.; Stenlid, J. Genotypes of Fraxinus excelsior with different susceptibility to the ash dieback pathogen Hymenoscyphus pseudoalbidus and their response to the phytotoxin viridiol—A metabolomic and microscopic study. Phytochemistry 2014, 102, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.C. Tannin-insect interactions. In Chemistry and Significance of condensed tannins, 3rd ed.; Hemingway, R.W., Karchesy, J.J., Branham, S.J., Eds.; Springer: New York, NY, USA, 1989; pp. 417–433. [Google Scholar]
- Abou-Zaid, M.M.; Helson, B.; Nozzolillo, C.; Arnason, J.T. Gallates from red maple, Acer rubrum as a source of resistance to forest tent caterpillar, Malacosoma disstria. J. Chem. Ecol. 2001, 27, 2517–2527. [Google Scholar] [CrossRef] [PubMed]
- Sollars, E.S.; Harper, A.L.; Kelly, L.J.; Sambles, C.M.; Ramirez-Gonzalez, R.H.; Swarbreck, D.; Kaithakottil, G.; Cooper, E.D.; Uauy, C.; Havlickova, L.; et al. Genome sequence and genetic diversity of European ash trees. Nature 2007, 541, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Sambles, C.M.; Salmon, D.L.; Florance, H.; Howard, T.P.; Smirnoff, N.; Nielsen, L.R.; McKinney, L.V.; Kjær, E.D.; Buggs, R.J.A.; Studholme, D.J.; et al. Ash leaf metabolomes reveal differences between trees tolerant and susceptible to ash dieback disease. Sci. Data 2017, 4, 170190. [Google Scholar] [CrossRef] [PubMed]
- Deu-Pons, J.; Schroeder, M.P.; Lopez-Bigas, N.J. Heatmap: An interactive heatmap viewer for the web. Bioinformatics 2014, 30, 1757–1758. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hirayama, C.; Yasui, H.; Nakamura, M. Enzymatic activation of oleuropein: A protein crosslinker used as a chemical defense in the privet tree. Proc. Natl. Acad. Sci. USA 1999, 96, 9159–9164. [Google Scholar] [CrossRef] [PubMed]
- Kumarasamy, Y.; Nahar, L.; Cox, P.J.; Jaspars, M.; Sarker, S.D. Bioactivity of secoiridoid glycosides from Centaurium erythraea. Phytomedicine 2003, 10, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B.; Debnath, S.; Harigaya, Y. Naturally occurring secoiridoids and bioactivity of naturally occurring iridoids and secoiridoids. A review, part 2. Chem. Pharm. Bull. 2007, 55, 689–728. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Whitehill, J.G.A.; Bonello, P.; Poland, T.M. Differential response in foliar chemistry of three ash species to Emerald Ash Borer adult feeding. J. Chem. Ecol. 2011, 37, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Terazawa, M.; Sasaya, T. Extractives of Yachidamo Fraxinus mandshurica RUPR. var. japonica MAXIM. III. Extractives of woods. Phenolic compounds in sapwood. Mokuzai Gakkaishi 1971, 17, 167–173. [Google Scholar]
- Baxter, H.; Harborne, J.B.; Moss, G.P. Phytochemical Dictionary. In A Handbook of Bioactive Compounds from Plants, 2nd ed.; Taylor and Francis LTD: London, UK, 1999. [Google Scholar]
- Miyazawa, M.; Ishikawa, Y.; Kasahara, H.; Yamanaka, J.I.; Kameoka, H. An insect growth inhibitory lignan from flower buds of Magnolia fargesii. Phytochemistry 1994, 35, 611–613. [Google Scholar] [CrossRef]
- Cabral, M.M.O.; Kelecom, A.; Garcia, E.S. Effects of the lignan pinoresinol on the moulting cycle of the bloodsucking bug Rhodnius prolixus and of the milkweed bug Oncopeltus fasciatus. Fitoterapia 1999, 70, 561–567. [Google Scholar] [CrossRef]
- Garcia, E.S.; Cabral, M.M.; Schaub, G.A.; Gottlieb, O.R.; Azambuja, P. Effects of lignoids on a hematophagous bug, Rhodnius prolixus: Feeding, ecdysis and diuresis. Phytochemistry 2000, 55, 611–616. [Google Scholar] [CrossRef]
- Macahig, R.A.; Harinantenaina, L.; Matsunami, K.; Otsuka, H.; Takeda, Y.; Shinzato, T. Secoiridoid and iridoid glucosides from the leaves of Fraxinus griffithii. J. Nat. Med. 2010, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gowda, H.; Ivanisevic, J.; Johnson, C.H.; Kurczy, M.E.; Benton, H.P.; Rinehart, D.; Nguyen, T.; Ray, J.; Kuehl, J.; Arevalo, B.; et al. Interactive XCMS Online: Simplifying advanced metabolomic data processing and subsequent statistical analyses. Anal. Chem. 2014, 86, 6931–6939. [Google Scholar] [CrossRef] [PubMed]
- Dey, P.M.; Harborne, B. Plethora of polyphenols Plant Phenolics. In Methods in Plant Biochemistry; Harborne, B., Ed.; Academic Press: London, UK, 1989; Volume 1. [Google Scholar]
- Fossen, T.; Andersen, O.M. Spectroscopic techniques applied to flavonoids. In Flavonoids: Chemistry, Biochemistry and Applications; Andersen, O.M., Markham, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 37–142. [Google Scholar]
- Benton, H.P.; Ivanisevic, J.; Mahieu, N.G.; Kurczy, M.E.; Johnson, C.H.; Franco, L.; Rinehart, D.; Valentine, E.; Gowda, H.; Ubhi, B.K.; et al. Autonomous metabolomics for rapid metabolite identification in global profiling. Anal. Chem. 2015, 87, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Ivanisevic, J.; Benton, P.H.; Rinehart, D.; Epstein, A.; Kurczy, M.E.; Boska, M.D.; Gendelman, H.E.; Siuzdak, G. An interactive cluster heat map to visualize and explore multidimensional metabolomic data. Metabolomics 2015, 11, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Qazi, S.S.; Li, D.; Briens, C.; Berruti, F.; Abou-Zaid, M.M. Antioxidant activity of the lignins derived from Fluidized-Bed Fast Pyrolysis. Molecules 2017, 22, 372. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dipetalae | Fraxinus | Melioides | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No | Compound Name | |||||||||
| Actual Mass | M + H | RT | Blue Ash | Manchurian Ash | Black Ash | Green Ash | Pumpkin Ash | White Ash | ||
| Coumarins | ||||||||||
| 1 | Esculin | 340.284 | 341.0857 | 2.8 | 0.31 | 0.12 | 0.37 | N.D. | N.D. | N.D. |
| 2 | Esculetin | 178.143 | 179.0335 | 3.55 | 0.07 | 0.32 | 0.08 | N.D. | N.D. | N.D. |
| 3 | Fraxetin | 208.169 | 209.0439 | 4.2 | 0.11 | 0.21 | 0.15 | N.D. | N.D. | N.D. |
| 4 | Fraxidin | 222.196 | 223.0595 | 5.64 | 0.46 | 0.17 | 0.05 | N.D. | N.D. | N.D. |
| 5 | Fraxin | 370.310 | 371.0962 | 5.7 | 0.05 | 0.42 | 0.32 | N.D. | N.D. | N.D. |
| 6 | Scopoletin | 192.170 | 193.0492 | 5.0 | 6.89 | 3.09 | 1.37 | N.D. | 0.09 | 0.08 |
| Phenolics/Phenolic Acids | ||||||||||
| 7 | Chlorogenic acid | 354.311 | 355.1018 | 3.5 | 0.08 | 0.13 | 0.07 | 0.04 | 0.06 | 0.17 |
| 8 | Caffeic acid | 180.159 | 181.0496 | 3.7 | 0.08 | 0.58 | N.D. | N.D. | N.D. | N.D. |
| 9 | Syringic acid | 198.174 | 199.0601 | 3.9 | 0.04 | 0.07 | N.D. | N.D. | N.D. | N.D. |
| 10 | Ferulic acid | 194.186 | 195.0651 | 5.2 | 0.84 | N.D. | N.D. | N.D. | N.D. | N.D. |
| 11 | p-Coumaric acid | 164.160 | 165.0546 | 4.8 | 0.79 | 0.1 | 0.08 | N.D. | 0.04 | 0.04 |
| 12 | Gentisic acid | 154.121 | 155.0338 | 5.74 | 0.24 | N.D. | N.D. | N.D. | N.D. | N.D. |
| Flavanols | ||||||||||
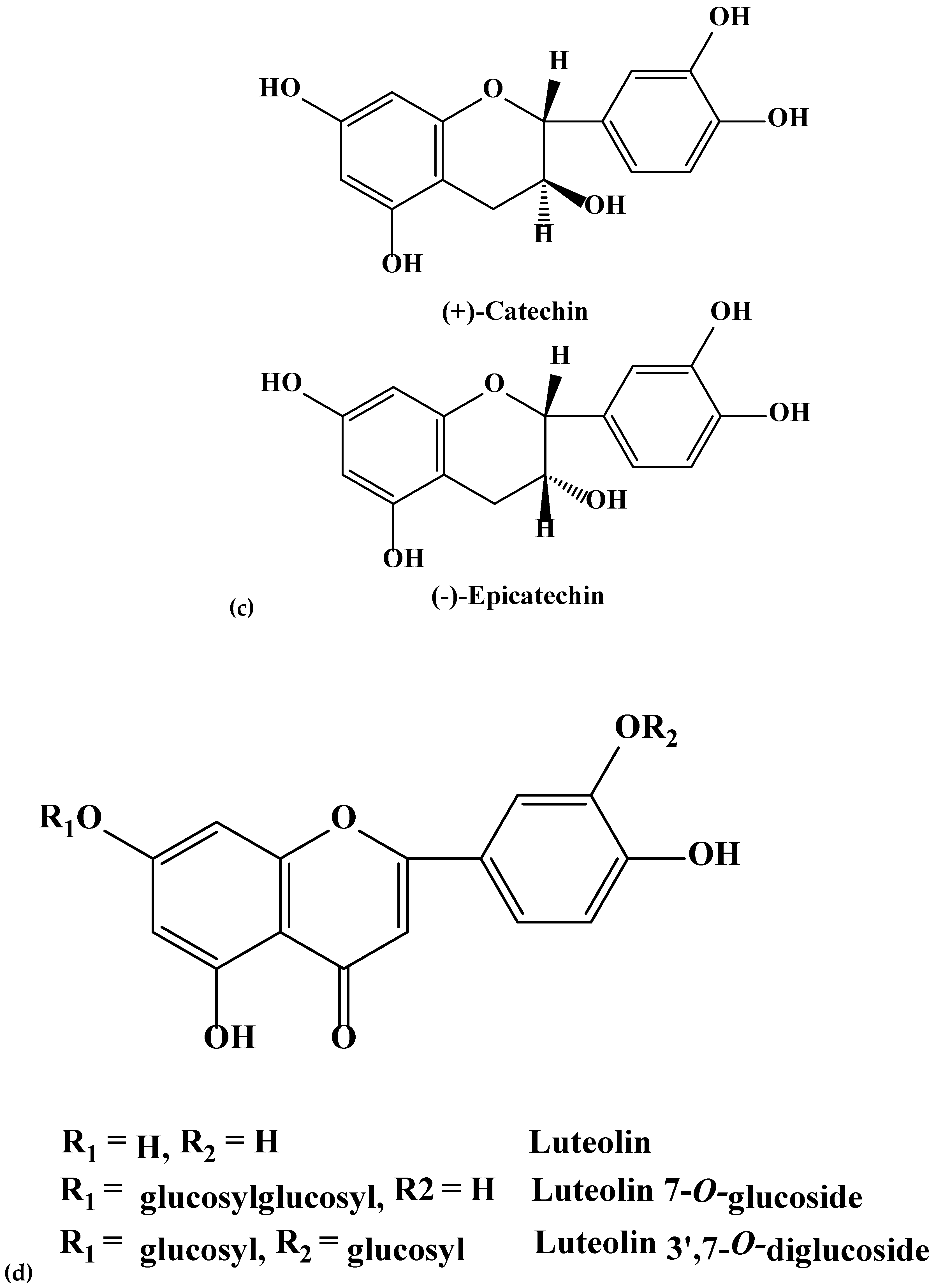
| 13 | (+) Catechin | 290.271 | 291.0859 | 10.0 | 0.15 | 0.04 | 0.02 | 0.18 | 0.01 | 0.03 |
| 14 | (−)-Epicatechin | 290.271 | 291.0859 | 15.3 | 0.09 | 0.12 | 0.06 | 0.15 | 0.01 | 0.09 |
| Flavones | ||||||||||
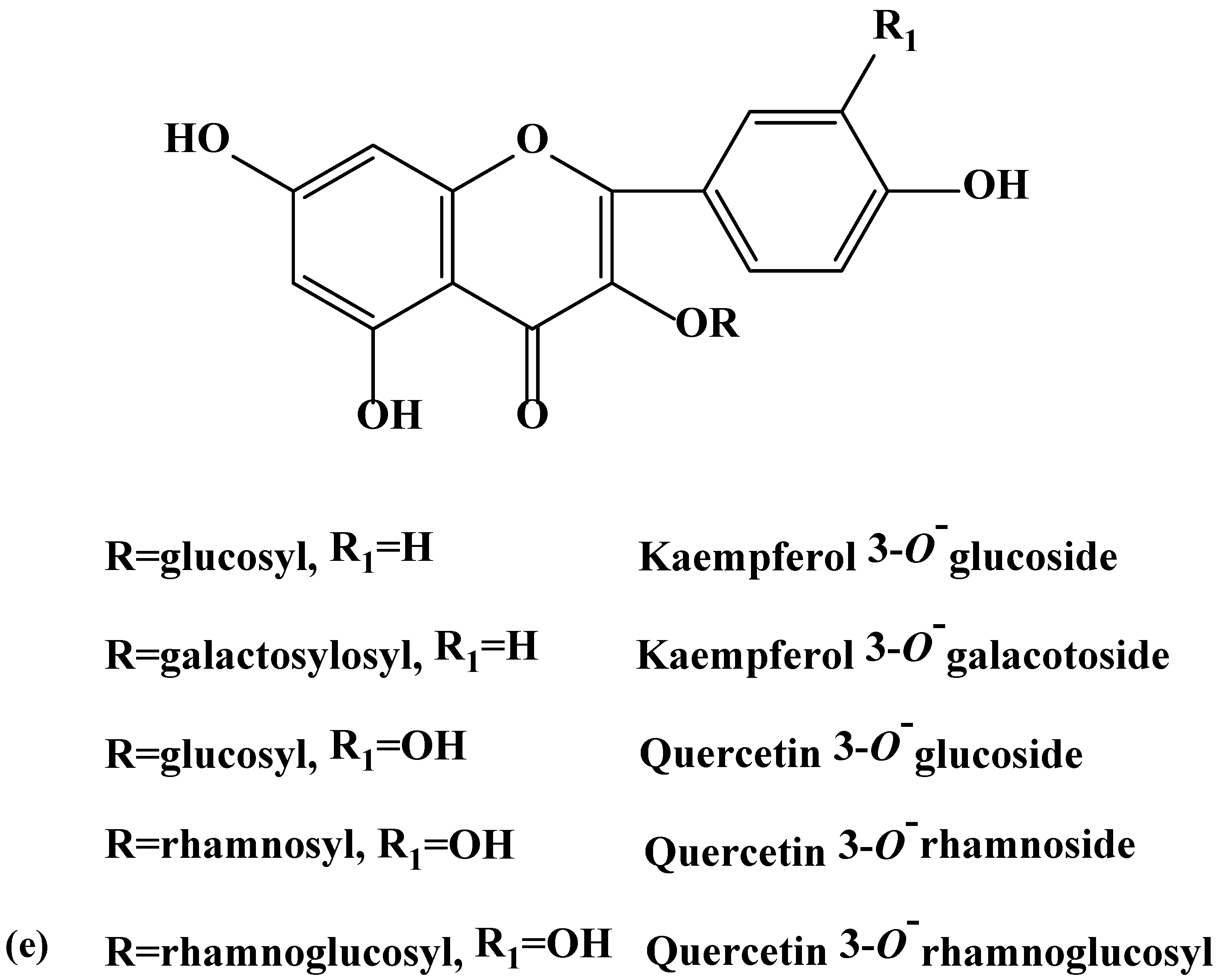
| 15 | Apigenin | 270.240 | 271.0597 | 8.6 | N.D. | N.D. | N.D. | 0.68 | 0.18 | N.D. |
| 16 | Luteolin | 286.239 | 287.0545 | 7.7 | N.D. | N.D. | N.D. | 0.1 | 0.05 | N.D. |
| 17 | Luteolin-7-O-glucoside | 448.380 | 449.1068 | 5.5 | N.D. | 0.89 | N.D. | 1.4 | 0.17 | 0.16 |
| 18 | Luteolin-3’,7-O-β-d-diglucoside | 610.518 | 611.1589 | 4.9 | N.D. | N.D. | N.D. | 0.04 | 0.02 | 0.21 |
| Flavonols | ||||||||||
| 19 | Quercetin-3-O-β-d-glucoside | 464.379 | 465.1018 | 5.6 | 0.57 | 0.35 | 0.1 | 0.37 | 0.11 | 0.84 |
| 20 | Quercetin-3-O-β-d-galactoside | 464.379 | 465.1018 | 5.48 | N.D. | 0.64 | 0.28 | 0.03 | 0.1 | 0.04 |
| 21 | Quercetin-3-O-β-l-rhamnoside | 448.380 | 449.1068 | 6.14 | 0.14 | 1.1 | 0.13 | 3.32 | 0.68 | 1.0 |
| 22 | Quercetin-3-O-rhamnoglucoside | 610.521 | 611.1589 | 5.4 | 0.34 | 0.15 | 0.2 | N.D. | 0.34 | 0.86 |
| 23 | Kaempferol-3-O-β-d-glucoside | 448.380 | 449.1070 | 6.08 | 0.26 | 0.07 | 0.1 | N.D. | N.D. | 0.13 |
| 24 | Kaempferol-3-O-rhamnoglucoside | 594.522 | 595.1643 | 5.9 | N.D. | 0.3 | 0.32 | N.D. | 0.3 | 0.2 |
| 25 | Kaempferol-2-B-coumaryl glucoside | 594.525 | 595.1432 | 7.8 | N.D. | N.D. | N.D. | N.D. | 0.06 | N.D. |
| Flavanone | ||||||||||
| 26 | Naringenin | 272.257 | 273.0752 | 8.4 | N.D. | N.D. | N.D. | 0.51 | N.D. | N.D. |
| No | Compound Name | Formula | Actual Mass | M + H | RT | Fold Change | Up/Down | p Value |
|---|---|---|---|---|---|---|---|---|
| 1 | Dicyclomine | C19H35NO2 | 309.487 | 310.2736 | 9.78 | 50 | DOWN | 0.00047 |
| 2 | N-hydroxyamphetamine | C9H13NO | 151.205 | 152.1069 | 8.03 | 13 | UP | 0.00055 |
| 3 | Norethynodrel | C20H26O2 | 298.419 | 299.2002 | 15.8 | 30 | UP | 0.00056 |
| 4 | 4-(3,4-Difluorophenyl) piperidine | C11H13F2N | 197.101 | 198.1086 | 10.38 | 30 | DOWN | 0.00059 |
| 5 | Erythronolide A | C17H30N4 | 418.527 | 419.2685 | 8.7 | 18 | UP | 0.00067 |
| 6 | Phenol, 4-amino-2,6-bis(1-piperidinylmethyl)- | C18H29N3O | 303.450 | 304.2314 | 14.31 | 32 | UP | 0.00082 |
| 7 | Butyronitrile, 2,2-diphenyl-4-(4-(3-hydroxymethyl-2-oxo-1-benzimidaizolinyl) piperidino)- | C29H30N4O2 | 466.585 | 467.5260 | 9.1 | 28 | UP | 0.00094 |
| 8 | Pinoresinol glucoside | C26H32O11 | 520.525 | 521.2012 | 6.39 | 122 | UP | 0.01517 |
| 9 | 8-hydroxypinoresinol 4-glucoside | C26H32O12 | 536.530 | 537.1959 | 4.75 | 34 | UP | 0.01435 |
| 10 | Mandarone A | C20H26O3 | 314.425 | 315.1953 | 8.31 | 7.0 | UP | 0.02711 |
| 11 | N,N-Dicyclohexyl-2,3,4,5,6-pentahydroxyhexanamide | C18H33NO6 | 359.463 | 360.2377 | 10.16 | 15 | DOWN | 0.00097 |
| 12 | 2,2’-Disulfanediylbis[Nt-(hepta-1,6-dien-4-yl) aniline] | C26H32N2S2 | 436.200 | 437.2079 | 13.6 | 58 | UP | 0.00098 |
| 13 | Matairesinol | C20H22O6 | 358.141 | 359.1482 | 7.9 | 6.7 | UP | 0.00550 |
| 14 | Benzamide, N-(2-(diethylamino) ethyl)-4-((1-oxopropyl) amino)-, monohydrochloride | C16H26ClN3O2 | 327.171 | 328.2055 | 12.99 | 156 | UP | 0.00112 |
| 15 | beta-Ionone | C13H20O | 192.151 | 193.1585 | 8.0 | 12 | UP | 0.00113 |
| 16 | Ligstroside | C25H32O12 | 524.519 | 525.2226 | 7.34 | 37 | UP | 0.00118 |
| 17 | 2-Methoxy-1-(phenylselanyl) decan-3-ol | C17H28O2Se C17H28O2Se | 344.125 | 345.1328 | 7.36 | 56 | UP | 0.00130 |
| 18 | Totarol | C20H30O | 286.229 | 287.2366 | 16.04 | 264 | UP | 0.00144 |
| 19 | Clovanediol diacetate | C19H30O4 | 322.214 | 323.2212 | 12.74 | 29 | DOWN | 0.00147 |
| 20 | Mulberrofuran T | C44H44O9 C44H44O9 | 716.298 | 717.3144 | 8.8 | 24 | UP | 0.00149 |
| 21 | ar-Artemisene | C20H30 | 270.234 | 271.2416 | 16.41 | 109 | UP | 0.00171 |
| 22 | 11,12-dihydroxy arachidic acid | C20H40O4 | 344.292 | 345.2994 | 8.22 | 20 | UP | 0.00174 |
| 23 | 1-Docosylpyridin-1-ium bromide | C27H50BrN | 467.312 | 468.3465 | 17.52 | 57 | DOWN | 0.00221 |
| 24 | Aphidicolin | C20H34O4 | 338.245 | 339.2525 | 10.29 | 35 | DOWN | 0.00236 |
| 25 | Parthenolide | C15H20O3 | 248.141 | 249.1481 | 10.46 | 20 | DOWN | 0.00291 |
| 26 | Pentaquine | C18H27N3O | 301.215 | 302.2156 | 9.74 | 56 | UP | 0.00293 |
| 27 | Famciclovir | C14H19N5O4 | 321.143 | 322.1773 | 8.42 | 24 | DOWN | 0.00302 |
| 28 | ar-Turmerone | C15H20O | 216.151 | 217.1583 | 14.92 | 22 | DOWN | 0.00309 |
| 29 | Myriocin | C21H39NO6 | 401.277 | 402.2846 | 10.4 | 20 | DOWN | 0.00439 |
| 30 | Coniferin | C16H22O8 | 342.131 | 343.1384 | 4.94 | 14 | UP | 0.07002 |
| 31 | Isosyringinoside | C23H34O14 | 534.194 | 535.1834 | 6.36 | 54 | UP | 0.03068 |
| 32 | Oleuropein | C25H32O13 | 540.184 | 541.2175 | 6.58 | 58 | UP | 0.00360 |
| 33 | Hydroxytyrosol 1-O-glucoside | C14H20O8 | 316.115 | 317.1492 | 1.25 | 16 | UP | 0.03709 |
| 34 | Verbenalin | C17H24O10 | 388.136 | 389.1437 | 3.11 | 64 | UP | 0.01169 |
| 35 | Fraxinol | C11H10O5 | 222.052 | 223.0599 | 5.2 | 9.3 | UP | 0.00060 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qazi, S.S.; Lombardo, D.A.; Abou-Zaid, M.M. A Metabolomic and HPLC-MS/MS Analysis of the Foliar Phenolics, Flavonoids and Coumarins of the Fraxinus Species Resistant and Susceptible to Emerald Ash Borer. Molecules 2018, 23, 2734. https://doi.org/10.3390/molecules23112734
Qazi SS, Lombardo DA, Abou-Zaid MM. A Metabolomic and HPLC-MS/MS Analysis of the Foliar Phenolics, Flavonoids and Coumarins of the Fraxinus Species Resistant and Susceptible to Emerald Ash Borer. Molecules. 2018; 23(11):2734. https://doi.org/10.3390/molecules23112734
Chicago/Turabian StyleQazi, Sohail S., Domenic A. Lombardo, and Mamdouh M. Abou-Zaid. 2018. "A Metabolomic and HPLC-MS/MS Analysis of the Foliar Phenolics, Flavonoids and Coumarins of the Fraxinus Species Resistant and Susceptible to Emerald Ash Borer" Molecules 23, no. 11: 2734. https://doi.org/10.3390/molecules23112734
APA StyleQazi, S. S., Lombardo, D. A., & Abou-Zaid, M. M. (2018). A Metabolomic and HPLC-MS/MS Analysis of the Foliar Phenolics, Flavonoids and Coumarins of the Fraxinus Species Resistant and Susceptible to Emerald Ash Borer. Molecules, 23(11), 2734. https://doi.org/10.3390/molecules23112734