
Oligonucleotides and ND-FISH Displaying Different Arrangements of Tandem Repeats and Identification of Dasypyrum villosum Chromosomes in Wheat Backgrounds

Abstract
:1. Introduction
2. Results
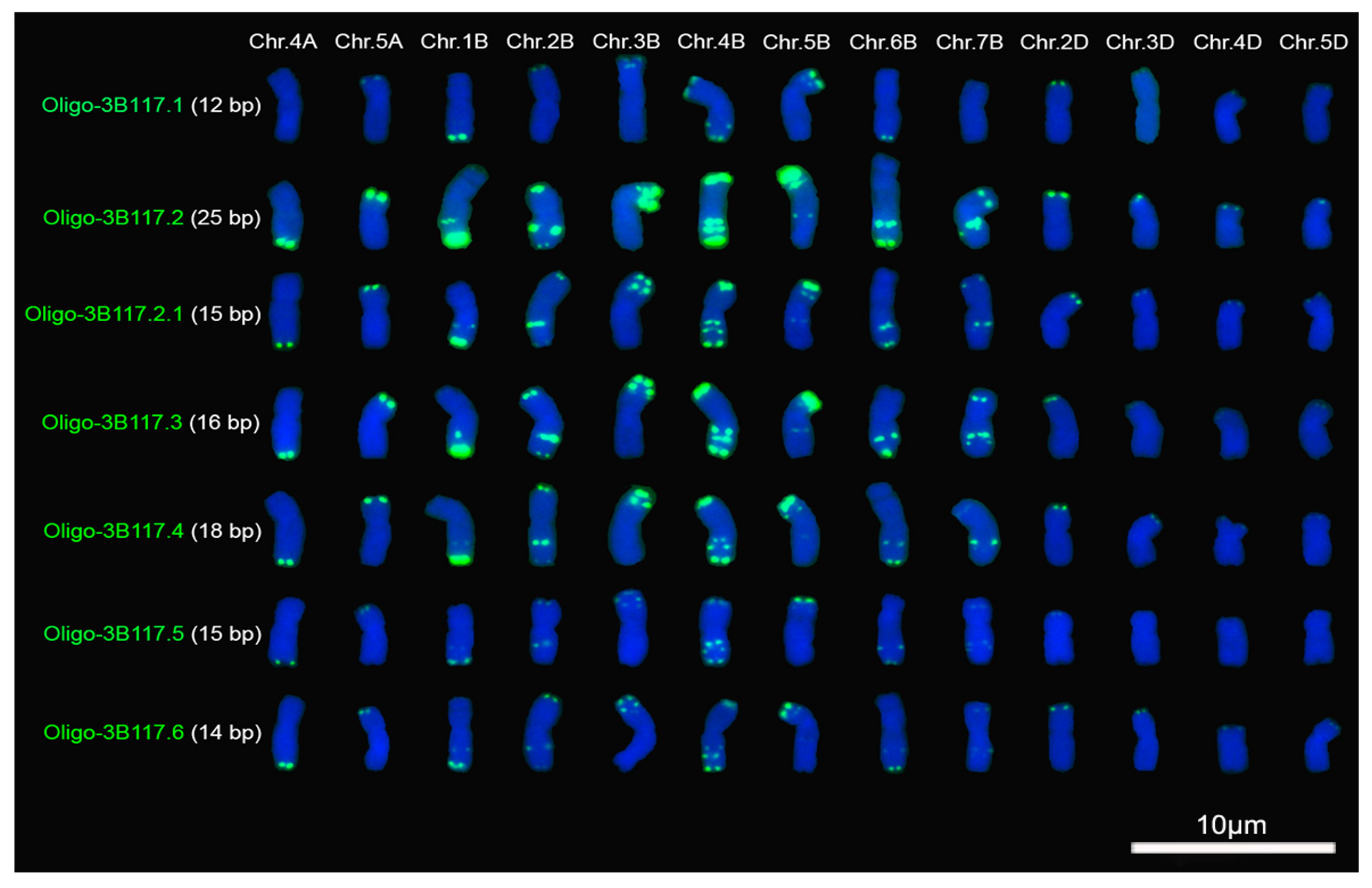
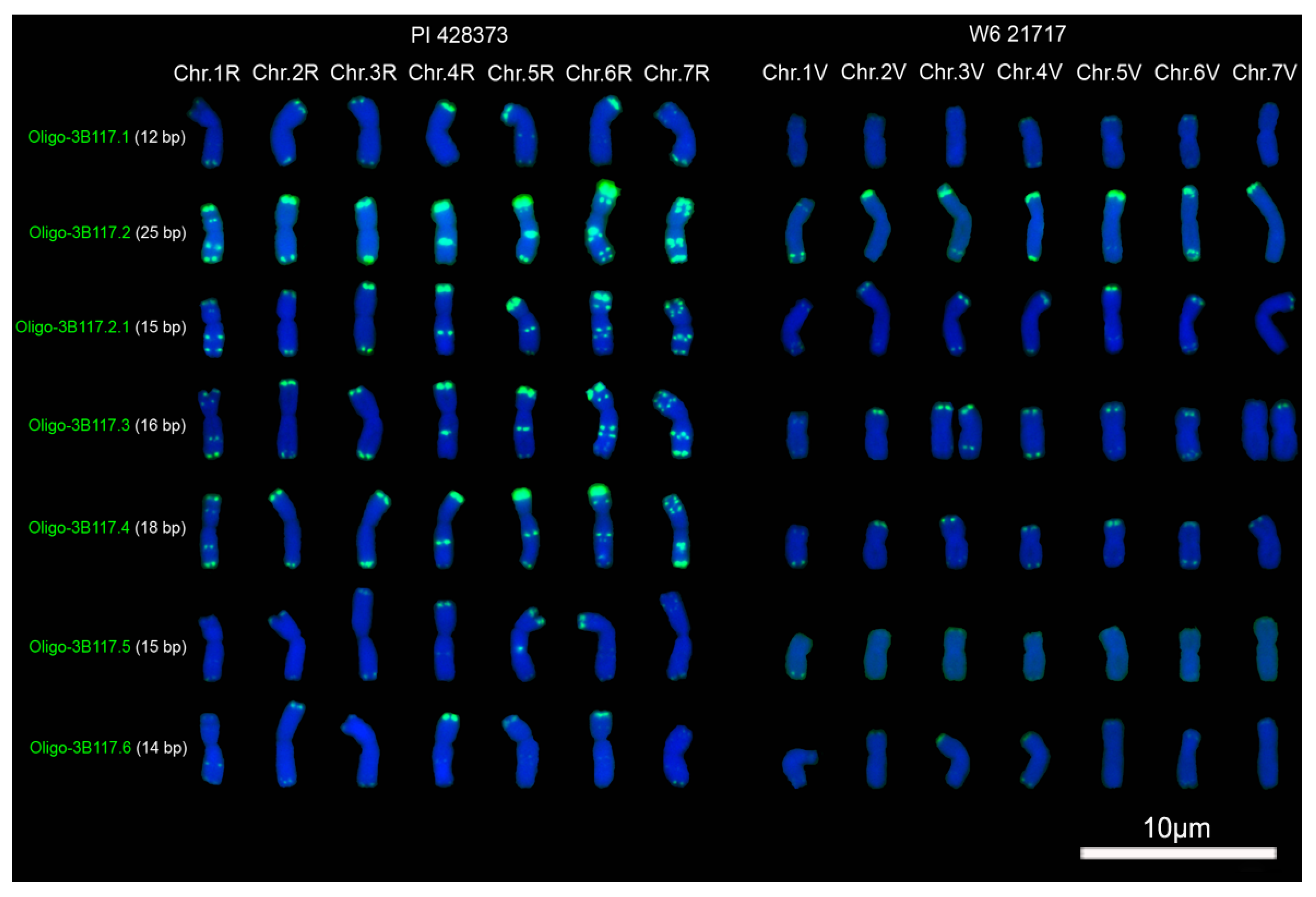
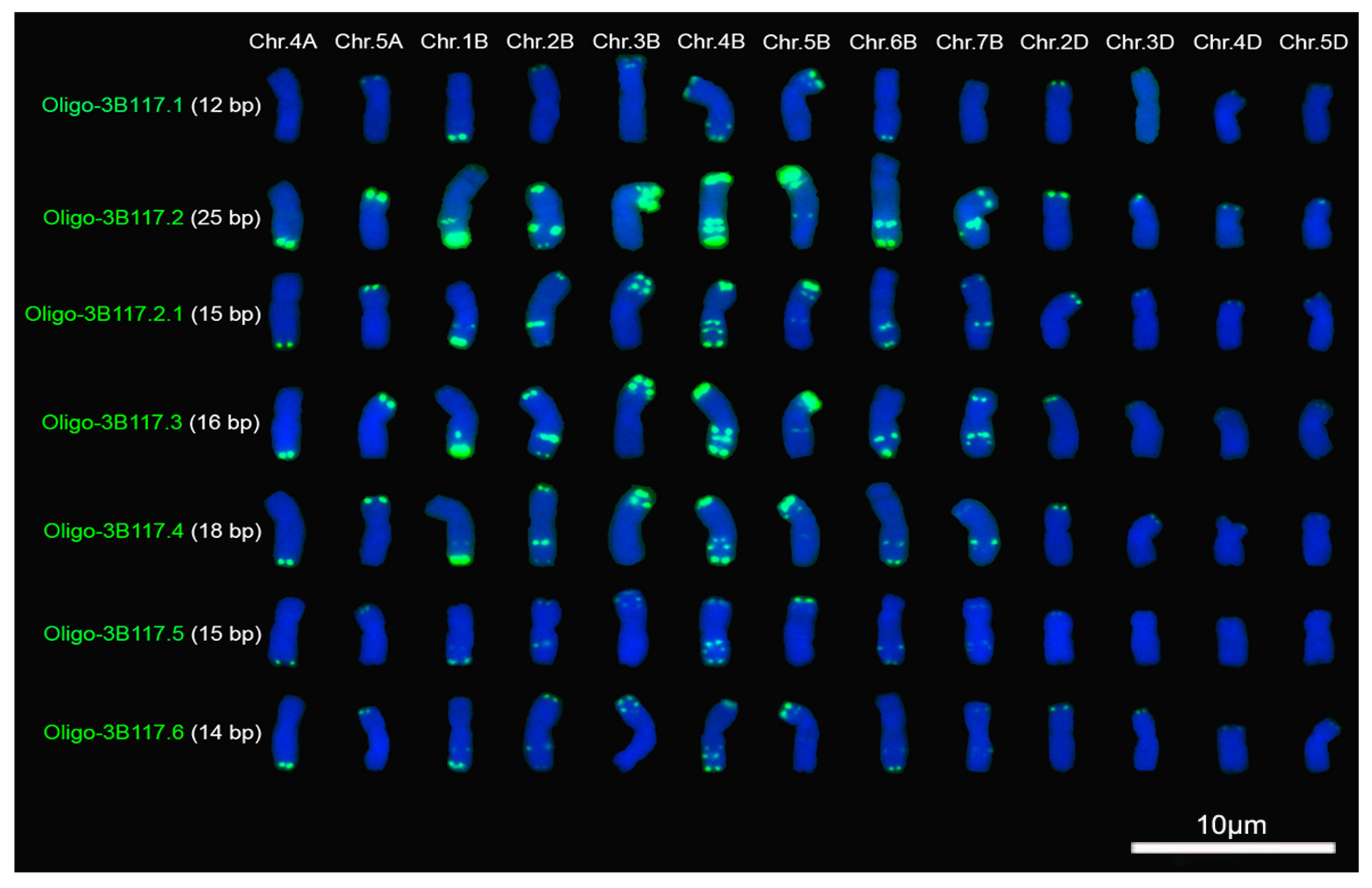
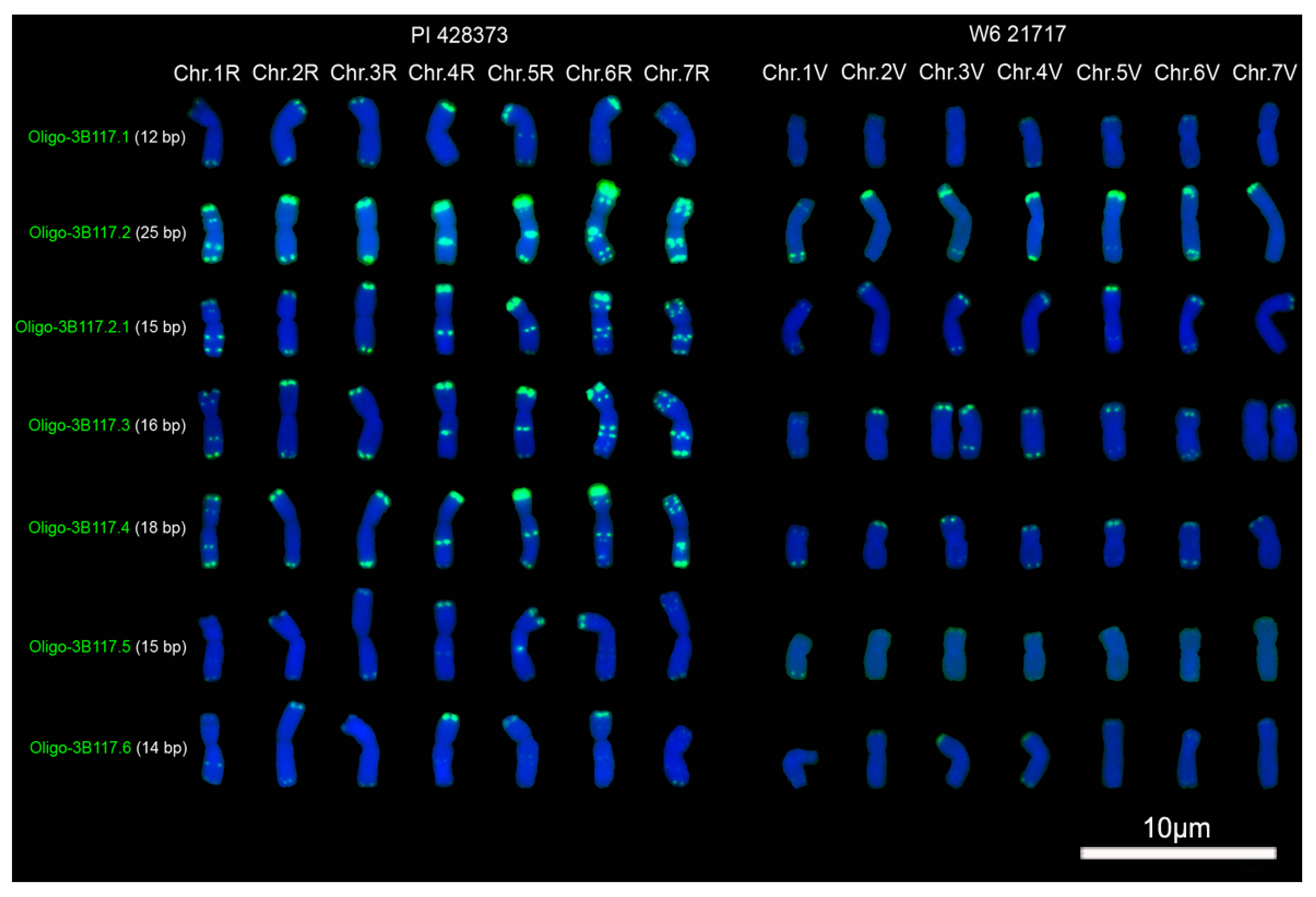
2.1. Signal Pattern of Different Segments of Tandem Repeat 3B117
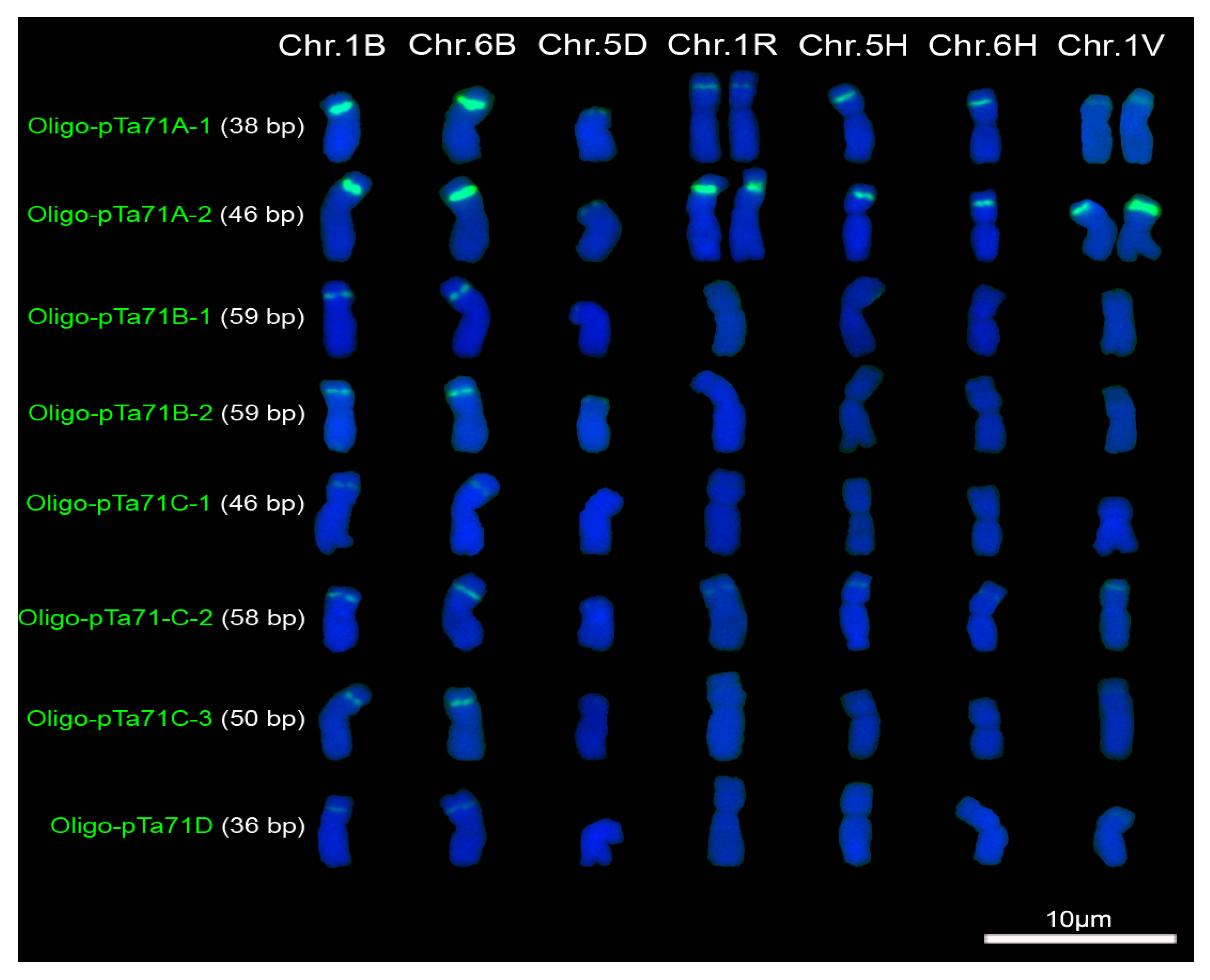
2.2. Signal Pattern of Different Repeated Family of pTa71
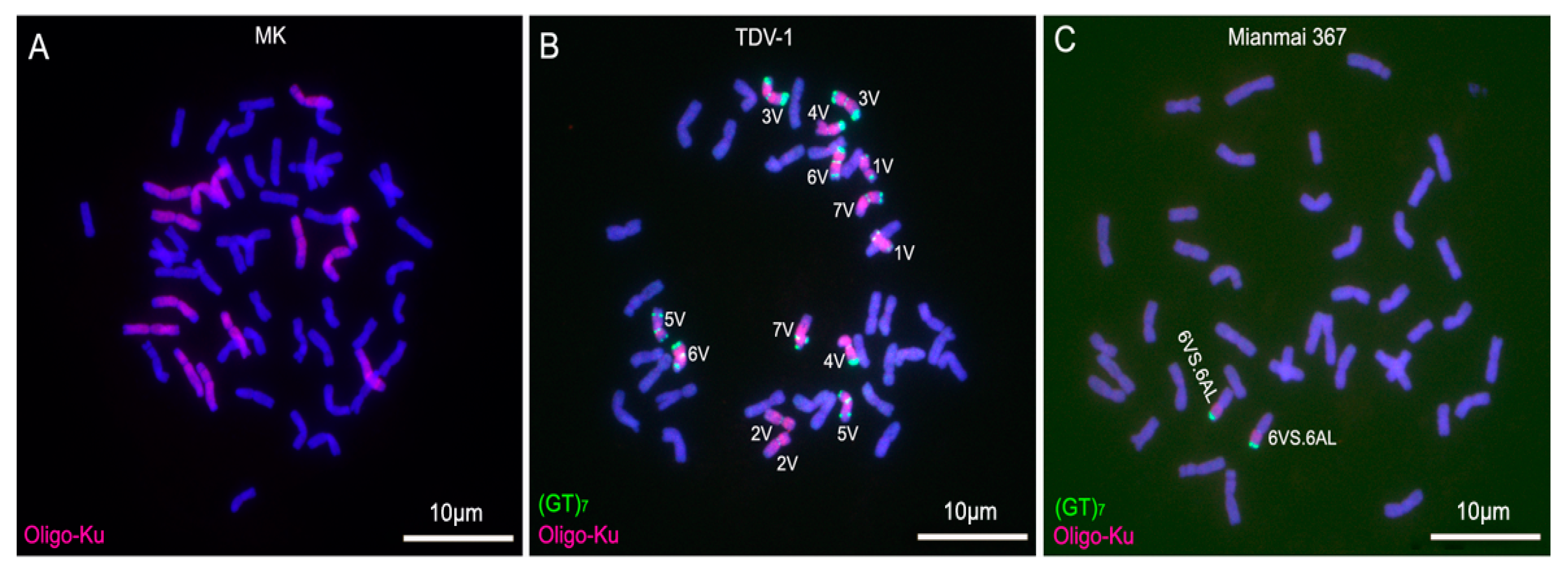
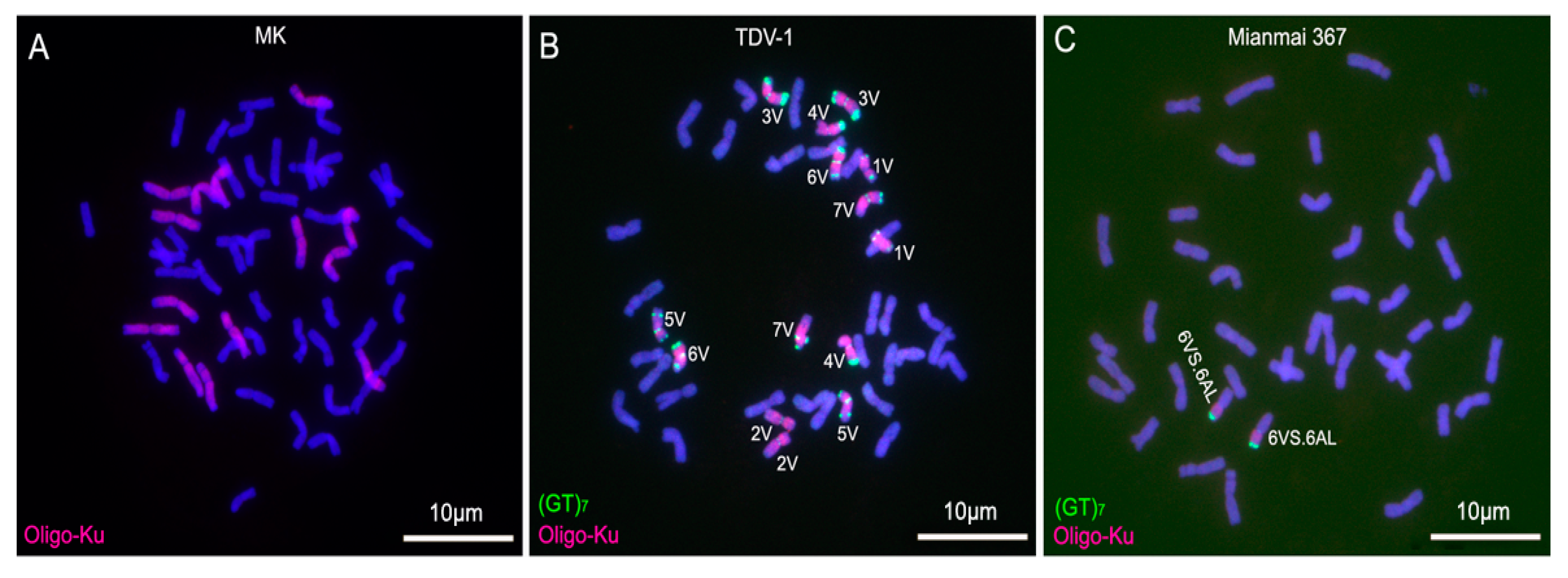
2.3. ND-FISH Analysis for Identifying D. villosum Chromosomes in Wheat Backgrounds
3. Discussion
3.1. The Factors Influencing the Signal Intensity of Oligonucleotide Probes
3.2. Oligonucleotide Probes and ND-FISH Reflecting Different Distribution Pattern of Tandem Repeats
3.3. Convenience of Oligonucleotide Probes and ND-FISH in Identifying D. villosum Chromosomes in Wheat Backgrounds
4. Materials and Methods
4.1. Plant Materials
4.2. Oligonucleotide Probe Development
4.3. ND-FISH Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cuadrado, Á.; Golczyk, H.; Jouve, N. A novel, simple and rapid nondenaturing FISH (ND-FISH) technique for the detection of plant telomeres. Potential used and possible target structures detected. Chromosome Res. 2009, 17, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, Á.; Jouve, N. Chromosomal detection of simple sequence repeats (SSRs) using nondenaturing FISH (ND-FISH). Chromosoma 2010, 119, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Pavia, I.; Carvalho, A.; Rocha, L.; Gaspar, M.J.; Lima-Brito, J. Physical location of SSR regions and cytogenetic instabilities in Pinus sylvestris chromosomes revealed by ND-FISH. J. Genet. 2014, 93, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Kirov, I.V.; Kiseleva, A.V.; Laere, K.V.; Roy, N.V.; Khrustaleva, L.I. Tandem repeats of Allium fistulosum associated with major chromosomal landmarks. Mol. Genet. Genom. 2017, 292, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Carmona, A.; Friero, E.; Bustos, A.D.; Jouve, N.; Cuadrado, A. Cytogenetic diversity of SSR motifs within and between Hordeum species carrying the H genome: H. vulgare L. and H. bulbosum L. Theor. Appl. Genet. 2013, 126, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Guedes-Pinto, H.; Lima-Brito, J. Polymorphism of the simple sequence repeat (AAC) 5 in the nucleolar chromosomes of old Portuguese wheat cultivars. J. Genet. 2013, 92, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Cabo, S.; Carvalho, A.; Martín, A.; Lima-Brito, J. Structural rearrangements detected in newly-formed hexaploid tritordeum after three sequential FISH experiments with repetitive DNA sequences. J. Genet. 2014, 93, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.X.; Yang, Z.J.; Fu, S.L. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.L.; Chen, L.; Wang, Y.Y.; Li, M.; Yang, Z.J.; Qiu, L.; Yan, B.J.; Ren, Z.L.; Tang, Z.X. Oligonucleotide probes for ND-FISH analysis to identify rye and wheat chromosomes. Sci. Rep. 2015, 5, 10552. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Amosova, A.V.; Goncharov, N.P.; Macas, J.; Ruban, A.S.; Grechishnikova, I.V.; Zoshchuk, S.A.; Houben, A. A set of cytogenetic markers allows the precise identification of all A-genome chromosomes in diploid and polyploidy wheat. Cytogenet. Genome Res. 2015, 146, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.; Carvalho, A.; Martín, A.C.; Martín, A.; Lima-Brito, J. Use of the synthetic Oligo-pTa535 and Oligo-pAs1 probes for identification of Hordeum chilense-origin chromosomes in hexaploid tritordeum. Genet. Resour. Crop 2016, 63, 945–951. [Google Scholar] [CrossRef]
- Li, G.R.; Gao, D.; Zhang, H.G.; Li, J.B.; Wang, H.J.; La, S.X.; Ma, J.W.; Yang, Z.J. Molecular cytogenetic characterization of Dasypyrum breviaristatum chromosomes in wheat background revealing the genomic divergence between Dasypyrum species. Mol. Cytogenet. 2016, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhuang, L.F.; Wang, Y.Z.; Yuan, Q.; Wang, D.R.; Dawadondup; Tan, L.J.; Shen, J.; Xu, H.B.; Zhao, H.; et al. Development of oligonucleotides and multiplex probes for quick and accurate identification of wheat and Thinopyrum bessarabicum chromosomes. Genome 2017, 60, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.Y.; Qiu, L.; Xiao, Z.Q.; Fu, S.L.; Tang, Z.X. New oligonucleotide probes for ND-FISH analysis to identify barley chromosomes and to investigate polymorphisms of wheat chromosomes. Genes 2016, 7, 118. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Luo, J.T.; Zhang, L.Q.; Yuan, Z.W.; Yang, Y.W.; Wu, M.; Chen, W.J.; Zheng, Y.L.; Zhang, H.G.; Liu, D.C. Production of hexaploid triticale by a synthetic hexaploid wheat-rye hybrid method. Euphytica 2013, 193, 347–357. [Google Scholar] [CrossRef]
- Mirzaghaderi, G.; Houben, A.; Badaeva, E. Molecular-cytogenetic analysis of Aegilops triuncialis and identification of its chromosomes in the background of wheat. Mol. Cytogenet. 2014, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.Q.; Zhang, M.Y.; Wang, X.E.; Chen, P.D. Introduction of chromosome segment carrying the seed storage protein genes from chromosome 1V of Dasypyrum villosum showed positive effect on bread-making quality of common wheat. Theor. Appl. Genet. 2014, 127, 523–533. [Google Scholar]
- Tang, Z.X.; Fu, S.L.; Ren, Z.L.; Zhang, T.; Zou, Y.T.; Yang, Z.J.; Li, G.R.; Zhou, J.P.; Zhang, H.Q.; Yan, B.J.; et al. Diversity and evolution of four dispersed repetitive DNA sequences in the genus Secale. Genome 2011, 54, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.F.; Harberd, N.P.; Jarvis, M.G.; Flavell, R.B. Structure and evolution of the intergenic region in a ribosomal DNA repeat unit of wheat. J. Mol. Biol. 1988, 201, 1–7. [Google Scholar] [CrossRef]
- Contento, A.; Hslop-Harrison, J.S.; Schwarzacher, T. Diversity of a major repetitive DNA sequence in diploid and polyploid Triticeae. Cytogenet. Genome Res. 2005, 109, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.J. A model of DNA repeat-assembled mitotic chromosomal skeleton. Genes 2011, 2, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Qiu, L.; Li, M.; Tang, S.Y.; Fu, S.L.; Tang, Z.X. Molecular detection and FISH analysis of powdery mildew resistance gene in wheat cultivars Mianmai 37 and Mianmai 367. J. Triticeae Crops 2015, 35, 1502–1505. [Google Scholar]
- Zhang, R.Q.; Sun, B.X.; Cheng, J.; Cao, A.Z.; Xing, L.P.; Feng, Y.G.; Lan, C.X.; Chen, P.D. Pm55, a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat. Theor. Appl. Genet. 2016, 129, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, C.L.; Pereira, S.; Moran, L.B.; Appels, R. New Secale cereale (rye) DNA derivatives for the detection of rye chromosome segments in wheat. Genome 1990, 33, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Han, F.P.; Lamb, J.C.; Birchler, A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe | Nucleotide Sequence and Fluorochrome Label | Length of Probe (bp) | Amount Used for Each Slide (pmol/slide) | Original Sequences Used to Develop Probes (GenBank Accession Number) |
|---|---|---|---|---|
| Oligo-Ku | Tamra-5′GATCG AGACT TCTAG CAATA GGCAA AAATA GTAAT GGTAT CCGGG TTCG 3′ | 49 | 0.68 | Rye dispersed repetitive sequence KU.D15.15 (GU318080.1) |
| Oligo-3B117.1 | 6-FAM-5′CCCCGGGGTGCG3′ | 12 | 2.0 | 3B117, obtained from sequence of IWGSC_RefSeq_V1_chr3B.fsa |
| Oligo-3B117.2 | 6-FAM-5′ACGTGTCGGTCATCAACACTCACAG3′ | 25 | 2.0 | |
| Oligo-3B117.2.1 | 6-FAM-5′ACGTGTCGGTCATCA3′ | 15 | 2.0 | |
| Oligo-3B117.3 | 6-FAM-5′GGCCGATTCTGGCCCG3′ | 16 | 2.0 | |
| Oligo-3B117.4 | 6-FAM-5′CGTGGACTATTACTCAGC3′ | 18 | 2.0 | |
| Oligo-3B117.5 | 6-FAM-5′GGGGTCCCAGAGTGA3′ | 15 | 2.0 | |
| Oligo-3B117.6 | 6-FAM-5′CCACGATTGACGAA3′ | 14 | 2.0 | |
| Oligo-pTa71A-1 | 6-FAM-5′CCGTG AACGG GCTGT ACGAG GACAC GGGAA AAAAC TGG3′ | 38 | 1.0 | Wheat ribosomal DNA (rDNA) 25S-18S intergenic region EcoRI-BamHI fragment, repeat family A (X07841.1) |
| Oligo-pTa71A-2 | 6-FAM-5′CCGAC GGCCG TCGTG GACGG AAGTT GACGC GCGCC ATGGA AAACT G3′ | 46 | 1.0 | |
| Oligo-pTa71B-1 | 6-FAM-5′AAATG GCTAA GTCCC TTGTA AGACA TACCC TTGGA CCCAA TAAGG CCAGT GGAAA CAAC3′ | 59 | 1.0 | Wheat ribosomal DNA (rDNA) 25S-18S intergenic region EcoRI-BamHI fragment, repeat family B (X07841.1) |
| Oligo-pTa71B-2 | 6-FAM-5′TACTT GGCCG ATTCA TGCGG ATGCC GTCGT CAGAG GCTAC ACTGC TAAGT CATGG TCAA3′ | 59 | 1.0 | |
| Oligo-pTa71C-1 | 6-FAM-5′GTCGC CTCCG GAAAA ACGTT GCCCC TCGGT GGCAA CGTCA TCGCT GT3′ | 46 | 1.0 | Wheat ribosomal DNA (rDNA) 25S-18S intergenic region EcoRI-BamHI fragment, repeat family C (X07841.1) |
| Oligo-pTa71C-2 | 6-FAM-5′TGTAC GTCTC AAGTG AAATT CTAAC CCAAC AGCCG AATGC GGCTC GGGAA ACAGG AAA3′ | 58 | 1.0 | |
| Oligo-pTa71C-3 | 6-FAM-5′CCCGT TGCGT ACACG ATCCG ACCGA CGGTA AACAG TCGCA ACGAT GTCCC3′ | 50 | 1.0 | |
| Oligo-pTa71D | 6-FAM-5′CATGT CTCAT GGCAA AAAAA CGCTG CCACG GCAGC G 3′ | 36 | 1.0 | Wheat ribosomal DNA (rDNA) 25S-18S intergenic region EcoRI-BamHI fragment, repeat family D (X07841.1) |
| (GT)7 | 6-FAM-5′GTGTGTGTGTGTGT3′ | 14 | 1.77 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Z.; Tang, S.; Qiu, L.; Tang, Z.; Fu, S. Oligonucleotides and ND-FISH Displaying Different Arrangements of Tandem Repeats and Identification of Dasypyrum villosum Chromosomes in Wheat Backgrounds. Molecules 2017, 22, 973. https://doi.org/10.3390/molecules22060973
Xiao Z, Tang S, Qiu L, Tang Z, Fu S. Oligonucleotides and ND-FISH Displaying Different Arrangements of Tandem Repeats and Identification of Dasypyrum villosum Chromosomes in Wheat Backgrounds. Molecules. 2017; 22(6):973. https://doi.org/10.3390/molecules22060973
Chicago/Turabian StyleXiao, Zhiqiang, Shuyao Tang, Ling Qiu, Zongxiang Tang, and Shulan Fu. 2017. "Oligonucleotides and ND-FISH Displaying Different Arrangements of Tandem Repeats and Identification of Dasypyrum villosum Chromosomes in Wheat Backgrounds" Molecules 22, no. 6: 973. https://doi.org/10.3390/molecules22060973
APA StyleXiao, Z., Tang, S., Qiu, L., Tang, Z., & Fu, S. (2017). Oligonucleotides and ND-FISH Displaying Different Arrangements of Tandem Repeats and Identification of Dasypyrum villosum Chromosomes in Wheat Backgrounds. Molecules, 22(6), 973. https://doi.org/10.3390/molecules22060973