Effects of Different Cultivation Parameters on the Production of Surfactin Variants by a Bacillus subtilis Strain

 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
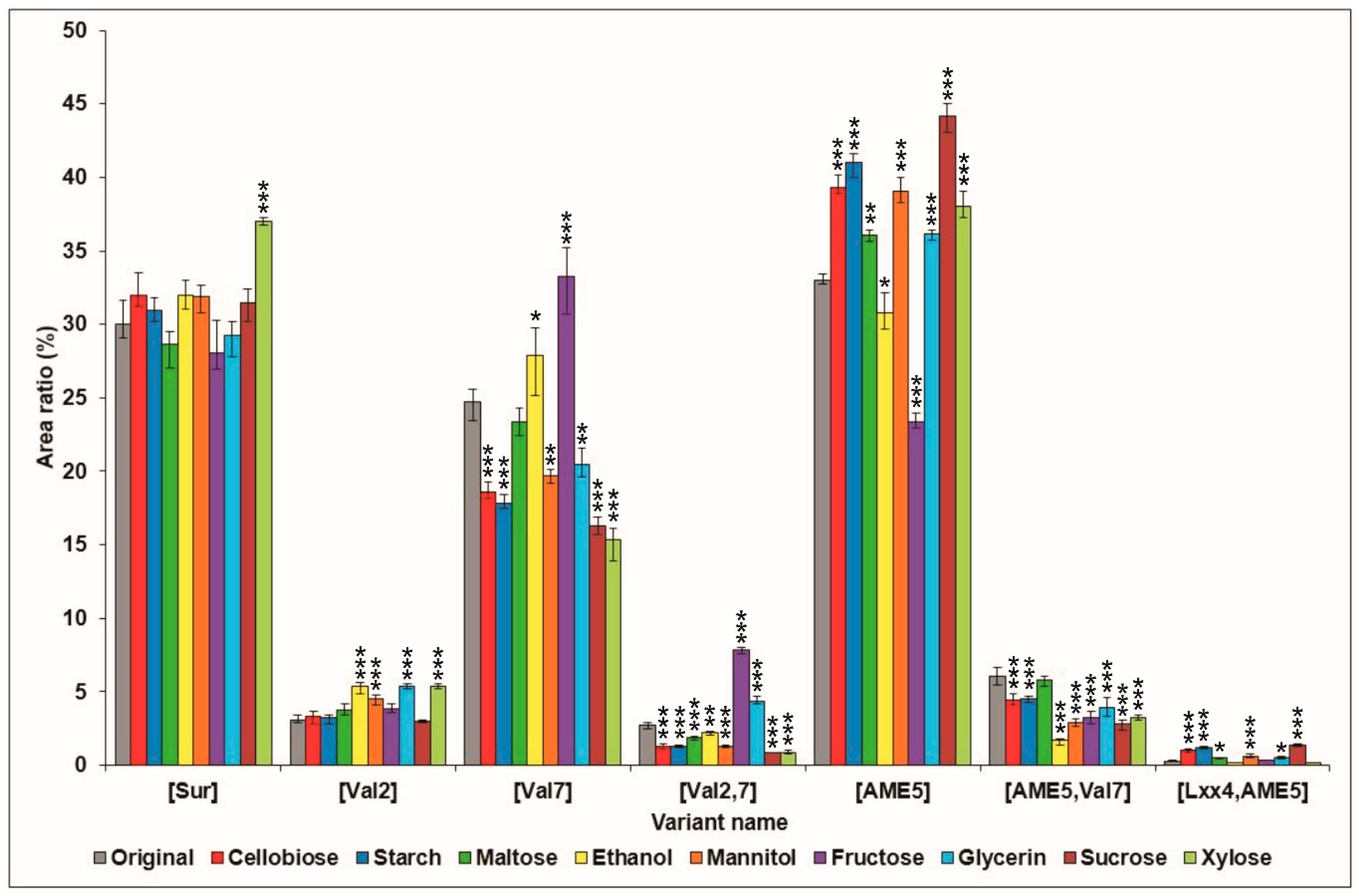
2.1. Effects of Various Carbon Sources on Surfactin Production
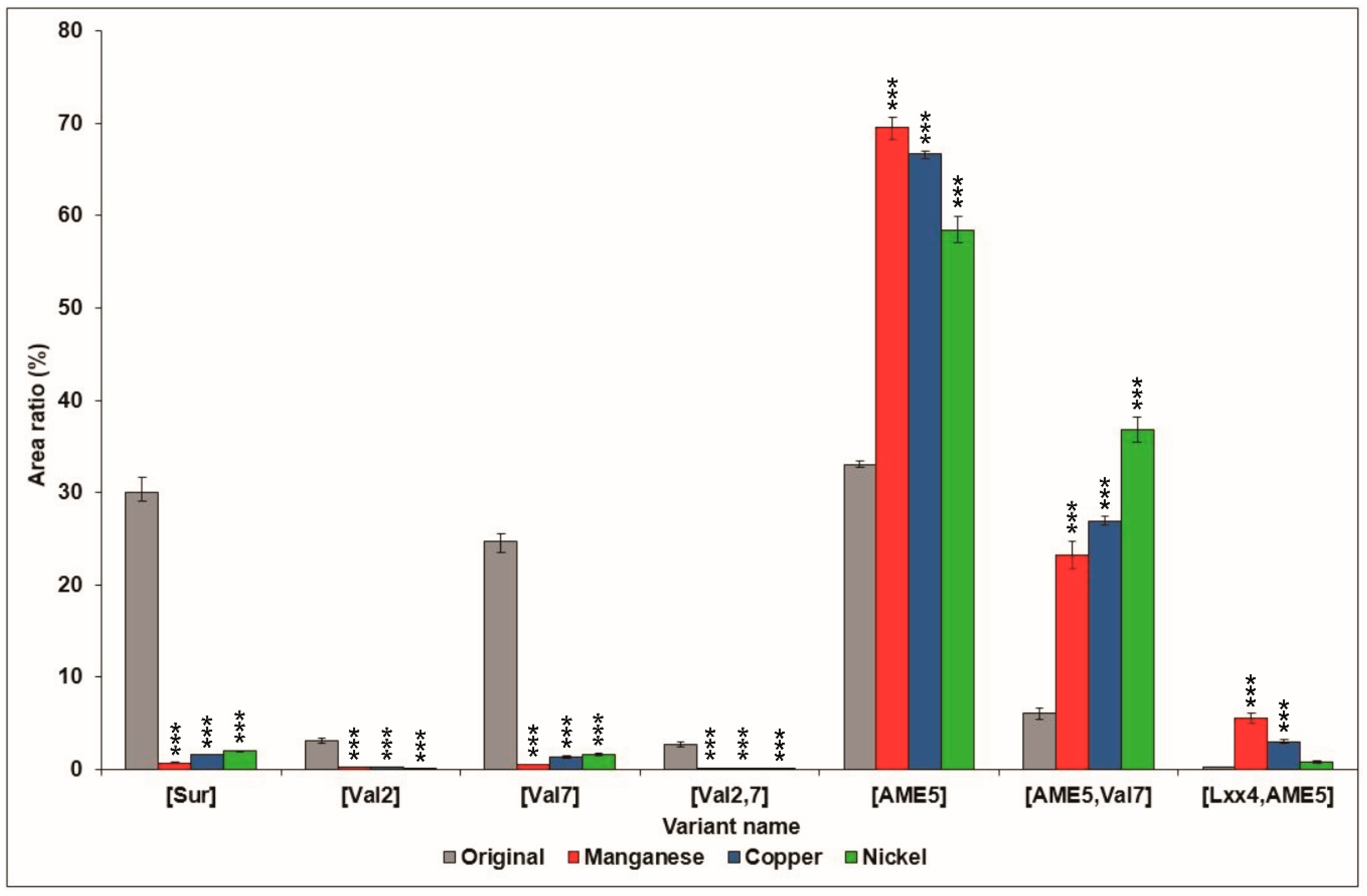
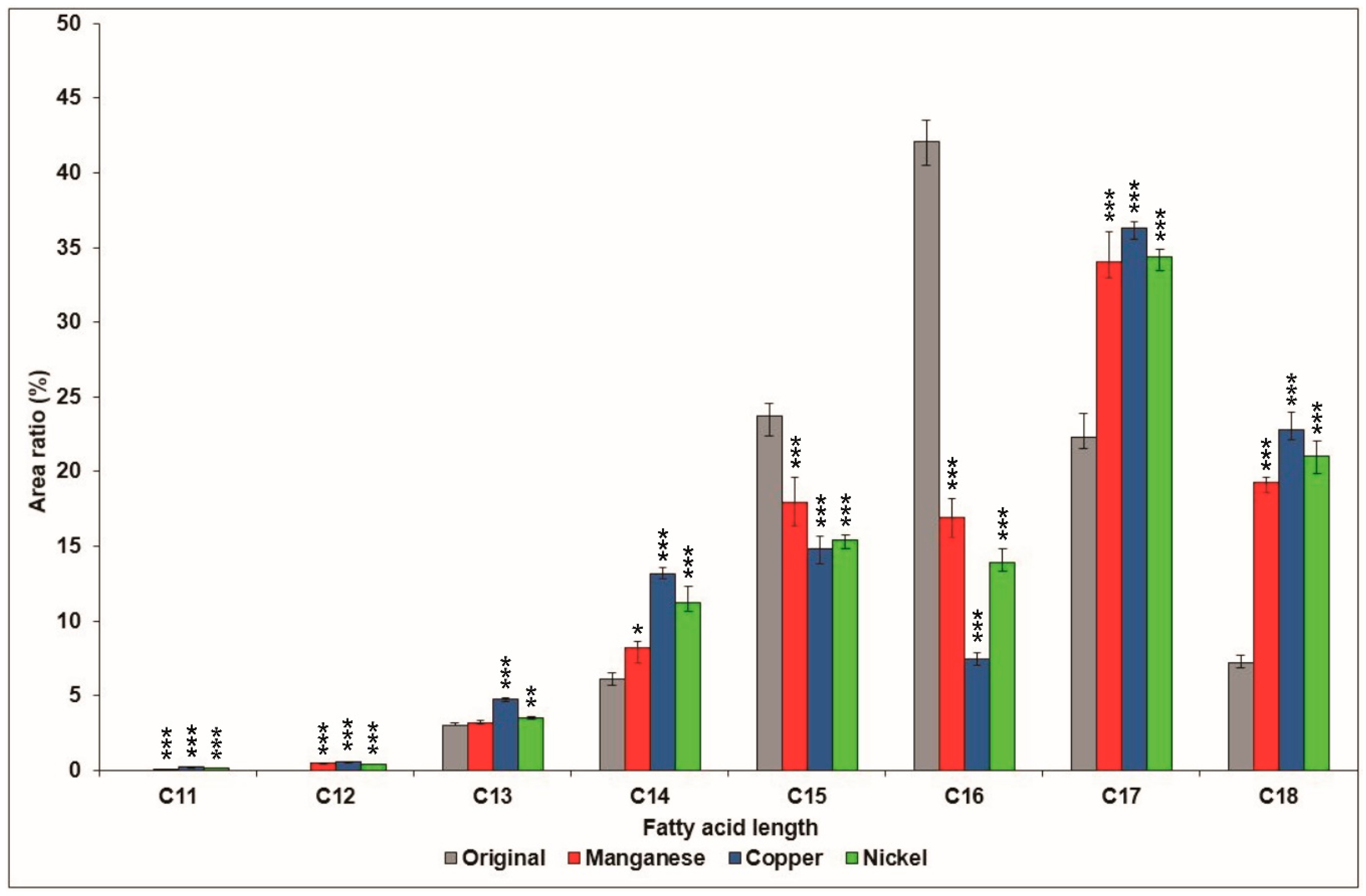
2.2. Effects of Culture Media Supplemented with Various Metal Ions on Surfactin Production
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Microorganism and Culture Conditions
3.3. Extraction and Measurement of Surfactin Variants
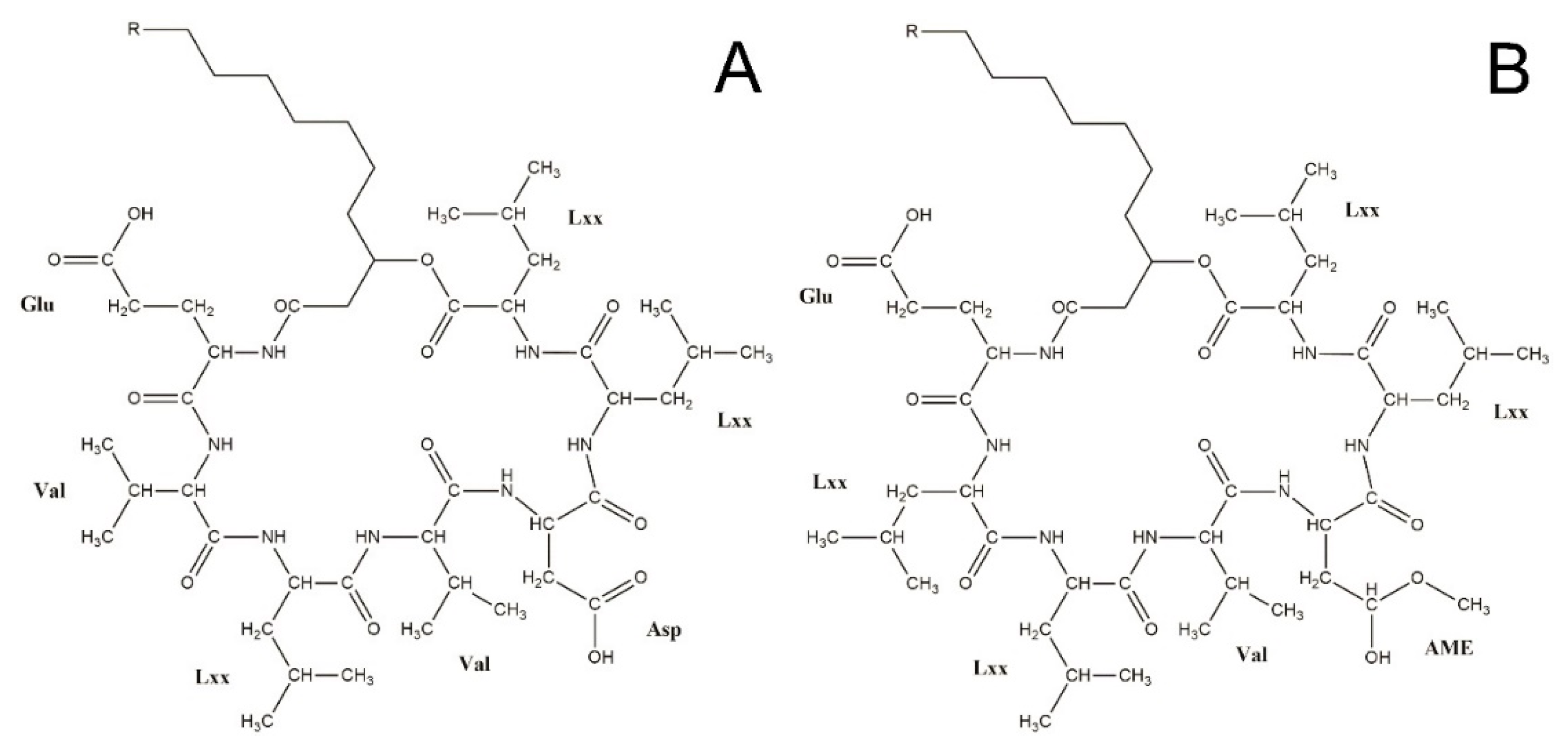
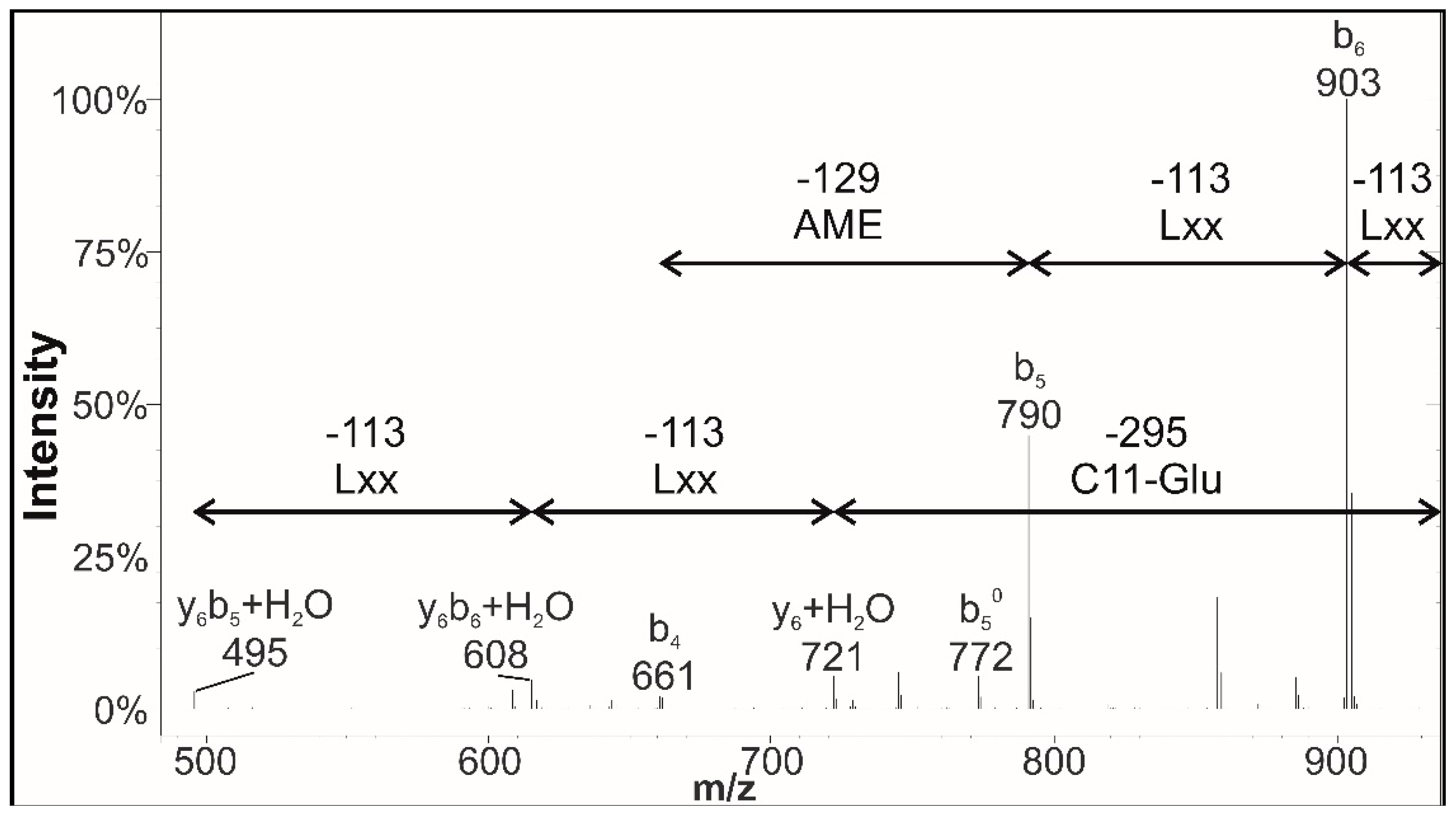
3.4. Structural Elucidation of Novel Surfactins
3.5. Nomenclature
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Arima, K.; Kakinuma, A.; Tamura, G. Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: Isolation, characterization and its inhibition of fibrin clot formation. Biochem. Biophys. Res. Commun. 1968, 31, 488–494. [Google Scholar] [CrossRef]
- Chen, W.-C.; Juang, R.-S.; Wei, Y.-H. Applications of a lipopeptide biosurfactant, surfactin, produced by microorganisms. Biochem. Eng. J. 2015, 103, 158–169. [Google Scholar] [CrossRef]
- Nieminen, T.; Rintaluoma, N.; Andersson, M.; Taimisto, A.-M.; Ali-Vehmas, T.; Seppälä, A.; Priha, O.; Salkinoja-Salonen, M. Toxinogenic Bacillus pumilus and Bacillus licheniformis from mastitic milk. Vet. Microbiol. 2007, 124, 329–339. [Google Scholar] [CrossRef] [PubMed]
- From, C.; Hormazabal, V.; Hardy, S.P.; Granum, P.E. Cytotoxicity in Bacillus mojavensis is abolished following loss of surfactin synthesis: Implications for assessment of toxicity and food poisoning potential. Inter. J. Food Microbiol. 2007, 117, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Pecci, Y.; Rivardo, F.; Martinotti, M.G.; Allegrone, G. LC/ESI-MS/MS characterisation of lipopeptide biosurfactants produced by the Bacillus licheniformis V9T14 strain. J. Mass Spectrom. 2010, 45, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, F.; Castro, M.; Príncipe, A.; Borioli, G.; Fischer, S.; Mori, G.; Jofré, E. The plant-associated Bacillus amyloliquefaciens strains MEP218 and ARP23 capable of producing the cyclic lipopeptides iturin or surfactin and fengycin are effective in biocontrol of sclerotinia stem rot disease. J. Appl. Microbiol. 2011, 112, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Vollenbroich, D.; Pauli, G.; Zel, M.O.; Vater, J. Antimycoplasma properties and application in cell culture of surfactin, a lipopeptide antibiotic from Bacillus subtilis. Appl. Environ. Microbiol. 1997, 63, 44–49. [Google Scholar] [PubMed]
- Wang, X.; Hu, W.; Zhu, L.; Yang, Q. Bacillus subtilis and surfactin inhibit the transmissible gastroenteritis virus from entering the intestinal epithelial cells. Biosci. Rep. 2017, 37, BSR20170082. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.; Gudiña, E.J.; Lima, C.F.; Rodrigues, L.R. Effects of biosurfactants on the viability and proliferation of human breast cancer cells. AMB Express 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, C.; Dong, B.; Ma, X.; Hou, L.; Cao, X.; Wang, C. Anti-inflammatory activity and mechanism of surfactin in lipopolysaccharide-activated macrophages. Inflammation 2015, 38, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Albino, J.D.; Nambi, I.M. Effect of biosurfactants on the aqueous solubility of PCE and TCE. J. Environ. Sci. Health Part A 2009, 44, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.-M.; Laprevote, O.; Peypoux, F. Diversity Among microbial cyclic lipopeptides: Iturins and surfactins. Activity-structure relationships to design new bioactive agents. Comb. Chem. High Throughput Screen. 2003, 6, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Peypoux, F.; Bonmatin, J.-M.; Labbe, H.; Grangemard, I.; Das, B.C.; Ptak, M.; Wallach, J.; Michel, G. [Ala4] Surfactin, a novel isoform from Bacillus subtilis studied by mass and NMR spectroscopies. Eur. J. Biochem. 1994, 224, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.-M.; Labbé, H.; Grangemard, I.; Peypoux, F.; Maget-Dana, R.; Ptak, M.; Michel, G. Production, isolation and characterization of [Leu4]- and [Ile4] surfactins from Bacillus subtilis. Lett. Pept. Sci. 1995, 2, 41–47. [Google Scholar] [CrossRef]
- Baumgart, F.; Kluge, B.; Ullrich, C.; Vater, J.; Ziessow, D. Identification of amino acid substitutions in the lipopeptide surfactin using 2D NMR spectroscopy. Biochem. Biophys. Res. Commun. 1991, 177, 998–1005. [Google Scholar] [CrossRef]
- Grangemard, I.; Peypoux, F.; Wallach, J.; Das, B.C.; Labbé, H.; Caille, A.; Genest, M.; Maget-Dana, R.; Ptak, M.; Bonmatin, J.-M. Lipopeptides with improved properties: Structure by NMR, purification by HPLC and structure–activity relationships of new isoleucyl-rich surfactins. J. Pept. Sci. 1997, 3, 145–154. [Google Scholar] [CrossRef]
- Kowall, M.; Vater, J.; Kluge, B.; Stein, T.; Franke, P.; Ziessow, D. Separation and characterization of surfactin isoforms produced by Bacillus subtilis OKB 105. J. Colloid Interface Sci. 1998, 204, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bóka, B.; Manczinger, L.; Kecskeméti, A.; Chandrasekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, C.; Szekeres, A. Ion trap mass spectrometry of surfactins produced by Bacillus subtilis SZMC 6179J reveals novel fragmentation features of cyclic lipopeptides. Rapid Commun. Mass Spectrom. 2016, 30, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Kecskeméti, A.; Bartal, A.; Bóka, B.; Kredics, L.; Manczinger, L.; Shine, K.; Alharby, N.S.; Khaled, J.M.; Varga, M.; Vágvölgyi, C.; et al. High-frequency occurrence of surfactin monomethyl isoforms in the ferment broth of a Bacillus subtilis strain revealed by ion trap mass spectrometry. Molecules 2018, 23, 2224. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.-S.; Gao, H.; Hong, K.; Yu, J.; Jiang, M.-M.; Lin, H.-P.; Ye, W.-C.; Yao, X.-S. Complete assignments of 1H and 13C NMR spectral data of nine surfactin isomers. Magn. Reson. Chem. 2007, 45, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.-S.; Zhao, F.; Gao, H.; Dai, Y.; Yao, Z.-H.; Hong, K.; Li, J.; Ye, W.-C.; Yao, X.-S. Characterization and online detection of surfactin isomers based on HPLC-MSn analyses and their inhibitory effects on the overproduction of nitric oxide and the release of TNF-α and IL-6 in LPS-induced macrophages. Mar. Drugs 2010, 8, 2605–2618. [Google Scholar] [CrossRef] [PubMed]
- Moro, G.V.; Almeida, R.T.R.; Napp, A.P.; Porto, C.; Pilau, E.J.; Lüdtke, D.S.; Moro, A.V.; Vainstein, M.H. Identification and ultra-high-performance liquid chromatography coupled with high-resolution mass spectrometry characterization of biosurfactants, including a new surfactin, isolated from oil-contaminated environments. Microb. Biotechnol. 2018, 11, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Yeh, M.-S.; Wei, Y.-H.; Chang, J.-S. Enhanced production of surfactin from Bacillus subtilis by addition of solid carriers. Biotechnol. Prog. 2008, 21, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mawgoud, A.M.; Aboulwafa, M.M.; Hassouna, N.A.-H. Optimization of surfactin production by Bacillus subtilis isolate BS5. Appl. Biochem. Biotechnol. 2008, 150, 305–325. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.I.A.; Wang, J.; Mu, B. Identification of a biosurfactant producing strain: Bacillus subtilis HOB2. Protein Pept. Lett. 2009, 16, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Swaminathan, T. Application of response-surface methodology to evaluate the optimum environmental conditions for the enhanced production of surfactin. Appl. Microbiol. Biotechnol. 1997, 47, 358–363. [Google Scholar] [CrossRef]
- Ghribi, D.; Ellouze-Chaabouni, S. Enhancement of Bacillus subtilis lipopeptide biosurfactants production through optimization of medium composition and adequate control of aeration. Biotechnol. Res. Int. 2011, 2011, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.-H.; Chu, I.-M. Mn2+ improves surfactin production by Bacillus subtilis. Biotechnol. Lett. 2002, 24, 479–482. [Google Scholar] [CrossRef]
- Slivinski, C.T.; Mallmann, E.; de Araújo, J.M.; Mitchell, D.A.; Krieger, N. Production of surfactin by Bacillus pumilus UFPEDA 448 in solid-state fermentation using a medium based on okara with sugarcane bagasse as a bulking agent. Process Biochem. 2012, 47, 1848–1855. [Google Scholar] [CrossRef]
- Joshi, S.J.; Al-Wahaibi, Y.M.; Al-Bahry, S.N.; Elshafie, A.E.; Al-Bemani, A.S.; Al-Mandhari, M.S. Production, characterization, and application of Bacillus licheniformis W16 biosurfactant in enhancing oil recovery. Front. Microbiol. 2016, 7, 1853. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.G.; Macdonald, C.R.; Duff, S.J.B.; Kosaric, N. Enhanced production of surfactin form Bacillus subtilis by continuous product removal and metal cation additions. Appl. Environ. Microbiol. 1981, 42, 408–412. [Google Scholar] [PubMed]
- Sheppard, J.D.; Cooper, D.G. The response of Bacillus subtilis ATCC 21332 to manganese during continuous-phased growth. Appl. Microbiol. Biotechnol. 1991, 35, 72–76. [Google Scholar] [CrossRef]
- Wei, Y.-H.; Chu, I.-M. Enhancement of surfactin production in iron-enriched media by Bacillus subtilis ATCC 21332. Enzyme Microb. Technol. 1998, 22, 724–728. [Google Scholar] [CrossRef]
- Wei, Y.-H.; Wang, L.-F.; Chang, J.-S.; Kung, S.-S. Identification of induced acification in iron-enriched cultures of Bacillus subtilis during biosurfactant fermentation. J. Biosci. Bioeng. 2003, 96, 174–178. [Google Scholar] [CrossRef]
- Wei, Y.-H.; Wang, L.-F.; Chang, J.-S. Optimizing iron supplement strategies for enhanced surfactin production with Bacillus subtilis. Biotechnol. Prog. 2004, 20, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, J.; Wang, Y.; Liu, J.; Lu, L. The positive effects of Mn2+ on nitrogen use and surfactin production by Bacillus subtilis ATCC 21332. Biotechnol. Biotechnol. Equip. 2015, 29, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.-H.; Lai, C.-C.; Chang, J.-S. Using Taguchi experimental design methods to optimize trace element composition for enhanced surfactin production by Bacillus subtilis ATCC 21332. Process Biochem. 2007, 42, 40–45. [Google Scholar] [CrossRef]
- Besson, F.; Chevanet, C.; Michel, G. Influence of the culture medium on the production of iturin A by Bacillus subtilis. J. Gen. Microbiol. 1987, 133, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Al-Ajlani, M.; Sheikh, M.; Ahmad, Z.; Hasnain, S. Production of surfactin from Bacillus subtilis MZ-7 grown on pharmamedia commercial medium. Microb. Cell Fact. 2007, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ren, B.; Gao, H.; Liu, M.; Dai, H.; Song, F.; Yu, Z.; Wang, S.; Hu, J.; Kokare, C.R.; et al. Optimization for the production of surfactin with a new synergistic antifungal activity. PLoS ONE 2012, 7, e34430. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.R.; Silva, A.J.R.; França, F.P.D.; Cardoso, V.L.; Sérvulo, E.F.C. Optimizing carbon/nitrogen ratio for biosurfactant production by a Bacillus subtilis strain. Appl. Biochem. Biotechnol. 2007, 136, 471–486. [Google Scholar]
- Akpa, E.; Jacques, P.; Wathelet, B.; Paquot, M.; Fuchs, R.; Budzikiewicz, H.; Thonart, P. Influence of culture conditions on lipopeptide production by Bacillus subtilis. In Twenty-Second Symposium on Biotechnology for Fuels and Chemicals; Humana Press: Totowa, NJ, USA, 2001; pp. 551–561. [Google Scholar]
- Peypoux, F.; Michel, G. Controlled biosynthesis of Val7- and Leu7-surfactins. Appl. Microbiol. Biotechnol. 1992, 36. [Google Scholar] [CrossRef]
- Romano, A.; Vitullo, D.; Di Pietro, A.; Lima, G.; Lanzotti, V. Antifungal lipopeptides from Bacillus amyloliquefaciens strain BO7. J. Nat. Prod. 2011, 74, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Shaligram, N.S.; Singhal, R.S. Surfactin–A Review on biosynthesis, fermentation, purification and applications. Food Technol. Biotechnol. 2010, 48, 119–134. [Google Scholar]
- Vágvölgyi, C.; Sajben-Nagy, E.; Bóka, B.; Vörös, M.; Berki, A.; Palágyi, A.; Krisch, J.; Škrbić, B.; Đurišić-Mladenović, N.; Manczinger, L. Isolation and characterization of antagonistic Bacillus strains capable to degrade ethylenethiourea. Curr. Microbiol. 2013, 66, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, P.; Fohlman, J. Proposal for a common nomenclature for sequence ions in mass spectra of peptides. Biol. Mass Spectrom. 1984, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Biemann, K. Sequencing of peptides by tandem mass spectrometry and high-energy collision-induced dissociation. Methods Enzymol. 1990, 193, 455–479. [Google Scholar] [PubMed]
- Hue, N.; Serani, L.; Laprévote, O. Structural investigation of cyclic peptidolipids from Bacillus subtilis by high-energy tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2001, 15, 203–209. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |

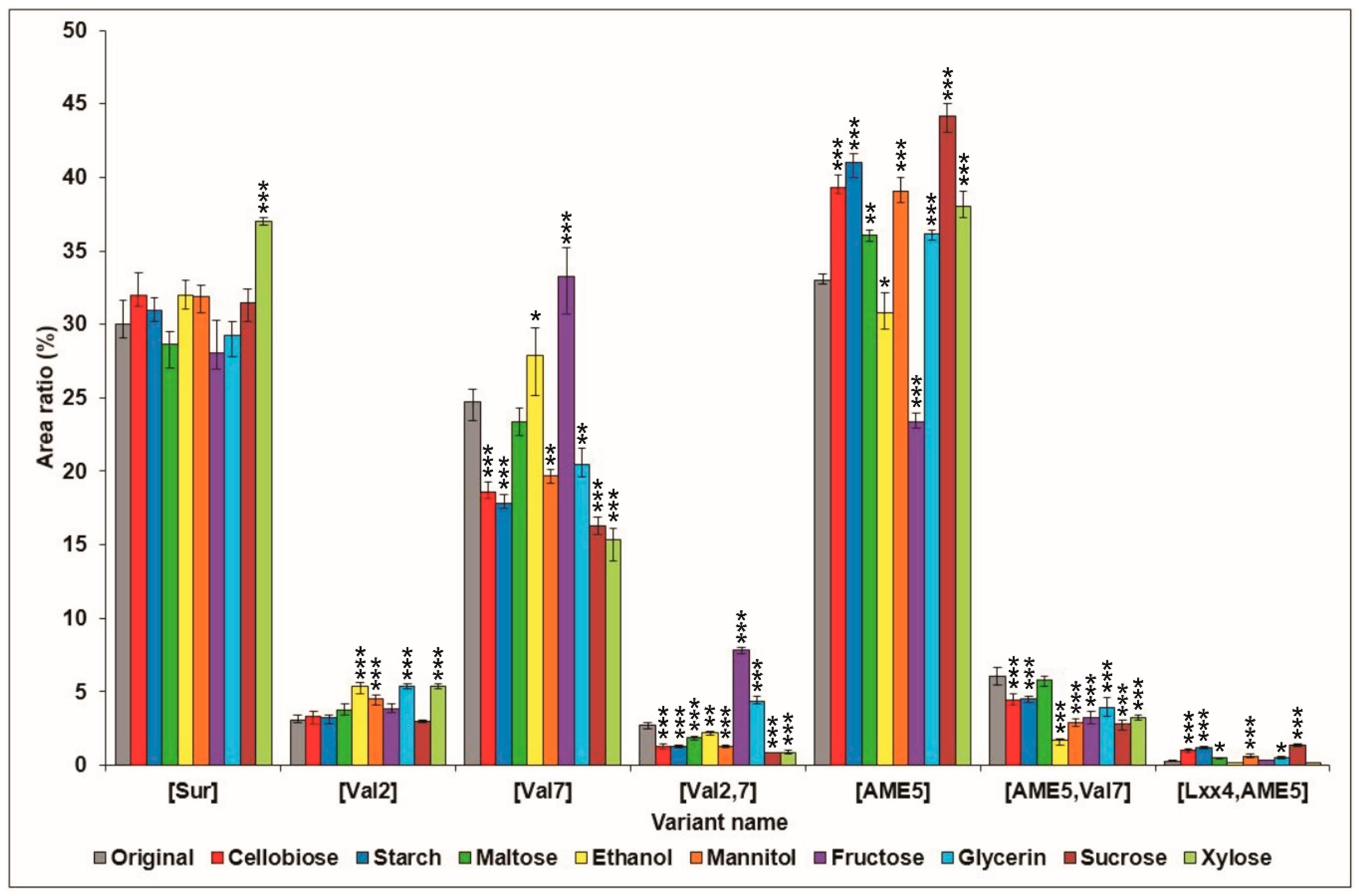





{kind=link}
{kind=link}
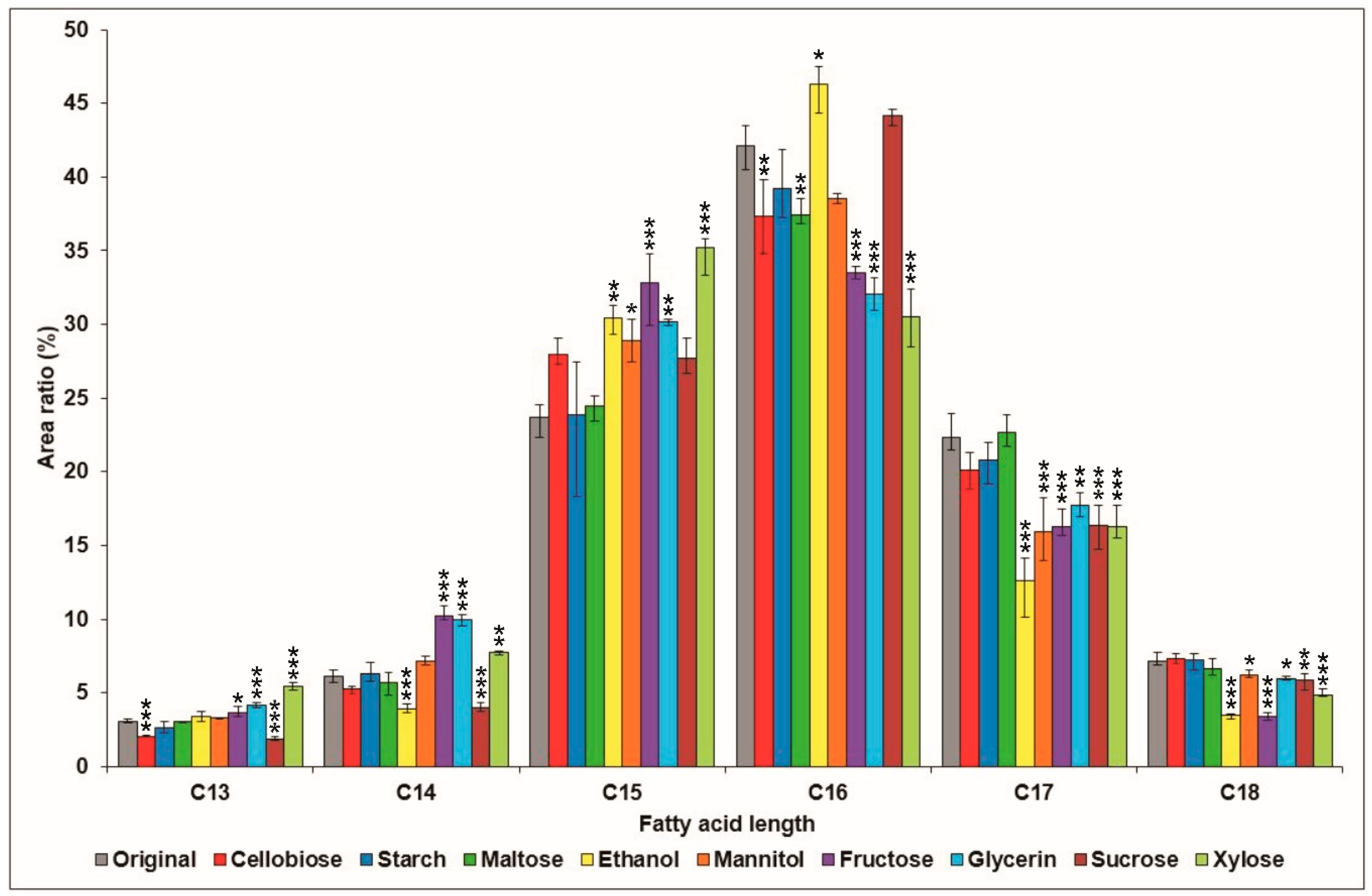{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Mass | Fatty Acid Chain | Peptide Sequence 1 |
|---|---|---|
| 993 | C13 | [Val2], [Val7] |
| C14 | [Val2,7] | |
| 1007 | C13 | [Sur] |
| C14 | [Val7] | |
| C15 | [Val2,7] | |
| 1021 | C14 | [Sur] |
| C15 | [Val2], [Val7] | |
| 1035 | C15 | [Sur] |
| C16 | [Val7] | |
| 1049 | C15 | [AME5] |
| C16 | [Sur] | |
| C17 | [Val7] | |
| 1063 | C16 | [AME5] |
| C17 | [AME5,Val7], [Sur] | |
| 1077 | C17 | [AME5] |
| C18 | [AME5,Val7] | |
| 1091 | C17 | [Lxx4,AME5] |
| C18 | [AME5] | |
| 1105 | C18 | [Lxx4,AME5] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartal, A.; Vigneshwari, A.; Bóka, B.; Vörös, M.; Takács, I.; Kredics, L.; Manczinger, L.; Varga, M.; Vágvölgyi, C.; Szekeres, A. Effects of Different Cultivation Parameters on the Production of Surfactin Variants by a Bacillus subtilis Strain. Molecules 2018, 23, 2675. https://doi.org/10.3390/molecules23102675
Bartal A, Vigneshwari A, Bóka B, Vörös M, Takács I, Kredics L, Manczinger L, Varga M, Vágvölgyi C, Szekeres A. Effects of Different Cultivation Parameters on the Production of Surfactin Variants by a Bacillus subtilis Strain. Molecules. 2018; 23(10):2675. https://doi.org/10.3390/molecules23102675
Chicago/Turabian StyleBartal, Attila, Aruna Vigneshwari, Bettina Bóka, Mónika Vörös, István Takács, László Kredics, László Manczinger, Mónika Varga, Csaba Vágvölgyi, and András Szekeres. 2018. "Effects of Different Cultivation Parameters on the Production of Surfactin Variants by a Bacillus subtilis Strain" Molecules 23, no. 10: 2675. https://doi.org/10.3390/molecules23102675
APA StyleBartal, A., Vigneshwari, A., Bóka, B., Vörös, M., Takács, I., Kredics, L., Manczinger, L., Varga, M., Vágvölgyi, C., & Szekeres, A. (2018). Effects of Different Cultivation Parameters on the Production of Surfactin Variants by a Bacillus subtilis Strain. Molecules, 23(10), 2675. https://doi.org/10.3390/molecules23102675