
Applications of Magnetotactic Bacteria, Magnetosomes and Magnetosome Crystals in Biotechnology and Nanotechnology: Mini-Review

 , ,
, ,
Abstract

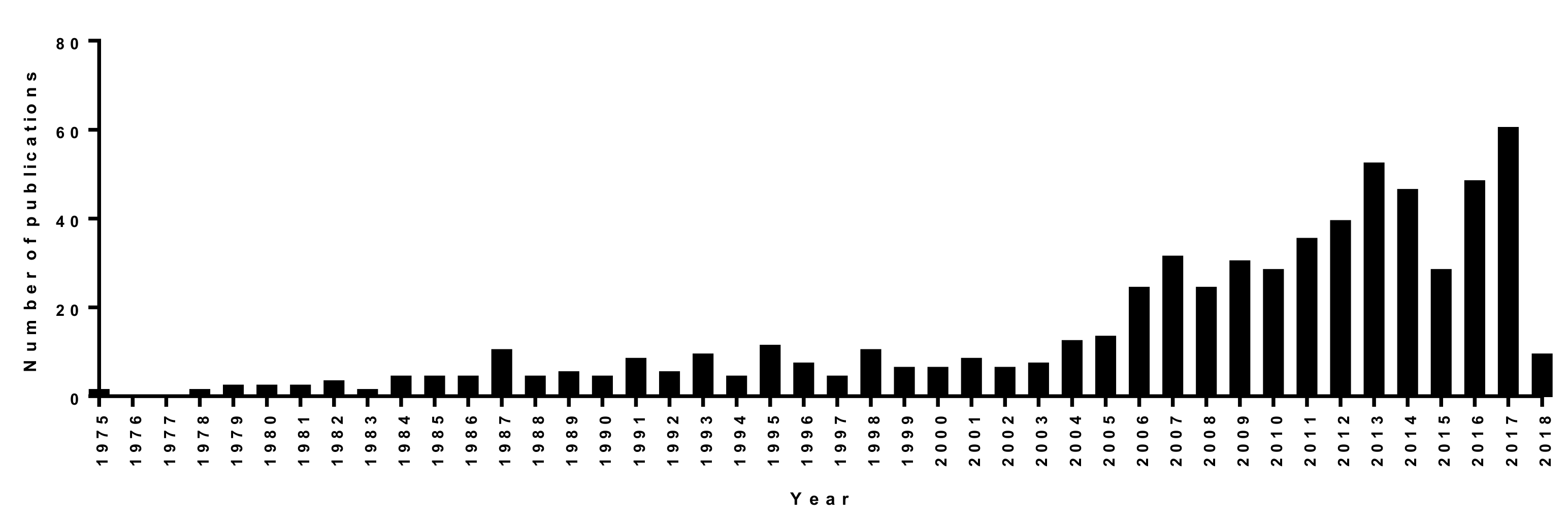
1. Introduction
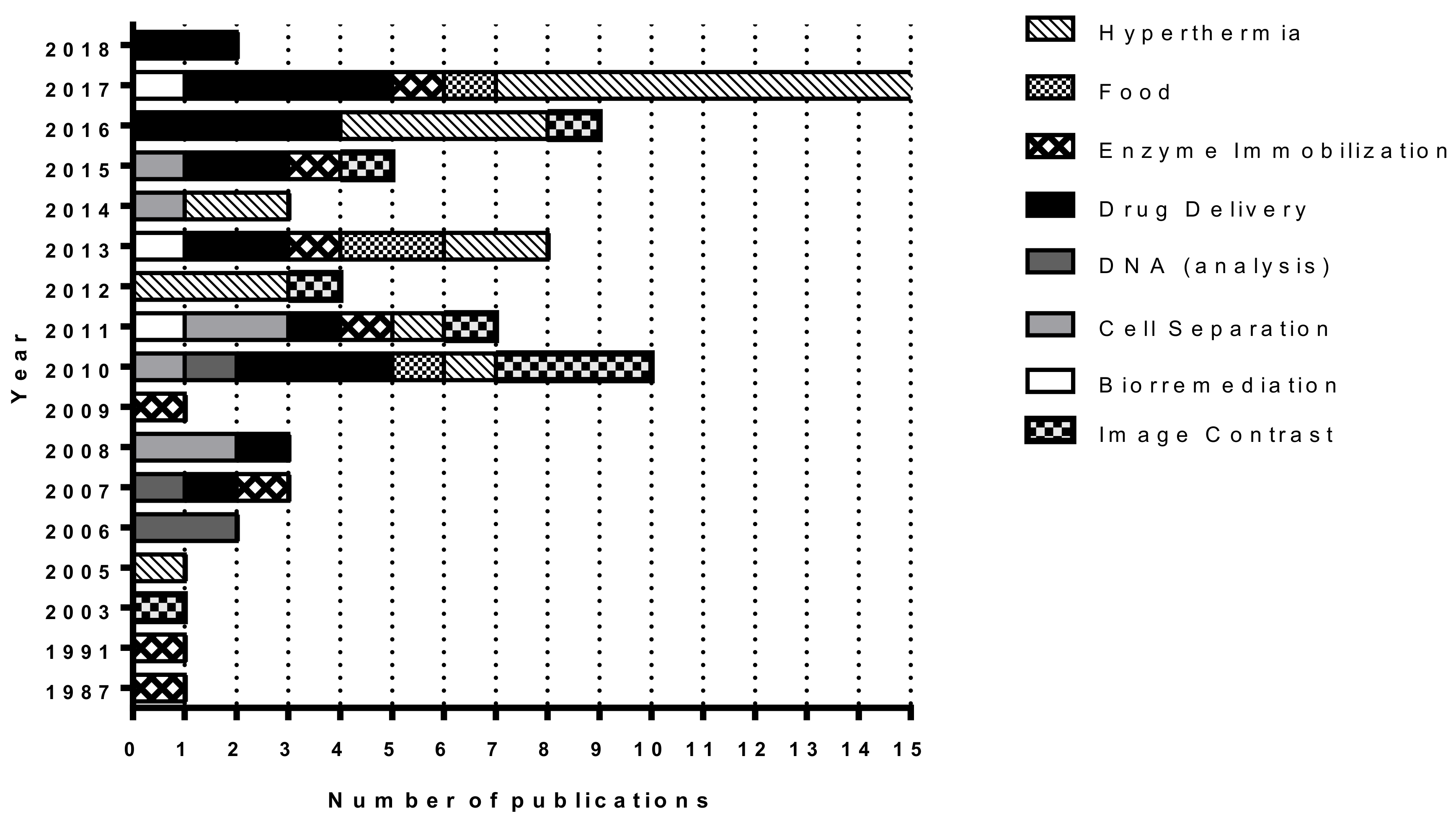
2. Results and Discussion
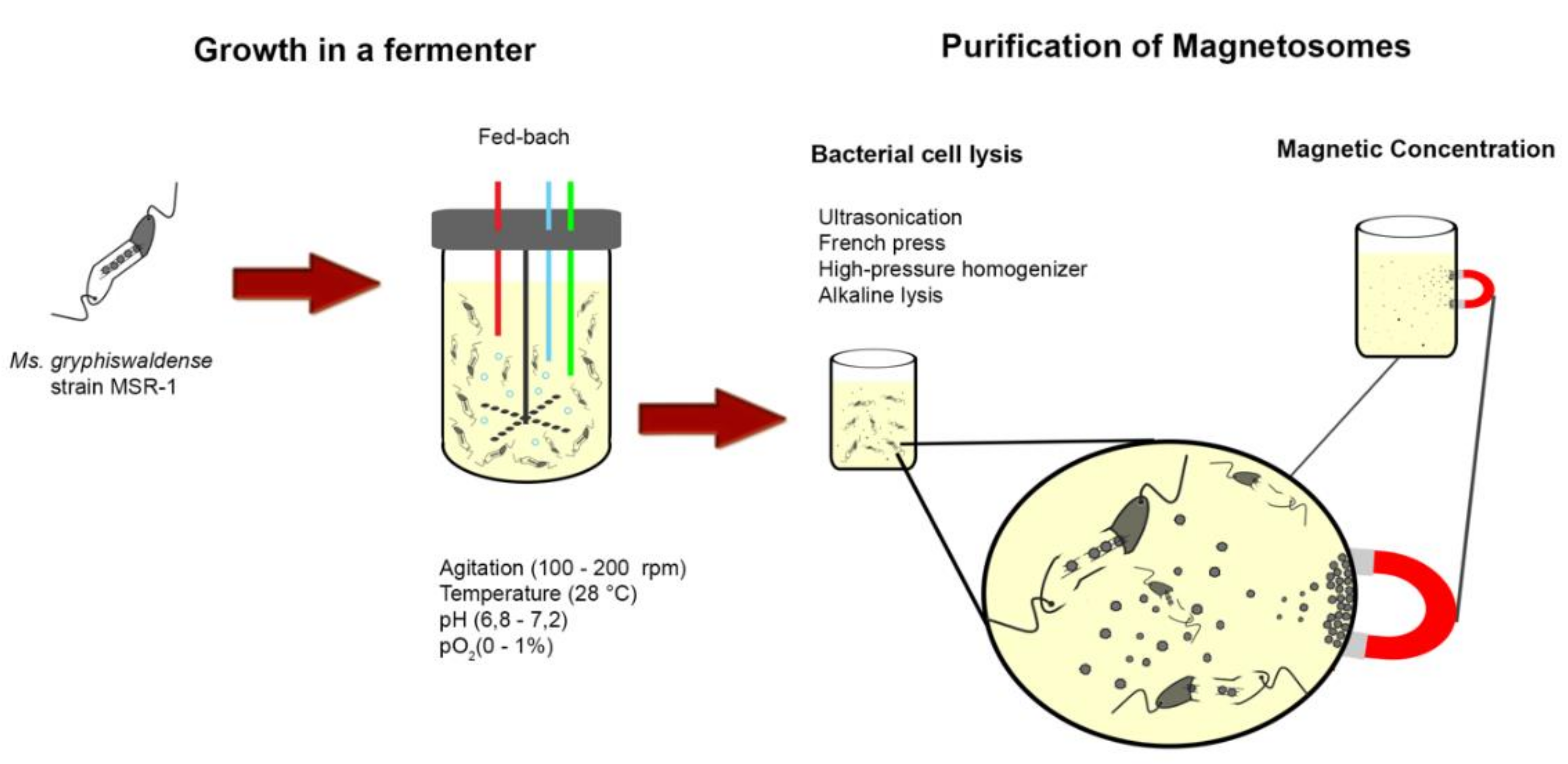
2.1. MTB and Magnetosomes Production
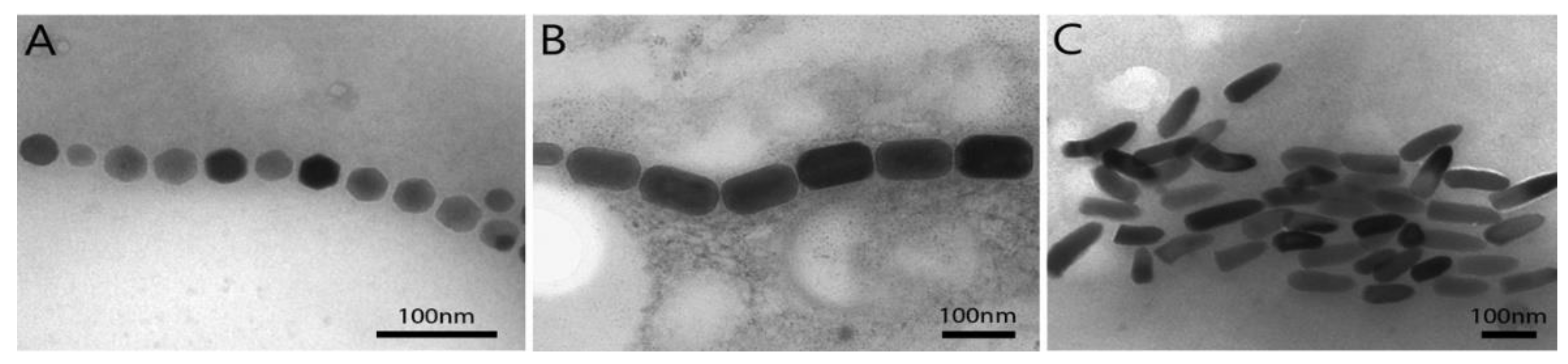
2.1.1. MTB and Magnetosomes
2.1.2. Magnetosome Production and Functionalization
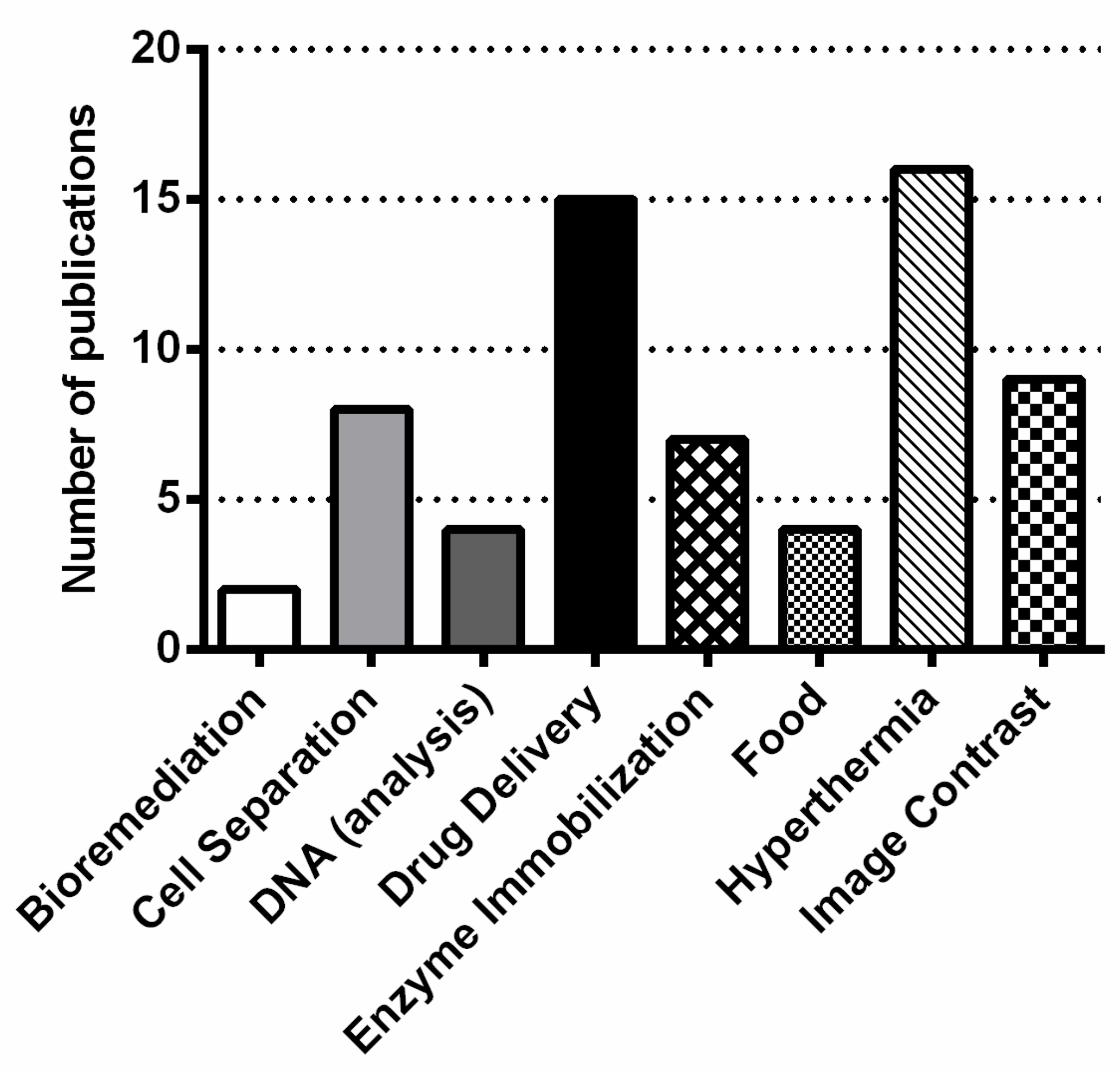
2.2. Applications of MTB and Magnetosomes
2.2.1. Applications of MTB
MTB in Drug Delivery
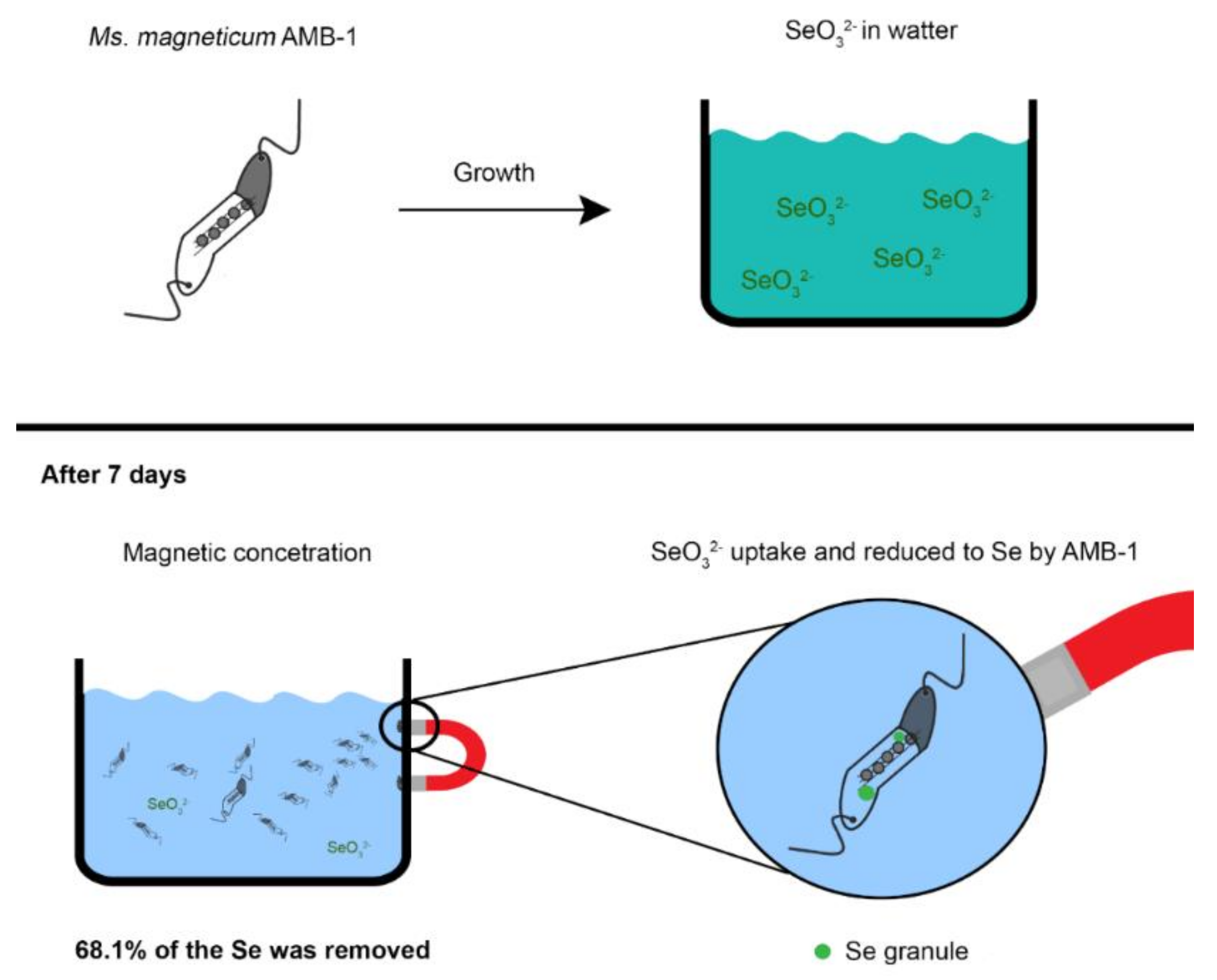
MTB in Bioremediation
Use of MTB in New, Novel Technologies
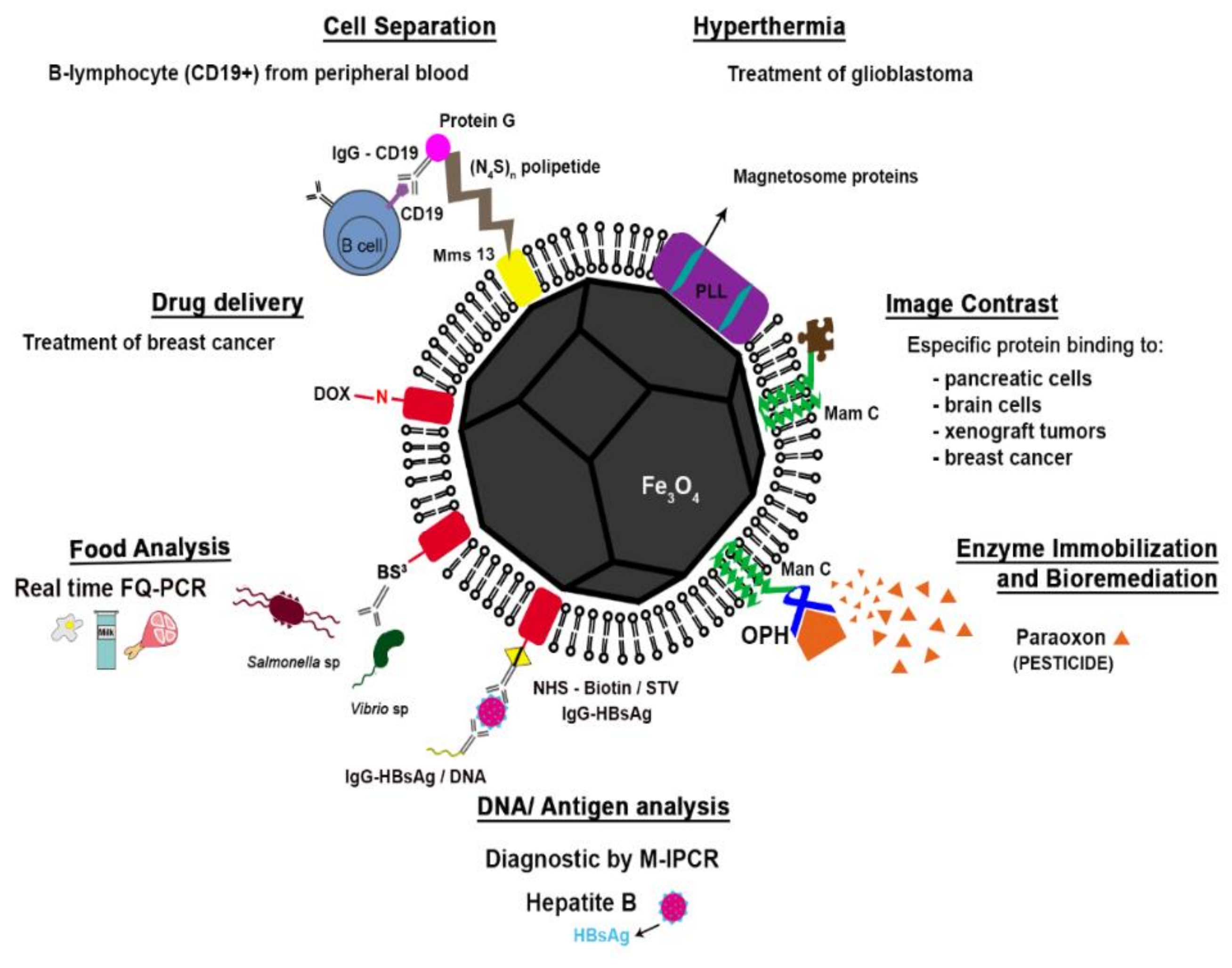
2.2.2. Applications of Magnetosomes
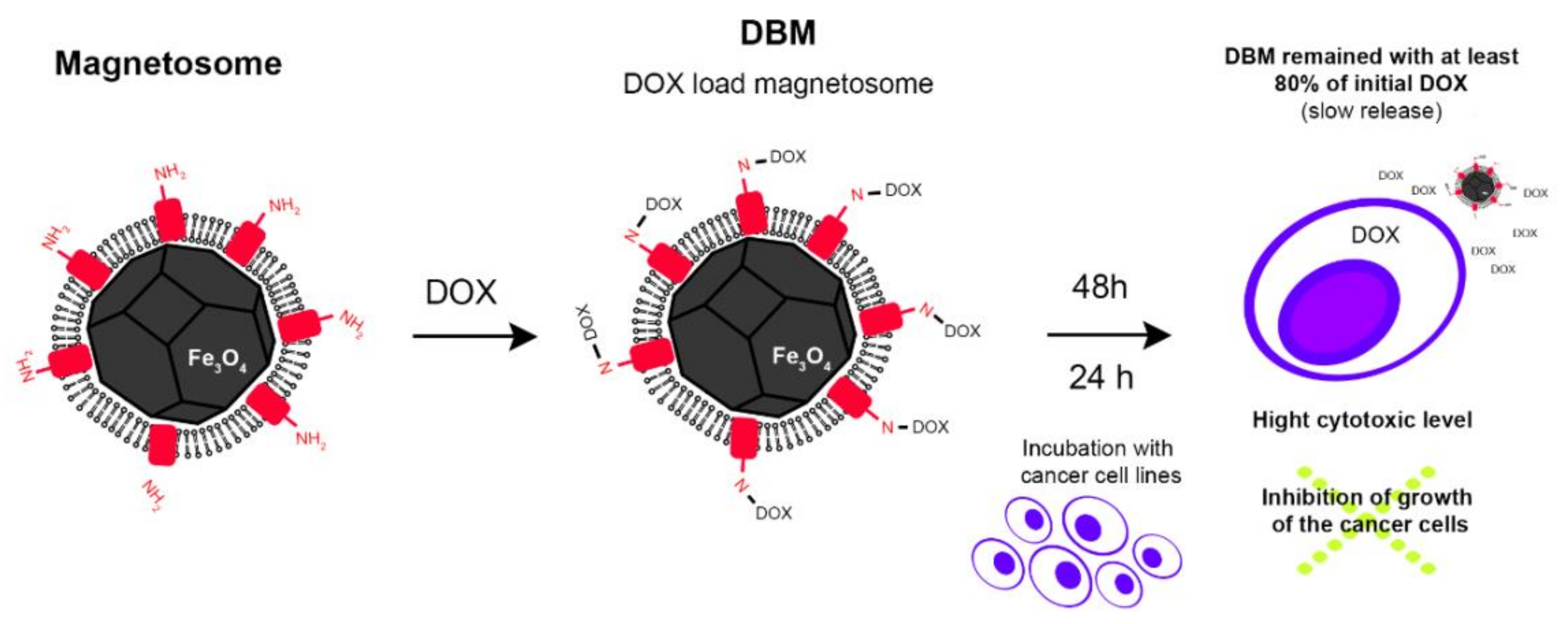
Magnetosomes in Drug Delivery
Magnetosomes in Cell Separation
Magnetosomes in Food Safety
Magnetosomes in DNA and Antigen Recovery/Detection Assays
Magnetosomes as Magnetic Resonance Imaging (MRI) Contrast Agents
Magnetosomes in Hyperthermia
Magnetosomes in Enzyme Immobilization
3. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bazylinski, D.A.; Frankel, R.B. Magnetosome formation in prokaryotes. Nat. Rev. Microbiol. 2004, 2, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Frankel, R.B.; Bazylinski, D.A. How magnetotactic bacteria make magnetosomes queue up. Trends Microbiol. 2006, 14, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Bellini, S. Ulteriori Studi Sui “batteri Magnetosensibili”. Universita di Pavia, Pavia, Italy, 1963, apud Frankel, R.B. The discovery of magnetotactic/magnetosensitive bactéria. Chin. J. Oceanol. Limnol. 2009, 27, 1–2. [Google Scholar]
- Blakemore, R. Magnetotactic bacteria. Science 1975, 190, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Lower, B.H.; Bazylinski, D.A. The bacterial magnetosome: A unique prokaryotic organelle. J. Mol. Microbiol. Biotechnol. 2013, 23, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, C.T.; Bazylinski, D.A. Ecology, diversity, and evolution of magnetotactic bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 497–526. [Google Scholar] [CrossRef] [PubMed]
- Web of science research platform. Clarivate Analytics: Philadelphia, PA, USA. Available online: https://login.webofknowledge.com/ (accessed on 1 March 2018).
- Scopus. Elsevier′s scopus, the largest abstract and citation database of peer-reviewed literature. In Elsevier′s Scopus; Elsevier: New York, NY, USA, 2018; Available online: https://www.scopus.com/ (accessed on 1 March 2018).
- Yan, L.; Da, H.; Zhang, S.; López, V.M.; Wang, W. Bacterial magnetosome and its potential application. Microbiol. Res. 2017, 203, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.J.; Suthindhiran, K. Magnetotactic bacteria and magnetosomes - scope and challenges. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 68, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Kamiya, S. Use of magnetic particles isolated from magnetotactic bacteria for enzyme immobilization. Appl. Microbiol. Biotechnol. 1987, 26, 328–332. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Beveridge, T.J.; Blakemore, R.P. Characterization of the bacterial magnetosome membrane. J. Bacteriol. 1988, 170, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Faivre, D.; Schuler, D. Magnetotactic bacteria and magnetosomes. Chem. Rev. 2008, 108, 4875–4898. [Google Scholar] [CrossRef] [PubMed]
- Murat, D.; Byrne, M.; Komeili, A. Cell biology of prokaryotic organelles. Cold Spring Harb. Perspect. Biol. 2010, 2, a000422. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.; Silva, K.T.; Leão, P.; Guedes, I.A.; Keim, C.N.; Farina, M.; Lins, U. Cell adhesion, multicellular morphology, and magnetosome distribution in the multicellular magnetotactic prokaryote Candidatus Magnetoglobus multicellularis. Microsc. Microanal. 2013, 19, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Komeili, A. Molecular mechanisms of compartmentalization and biomineralization in magnetotactic bacteria. FEMS Microbiol. Rev. 2012, 36, 232–255. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Kale, A.A.; Banpurkar, A.G.; Kulkarni, G.R.; Ogale, S.B. On the change in bacterial size and magnetosome features for Magnetospirillum magnetotacticum (MS-1) under high concentrations of zinc and nickel. Biomaterials 2009, 30, 4211–4218. [Google Scholar] [CrossRef] [PubMed]
- Grünberg, K.; Müller, E.C.; Otto, A.; Reszka, R.; Linder, D.; Kube, M.; Reinhardt, R.; Schüler, D. Biochemical and proteomic analysis of the magnetosome membrane in Magnetospirillum gryphiswaldense. Appl. Environ. Microbiol. 2004, 70, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, Y.; Arakaki, A.; Staniland, S.S.; Tanaka, T.; Matsunaga, T. Controlled formation of magnetite crystal by partial oxidation of ferrous hydroxide in the presence of recombinant magnetotactic bacterial protein Mms6. Biomaterials 2007, 28, 5381–5389. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, A.; Webb, J.; Matsunaga, T. A novel protein tightly bound to bacterial magnetic particles in Magnetospirillum magneticum strain AMB-1. J. Biol. Chem. 2003, 278, 8745–8750. [Google Scholar] [CrossRef] [PubMed]
- Murat, D.; Quinlan, A.; Vali, H.; Komeili, A. Comprehensive genetic dissection of the magnetosome gene island reveals the step-wise assembly of a prokaryotic organelle. Proc. Natl. Acad. Sci. USA 2010, 107, 5593–5598. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.; Murat, D.; Vali, H.; Komeili, A. The htra/degp family protease mame is a bifunctional protein with roles in magnetosome protein localization and magnetite biomineralization. Mol. Microbiol. 2011, 80, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Scheffel, A.; Gärdes, A.; Grünberg, K.; Wanner, G.; Schüler, D. The major magnetosome proteins mamgfdc are not essential for magnetite biomineralization in Magnetospirillum gryphiswaldense but regulate the size of magnetosome crystals. J. Bacteriol. 2008, 190, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Okamura, Y.; Arakaki, A.; Tanaka, T.; Takeyama, H.; Matsunaga, T. Origin of magnetosome membrane: Proteomic analysis of magnetosome membrane and comparison with cytoplasmic membrane. Proteomics 2006, 6, 5234–5247. [Google Scholar] [CrossRef] [PubMed]
- Faivre, D.; Godec, U.K. From bacteria to mollusks: The principles underlying the biomineralization of iron oxide materials. Angew. Chem. Int. Ed. 2015, 54, 4728–4747. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Lefèvre, C.T.; Schüler, D. Magnetotactic Bacteria. In The Prokaryotes; DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Baumgartner, J.; Morin, G.; Menguy, N.; Gonzalez, T.P.; Widdrat, M.; Cosmidis, J.; Faivre, D. Magnetotactic bacteria form magnetite from a phosphate-rich ferric hydroxide via nanometric ferric (oxyhydr) oxide intermediates. Proc. Natl. Acad. Sci. USA 2013, 110, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Pósfai, M.; Buseck, P.R.; Bazylinski, D.A.; Frankel, R.B. Iron sulfides from magnetotactic bacteria: Structure, composition, and phase transitions. Am. Mineral. 1998, 83, 1469–1481. [Google Scholar] [CrossRef]
- Li, Z.; Kawashita, M.; Araki, N.; Mitsumori, M.; Hiraoka, M.; Doi, M. Magnetite nanoparticles with high heating efficiencies for application in the hyperthermia of cancer. Mater. Sci Eng. 2010, 30, 990–996. [Google Scholar] [CrossRef]
- Sun, S.; Zheng, H. Size-Controlled Synthesis of Magnetite Nanoparticles. J. Am. Chem. Soc. 2002, 124, 8204–8205. [Google Scholar] [CrossRef] [PubMed]
- Schwab, S.T.; Stewart, C.A.; Dudeck, K.W.; Kozmina, S.M.; Katz, J.D.; Bartram, B.; Wuchina, E.J.; Kroenke, W.J.; Courtin, G. Polymeric precursors to refractory metal borides. J. Mat. Sci. 2004, 39, 6051–6055. [Google Scholar] [CrossRef]
- Qi, H.; Yan, B.; Lu, W.; Li, C.; Yang, Y. A non-alkoxide sol-gel method for the preparation of magnetite (Fe3O4) nanoparticles. Curr. Nanosci. 2011, 7, 381. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Z.; Qu, J.; Wang, Z.; Qiu, J. Shape-control and characterization of magnetite prepared via a one-step solvothermal route. Cryst. Growth Des. 2010, 10, 2863. [Google Scholar] [CrossRef]
- Peigneux, A.; Valverde-Tercedor, C.; Lopez-Moreno, R.; Perez-Gonzalez, T.; Fernandez-Vivas, M.A.; Jimenez-Lopez, C. Learning from magnetotactic bacteria: A review on the synthesis of biomimetic nanoparticles mediated by magnetosome-associated proteins. J. Struct. Biol. 2016, 196, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E. Applications of magnetosomes synthesized by magnetotactic bacteria in medicine. Front. Bioen. Biotechnol. 2014, 2, 5. [Google Scholar]
- Bazylinski, D.A.; Frankel, R.B.; Heywood, B.R.; Mann, S.; King, J.W.; Donaghay, P.L.; Hanson, A.K. Controlled biomineralization of magnetite Fe3O4 and greigite Fe3O4 in a magnetotactic bacterium. Appl. Environ. Microbiol. 1995, 61, 3232–3239. [Google Scholar] [PubMed]
- Bazylinski, D.A.; Garratt-Reed, A.J.; Frankel, R.B. Electron microscopic studies of magnetosomes in magnetotactic bacteria. Microsc. Res. Tech. 1994, 27, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, D.L.; Maratea, D.; Blakemore, R.P. Ultra-structure of a magnetotactic spirillum. J. Bacteriol. 1980, 141, 1399–1408. [Google Scholar] [PubMed]
- Mann, S.; Frankel, R.B.; Blakemore, R.P. Structure, morphology and crystal growth of bacterial magnetite. Nature 1984, 310, 405–407. [Google Scholar] [CrossRef]
- Mann, S.; Moench, T.T.; Williams, R.J.P. A high resolution electron microscopic investigation of bacterial magnetite. Implications for crystal growth. Proc. R. Soc. Lond. B. 1984, 310, 385–393. [Google Scholar] [CrossRef]
- Kiseleva, L.; Briliute, J.; Khilyas, I.V.; Simpson, D.J.; Fedorovich, V.; Cohen, M.; Goryanin, I. Magnet-facilitated selection of electrogenic bacteria from marine sediment. Biomed. Res. Int. 2015, 2015, 582471. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, C.T.; Frankel, R.B.; Abreu, F.; Lins, U.; Bazylinski, D.A. Culture-independent characterization of a novel, uncultivated magnetotactic member of the nitrospirae phylum. Environ. Microbiol. 2011, 13, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Menguy, N.; Gatel, C.; Boureau, V.; Snoeck, E.; Patriarche, G.; Leroy, E.; Pan, Y. Crystal growth of bullet-shaped magnetite in magnetotactic bacteria of the nitrospirae phylum. J. R Soc. Interface 2015, 12, 20141288. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.; Sparks, N.H.; Walker, M.M.; Kirschvink, J.L. Ultrastructure, morphology and organization of biogenic magnetite from sockeye salmon, oncorhynchus nerka: Implications for magnetoreception. J. Exp. Biol. 1988, 140, 35–49. [Google Scholar] [PubMed]
- Bazylinski, D.A.; Frankel, R.B. Biologically controlled mineralization of magnetic iron minerals by MTB. In Environmental Microbe-Mineral Interactions; Lovely, D.R., Ed.; ASM Press: Washington, DC, USA, 2000. [Google Scholar]
- Bazylinski, D.A.; Frankel, R.B. Magnetic iron oxide and iron sulfide minerals within organisms. In Biomineralization: From Biology to Biotechnology and Medical Application; Baeuerlein, E., Ed.; Wiley-VCH: Weinheim, Germany, 2000; pp. 25–46. [Google Scholar]
- Lins, U.; McCartney, M.R.; Farina, M.; Frankel, R.B.; Buseck, P.R. Habits of magnetosome crystals in coccoid magnetotactic bacteria. Appl. Environ. Microbiol. 2005, 71, 4902–4905. [Google Scholar] [CrossRef] [PubMed]
- Spring, S.; Lins, U.; Amann, R.; Schleifer, K.H.; Ferreira, L.C.; Esquivel, D.M.; Farina, M. Phylogenetic affiliation and ultrastructure of uncultured magnetic bacteria with unusually large magnetosomes. Arch. Microbiol. 1998, 169, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Janke, V.; Sievers, S.; Siegner, U.; Schueler, D.; Heyen, U. Scanning force microspy study of biogenic nanoparticles for medical applications. J. Magn. Magn. Mater. 2005, 290–291, 269–271. [Google Scholar] [CrossRef]
- Butler, R.F.; Banerjee, S.K. Theoretical single-domain grain size range in magnetite and titanomagnetite. J. Geophysics 1975, 80, 4049–4053. [Google Scholar] [CrossRef]
- Frankel, R.B.; Blakemore, R.P. Navigational compass in magnetic bacteria. J. Magn. Magn. Mater 1980, 1562, 15–18. [Google Scholar] [CrossRef]
- Moskowitz, B.M. Biomineralization of magnetic minerals. Rev. Geophys. 1995, 33, 123–128. [Google Scholar] [CrossRef]
- Feng, M.; Lu, Y.; Yang, Y.; Zhang, M.; Xu, Y.J.; Gao, H.L.; Dong, L.; Xu, W.P.; Yu, S.H. Bioinspired Greigite Magnetic Nanocrystals: Chemical Synthesis and Biomedicine Applications. Sci. Rep. 2013, 3, 2994. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Chen, K.; Chen, X. Production, modification and bio-applications of magnetic nanoparticles gestated by magnetotactic bacteria. Nano Res. 2009, 2, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhang, S.; Chen, P.; Liu, H.; Yin, H.; Li, H. Magnetotactic bacteria, magnetosomes and their application. Microbiol. Res. 2012, 167, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Heyen, U.; Schüler, D. Growth and magnetosome formation by microaerophilic Magnetospirillum strains in an oxygen-controlled fermentor. Appl. Microbiol. Biotechnol. 2003, 61, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-K.; Zhang, F.-F.; Sun, J.-J.; Sheng, J.; Wang, F.; Sun, M. Bio and nanomaterials based on Fe3O4. Molecules 2014, 19, 21506–21528. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, G.R.; Guo, F.F.; Jiang, W.; Li, Y.; Li, L.J. Large-scale production of magnetosomes by chemostat culture of Magnetospirillum gryphiswaldense at high cell density. Microb. Cell. Fact. 2010, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Jiang, W.; Li, Y.; Li, J. Semicontinuous culture of Magnetospirillum gryphiswaldense MSR-1 cells in an autofermentor by nutrient-balanced and isosmotic feeding strategies. Appl. Environ. Microbiol. 2011, 77, 5851–5856. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.B.; Zhao, F.; Tang, T.; Jiang, W.; Tian, J.S.; Li, Y.; Li, J.L. High-yield growth and magnetosome formation by Magnetospirillum gryphiswaldense MSR-1 in an oxygen-controlled fermentor supplied solely with air. Appl. Microbiol. Biotechnol. 2008, 79, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.C.; Abreu, F.; Silva, K.T.; Bazylinski, D.A.; Lins, U. Magnetotactic bacteria as potential sources of bioproducts. Mar. Drugs 2015, 13, 389–430. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Wei, J.; Jianbo, S.; Guili, W.; Feng, G.; Ying, L. Purified and sterilized magnetosomes from Magnetospirillum gryphiswaldense MSR-1 were not toxic to mouse fibroblasts in vitro. Lett. Appl. Microbiol. 2007, 45, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.B.; Duan, J.H.; Dai, S.L.; Ren, J.; Guo, L.; Jiang, W.; Li, Y. Preparation and anti-tumor efficiency evaluation of doxorubicin-loaded bacterial magnetosomes: Magnetic nanoparticles as drug carriers isolated from Magnetospirillum gryphiswaldense. Biotechnol. Bioeng. 2008, 101, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Amor, M.; Busigny, V.; Durand-Dubief, M.; Tharaud, M.; Ona-Nguema, G.; Gelabert, A.; Alphandery, E.; Menguy, N.; Benedetti, M.F.; Chebbi, I.; et al. Chemical signature of magnetotactic bacteria. Proc. Natl. Acad. Sci. USA 2015, 112, 1699–1703. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Y.; Liang, X.J.; Wang, P.C. Bacterial magnetosome: A novel biogenetic magnetic targeted drug carrier with potential multifunctions. J. Nanomater 2011, 2011, 469031–469043. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Matsunaga, T. Development of efficient expression system for protein display on bacterial magnetic particles. Biochem. Biophys. Res. Commun. 2005, 338, 1678–1681. [Google Scholar] [CrossRef] [PubMed]
- Kuhara, M.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Magnetic cell separation using antibody binding with protein a expressed on bacterial magnetic particles. Anal. Chem. 2004, 76, 6207–6213. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Matsunaga, T. Fully automated chemiluminescence immunoassay of insulin using antibody-Protein A–Bacterial magnetic particle complexes. Anal. Chem. 2000, 72, 3518–3522. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, D.; Schüler, D. Development of a genetic system for Magnetospirillum gryphiswaldense. Arch. Microbiol. 2003, 179, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Mickoleit, F.; Borkner, C.B.; Toro-Nahuelpan, M.; Herold, H.M.; Maier, D.S.; Plitzko, J.M.; Scheibel, T.; Schuler, D. In vivo coating of bacterial magnetic nanoparticles by magnetosome expression of spider silk-inspired peptides. Biomacromolecules 2018, 19, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Ceyhan, B.; Alhorn, P.; Lang, C.; Schüler, D.; Niemeyer, C.M. Semisynthetic biogenic magnetosome nanoparticles for the detection of proteins and nucleic acids. Small 2006, 2, 1251–1255. [Google Scholar] [CrossRef] [PubMed]
- Wacker, R.; Ceyhan, B.; Alhorn, P.; Schueler, D.; Lang, C.; Niemeyer, C.M. Magneto immuno-PCR: A novel immunoassay based on biogenic magnetosome nanoparticles. Biochem. Biophys. Res. Commun. 2007, 357, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.G.; Dai, Q.L.; Wang, S.B.; Deng, Q.J.; Wu, W.G.; Chen, A.Z. Preparation and in vitro antitumor effects of cytosine arabinoside-loaded genipin-poly-l-glutamic acid-modified bacterial magnetosomes. Int. J. Nanomed. 2015, 10, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Ke, Y.; Yu, S.; Jing, J. Co-delivery of doxorubicin and recombinant plasmid phsp70-plk1-shrna by bacterial magnetosomes for osteosarcoma therapy. Int. J. Nanomed. 2016, 11, 5277–5286. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Li, X.; Guo, J.; Yang, G.; Li, X. Ganglioside-magnetosome complex formation enhances uptake of gangliosides by cells. Int. J. Nanomed. 2015, 10, 6919–6930. [Google Scholar]
- Li, A.; Zhang, H.; Zhang, X.; Wang, Q.; Tian, J.; Li, Y.; Li, J. Rapid separation and immunoassay for low levels of Salmonella in foods using magnetosome-antibody complex and real-time fluorescence quantitative pcr. J. Sep. Sci. 2010, 33, 3437–3443. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Gao, B.; Zhang, F.; Sun, X.; Zhang, Y.; Li, Z. A novel electrochemical immunosensor based on magnetosomes for detection of Staphylococcal enterotoxin B in milk. Talanta 2013, 106, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Felfoul, O.; Mohammadi, M.; Taherkhani, S.; De Lanauze, D.; Xu, Y.Z.; Loghin, D.; Essa, S.; Jancik, S.; Houle, D.; Lafleur, M.; et al. Magneto-aerotactic bacteria deliver drug-containing nanoliposomes to tumour hypoxic regions. Nat. Nanotechnol. 2016, 11, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Shimoshige, H.; Nakajima, Y.; Kobayashi, H.; Yanagisawa, K.; Nagaoka, Y.; Shimamura, S.; Mizuki, T.; Inoue, A.; Maekawa, T. Formation of core-shell nanoparticles composed of magnetite and samarium oxide in Magnetospirillum magneticum strain RSS-1. PLoS ONE 2017, 12, e0170932. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lisowski, W.; Zhou, Y.; Jern, N.W.; Huang, K.; Fong, E. Genetic improvement of Magnetospirillum gryphiswaldense for enhanced biological removal of phosphate. Biotechnol. Lett. 2017, 39, 1509–1514. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Brown, R.; Hondow, N.; Arakaki, A.; Matsunaga, T.; Staniland, S. Highest levels of Cu, Mn and Co doped into nanomagnetic magnetosomes through optimized biomineralisation. J. Mater. Chem. 2012, 22, 11919–11921. [Google Scholar] [CrossRef]
- Prozorov, T.; Palo, P.; Wang, L.; Nilsen-Hamilton, M.; Jones, D.; Orr, D.; Mallapragada, S.K.; Narasimhan, B.; Canfield, P.C.; Prozorov, R. Cobalt ferrite nanocrystals: Out-performing magnetotactic bacteria. ACS Nano 2007, 1, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Staniland, S.; Williams, W.; Telling, N.; Van Der Laan, G.; Harrison, A.; Ward, B. Controlled cobalt doping of magnetosomes in vivo. Nat. Nanotechnol. 2008, 3, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Arakaki, A.; Staniland, S.S.; Matsunaga, T. Simultaneously discrete biomineralization of magnetite and tellurium nanocrystals in magnetotactic bacteria. Appl. Environ. Microbiol. 2010, 76, 5526–5532. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Knowles, W.; Brown, R.; Hondow, N.; Arakaki, A.; Baldwin, S.; Staniland, S.; Matsunaga, T. Biomagnetic recovery and bioaccumulation of selenium granules in magnetotactic bacteria. Appl. Environ. Microbiol. 2016, 82, 3886–3891. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Peng, C.; Khan, Z.M.; Naz, I.; Sultan, M. An overview of heavy metal removal from wastewater using magnetotactic bacteria. J. Chem. Technol. Biotechnol. 2018, 93, 2817–2832. [Google Scholar] [CrossRef]
- Arakaki, A.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Cadmium recovery by a sulfate-reducing magnetotactic bacterium, Desulfovibrio magneticus RS-1, using magnetic separation. Appl. Biochem. Biotechnol. 2002, 98–100, 833–840. [Google Scholar] [CrossRef]
- Smit, B.A.; Van Zyl, E.; Joubert, J.J.; Meyer, W.; Preveral, S.; Lefevre, C.T.; Venter, S.N. Magnetotactic bacteria used to generate electricity based on faraday′s law of electromagnetic induction. Lett. Appl. Microbiol. 2018, 66, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Long, R.; Liu, Y.; Dai, Q.; Wang, S.; Deng, Q.; Zhou, X. A natural bacterium-produced membrane-bound nanocarrier for drug combination therapy. Materials 2016, 9, 889. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, J.G.; Geng, Y.Y.; Wang, J.J.; Zhang, X.M.; Yang, S.S.; Jiang, W.; Liu, W.Q. An enhanced anti-tumor effect of apoptin-cecropin b on human hepatoma cells by using bacterial magnetic particle gene delivery system. Biochem. Biophys. Res. Commun. 2018, 496, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Hirabe, H.; Takahashi, M.; Kuhara, M.; Takeyama, H.; Matsunaga, T. Magnetic cell separation using nano-sized bacterial magnetic particles with reconstructed magnetosome membrane. Biotechnol. Bioeng. 2008, 101, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yoshino, T.; Matsunaga, T. Surface modification of magnetic nanoparticles using asparagines-serine polypeptide designed to control interactions with cell surfaces. Biomaterials 2010, 31, 4952–4957. [Google Scholar] [CrossRef] [PubMed]
- Grouzdev, D.S.; Dziuba, M.V.; Kurek, D.V.; Ovchinnikov, A.I.; Zhigalova, N.A.; Kuznetsov, B.B.; Skryabin, K.G. Optimized method for preparation of igg-binding bacterial magnetic nanoparticles. PLoS ONE 2014, 9, e109914. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Hu, J.; Liu, L.; Li, L.; Wang, X.; Zhang, H.; Jiang, W.; Tian, J.; Li, Y.; Li, J. Surface expression of protein a on magnetosomes and capture of pathogenic bacteria by magnetosome/antibody complexes. Front. Microbiol. 2014, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Kim, H.; Choi, S.H.; Park, M.; Kim, D.; Kim, H.C.; Choi, Y.; Lin, S.; Kim, B.H.; Jung, H.S.; et al. Magnetosome-like ferrimagnetic iron oxide nanocubes for highly sensitive MRI of single cells and transplanted pancreatic islets. Proc. Natl. Acad. Sci. USA 2011, 108, 2662–2667. [Google Scholar] [CrossRef] [PubMed]
- Orlando, T.; Mannucci, S.; Fantechi, E.; Conti, G.; Tambalo, S.; Busato, A.; Innocenti, C.; Ghin, L.; Bassi, R.; Arosio, P.; et al. Characterization of magnetic nanoparticles from Magnetospirillum gryphiswaldense as potential theranostics tools. Contrast Media Mol Imaging 2016, 11, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Boucher, M.; Geffroy, F.; Prévéral, S.; Bellanger, L.; Selingue, E.; Adryanczyk-Perrier, G.; Péan, M.; Lefèvre, C.T.; Pignol, D.; Ginet, N.; Mériaux, S. Genetically tailored magnetosomes used as MRI probe for molecular imaging of brain tumor. Biomaterials 2017, 121, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Goldhawk, D.E.; Neil Gelman, T.R.T.; Prato, F.S. Forming magnetosome-like nanoparticles in mammalian cells for molecular MRI. In Design and Applications of Nanoparticles in Biomedical Imaging; Springer International Publishing: Basel, Switzerland, 2017; pp. 187–203. [Google Scholar]
- Xiang, Z.; Yang, X.; Xu, J.; Lai, W.; Wang, Z.; Hu, Z.; Tian, J.; Geng, L.; Fang, Q. Tumor detection using magnetosome nanoparticles functionalized with a newly screened egfr/her2 targeting peptide. Biomaterials 2017, 115, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Idbaih, A.; Adam, C.; Delattre, J.Y.; Schmitt, C.; Guyot, F.; Chebbi, I. Development of non-pyrogenic magnetosome minerals coated with poly-l-lysine leading to full disappearance of intracranial U87-luc glioblastoma in 100% of treated mice using magnetic hyperthermia. Biomaterials 2017, 141, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Faure, S.; Seksek, O.; Guyot, F.; Chebbi, I. Chains of magnetosomes extracted from AMB-1 magnetotactic bacteria for application in alternative magnetic field cancer therapy. ACS Nano 2011, 5, 6279–6296. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, S.; Tambalo, S.; Conti, G.; Ghin, L.; Milanese, A.; Carboncino, A.; Nicolato, E.; Marinozzi, M.R.; Benati, D.; Bassi, R.; et al. Magnetosomes extracted from Magnetospirillum gryphiswaldense as theranostic agents in an experimental model of glioblastoma. Contrast Media Mol. Imaging 2018, 2018, 2198703. [Google Scholar] [CrossRef] [PubMed]
- Ginet, N.; Pardoux, R.; Adryanczyk, G.; Garcia, D.; Brutesco, C.; Pignol, D. Single-step production of a recyclable nanobiocatalyst for organophosphate pesticides biodegradation using functionalized bacterial magnetosomes. PLoS ONE 2011, 6, e21442. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Tanaka, T.; Yoshino, T. Stoichiometrically controlled immobilization of multiple enzymes on magnetic nanoparticles by the magnetosome display system for efficient cellulose hydrolysis. Biomacromolecules 2015, 16, 3863–3868. [Google Scholar] [CrossRef] [PubMed]
- Martel, S. Targeting active cancer cells with smart bullets. Ther. Deliv. 2017, 8, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Hoshino, K.; Kawai, T.; Sanjo, H.; Takada, H.; Ogawa, T.; Takeda, K.; Akira, S. Differential roles of tlr2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components. Immunity 1999, 11, 443–451. [Google Scholar] [CrossRef]
- Lüderitz, O.T.T.O.; Westphal, O.T.T.O.; Staub, A.M.; Nikaido, H.I.R.O.S.H.I. Isolation and chemical and immunological characterization of bacterial lipopolysaccharides. In Microbial Toxins, 1st ed.; Weinbaum, G., Kadis, S., Ajl, S.J., Eds.; Academic Press: New York, NY, USA, 1971; Volume 4, pp. 145–233. [Google Scholar]
- Wang, X.; Quinn, P.J. Endotoxins: Lipopolysaccharides of gram-negative bacteria. Subcell. Biochem. 2010, 53, 3–25. [Google Scholar] [PubMed]
- Tanaka, M.; Nakata, Y.; Mori, T.; Okamura, Y.; Miyasaka, H.; Takeyama, H.; Matsunaga, T. Development of a cell surface display system in a magnetotactic bacterium, “Magnetospirillum magneticum” AMB-1. Appl. Environ. Microbiol. 2008, 74, 3342–3348. [Google Scholar] [CrossRef] [PubMed]
- Blondeau, M.; Guyodo, Y.; Guyot, F.; Gatel, C.; Menguy, N.; Chebbi, I.; Haye, B.; Durand-Dubief, M.; Alphandery, E.; Brayner, R.; et al. Magnetic-field induced rotation of magnetosome chains in silicified magnetotactic bacteria. Sci. Rep. 2018, 8, 7699. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.J.; Mumper, E.; Brown, E.E.; Brangham, J.T.; Lower, B.H.; Lower, S.K.; Yang, F.Y.; Sooryakumar, R. Tuning bacterial hydrodynamics with magnetic fields. Phys. Rev. E 2017, 95, 062612. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.B.; Duan, J.H.; Dai, S.L.; Ren, J.; Zhang, Y.D.; Tian, J.S.; Li, Y. In vitro and in vivo antitumor effects of doxorubicin loaded with bacterial magnetosomes (DBMs) on H22 cells: The magnetic bio-nanoparticles as drug carriers. Cancer Lett. 2007, 258, 109–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, G.; Cong, Y.; Tong, G.; Lin, Z.; Yin, Y.; Zhang, C. The preparation and characterization of micelles from poly (γ-glutamic acid)-graft-poly(l-lactide) and the cellular uptake thereof. J. Mater. Sci. Mater. Med. 2015, 26, 187. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Kamiya, S. Indroduction of bacterial magnetic particles into red blood cells with cell fusion. In Proceedings of the 6th International Conference on Biomagnetisms, Toyko, Japan, 27–30 August 1987; Tokyo Denki University Press: Toyko, Japan, 1988; pp. 50–51. [Google Scholar]
- Matsunaga, T.; Hashimoto, K.; Nakamura, N.; Nakamura, K.; Hashimoto, S. Phagocytosis of bacterial magnetite by leucocytes. Appl. Microbiol. Biotechnol. 1989, 31, 401–405. [Google Scholar] [CrossRef]
- Herborn, C.U.; Papanikolaou, N.; Reszka, R.; Grunberg, K.; Schuler, D.; Debatin, J.F. Magnetosomes as biological model for iron binding: Relaxivity determination with MRI. RoFo 2003, 175, 830–834. [Google Scholar] [PubMed]
- Mériaux, S.; Boucher, M.; Marty, B.; Lalatonne, Y.; Prévéral, S.; Motte, L.; Lefèvre, C.T.; Geffroy, F.; Lethimonnier, F.; Péan, M.; et al. Magnetosomes, biogenic magnetic nanomaterials for brain molecular imaging with 17.2 t MRI scanner. Adv. Healthc. Mater. 2015, 4, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Kraupner, A.; Eberbeck, D.; Heinke, D.; Uebe, R.; Schuler, D.; Briel, A. Bacterial magnetosomes - nature’s powerful contribution to mpi tracer research. Nanoscale 2017, 9, 5788–5793. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, R.K.; Medal, R.; Shorey, W.D.; Hanselman, R.C.; Parrott, J.C.; Taylor, C.B. Selective inductive heating of lymph nodes. Ann. Surg. 1957, 146, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Moroz, P.; Jones, S.K.; Gray, B.N. Magnetically mediated hyperthermia: Current status and future directions. Int. J. Hyperthermia 2002, 18, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Idbaih, A.; Adam, C.; Delattre, J.Y.; Schmitt, C.; Guyot, F.; Chebbi, I. Chains of magnetosomes with controlled endotoxin release and partial tumor occupation induce full destruction of intracranial U87-luc glioma in mice under the application of an alternating magnetic field. J. Control. Release 2017, 262, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ahn, J.H.; Choi, H.; Seo, J.M.; Cho, D.; Koo, K. Natural magnetic nanoparticle containing droplet for smart drug delivery and heat treatment. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2015, 2015, 3541–3544. [Google Scholar] [PubMed]
- Mathuriya, A.S. Magnetotactic bacteria: Nanodrivers of the future. Crit. Rev. Biotechnol. 2016, 36, 788–802. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Chebbi, I.; Guyot, F.; Durand-Dubief, M. Use of bacterial magnetosomes in the magnetic hyperthermia treatment of tumours: A review. Int. J. Hyperthermia 2013, 29, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Guyot, F.; Chebbi, I. Preparation of chains of magnetosomes, isolated from Magnetospirillum magneticum strain AMB-1 magnetotactic bacteria, yielding efficient treatment of tumors using magnetic hyperthermia. Int. J. Pharm. 2012, 434, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Shimada, T.; Ito, Y.; Honda, T.; Maeda, Y.; Matsunaga, T.; Tanaka, T. Biosynthesis of thermoresponsive magnetic nanoparticles by magnetosome display system. Bioconjug. Chem. 2018, 29, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Hergt, R.; Hiergeist, R.; Zeisberger, M.; Schüler, D.; Heyen, U.; Hilger, I.; Kaiser, W.A. Magnetic properties of bacterial magnetosomes as potential diagnostic and therapeutic tools. J. Magn. Magn. Mater. 2005, 293, 80–86. [Google Scholar] [CrossRef]
- Alphandery, E.; Abi Haidar, D.; Seksek, O.; Guyot, F.; Chebbi, I. Fluorescent magnetosomes for controlled and repetitive drug release under the application of an alternating magnetic field under conditions of limited temperature increase (<2.5 °C). Nanoscale 2018, 10, 10918–10933. [Google Scholar] [CrossRef] [PubMed]
- Plan Sangnier, A.; Preveral, S.; Curcio, A.; Silva, A.K.A.; Lefevre, C.T.; Pignol, D.; Lalatonne, Y.; Wilhelm, C. Targeted thermal therapy with genetically engineered magnetite magnetosomes@RGD: Photothermia is far more efficient than magnetic hyperthermia. J. Control. Release 2018, 279, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.A.; Park, H.J.; Driscoll, A.J. Enzyme nanoparticle fabrication: Magnetic nanoparticle synthesis and enzyme immobilization. Methods Mol. Biol. 2011, 679, 183–191. [Google Scholar] [PubMed]
- Kolinko, I.; Lohße, A.; Borg, S.; Raschdorf, O.; Jogler, C.; Tu, Q.; Pósfai, M.; Tompa, E.; Plitzko, J.M.; Brachmann, A.; et al. Biosynthesis of magnetic nanostructures in a foreign organism by transfer of bacterial magnetosome gene clusters. Nat. Nanotechnol. 2014, 9, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Zhang, J.; An, X.; Zhang, B. In situ synthesis of fluorescent magnetosomes using an organic membrane as a soft template. Nanoscale 2017, 9, 5408–5412. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Applications of whole MTB | ||||||
| Field | Application | Ref. | Advantages | Disadvantages | ||
| Drug delivery | Drug-loaded nanoliposomes attached to Mc. marinus cells for targeted tumor treatment | [78] | Dispenses cell lysis; Uses cell’s own magnetotaxis | Potentially immunogenic due to outer LPS | ||
| Bioremediation | Wastewater treatment; Removal of heavy metals (Cd, Te, Se) | [79,80,81,82,83,84,85,86,87] | Magnetic crystal doping possible; Recovery of removed minerals | Poor growth of MTB in contaminated media; Biomineralization may be affected | ||
| Energy generation | Electricity generation by cells and magnetosomes of Ms. magneticum AMB-1 by means of electromagnetic induction | [88] | Green energy technology | Only millivolts generated; Expensive | ||
| Applications of magnetosomes | ||||||
| Field | Application | Ref. | Functionalization Method | Advantages | Disadvantages | |
| Drug delivery | Delivery of antitumor drugs: doxorubicin, cytarabine, daunorubicin; delivery of gangliosides; Antitumor gene delivery | [63,74,75,89,90] | Chemical crosslinking with glutaraldehyde and genipin/PLGA; Surface adsorption of plasmids | Targeted drug delivery; Reduction of drug toxicity; Tissue specificity; Easy functionalization | Possible activity alteration; Unclear biological fate; Endotoxin test needed | |
| Cell separation | Sorting of blood cells; | [91,92,93] | Binding protein expression by vector cloning; Insertion of modified binding protein into membrane | Reutilization of capture complex; High specificity separation | Difficult steps of cloning and expression; Alteration of cell viability after capture | |
| Food safety | Capture of Salmonella and Vibrio cells; Enterotoxin detection | [76,77,94] | Crosslinking of antibodies | Reutilization of capture complex; High sensitivity | Antibody specificity | |
| MRI contrast agent | Diagnostic detection of tumors | [95,96,97,98,99] | No functionalization; Chemical coupling of targeting peptide | May also be used as therapeutic tool (by hyperthermia, drug delivery); High affinity to target cells; High detection sensitivity | Unclear biological fate; Endotoxin test needed | |
| DNA/Antigen Recovery/Detection | Capture of oligonucleotides and antibodies; Hepatitis B antigen detection | [71,72] | Biotinylation by chemical crosslinking with NHS | High sensitivity and recovery efficiency | Complex technology | |
| Hyperthermia | Treatment of tumors | [100,101,102] | No functionalization, generally | Less significant side-effects than chemotherapy and radiotherapy; Tissue specificity; May also be used as diagnostic tool | Unclear biological fate; Endotoxin test needed | |
| Enzyme immobilization | Bioremediation of organophosphate pesticides; Cellulose degradation | [103,104] | Enzyme expression by vector cloning | Reutilization of nanobiocatalyst; Immobilization of multiple catalysts | Difficult steps of cloning and expression; Possible loss of activity due to immobilization | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas, G.; Cypriano, J.; Correa, T.; Leão, P.; Bazylinski, D.A.; Abreu, F. Applications of Magnetotactic Bacteria, Magnetosomes and Magnetosome Crystals in Biotechnology and Nanotechnology: Mini-Review. Molecules 2018, 23, 2438. https://doi.org/10.3390/molecules23102438
Vargas G, Cypriano J, Correa T, Leão P, Bazylinski DA, Abreu F. Applications of Magnetotactic Bacteria, Magnetosomes and Magnetosome Crystals in Biotechnology and Nanotechnology: Mini-Review. Molecules. 2018; 23(10):2438. https://doi.org/10.3390/molecules23102438
Chicago/Turabian StyleVargas, Gabriele, Jefferson Cypriano, Tarcisio Correa, Pedro Leão, Dennis A. Bazylinski, and Fernanda Abreu. 2018. "Applications of Magnetotactic Bacteria, Magnetosomes and Magnetosome Crystals in Biotechnology and Nanotechnology: Mini-Review" Molecules 23, no. 10: 2438. https://doi.org/10.3390/molecules23102438
APA StyleVargas, G., Cypriano, J., Correa, T., Leão, P., Bazylinski, D. A., & Abreu, F. (2018). Applications of Magnetotactic Bacteria, Magnetosomes and Magnetosome Crystals in Biotechnology and Nanotechnology: Mini-Review. Molecules, 23(10), 2438. https://doi.org/10.3390/molecules23102438