A Study on Neonatal Intake of Oleanolic Acid and Metformin in Rats (Rattus norvegicus) with Metabolic Dysfunction: Implications on Lipid Metabolism and Glucose Transport

 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Growth Development of the Rats
2.2. Saturated Free Fatty Acid
2.3. Monounsaturated Free Fatty Acids
2.4. Polyunsaturated Free Fatty Acid
2.5. Effects of Neonatal OA and MET Treatment on the Expression of Glut-4
2.6. Effects of Neonatal OA and MET Treatment on the Expression of Nrf-1
2.7. Effects of Neonatal OA and MET Treatment on the Expression of Acc-1
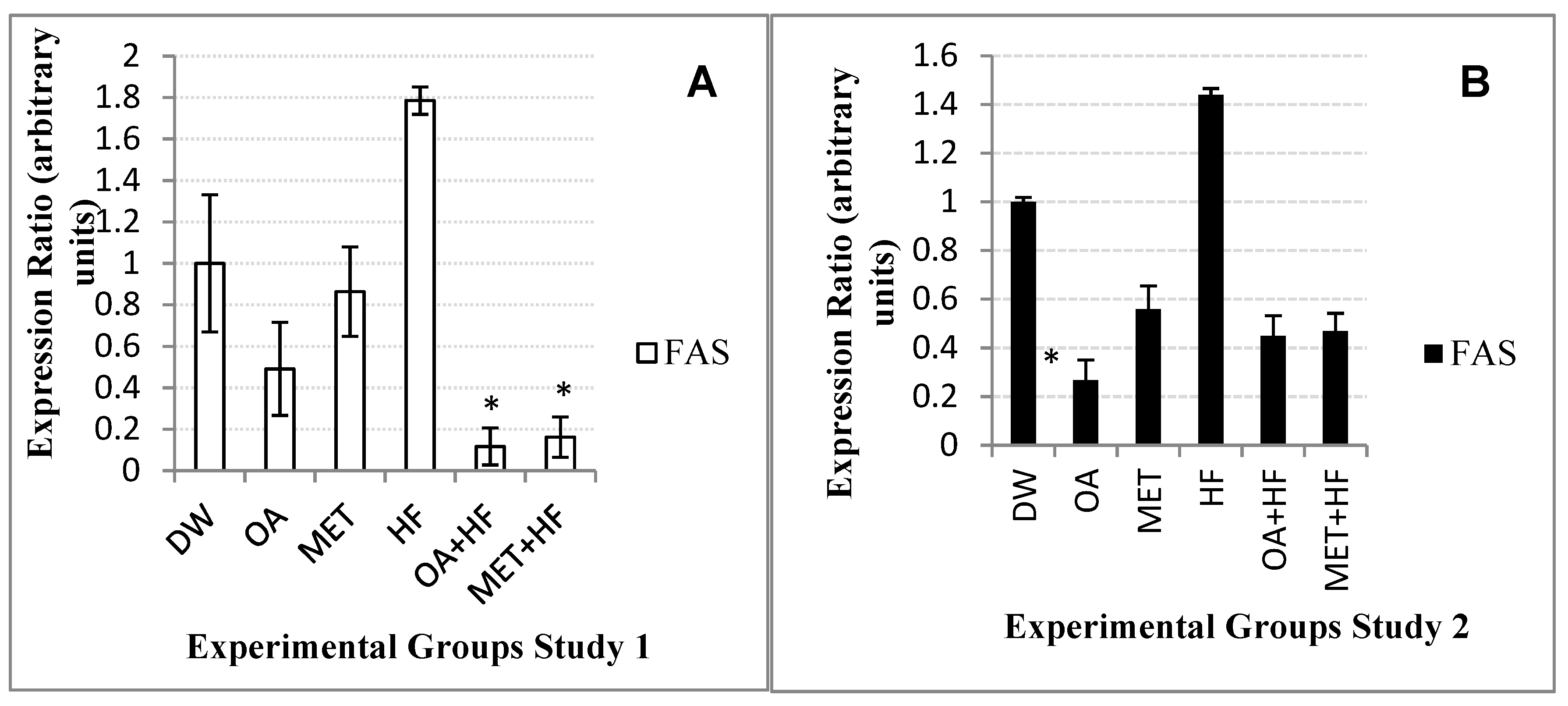
2.8. Effects of Neonatal OA and MET Treatment on the Expression of Fas
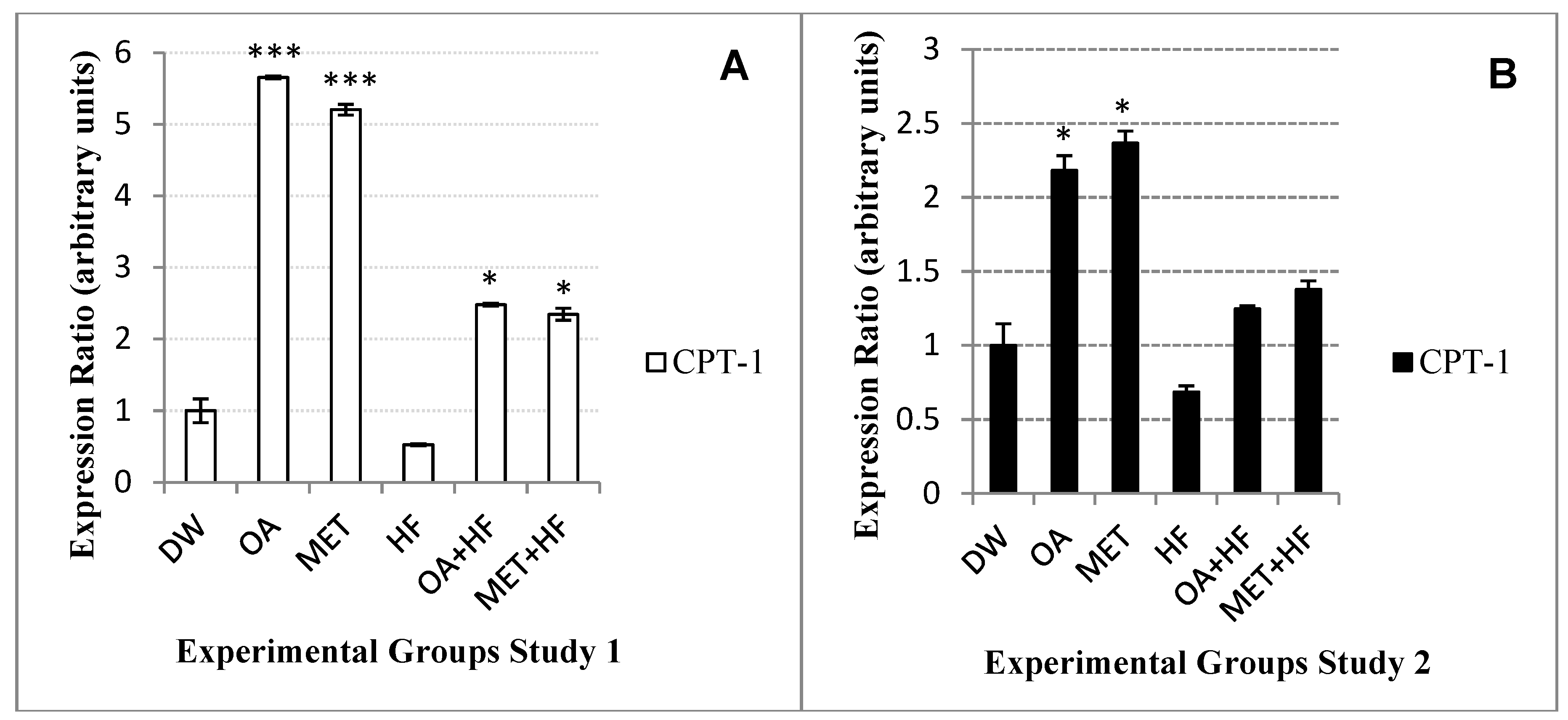
2.9. Effects Neonatal OA and MET Treatment on Expression of Cpt-1
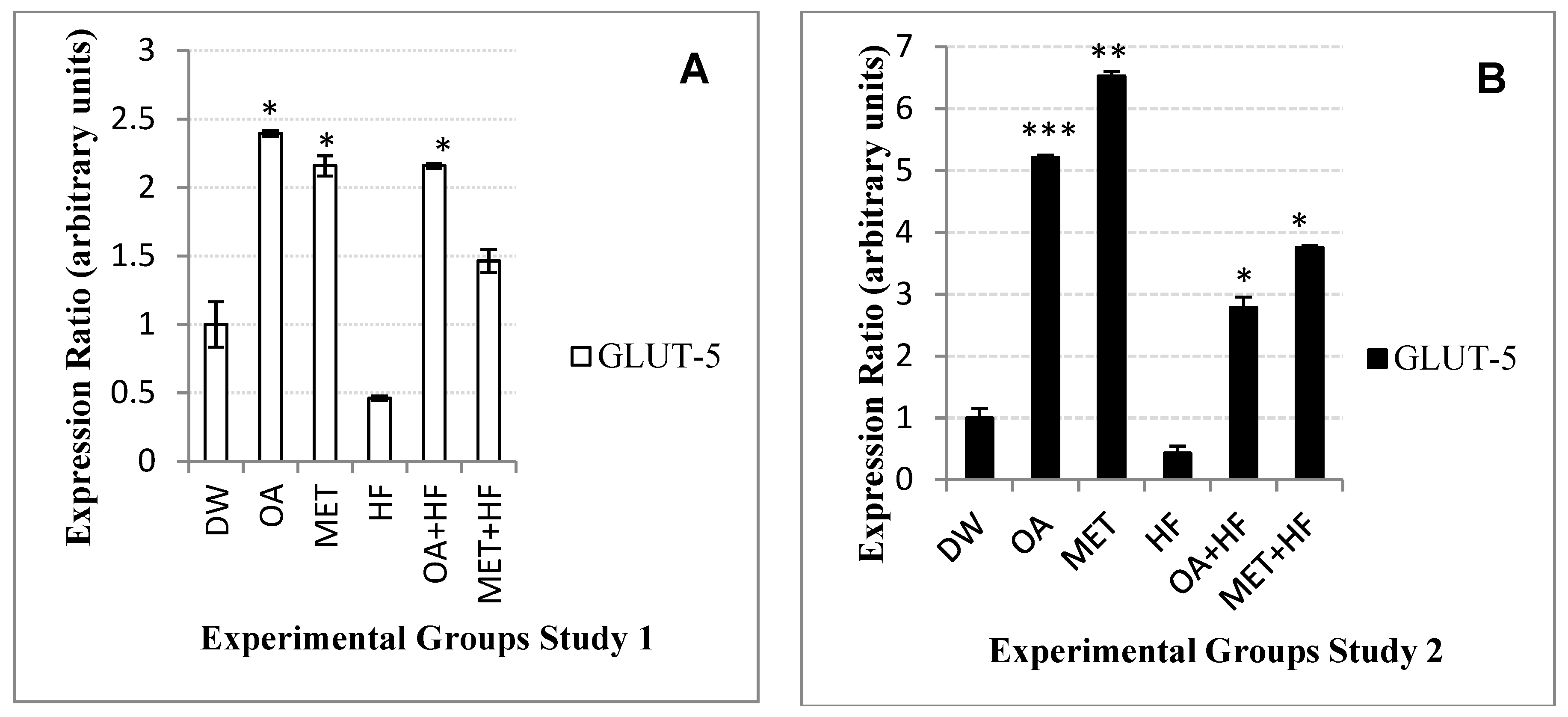
2.10. Effects Neonatal OA and MET Treatment on Expression of Glut-5
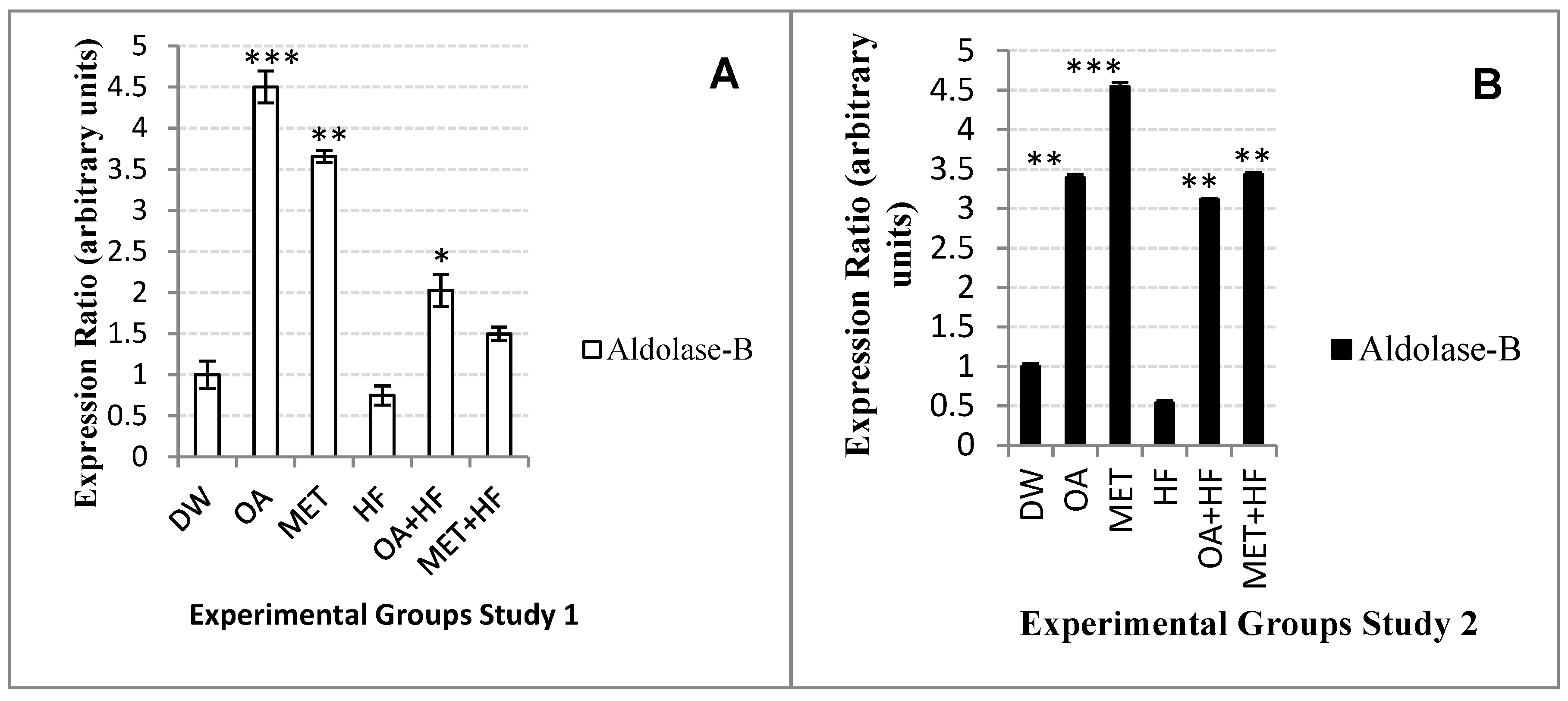
2.11. Effects of Neonatal OA and MET Treatment on the Expression of Aldolase-b
3. Discussion
4. Materials and Methods
4.1. Ethical Clearance
4.2. Experimental Design
4.3. Terminal Procedure and Sample Collection
4.4. Lipid Extraction, Purification and Identification by Gas Chromatography-Mass Spectrometry
4.5. RNA Extraction
4.6. Gel Electrophoresis
4.7. cDNA Synthesis
4.8. Quantitative Real Time PCR (RT-qPCR)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luo, M.; Luo, P.; Zhang, Z.; Payne, K.; Watson, S.; Wu, H.; Tan, H.; Ding, Y.; Sun, W.; Yin, X.; et al. Zinc delays the progression of obesity-related glomerulopathy in mice via down-regulating P38 MAPK-mediated inflammation. J. Obes. 2016, 24, 1244–1256. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Gomez, C.I.; Rocha-Guzman, N.E.; Gallegos-Infante, J.A.; Moreno-Jimenez, M.R.; Vazquez-Cabral, B.D.; Gonzalez-Laredo, R.F. Plants with potential use on obesity and its complications. EXCLI J. 2015, 14, 809–831. [Google Scholar] [PubMed]
- Groop, L. Genetics of the metabolic syndrome. Br. J. Nutr. 2000, 83, 39–48. [Google Scholar] [CrossRef]
- Roche, H.M.; Phillips, C.; Gibney, M.J. The metabolic syndrome: The crossroads of diet and genetics. Proc. Nutr. Soc. 2005, 64, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.A.; Katz, R. n-3 long-chain polyunsaturated fatty acids in type 2 diabetes: A review. J. Am. Diet. Assoc. 2005, 105, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Malik, V.S. Sugar-sweetened beverages and risk of obesity and Type 2 diabetes: Epidemiologic evidence. J. Physiol. Behav. 2010, 100, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Vance, D.E.; Vance, J.E. Biochemistry of Lipids, Lipoproteins and Membranes, 4th ed.; Elsevier Science: New York, NY, USA, 2002. [Google Scholar]
- Castellano, J.M.; Guinda, A.; Delgado, T.; Rada, M.; Jose, A.C. Biochemical basis of the antidiabetic activity of Oleanolic acid and related pentacyclic triterpenes. Diebetes 2013, 62, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yupeng, C.; Abdelkader, D.; Hasaan, H.S.; Liu, J. Combination therapy with Oleanolic acid and Metformin as a synergistic treatment for diabetes. J. Diabetes Res. 2015, 2015, 973287. [Google Scholar] [CrossRef] [PubMed]
- Ayeleso, T.B.; Matumba, M.G.; Mukwevho, E. Oleanolic acid and its derivatives: Biological activities and therapeutic potential in chronic diseases. Molecules 2017, 22, 1915. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujji, N.; et al. Role of AMP-activated protein kinase in mechanisms of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, J.M.; Kliewer, S.A.; Paulik, M.A.; Plunket, K.D.; Lehmann, J.M.; Weiel, J.E. Effects of troglitazone and metformin on glucose and lipid metabolism. Biochem. Pharmacol. 1997, 54, 801–808. [Google Scholar] [CrossRef]
- De Melo, C.L.; Queiroz, M.G.; Fonseca, S.G.; Bizerra, A.M.; Lemos, T.L.; Melo, T.S.; Santos, F.A.; Rao, V.S. Oleanolic acid, a naturaltriterpenoid improves blood glucose tolerance in normal mice and ameliorates visceral obesity in mice fed a high-fat diet. Chem. Biol. Interact. 2010, 185, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.E.; Bucarey, J.L.; Espinosa, A. Muscle lipid metabolism: Role of lipid droplets and metabolism. J. Diabetes Res. 2017, 2017, 1789395. [Google Scholar] [CrossRef] [PubMed]
- Steyn, K.; Fourie, J.; Temple, N. Chronic Diseases of Lifestyle in South Africa Since: 1995–2005; Technical Report; South African Medical Research Council: Cape Town, South Africa, 2006; pp. 65–74. [Google Scholar]
- Le, K.-A.; Ith, M.; Kreis, R.; Faeh, D.; Bortolotti, M.; Tran, C.; Boesch, C.; Tappy, L. Fructose overconsumption causes dyslipidemia and ectopic lipid deposition in healthy subjects with and without a family history of type 2 diabetes. Am. J. Clin. Nutr. 2009, 89, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Mark, A.H.; Varman, T.S. The sweet path to metabolic demise: Fructose and lipid synthesis. Trends Endocrinol. Metab. 2016, 27, 719–730. [Google Scholar]
- Voet, D.; Voet, J.G. Biochemistry, 3rd ed.; Wiley: New York, NY, USA, 2004. [Google Scholar]
- Im, S.; Kwon, S.; Kim, T.; Kim, H.; Ahn, Y. Regulation of glucose Transporter type 4 isoform gene expression in muscle and adipocytes. IUBMB Life 2007, 59, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Rabie, E.M.; Heeba, G.H.; Abouzied, M.M.; Khalifa, M.M.A. Comparative effects of Aliskiren and Telmisartan in high fructose diet-induced metabolic syndrome in rats. Eur. J. Pharmacol. 2015, 760, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Kain, J.; Corvalan, C.; Lera, L.; Galvan, M.; Uauy, R. Accelerated growth in early life and obesity in preschool Chilean children. J. Obes. 2009, 17, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.; Corvalan, C.; Gallehuillos, B.; Kain, J.; Gonzalez, L.; Uauy, R.; Garmendia, M.L.; Mericq, V. The effects of pre-pregnancy BMI and maternal factors on the timing of adiposity rebound in offspring. J. Obes. 2016, 24, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Rolland-Cachera, M.F.; Deheeger, M.; Bellisle, F.; Semple, M.; Guilloud-Bataille, M.; Patois, E. Adiposity rebound in children: A simple indicator for predicting obesity. Am. J. Clin. Nutr. 1984, 39, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Mukwevho, E.; Joseph, J.S. Calmodulin dependent protein kinase II activation by exercise regulates saturated and unsaturated fatty acids and improves some metabolic syndrome markers. Life Sci. 2014, 111, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Adugna, S.; Alemu, L.A.M.; Kelemu, T.; Tekola, H.; Kibret, B.; Genet, S. Medical Biochemistry; Ethiopia Public Health Training Initiative, The Carter Center, The Ethiopia Ministry of Health, The Ethiopia Ministry of Education: Addis Ababa, Ethiopia, 2004.
- Delarue, J.; Magnan, C. Free fatty acids and insulin resistance. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Welters, H.J.; Tadayyon, M.; Scarpello, J.H.B.; Smith, S.A.; Morgan, N.G. Mono-unsaturated fatty acids protect against β-cell apoptosis induced by saturated fatty acids, serum withdrawal or cytokine exposure. FEBS Lett. 2004, 560, 103–108. [Google Scholar] [CrossRef]
- Klip, A.; Volchuk, A.; He, L.; Tsakiridis, T. The glucose transporters of Skeletal muscle. Cell Dev. Biol. 1996, 7, 229–237. [Google Scholar] [CrossRef]
- Caponi, P.W.; Lehnen, A.M.; Pinto, G.H.; Borges, J.; Markoski, M.; Machado, U.F.; Schaan, B.D. Aerobic exercise training induces metabolic benefits in rats with betabolic syndrome independent of dietary changes. Clinics 2013, 68, 1010–1017. [Google Scholar] [CrossRef]
- Gould, G.W.; Holman, G.D. The glucose transport transporter family:structure, function and tissue-specific expression. Biochemistry 1993, 295, 329–341. [Google Scholar] [CrossRef]
- Huang, S.; Czech, M. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef] [PubMed]
- O’neill, H.M.; Lally, S.; Galic, S.; Thomas, M.; Azizi, P.D.; Fullerton, M.D.; Smith, B.K.; Pulinilkunnil, T.; Chen, Z.; Samaan, M.C.; et al. AMPK phosphorylation of ACC-2 is required for skeletal muscle fatty acid oxidation and insulin sensitivity in mice. Diabetologia 2014, 57, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, L.P.; Fisher, J.S. Skeletal muscle insulin resistance: Role of fatty acid metabolism and exercise. Phys. Ther. 2008, 88, 1279–1296. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.; Ren, J.M.; Young, L.H.; Pypaert, M.; Mu, J.; Birnbaum, M.J.; Shulman, G.I. AMP kinase is required for mitochondrial biogenesis in skeletal muscle in response to chronic energy deprivation. Proc. Natl. Acad. Sci. USA 2002, 99, 15983–15987. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, M.R. Normal roles for dietary fructose in carbohydrate metabolism. J. Nutr. 2014, 6, 3117–3129. [Google Scholar] [CrossRef] [PubMed]
- Nyakudya, T.T.; Mukwevho, E.; Nkomozepi, P.; Erlwanger, K.H. Neonatal intake of oleanolic acid attenuates the subsequent development of high fructose diet-induced non-alcoholic fatty liver disease in rats. J. Dev. Orig. Health Dis. 2018, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Musabayane, C.T.; Mahlalela, N.; Shode, F.O.; Ojewole, J.A.O. Effects of Syzygium cordatum (Hochst.) [Myrtaceae] leaf extract on plasma glucose and hepatic glycogen in streptozotocin-induced diabetic rats. Ethnopharmacology 2005, 97, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H.S. A simple method for the isolation and Purification of the total lipids from animal tissues. J. Biol. 1956, 226, 497–509. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | Induction Body Mass (g) | Terminal Body Mass (g) |
|---|---|---|
| DW | 15.11 ± 1.54 | 17.01 ± 1.71 |
| OA | 16.60 ± 1.22 | 18.60 ± 0.86 |
| MET | 15.79 ± 1.42 | 16.75 ± 1.85 |
| HF | 16.46 ± 1.42 | 18.15 ± 1.31 |
| OA + HF | 16.14 ± 2.26 | 17.78 ± 2.52 |
| MET + HF | 16.24 ± 1.96 | 17.01 ± 1.71 |
| Experimental Groups | Study 1 (Mean ± SD) | Study 2 (Mean ± SD) |
|---|---|---|
| DW | 2.30 ± 1.72 | 1.29 ± 0.77 |
| OA | 1.01 ± 1.04 | 0.75 ± 0.89 * |
| MET | 0.75 ± 0.89 * | 0.38 ± 0.23 ** |
| HF | 4.70 ± 7.19 * | 1.92 ± 1.31 * |
| OA + HF | 0.56 ± 0.48 ** | 0.37 ± 0.17 ** |
| MET + HF | 0.24 ± 0.19 *** | 0.20 ± 0.16 *** |
| Experimental Groups | Study 1 (Mean (Ra) ± SD) | Study 2 (Mean (Ra) ± SD) |
|---|---|---|
| DW | 0.00 ± 0.00 | 0.00 ± 0.00 |
| OA | 1.55 ± 1.03*** | 1.39 ± 0.26*** |
| MET | 0.00 ± 0.00 | 0.67 ± 0.47*** |
| HF | 0.00 ± 0.00 | 0.00 ± 0.00 |
| OA + HF | 0.09 ± 0.08* | 0.17 ± 0.03* |
| MET + HF | 0.00 ± 0.00 | 0.06 ± 0.03 |
| Experimental Groups | Study 1 (Mean (Ra) ± SD) | Study 2 (Mean (Ra) ± SD) |
|---|---|---|
| DW | 0.35 ± 0.33 | 0.43 ± 0.58 |
| OA | 2.96 ± 3.12 *** | 1.78 ± 1.06 *** |
| MET | 1.07 ± 0.60 *** | 0.75 ± 0.58 |
| HF | 0.08 ± 0.09 * | 0.00 ± 0.00 |
| OA + HF | 0.43 ± 0.58 | 0.24 ± 0.17 |
| MET + HF | 0.51 ± 0.53 | 0.20 ± 0.15 |
| Internal Standard Mix (IS) | |||
|---|---|---|---|
| Weight (mg) | Methanol (µL) | Water (µL) | IS (µL) |
| 50 | 400 | 75 | 50 |
| After Homogenization | |||
| Water (µL) | Chloroform (µL) | Total polar (~µL) | Total apolar (~µL) |
| 200 | 400 | 725 | 400 |
| Gene For: | Primers Orientation | |
|---|---|---|
| Forward 5′–3′ | Reverse 3′–5′ | |
| cpt-1 | GCA AAC TGG ACC GAG AAG AG | TCC ATG AGG GAT GGA CTC TC |
| nrf-1 | GTT GGA TCC CTC TCA CCC ATT G | CCA AGT CGA GAC TTA ATT CC |
| acc-1 | AGG AGG GAA GGG AAT CAG AA | TGT GCT GCA GGA AGA TTG AC |
| Fas | GCT TTG CTG CCG TGT CCT TCT | GTG TCT GCT GGG GTC CTC GTT |
| glut-4 | GCA GCG AGT GAC TGG AAC A | CCA GCC ACG TTG CAT TGT AG |
| glut-5 | CCA GAG AAG CAT GGA GCA | AGG ATG ACC CAA AGG CAG |
| Aldolase-b | GCC ACC TCA CAC AGC TTC TG | TCG GTG AGC CAT GAT GAC A |
| Actin | AGC CAT GTA CGT AGC CAT CC | CTC TCA GCT GTG GTGGTG AA |
| Gapdh | GCA CAG TCA AGG CCG AGA AT | GCC TTC TCC ATG GTG GTG AA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molepo, M.; Ayeleso, A.; Nyakudya, T.; Erlwanger, K.; Mukwevho, E. A Study on Neonatal Intake of Oleanolic Acid and Metformin in Rats (Rattus norvegicus) with Metabolic Dysfunction: Implications on Lipid Metabolism and Glucose Transport. Molecules 2018, 23, 2528. https://doi.org/10.3390/molecules23102528
Molepo M, Ayeleso A, Nyakudya T, Erlwanger K, Mukwevho E. A Study on Neonatal Intake of Oleanolic Acid and Metformin in Rats (Rattus norvegicus) with Metabolic Dysfunction: Implications on Lipid Metabolism and Glucose Transport. Molecules. 2018; 23(10):2528. https://doi.org/10.3390/molecules23102528
Chicago/Turabian StyleMolepo, Mmahiine, Ademola Ayeleso, Trevor Nyakudya, Kennedy Erlwanger, and Emmanuel Mukwevho. 2018. "A Study on Neonatal Intake of Oleanolic Acid and Metformin in Rats (Rattus norvegicus) with Metabolic Dysfunction: Implications on Lipid Metabolism and Glucose Transport" Molecules 23, no. 10: 2528. https://doi.org/10.3390/molecules23102528
APA StyleMolepo, M., Ayeleso, A., Nyakudya, T., Erlwanger, K., & Mukwevho, E. (2018). A Study on Neonatal Intake of Oleanolic Acid and Metformin in Rats (Rattus norvegicus) with Metabolic Dysfunction: Implications on Lipid Metabolism and Glucose Transport. Molecules, 23(10), 2528. https://doi.org/10.3390/molecules23102528