Measuring Artificial Sweeteners Toxicity Using a Bioluminescent Bacterial Panel

 , , ,
, , , 

Abstract
1. Introduction
2. Results
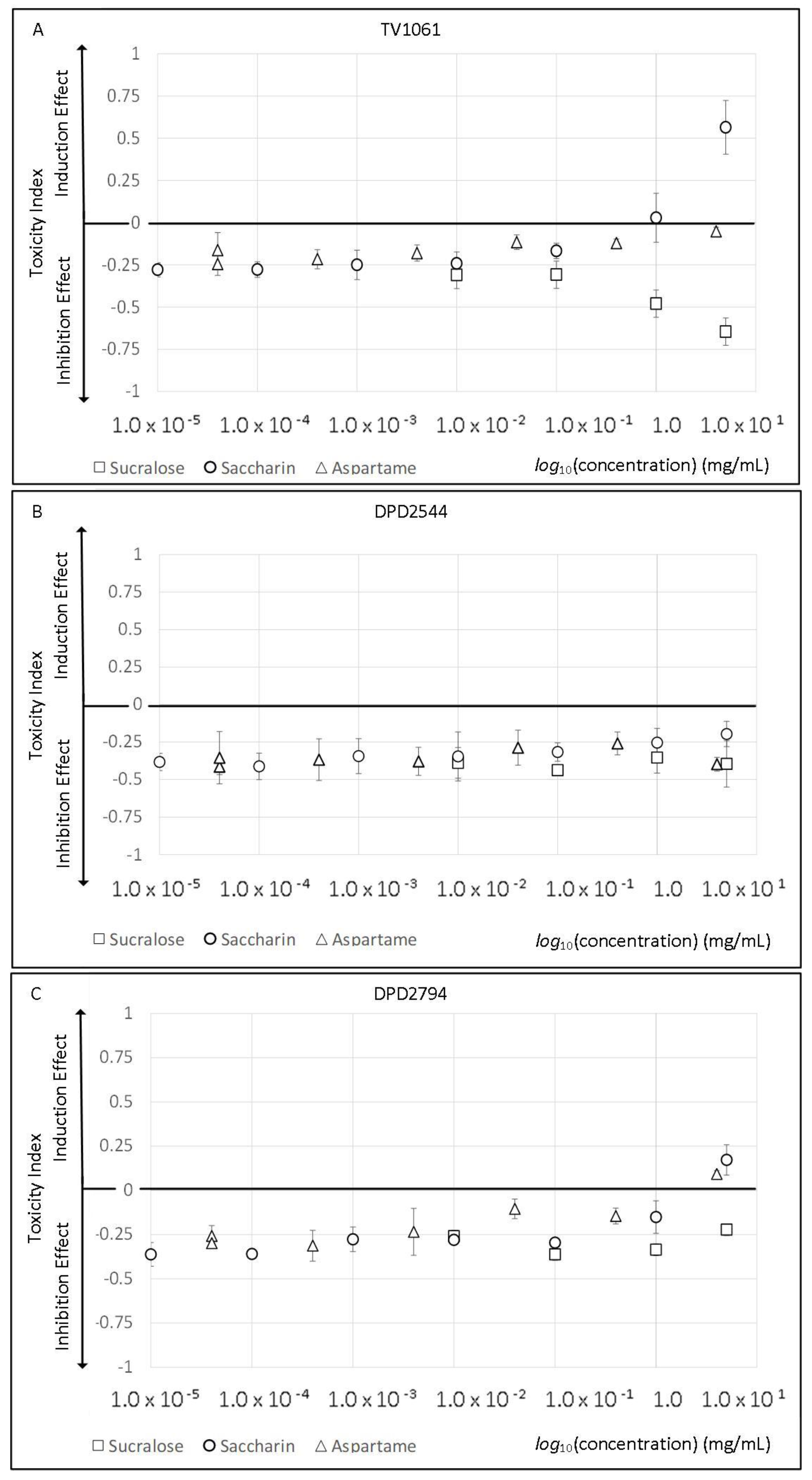
2.1. Artificial Sweeteners Toxicity and Viability Effect
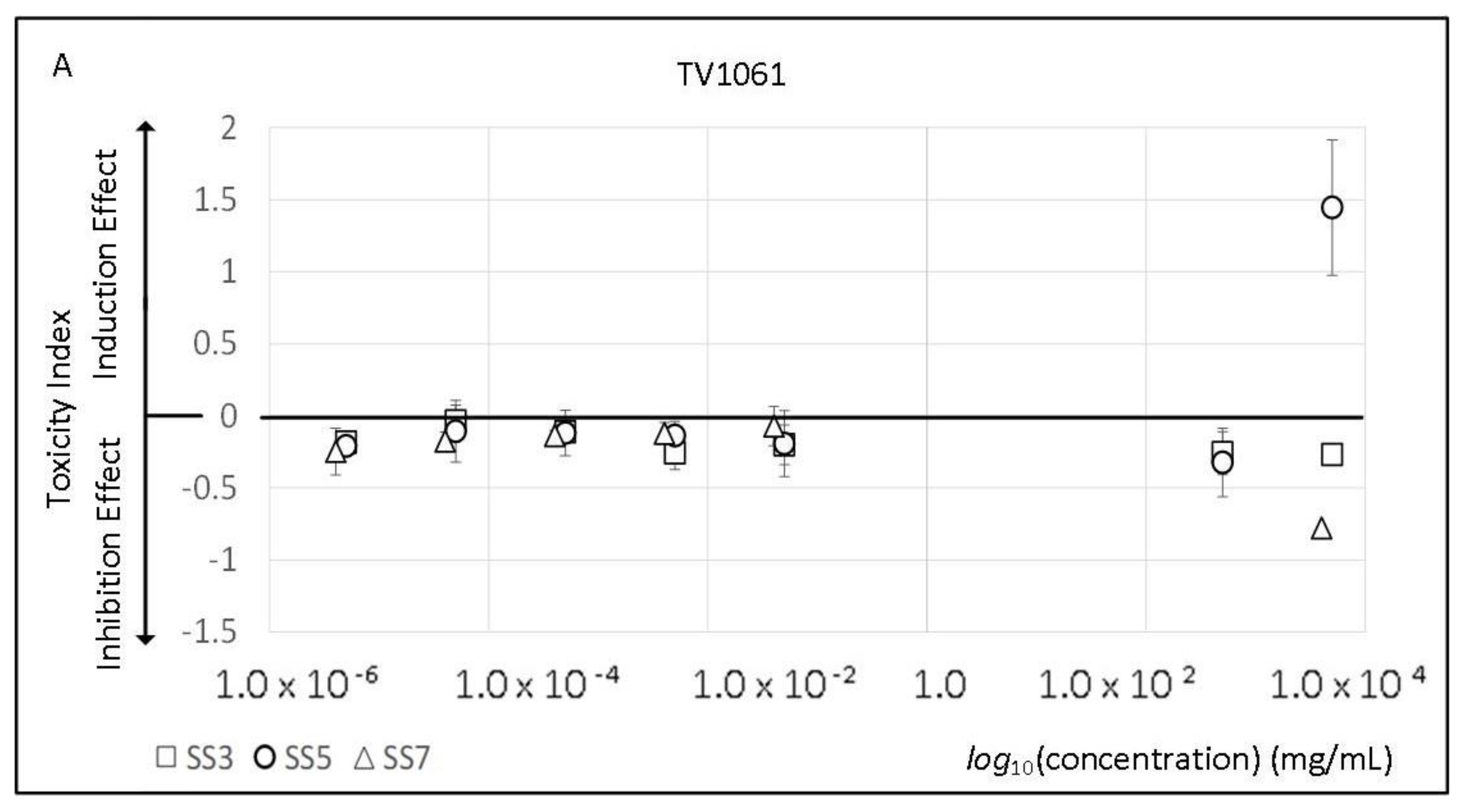
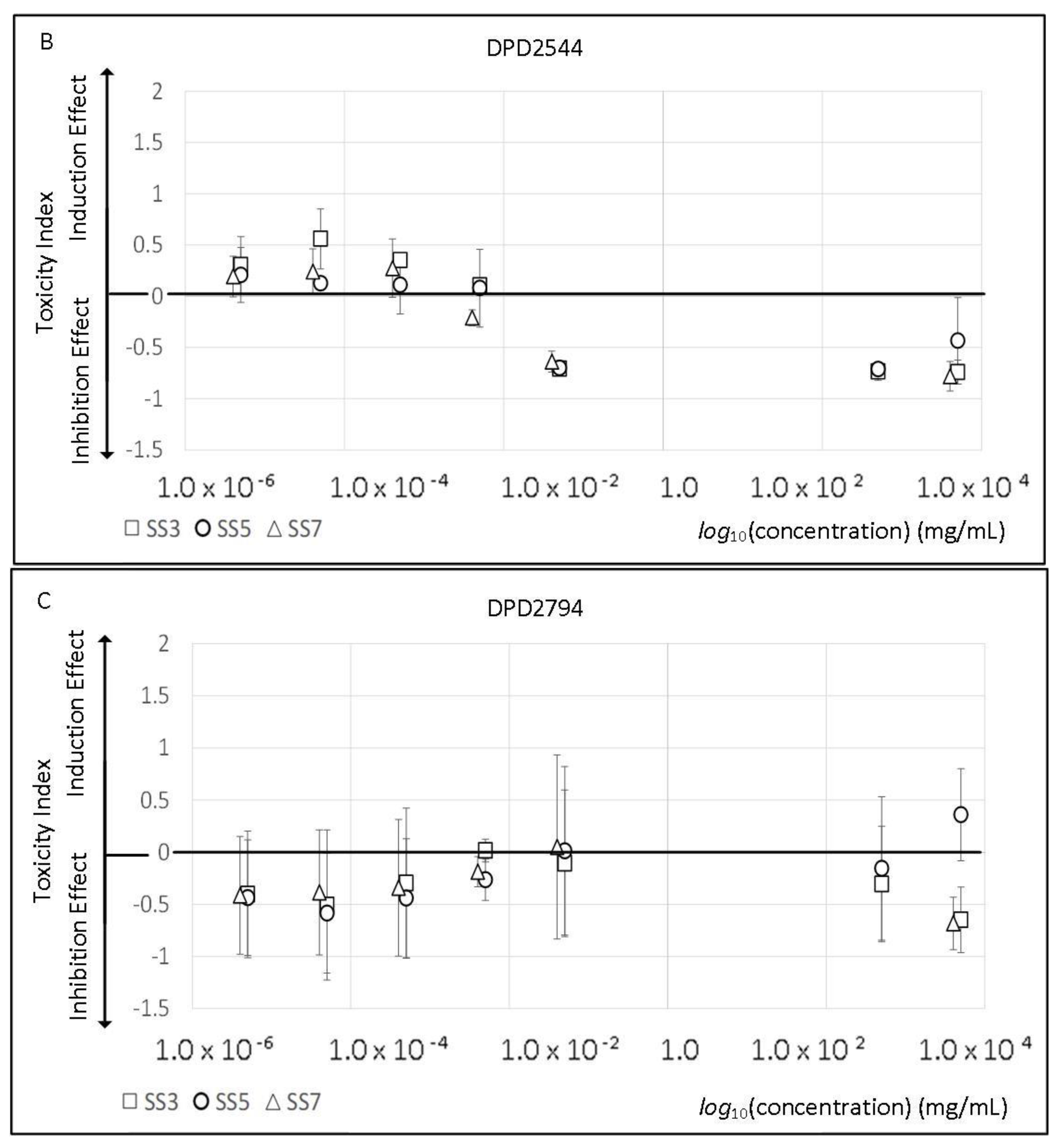
2.2. Sport Supplements Toxicity and Viability Effect
3. Discussion
3.1. Artificial Sweeteners’ Toxicity and Viability Effect
3.2. Sport Supplements Toxicity and Viability Effect
4. Materials and Methods
4.1. Materials
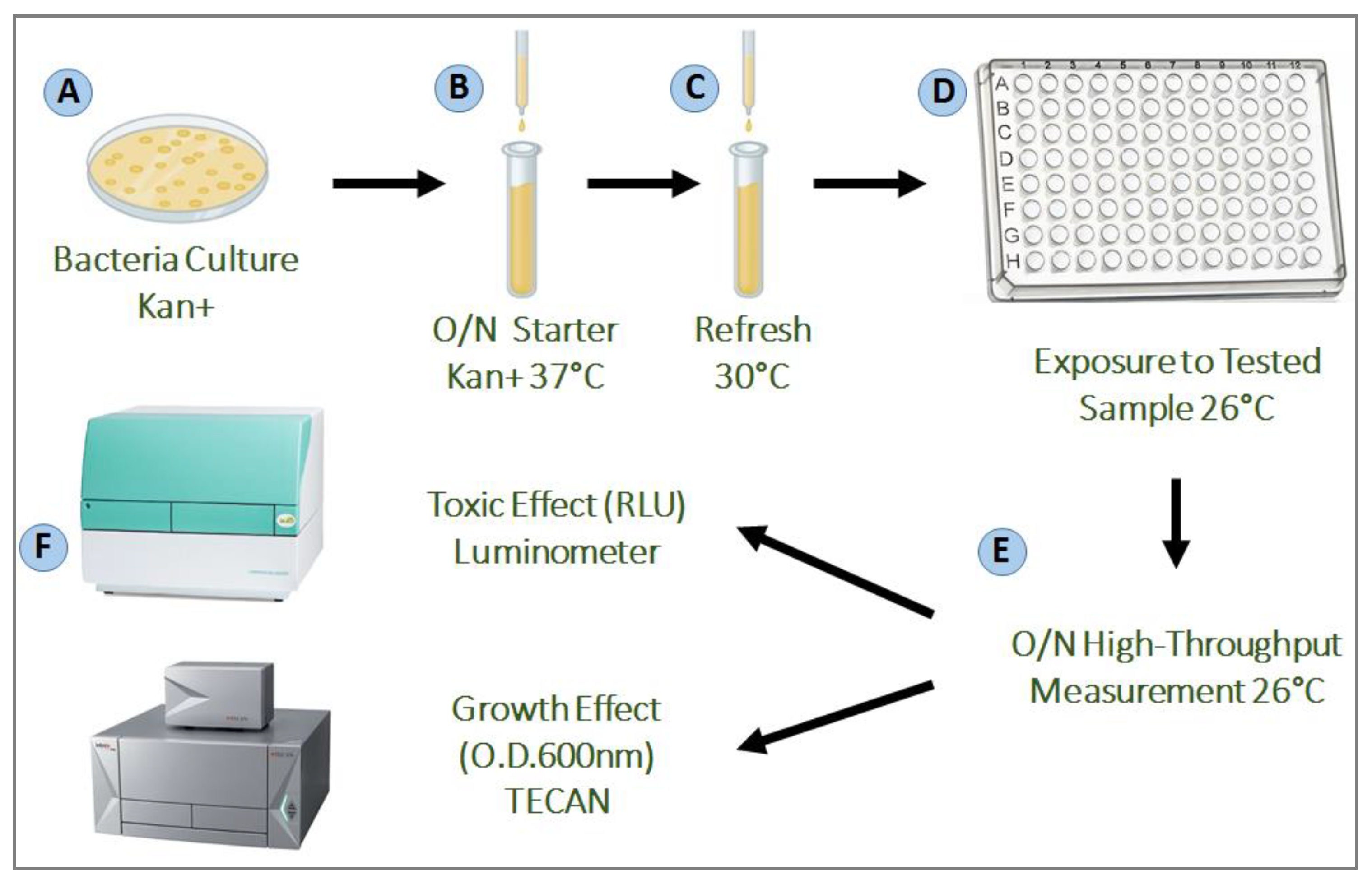
4.2. Bioluminescent Bacteria from E. coli
4.3. Growth Conditions
4.4. Bioluminescence Assay
4.5. Growth Assay
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swithers, S.E. Not-so-healthy sugar substitutes? Curr. Opin. Behav. Sci. 2016, 9, 106–110. [Google Scholar] [CrossRef] [PubMed]
- FDA, High-Intensity Sweeteners U.S. Food and Drug Administration. Available online: https://www.fda.gov/food/ingredientspackaginglabeling/foodadditivesingredients/ucm397716.htm (accessed on 19 May 2014).
- EFSA, Sugars and Sweeteners European Food Safety Authority. Available online: https://ec.europa.eu/jrc/en/health-knowledge-gateway/promotion-prevention/nutrition/sugars-sweeteners (accessed on 30 January 2018).
- Sylvetsky, A.C.; Rother, K.I. Trends in the consumption of low-calorie sweeteners. Physiol. Behav. 2016, 164, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Lugasi, A. Safety of intensive sweetener. Orvosi Hetil. 2016, 157 (Suppl. 1), 14–28. [Google Scholar] [CrossRef]
- Ranchordas, M.K. Nutrition for adventure racing. Sports Med. 2012, 42, 915–927. [Google Scholar] [PubMed]
- Ko, S.Y. Electrolyte Drink. U.S. Patent 06/154,259, 29 May 1980. [Google Scholar]
- Kampinga, J.; Colaco, C. Compositions for Use in Rehydration and Nutrition during Athletic Exercise and Methods of Making Same. U.S. Patent 08/899,012, 23 July 1997. [Google Scholar]
- Ross, N.; Reyman, J. Chewable Electrolyte Tablet. U.S. Patent 10/954,874, 30 March 2006. [Google Scholar]
- Stone, K.R. Cartilage Enhancing Food Supplements and Methods of Preparing the Same. U.S. Patent 09/598,634, 21 June 2000. [Google Scholar]
- Howard, A.N.; Harris, R. Compositions Containing Creatine in Suspension, 2001. U.S. Patent 09/419,922, 2 June 1999. [Google Scholar]
- Bakal, A.I.; Crossman, T.L. Use of Lactose-Hydrolyzed Whey in Chewing Gum. U.S. Patent 06/472,734, 7 March 1983. [Google Scholar]
- Badalov, C. Super Sweet Sugar Crystals and Syrups for Health and Method. U.S. Patent 11/487,933, 17 Janurary 2008. [Google Scholar]
- Nuralam, M. Nutritional Supplement Composition Comprising Creatine and Method for Making the Same. U.S. Patent 11/604,562, 29 May 2008. [Google Scholar]
- Tandel, K.R. Sugar substitutes: Health controversy over perceived benefits. J. Pharmacol. Pharmacother. 2011, 2, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.M.V.; Mahajan, S.; Tandon, V.R. Artificial sweeteners. JK Sci. 2012, 14, 1–4. [Google Scholar]
- Marinovich, M.; Galli, C.L.; Bosetti, C.; Gallus, S.; La Vecchia, C. Aspartame, low-calorie sweeteners and disease: Regulatory safety and epidemiological issues. Food Chem. Toxicol. 2013, 60, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Ahmed, K.; Froghi, S.; Dasgupta, P. Systematic review of the relationship between artificial sweetener consumption and cancer in humans: Analysis of 599,741 participants. Int. J. Clin. Pract. 2015, 69, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Schernhammer, E.S.; Bertrand, K.A.; Birmann, B.M.; Sampson, L.; Willett, W.C.; Feskanich, D. Consumption of artificial sweetener- and sugar-containing soda and risk of lymphoma and leukemia in men and women. Am. J. Clin. Nutr. 2012, 96, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Curhan, G.C. Associations of sugar and artificially sweetened soda with albuminuria and kidney function decline in women. Clin. J. Am. Soc. Nephrol. 2011, 6, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Gardener, H.R.T.; Markert, M.; Wright, C.B.; Elkind, M.S.V.; Sacco, R.L. Diet soft drink consumption is associated with an increased risk of vascular events in the northern manhattan study. J. Gen. Intern. Med. 2012, 27, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, G.L.; Kanders, B.S.; Lavin, P.T.; Keller, S.D.; Whatley, J. The effect of aspartame as part of a multidisciplinary weight-control program on short- and long-term control of body weight. Am. J. Clin. Nutr. 1997, 65, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Raben, A.; Vasilaras, T.H.; Moller, A.C.; Astrup, A. Sucrose compared with artificial sweeteners: Different effects on ad libitum food intake and body weight after 10 wk of supplementation in overweight subjects. Am. J. Clin. Nutr. 2002, 76, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Hampton, T. Sugar substitutes linked to weight gain. JAMA 2008, 299, 2137–2138. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, S.S.; Rother, K.I. Sucralose, a synthetic organochlorine sweetener: Overview of biological issues. J. Toxicol. Environ. Health Part B Crit. Rev. 2013, 16, 399–451. [Google Scholar] [CrossRef] [PubMed]
- DeNoon, D.J.R.b.C.G.M.M. Drink More Diet Soda, Gain More Weight? Overweight Risk Soars 41% with Each Daily Can of Diet Soft Drink. Web MD Medical News 2005. Available online: https://www.webmd.com/diet/news/20050613/drink-more-diet-soda-gain-more-weight#1 (accessed on 11 February 2011).
- Swithers, S.E.; Davidson, T.L. A role for sweet taste: Calorie predictive relations in energy regulation by rats. Behav. Neurosci. 2008, 122, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Darby, A.C.; Hall, N.; Nau, A.; Bravo, D.; Shirazi-Beechey, S.P. Dietary supplementation with lactose or artificial sweetener enhances swine gut lactobacillus population abundance. Br. J. Nutr. 2014, 111, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Darby, A.C.; Shirazi-Beechey, S.P. Low calorie sweeteners and gut microbiota. Physiol. Behav. 2016, 164, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Lange, F.T.; Scheurer, M.; Brauch, H.-J. Artificial sweeteners—A recently recognized class of emerging environmental contaminants: A review. Anal. Bioanal. Chem. 2012, 403, 2503–2518. [Google Scholar] [CrossRef] [PubMed]
- Kokotou, M.G.; Asimakopoulos, A.G.; Thomaidis, N.S. Artificial sweeteners as emerging pollutants in the environment: Analytical methodologies and environmental impact. Anal. Methods 2012, 4, 3057–3070. [Google Scholar] [CrossRef]
- Sang, Z.; Jiang, Y.; Tsoi, Y.-K.; Leung, K.S.-Y. Evaluating the environmental impact of artificial sweeteners: A study of their distributions, photodegradation and toxicities. Water Res. 2014, 52, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.; Gawlik, B.M.; Boettcher, K.; Locoro, G.; Contini, S.; Bidoglio, G. Sucralose screening in european surface waters using a solid-phase extraction-liquid chromatographytriple quadrupole mass spectrometry method. J. Chromatogr. A 2009, 1216, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Buerge, I.J.; Buser, H.R.; Kahle, M.; Muller, M.D.; Poiger, T. Ubiquitous occurrence of the artificial sweetener acesulfame in the aquatic environment: An ideal chemical marker of domestic wastewater in groundwater. Environ. Sci. Technol. 2009, 43, 4381–4385. [Google Scholar] [CrossRef] [PubMed]
- Scheurer, M.; Brauch, H.J.; Lange, F.T. Analysis and occurrence of seven artificial sweeteners in German waste water and surface water and in soil aquifer treatment (sat). Anal. Bioanal. Chem. 2009, 394, 1585–1594. [Google Scholar] [CrossRef] [PubMed]
- Mawhinney, D.B.; Young, R.B.; Vanderford, B.J.; Borch, T.; Snyder, S.A. Artificial sweetener sucralose in U.S. Drinking water systems. Environ. Sci. Technol. 2011, 45, 8716–8722. [Google Scholar] [CrossRef] [PubMed]
- Van Stempvoort, D.R.; Roy, J.W.; Brown, S.J.; Bickerton, G. Artificial sweeteners as potential tracers in groundwater in urban environments. J. Hydrol. 2011, 401, 126–133. [Google Scholar] [CrossRef]
- Gan, Z.; Sun, H.; Feng, B.; Wang, R.; Zhang, Y. Occurrence of seven artificial sweeteners in the aquatic environment and precipitation of Tianjin, China. Water Res. 2013, 47, 4928–4937. [Google Scholar] [CrossRef] [PubMed]
- Stolte, S.; Steudte, S.; Schebb, N.H.; Willenberg, I.; Stepnowski, P. Ecotoxicity of artificial sweeteners and stevioside. Environ. Int. 2013, 60, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, C.R.; Boullata, J.; McCauley, L.A. The potential toxicity of artificial sweeteners. Aaohn J. 2008, 56, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Eltzov, E.; Ben-Yosef, D.Z.; Kushmaro, A.; Marks, R. Detection of sub-inhibitory antibiotic concentrations via luminescent sensing bacteria and prediction of their mode of action. Sen. Actuators B Chem. 2008, 129, 685–692. [Google Scholar] [CrossRef]
- Eltzov, E.; Marks, R.S. Fiber-optic based cell sensors. In Whole Cell Sensing Systems I: Reporter Cells and Devices; Springer: Berlin/Heidelberg, Germany, 2010; Volume 117, pp. 131–154. [Google Scholar]
- Nordeen, S.K. Luciferase reporter gene vectors for analysis of promoters and enhancers. Biotechniques 1988, 6, 454–458. [Google Scholar] [PubMed]
- Ivask, A.; Virta, M.; Kahru, A. Construction and use of specific luminescent recombinant bacterial sensors for the assessment of bioavailable fraction of cadmium, zinc, mercury and chromium in the soil. Soil Biol. Biochem. 2002, 34, 1439–1447. [Google Scholar] [CrossRef]
- Tauriainen, S.; Karp, M.; Chang, W.; Virta, M. Luminescent bacterial sensor for cadmium and lead. Biosens. Bioelectron. 1998, 13, 931–938. [Google Scholar] [CrossRef]
- Ivask, A.; Green, T.; Polyak, B.; Mor, A.; Kahru, A.; Virta, M.; Marks, R. Fibre-optic bacterial biosensors and their application for the analysis of bioavailable hg and as in soils and sediments from aznalcollar mining area in spain. Biosens. Bioelectron. 2007, 22, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Michelini, E.; Leskinen, P.; Virta, M.; Karp, M.; Roda, A. A new recombinant cell-based bioluminescent assay for sensitive androgen-like compound detection. Biosens. Bioelectron. 2005, 20, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Fine, T.; Leskinen, P.; Isobe, T.; Shiraishi, H.; Morita, M.; Marks, R.S.; Virta, M. Luminescent yeast cells entrapped in hydrogels for estrogenic endocrine disrupting chemical biodetection. Biosens. Bioelectron. 2006, 21, 2263–2269. [Google Scholar] [CrossRef] [PubMed]
- Belkin, S.; Smulski, D.R.; Vollmer, A.C.; Van Dyk, T.K.; LaRossa, R.A. Oxidative stress detection with Escherichia coli harboring a katg’: Lux fusion. Appl. Environ. Microbiol. 1996, 62, 2252–2256. [Google Scholar] [PubMed]
- Gu, M.B.; Min, J.; Kim, E.J. Toxicity monitoring and classification of endocrine disrupting chemicals (EDCs) using recombinant bioluminescent bacteria. Chemosphere 2002, 46, 289–294. [Google Scholar] [CrossRef]
- Choi, S.H.; Gu, M.B. A portable toxicity biosensor using freeze-dried recombinant bioluminescent bacteria. Biosens. Bioelectron. 2002, 17, 433–440. [Google Scholar] [CrossRef]
- Bechor, O.; Smulski, D.R.; Van Dyk, T.K.; LaRossa, R.A.; Belkin, S. Recombinant microorganisms as environmental biosensors: Pollutants detection by Escherichia coli bearing fabA’:: Lux fusions. J. Biotechnol. 2002, 94, 125–132. [Google Scholar] [CrossRef]
- Durand, M.J.; Thouand, G.; Dancheva-Ivanova, T.; Vachon, P.; DuBow, M. Specific detection of organotin compounds with a recombinant luminescent bacteria. Chemosphere 2003, 52, 103–111. [Google Scholar] [CrossRef]
- Polyak, B.; Bassis, E.; Novodvorets, A.; Belkin, S.; Marks, R.S. Optical fiber bioluminescent whole-cell microbial biosensors to genotoxicants. Water Sci. Technol. 2000, 42, 305–311. [Google Scholar] [CrossRef]
- Polyak, B.; Geresh, S.; Marks, R.S. Synthesis and characterization of a biotin-alginate conjugate and its application in a biosensor construction. Biomacromolecules 2004, 5, 389–396. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.F. Microbial biosensors. Biosens. Bioelectron. 2001, 16, 337–353. [Google Scholar] [CrossRef]
- Van der Meer, J.R.; Belkin, S. Where microbiology meets microengineering: Design and applications of reporter bacteria. Nat. Rev. Microbiol. 2010, 8, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Zygler, A.; Wasik, A.; Namieśnik, J. Analytical methodologies for determination of artificial sweeteners in foodstuffs. TrAC Trends Anal. Chem. 2009, 28, 1082–1102. [Google Scholar] [CrossRef]
- Shankar, P.; Ahuja, S.; Sriram, K. Non-nutritive sweeteners: Review and update. Nutrition 2013, 29, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Palmnas, M.S.A.; Cowan, T.E.; Bomhof, M.R.; Su, J.; Reimer, R.A.; Vogel, H.J.; Hittel, D.S.; Shearer, J. Low-dose aspartame consumption differentially affects gut microbiota-host metabolic interactions in the diet-induced obese rat. PLoS ONE 2014, 9, e109841. [Google Scholar] [CrossRef] [PubMed]
- Rettig, S.; Tenewitz, J.; Ahearn, G.; Coughlin, C. Sucralose causes a concentration dependent metabolic inhibition of the gut flora bacteroides, B. fragilis and B. uniformis not observed in the firmicutes, E. faecalis and C. sordellii. FASEB J. 2014, 28, 1118. [Google Scholar]
- Abou-Donia, M.B.; El-Masry, E.M.; Abdel-Rahman, A.A.; McLendon, R.E.; Schiffman, S.S. Splenda alters gut microflora and increases intestinal p-glycoprotein and cytochrome p-450 in male rats. J. Toxicol. Environ. Health Part A 2008, 71, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Renneberg, R.; Riedel, K.; Scheller, F. Microbial sensor for aspartame. Appl. Microbiol. Biotechnol. 1985, 21, 180–181. [Google Scholar] [CrossRef]
- Labare, M.; Alexander, M. Microbial cometabolism of sucralose, a chlorinated disaccharide, in environmental samples. Appl. Microbiol. Biotechnol. 1994, 42, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Bowen, W. The influence of sucralose on bacterial metabolism. J. Dent. Res. 1990, 69, 1480–1484. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, B.A.; Roberts, A.; Nestmann, E.R. Critical review of the current literature on the safety of sucralose. Food Chem. Toxicol. 2017, 106, 324–355. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, D.A.; Phil Comer, C.; Wayne Stargel, W. Food consumption and body weight changes with neotame, a new sweetener with intense taste: Differentiating effects of palatability from toxicity in dietary safety studies. Regul. Toxicol. Pharmacol. 2003, 38, 124–143. [Google Scholar] [CrossRef]
- Carocho, M.; Morales, P.; Ferreira, I.C.F.R. Sweeteners as food additives in the xxi century: A review of what is known, and what is to come. Food Chem. Toxicol. 2017, 107, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Hanina, M.; Shahril, M.H.; Asyikin, I.I.N.; Jalil, A.A.; Salina, M.; Maryam, M.; Rosfarizan, M. Extracellular protein secreted by Bacillus subtilis atcc21332 in the presence of streptomycin sulfate. World Acad. Sci. Eng. Technol. Int. J. Biol. Biomol. Agric. Food Biotechnol. Eng. 2014, 8, 820–824. [Google Scholar]
- Yang, D.; Chen, B. Determination of neotame in beverages, cakes and preserved fruits by column-switching high-performance liquid chromatography. Food Addit. Contam. Part A 2010, 27, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Chakrabarti, J. In vivo cytogenetic studies on mice exposed to acesulfame-k—A non-nutritive sweetener. Food Chem. Toxicol. 1997, 35, 1177–1179. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Mukherjee, A.; Chakrabarti, J. In vivo cytogenetic studies on blends of aspartame and acesulfame-k. Food Chem. Toxicol. 2000, 38, 75–77. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Raychaudhuri, U.; Chakraborty, R. Artificial sweeteners—A review. J. Food Sci. Technol. 2014, 51, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, D.; Gatehouse, D. Aspartame: A review of genotoxicity data. Food Chem. Toxicol. 2015, 84, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.; Ghoshal, S.; Mukherjee, A. Genotoxicity testing of low-calorie sweeteners: Aspartame, acesulfame-k, and saccharin. Drug Chem. Toxicol. 2008, 31, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Weihrauch, M.R.; Diehl, V. Artificial sweeteners—Do they bear a carcinogenic risk? Ann. Oncol. 2004, 15, 1460–1465. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Arnold, L.L.; Emerson, J.L. Safety of saccharin. Agro Food Ind. Hi Tech 2008, 19, 26–29. [Google Scholar]
- Eltzov, E.; Cohen, A.; Marks, R.S. Bioluminescent liquid light guide pad biosensor for indoor air toxicity monitoring. Anal. Chem. 2015, 87, 3655–3661. [Google Scholar] [CrossRef] [PubMed]
- Hakkila, K.; Green, T.; Leskinen, P.; Ivask, A.; Marks, R.; Virta, M. Detection of bioavailable heavy metals in eilatox-oregon samples using whole-cell luminescent bacterial sensors in suspension or immobilized onto fibre-optic tips. J. Appl. Toxicol. 2004, 24, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Eltzov, E.; Slobodnik, V.; Ionescu, R.E.; Marks, R.S. On-line biosensor for the detection of putative toxicity in water contaminants. Talanta 2015, 132, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Gu, M.B. A whole cell bioluminescent biosensor for the detection of membrane-damaging toxicity. Biotechnol. Bioprocess Eng. 1999, 4, 59–62. [Google Scholar] [CrossRef]
- Premkumar, J.R.; Lev, O.; Marks, R.S.; Polyak, B.; Rosen, R.; Belkin, S. Antibody-based immobilization of bioluminescent bacterial sensor cells. Talanta 2001, 55, 1029–1038. [Google Scholar] [CrossRef]
- Van Dyk, T.K.; Majarian, W.R.; Konstantinov, K.B.; Young, R.M.; Dhurjati, P.S.; LaRossa, R.A. Rapid and sensitive pollutant detection by induction of heat shock gene-bioluminescence gene fusions. Appl. Environ. Microbiol. 1994, 60, 1414–1420. [Google Scholar] [PubMed]
- Vollmer, A.C.; Belkin, S.; Smulski, D.R.; Van Dyk, T.K.; LaRossa, R.A. Detection of DNA damage by use of Escherichia coli carrying reca’::Lux, uvra’::Lux, or alka’::Lux reporter plasmids. Appl. Environ. Microbiol. 1997, 63, 2566–2571. [Google Scholar] [PubMed]
- Eltzov, E.; Marks, R.S.; Voost, S.; Wullings, B.A.; Heringa, M.B. Flow-through real time bacterial biosensor for toxic compounds in water. Sens. Actuators B Chem. 2009, 142, 11–18. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.D. The indigenous gastrointestinal microflora. Trends Microbiol. 1996, 4, 430–435. [Google Scholar] [CrossRef]
Sample Availability: Not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MLIC | MLIndC | MGIC | MGIndC | |
|---|---|---|---|---|---|
| Aspartame | TV1061 | N.E. | N.E. | N.E. | N.E. |
| DPD2544 | N.E. | N.E. | N.E. | N.E. | |
| DPD2794 | N.E. | 4 | N.E. | N.E. | |
| Sucralose | TV1061 | 1 | N.E. | 50 | N.E. |
| DPD2544 | 50 | N.E. | 50 | N.E. | |
| DPD2794 | 100 | N.E. | 50 | N.E. | |
| Saccharin | TV1061 | N.E. | 5 | 5 | N.E. |
| DPD2544 | N.E. | N.E. | N.E. | N.E. | |
| DPD2794 | N.E. | 5 | N.E. | N.E. | |
| Advantame | TV1061 | N.E. | N.E. | N.E. | 2 |
| DPD2544 | N.E. | N.E. | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | N.E. | |
| Neotame | TV1061 | N.E. | 2 | N.E. | N.E. |
| DPD2544 | 2 | N.E. | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | N.E. | |
| Ace-K | TV1061 | N.E. | N.E. | N.E. | N.E. |
| DPD2544 | N.E. | N.E. | N.E. | N.E. | |
| DPD2794 | N.E. | 10 | N.E. | N.E. |
| Strain | MLIC | MLIndC | MGIC | MGIndC | |
|---|---|---|---|---|---|
| SS1 | TV1061 | N.E. | 2000 | N.E. | N.E. |
| DPD2544 | 2 × 10−3 | 2 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | 2000 | N.E. | |
| SS2 | TV1061 | N.E. | N.E. | N.E. | N.E. |
| DPD2544 | 1 × 10−3 | 1 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | 1000 | N.E. | |
| SS3 | TV1061 | N.E. | N.E. | N.E. | 4000 |
| DPD2544 | 4 × 10−3 | 4 × 10−6 | N.E. | N.E. | |
| DPD2794 | 4000 | N.E. | N.E. | 4000 | |
| SS4 | TV1061 | N.E. | N.E. | N.E. | 5000 |
| DPD2544 | 5 × 10−3 | 5 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | 5000 | |
| SS5 | TV1061 | N.E. | 5000 | N.E. | N.E. |
| DPD2544 | 5 × 10−3 | 5 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | 5000 | 5000 | N.E. | |
| SS6 | TV1061 | N.E. | N.E. | N.E. | 3000 |
| DPD2544 | 3 × 10−3 | 3 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | N.E. | |
| SS7 | TV1061 | 5000 | 500 | 5000 | N.E. |
| DPD2544 | 5 × 10−3 | 5 × 10−6 | N.E. | N.E. | |
| DPD2794 | 5000 | N.E. | 5000 | N.E. | |
| SS8 | TV1061 | N.E. | N.E. | N.E. | 2000 |
| DPD2544 | 2 × 10−3 | 2 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | 2000 | |
| SS9 | TV1061 | N.E. | 3000 | N.E. | N.E. |
| DPD2544 | 3 × 10−3 | 3 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | N.E. | |
| SS10 | TV1061 | N.E. | N.E. | N.E. | 3000 |
| DPD2544 | 3 × 10−3 | 3 × 10−6 | N.E. | N.E. | |
| DPD2794 | N.E. | N.E. | N.E. | 3000 |
| Artificial Sweeteners Content | Recommended Amount for Consumption (1 oz = 30 mL) | Ingredients | |
|---|---|---|---|
| SS1 | Sucralose | 2 tablets (5 g), recommended to drink a lot of water | Creatine Hydrochloride, Cellulose, Dicalcium phosphate, Enteric Coating (Cellulose, Sodium Alginate, Medium Chain Triglycerides, Oleic and Stearic Acid), Natural Mint Flavor, Sucralose, Titanium Dioxide |
| SS2 | Acesulfame Potassium-K and Sucralose | 2 (7 g) to 8 (28 g) scoops in 8–10 oz per serving (2 scoops) | Black Tea Extract, Green Tea Extract, Green Coffee Extract, Micronized Taurine, Micronized l-Glutamine, Micronized l-Arginine, Micronized l-Leucine, Beta-Alanine (as CarnoSyn®), Micronized Citrulline, Micronized l-Isoleucine, Micronized l-Valine, Micronized l-Tyrosine, Micronized l-Histidine, Micronized l-Lysine, Micronized l-Phenylalanine, Micronized l-Threonine, Micronized l-Methionine Other Ingredients: Inulin, Acesulfame Potassium, Citric Acid, FD&C Red #40, Malic Acid, Natural and Artificial Flavors, Sucralose, Silion Dioxide |
| SS3 | Acesulfame Potassium-K and Sucralose | 1 (31 g) to 2 (62 g) scoops in 6–8 oz per scoop | Calcium, Cholesterol, Dietary Fibers, Potassium, Protein, Saturated Fat, Sodium, Sugars, Trans Fat Other Ingredients: Acesulfame Potassium, Cocoa (Processed with Alkali), Enzyme Blend (Aminogen®, Lactase), Lecithin, Natural and Artificial Flavors, Salt, Sucralose, Whey Protein Blend (Whey Protein Isolate, Whey Protein Concentrate, Whey Protein Hydrolysate), Xanthan Gum |
| SS4 | Sucralose | 1 (31 g) to 2 (62 g) scoops in 4–10 oz per scoop | Calcium, Cholesterol, proteins, Sodium, Saturated Fat, sugars, Trans Fat Other Ingredients: Citric Acid, FD&C Red #40 Lake, Lactase, Sucralose, Natural and Artificial Flavors, Soy Lecithin, Whey Protein Isolate, Whey Protein Concentrate, Whey Peptides |
| SS5 | Sucralose | 2 (9 g) to 6 (27 g) scoops in 10–12 oz per serving (2 scoops) | Caffeine, Green Tea Extract, Green Coffee Extract, Micronized Taurine, Micronized l-Glutamine, Micronized l-Arginine, Micronized l-Leucine, Beta-Alanine (as CarnoSyn®), Micronized Citrulline, Micronized l-Isoleucine, Micronized l-Valine, Micronized l-Tyrosine, Micronized l-Histidine, Micronized l-Lysine HCI, Micronized l-Phenylalanine, Micronized l-Threonine, Micronized l-Methionine Other Ingredients: Calcium Citrate, Calcium Silicate, Citric Acid, Gum Blend (Cellulose Gum, Xanthan Gum, Carrageenan), FD&C Blue #2, FD&C Red #40, Inulin, Lecithin, Malic Acid, Natural and Artificial Flavors, Silicon Dioxide, Sucralose, Tartaric Acid |
| SS6 | Acesulfame Potassium-K | 1 (29.4 g) rounded scoop in 4–10 oz | Calcium, Protein, Saturated Fat, Sodium, Sugars, Trans Fat Other Ingredients: Acesulfame Potassium, Aminogen®, Lactase, Lecithin, Natural and Artificial Flavor, Whey Protein Isolate, Whey Protein Concentrate, Whey Peptides |
| SS7 | Acesulfame Potassium-K and Sucralose | 1 (49 g) to 2 (98 g) scoops in 6 oz per scoop | Alpha lipoic acid, Calcium, Citric Acid, Creatine Monohydrate, Creatine HCI, Dicalcium Phosphate, Dextrose, l-alanine, l-Isoleucine, l-Leucine, l-Valine, Magnesium Oxide, Potassium, Sodium, Sugar, Taurine, Vitamin B6, Vitamin C, Vitamin B12 Other Ingredients: Acesulfame-Potassium, Dextrose, Ethyl-Cellulose, Glucose Polymers, Modcarb™ [Oat Bran, Amaranth, Quinoa, Buckwheat, Millet, Chia], Natural Flavors, Calcium Silicate, Salt, Sucralose, FD&C Yellow No. 6, Soy Lecithin, FD&C Yellow No. 5, Waxy Maize (Corn Starch), (Cluster Dextrin) |
| SS8 | Acesulfame Potassium-K and Sucralose | 1 (34 g) scoop in 6 oz water or skim milk | Calcium, Cholesterol, Dietary Fiber, Iron, Protein, Saturated Fat, Sodium, Sugar Other Ingredients: Acesulfame-Potassium, Alkalized Cocoa Powder, Calcium Carbonate, Gum Blend (Cellulose Gum, Xanthan Gum, Carrageenan), Natural and Artificial Flavors, Salt, Soy Lecithin, Sucralose, Sunflower-based Creamer (Sunflower oil, Corn syrup solids, Sodium Caseinate, Mono-Diglycerides, Dipotassium Phosphate, Tocopherols), Tricalcium Phosphate, Whey Protein Isolate, Whey Peptides, whey Protein Concentrate |
| SS9 | Acesulfame Potassium-K and Sucralose | 1 (32.4 g) to 2 (64.8 g) scoops in 8–12 oz | Calcium, Cholesterol, Dietary Fiber, Iron, Potassium, Protein, Saturated Fat, Sodium, Sugar, Trans Fat, Vitamin A, Vitamin C Other Ingredients: Acesulfame-Potassium, Amino Matrix (l-Glycine, l-Taurine, BCAAs (Leucine, Iso-Leucine, Valine), l-Glutamine), Flax Seed Oil, Glucose Polymers, Lactase, Natural and Artificial Flavors, Sucralose, Sea Salt, Suspension Matrix (Xanthan Gum, Cellulose Gum, Guar Gum), Whey Protein Concentrate, Whey Protein Isolate, Whey Protein Hydrolysate |
| SS10 | Acesulfame Potassium-K and Sucralose | 1 (34.9 g) to 2 (69.8 g) scoops in 8–12 oz | Calcium, Cholesterol, Dietary Fiber, Iron, Multi-level Amino Acid Growth Matrix, Potassium, Protein, Saturated Fat, Sodium, Trans Fat Other Ingredients: Alanine, Arginine, Aspartic Acid, BCAAs (l-Leucine, l-isoleucine, l-Glutamine, l-valine), Cystine, Digestive Enzyme Blend, Egg Albumen, Glycine, Histidine, Lactase, Lysine, Methionine, Micellar Casein, Partially-hydrolyzed Whey Concentrate, Phenylalanine, Proline, Protease, Serine, Tyrosine, Threonine, Tryptophan, Whey Protein Isolate, Whey Protein Concentrate |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harpaz, D.; Yeo, L.P.; Cecchini, F.; Koon, T.H.P.; Kushmaro, A.; Tok, A.I.Y.; Marks, R.S.; Eltzov, E. Measuring Artificial Sweeteners Toxicity Using a Bioluminescent Bacterial Panel. Molecules 2018, 23, 2454. https://doi.org/10.3390/molecules23102454
Harpaz D, Yeo LP, Cecchini F, Koon THP, Kushmaro A, Tok AIY, Marks RS, Eltzov E. Measuring Artificial Sweeteners Toxicity Using a Bioluminescent Bacterial Panel. Molecules. 2018; 23(10):2454. https://doi.org/10.3390/molecules23102454
Chicago/Turabian StyleHarpaz, Dorin, Loo Pin Yeo, Francesca Cecchini, Trish H. P. Koon, Ariel Kushmaro, Alfred I. Y. Tok, Robert S. Marks, and Evgeni Eltzov. 2018. "Measuring Artificial Sweeteners Toxicity Using a Bioluminescent Bacterial Panel" Molecules 23, no. 10: 2454. https://doi.org/10.3390/molecules23102454
APA StyleHarpaz, D., Yeo, L. P., Cecchini, F., Koon, T. H. P., Kushmaro, A., Tok, A. I. Y., Marks, R. S., & Eltzov, E. (2018). Measuring Artificial Sweeteners Toxicity Using a Bioluminescent Bacterial Panel. Molecules, 23(10), 2454. https://doi.org/10.3390/molecules23102454