
Quercetin from Polygonum capitatum Protects against Gastric Inflammation and Apoptosis Associated with Helicobacter pylori Infection by Affecting the Levels of p38MAPK, BCL-2 and BAX

Abstract
:1. Introduction
2. Results and Discussion
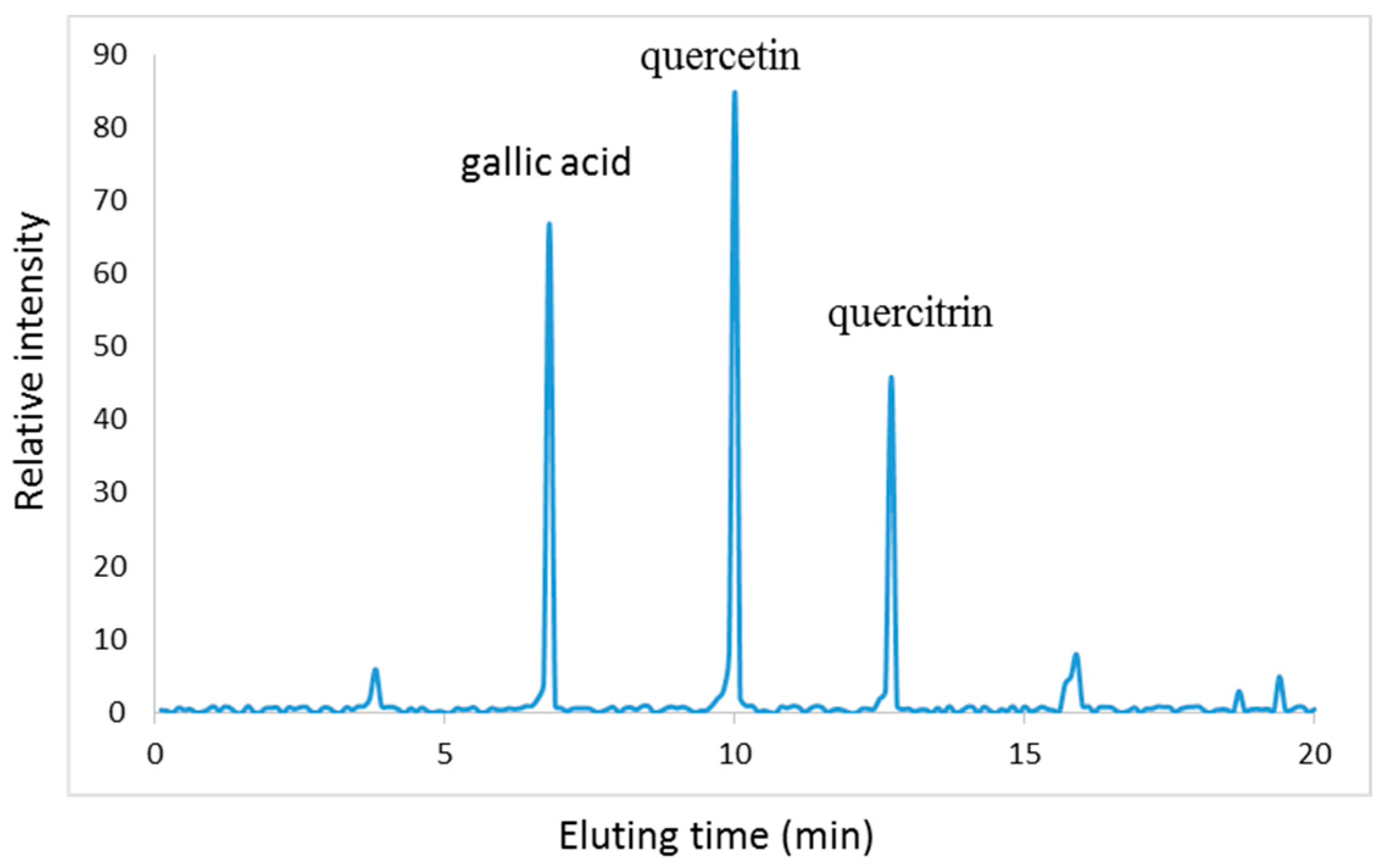
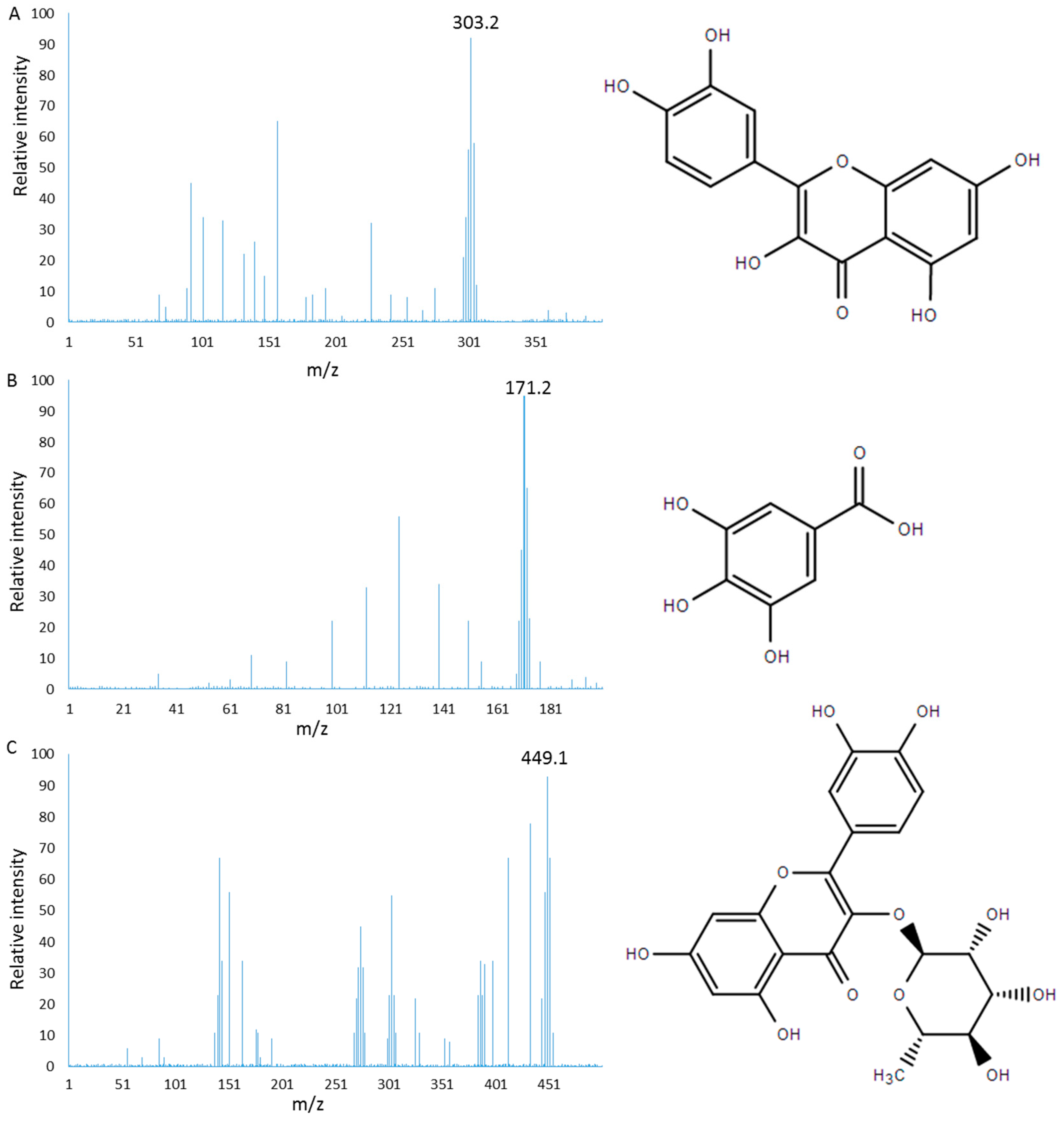
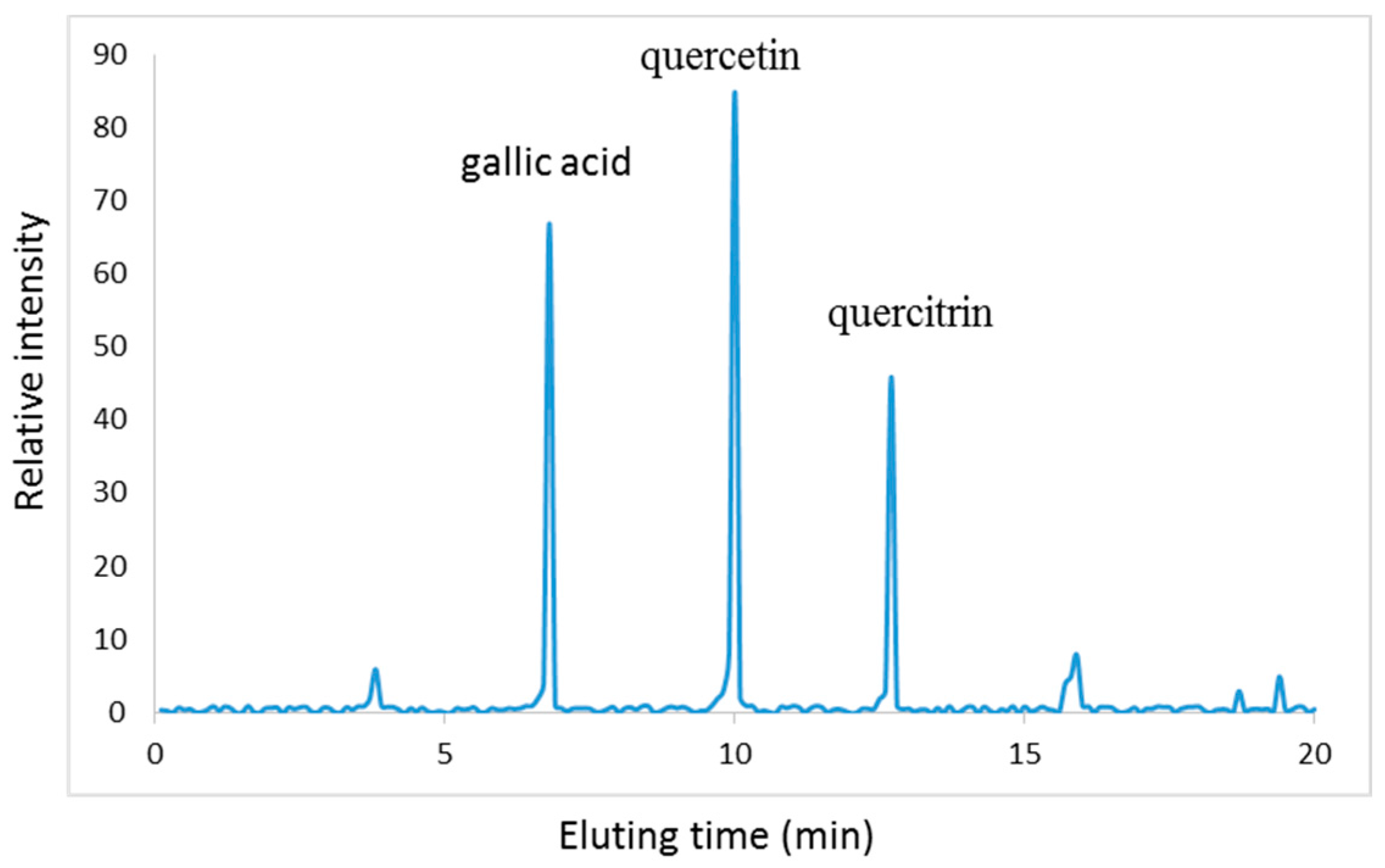
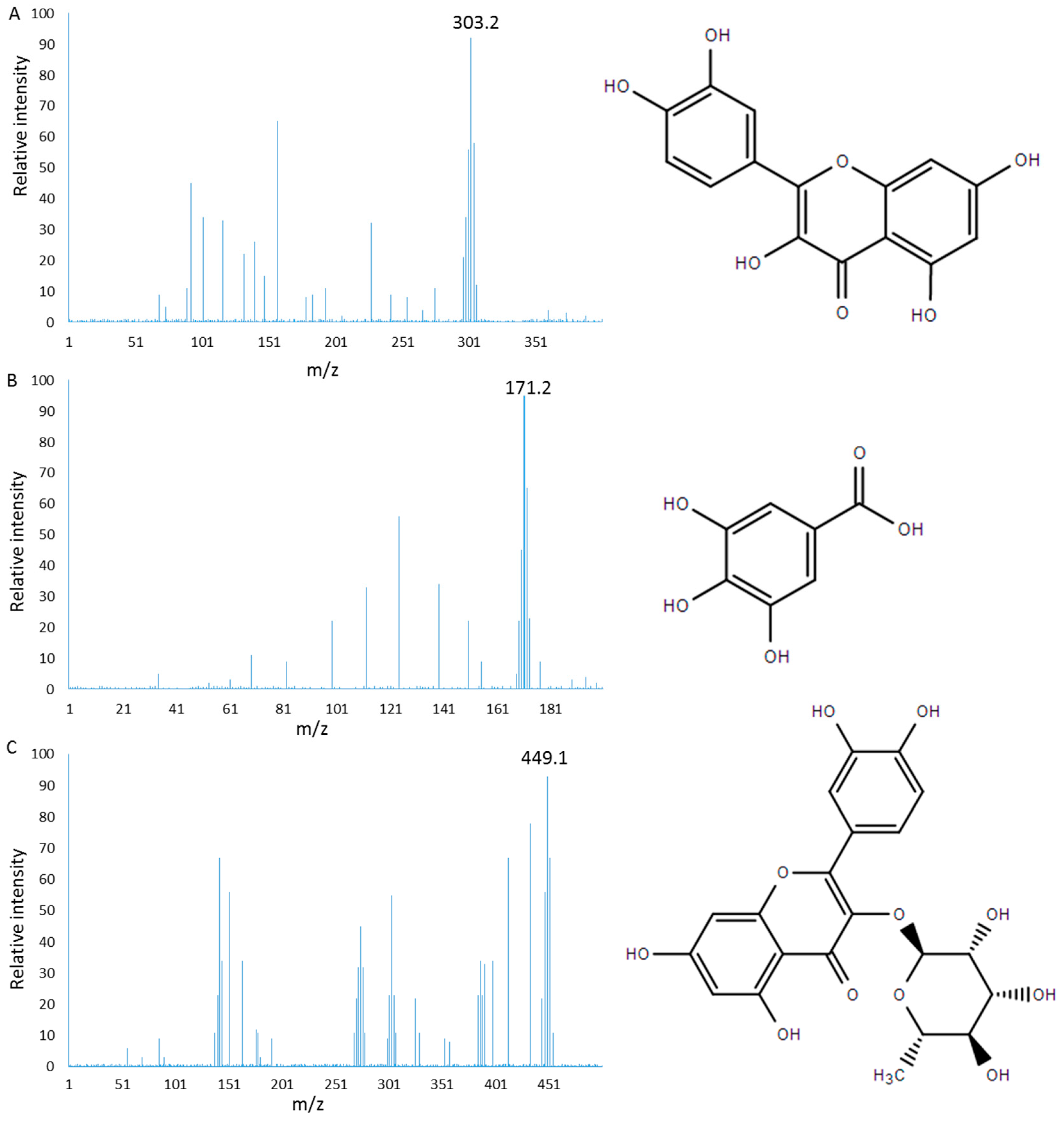
2.1. Characterization of PC
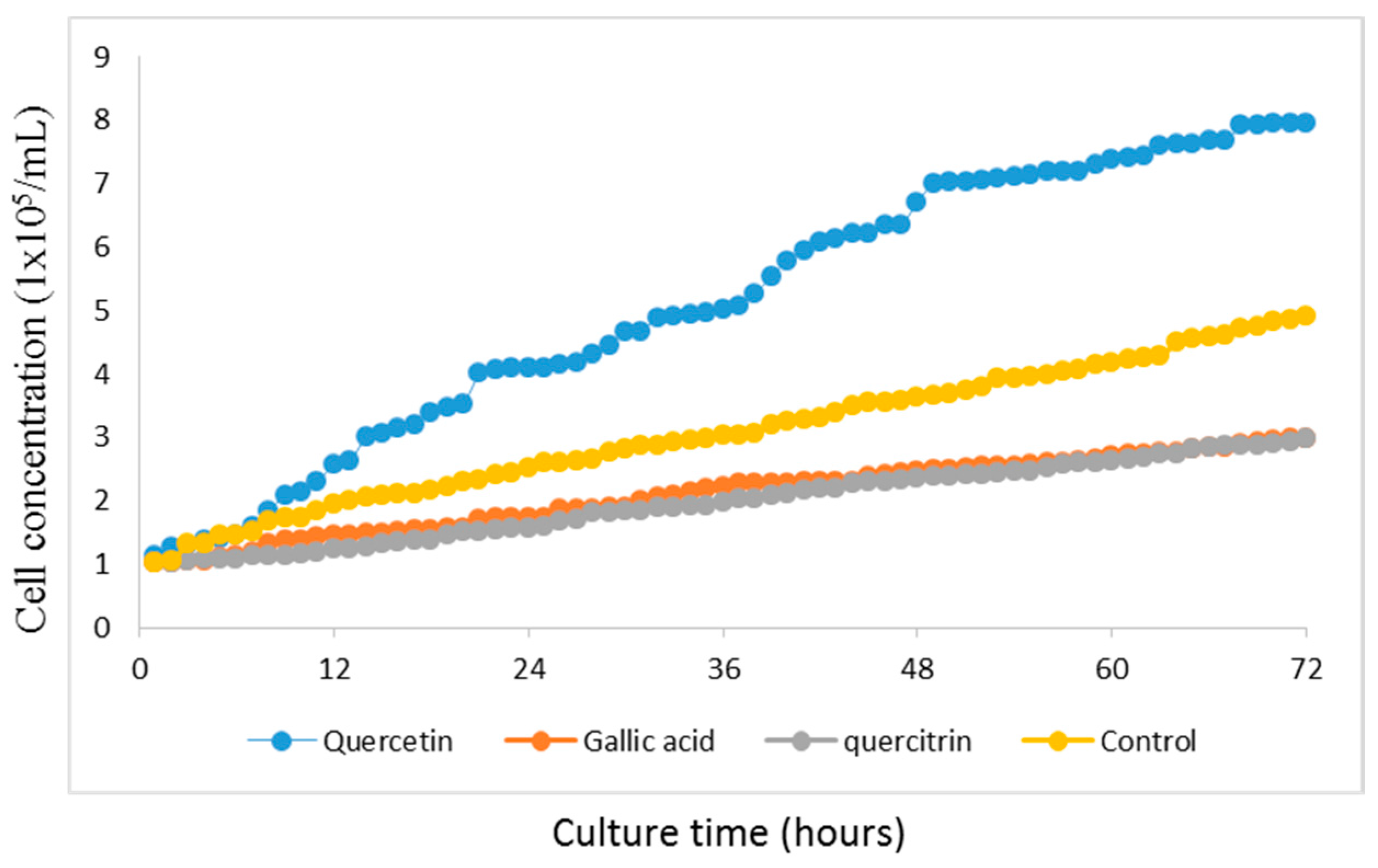
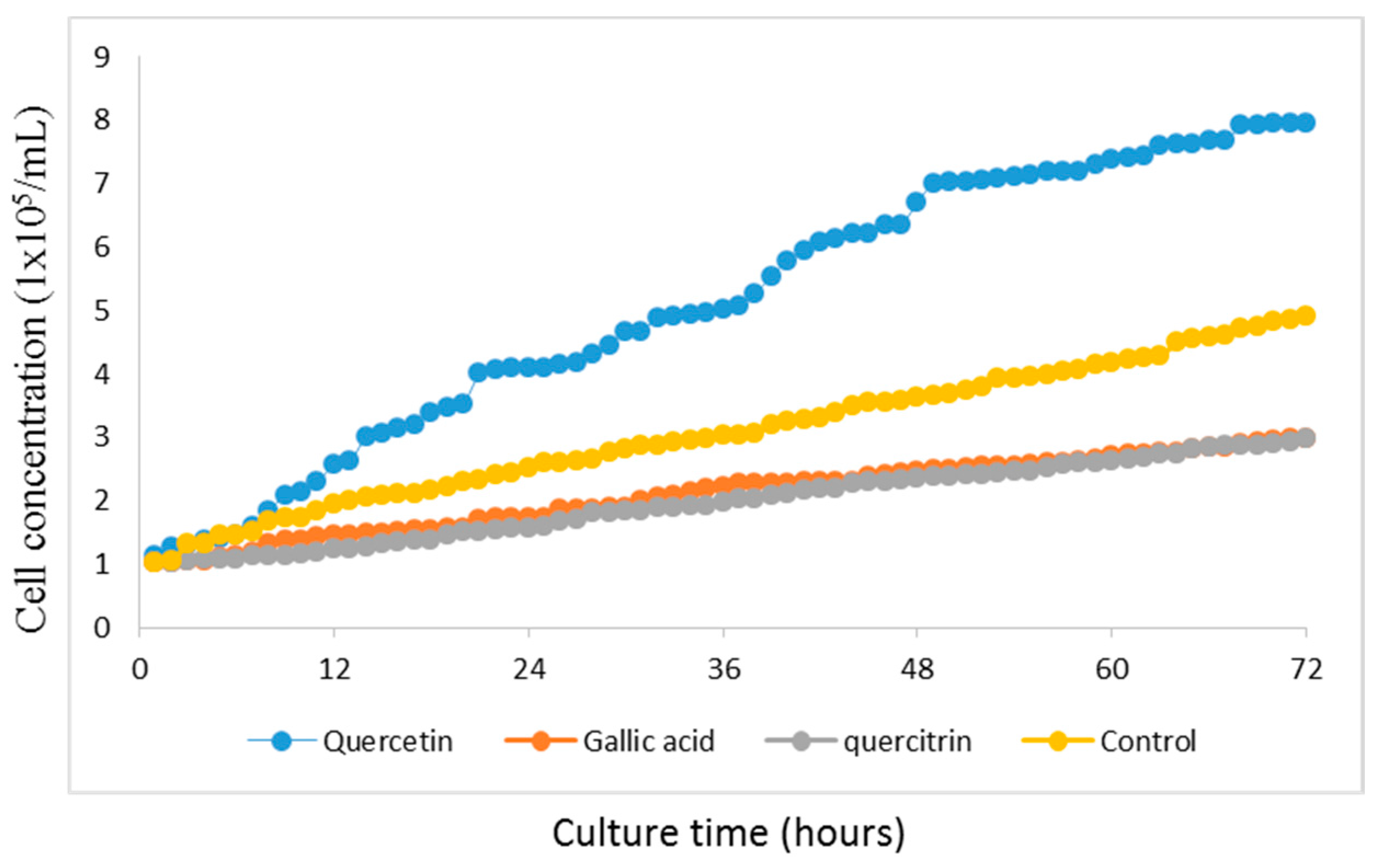
2.2. GES-1 Growth Curve
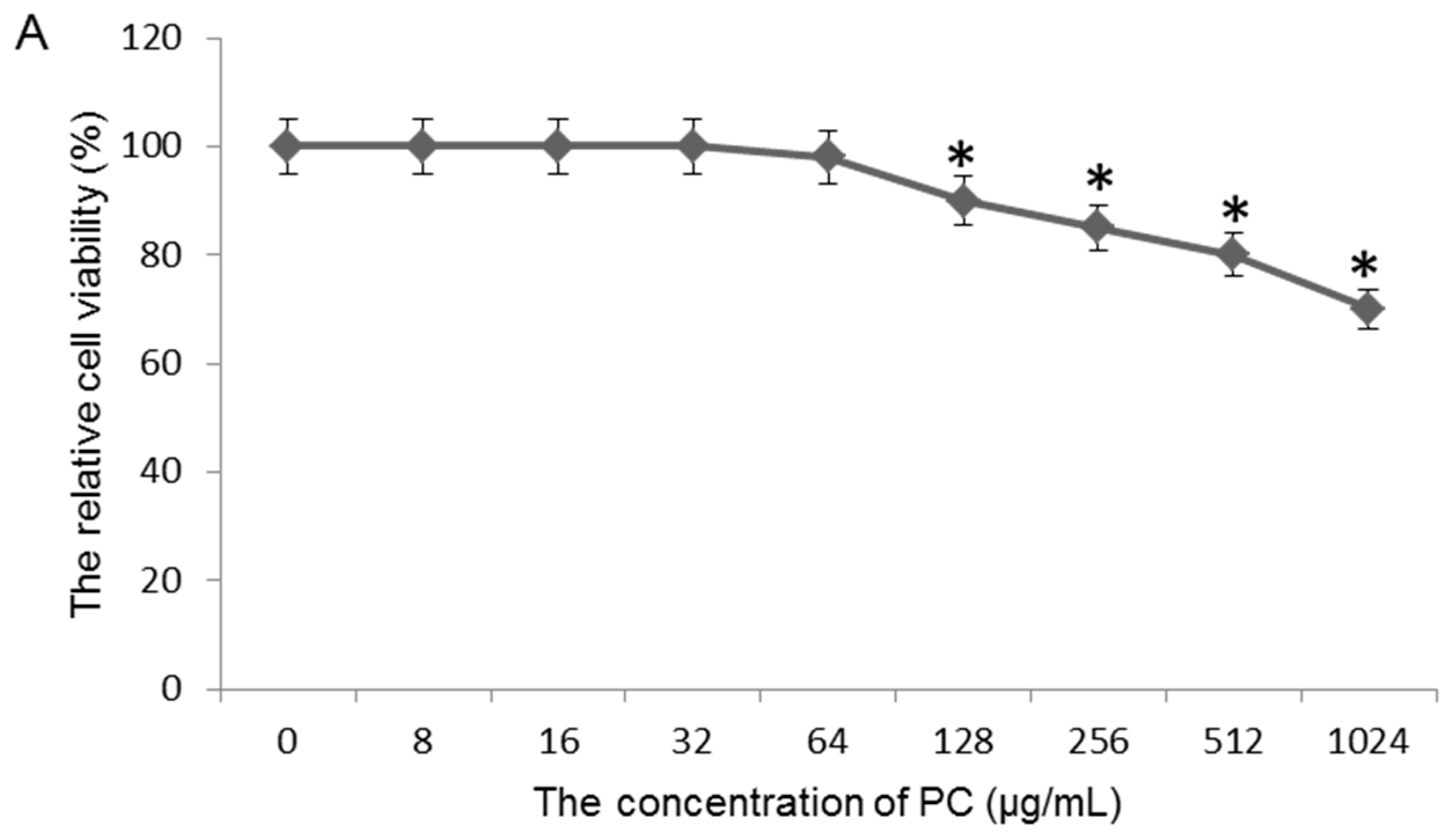
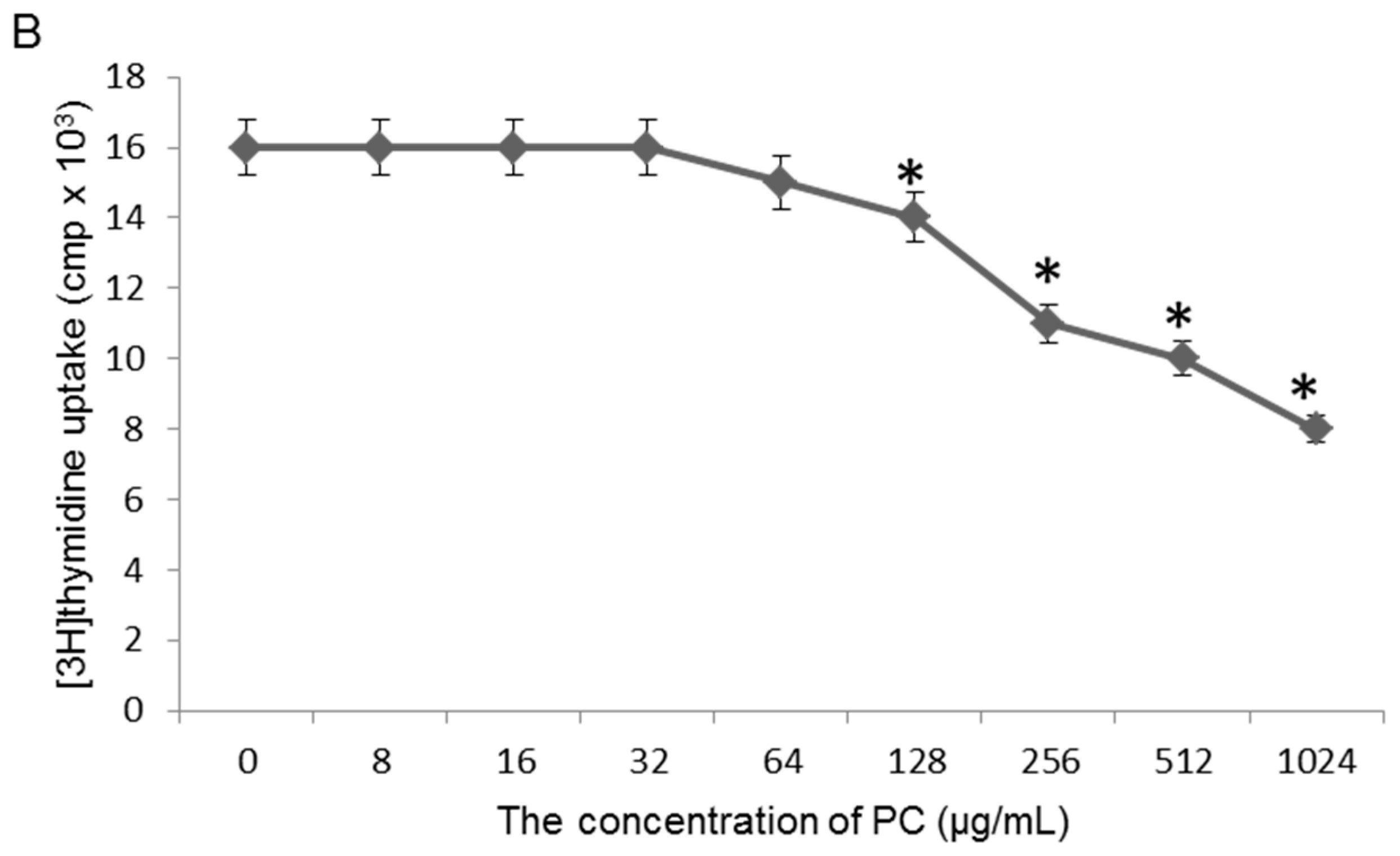
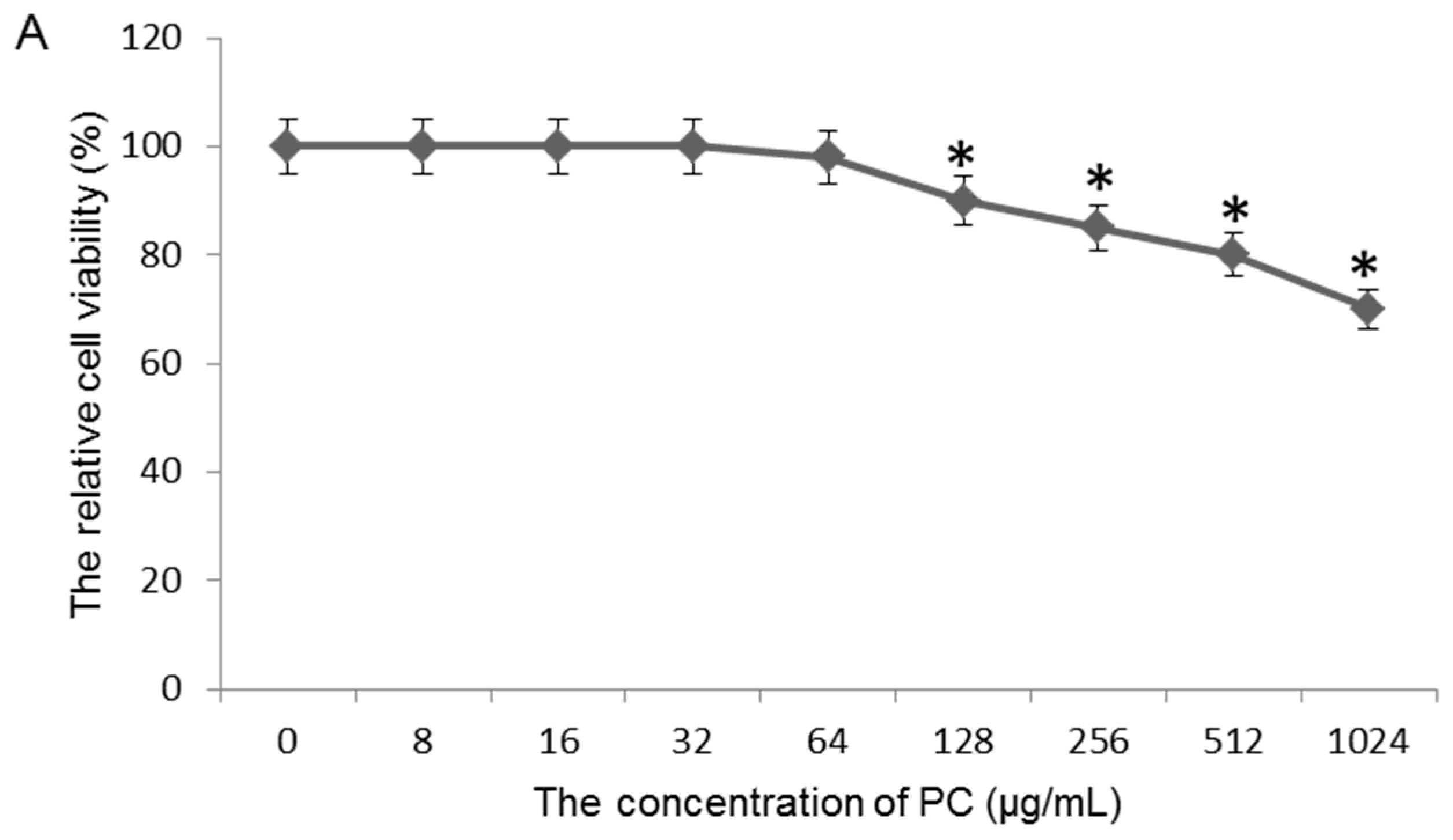
2.3. Effects of Quercetin on GES-1 Cells
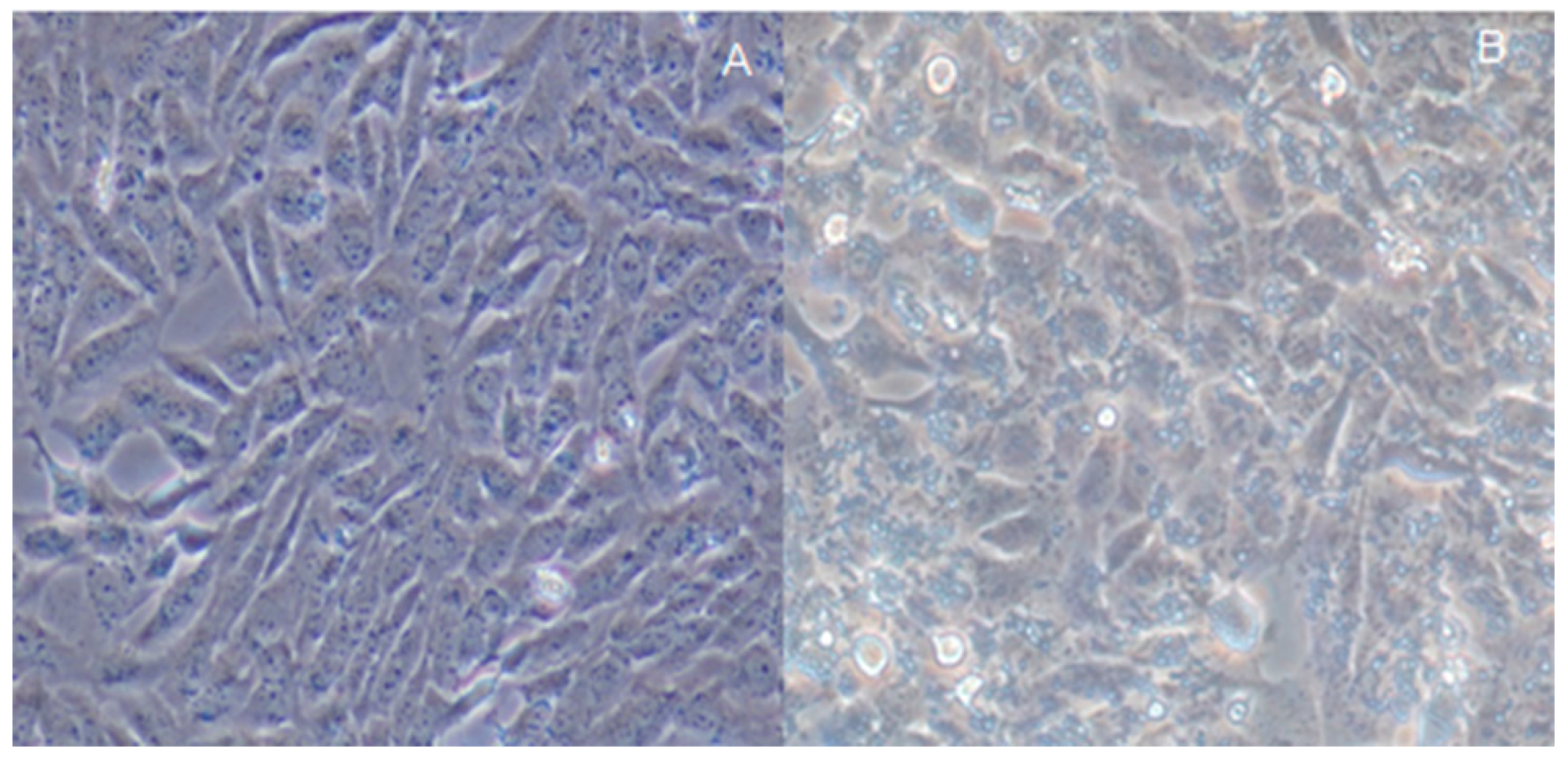
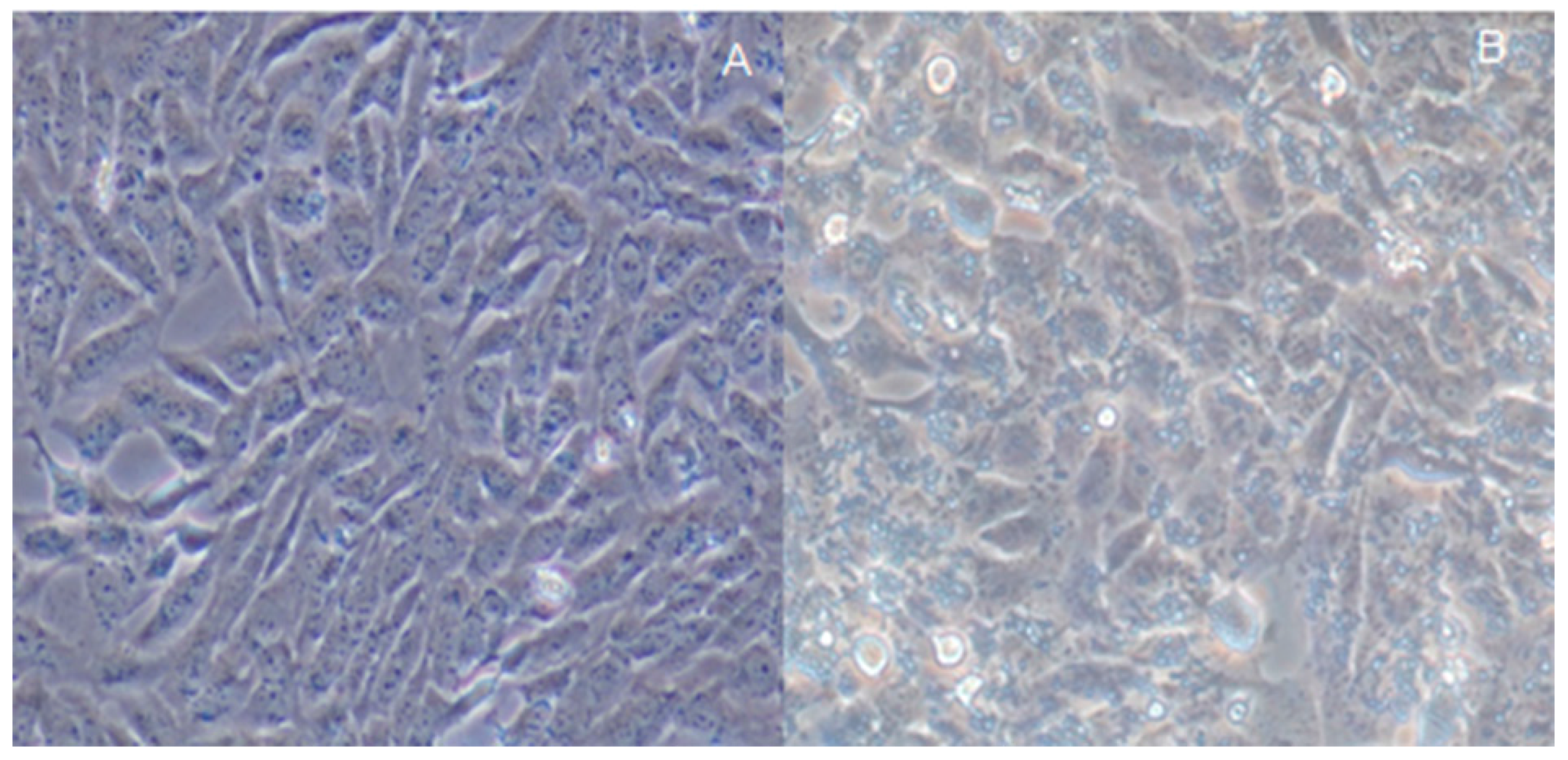
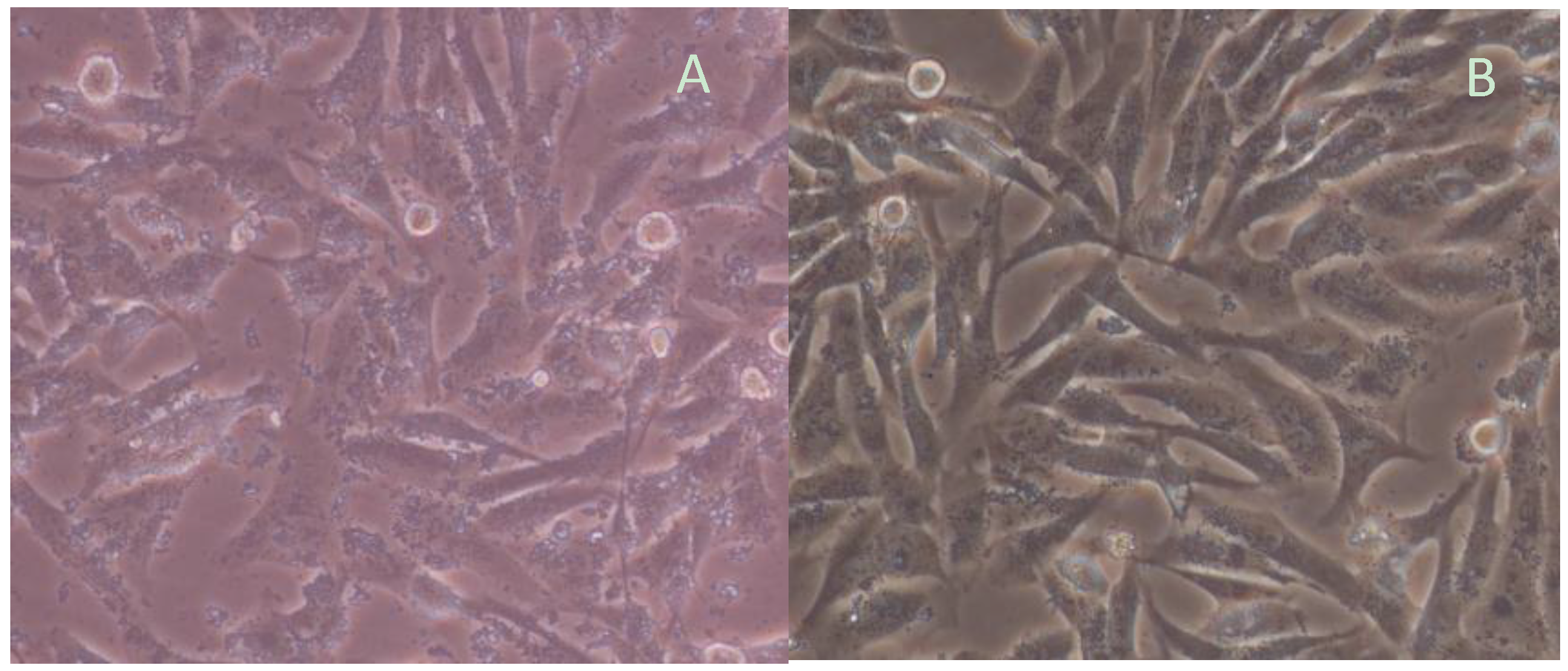
2.4. Assay of H. pylori-Infected Mouse Model
2.5. Effects of Quercetin on Mouse Model Inflammation
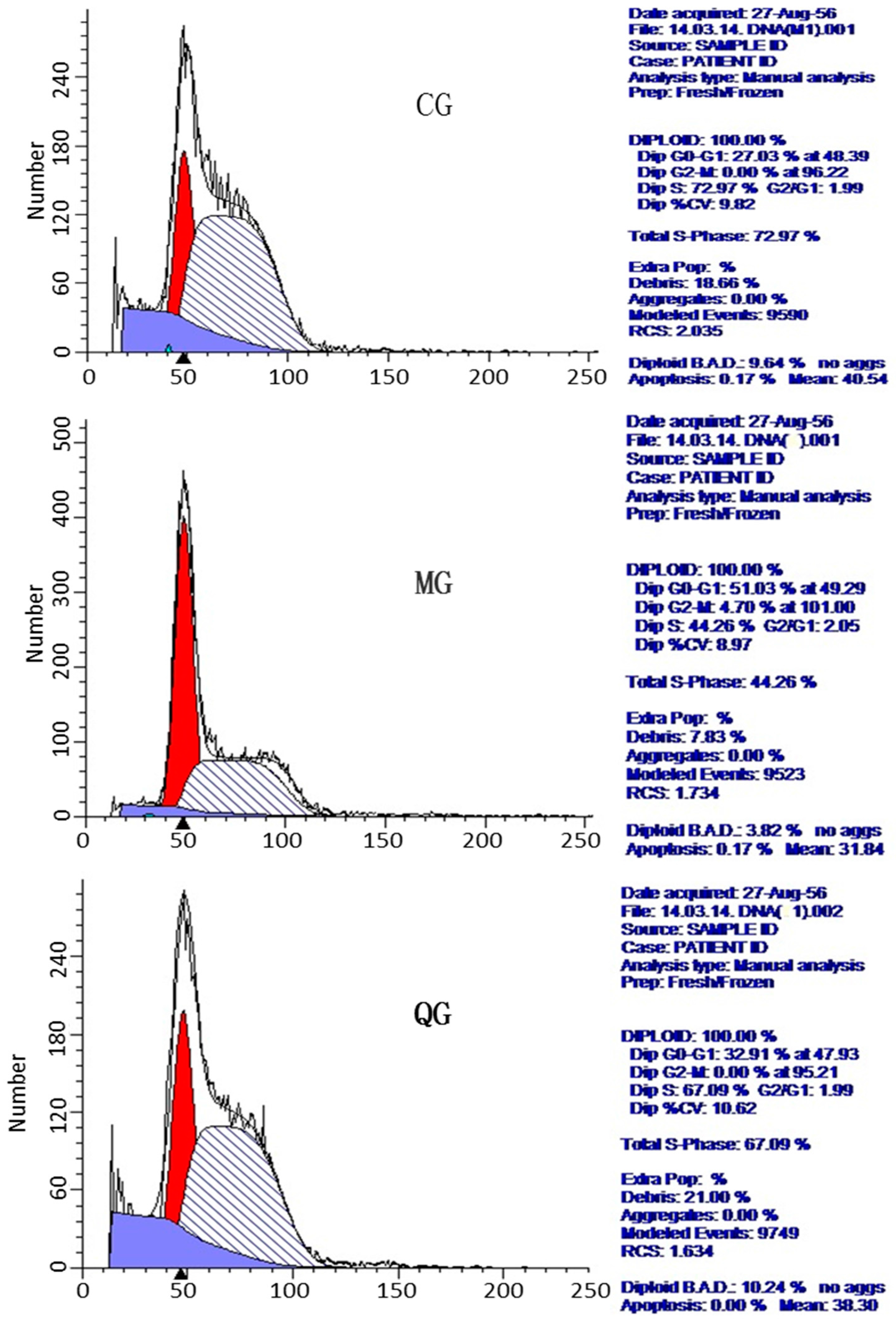
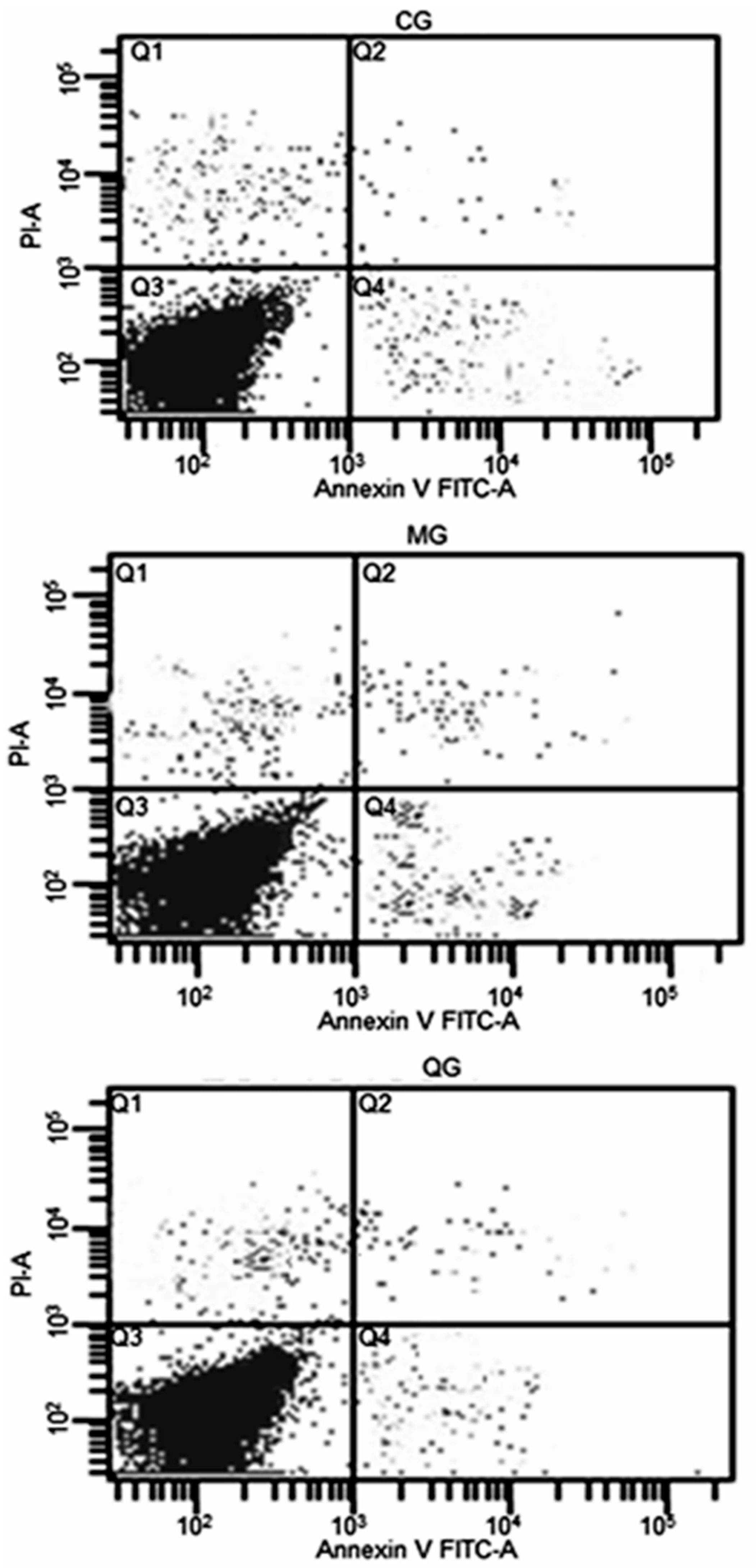
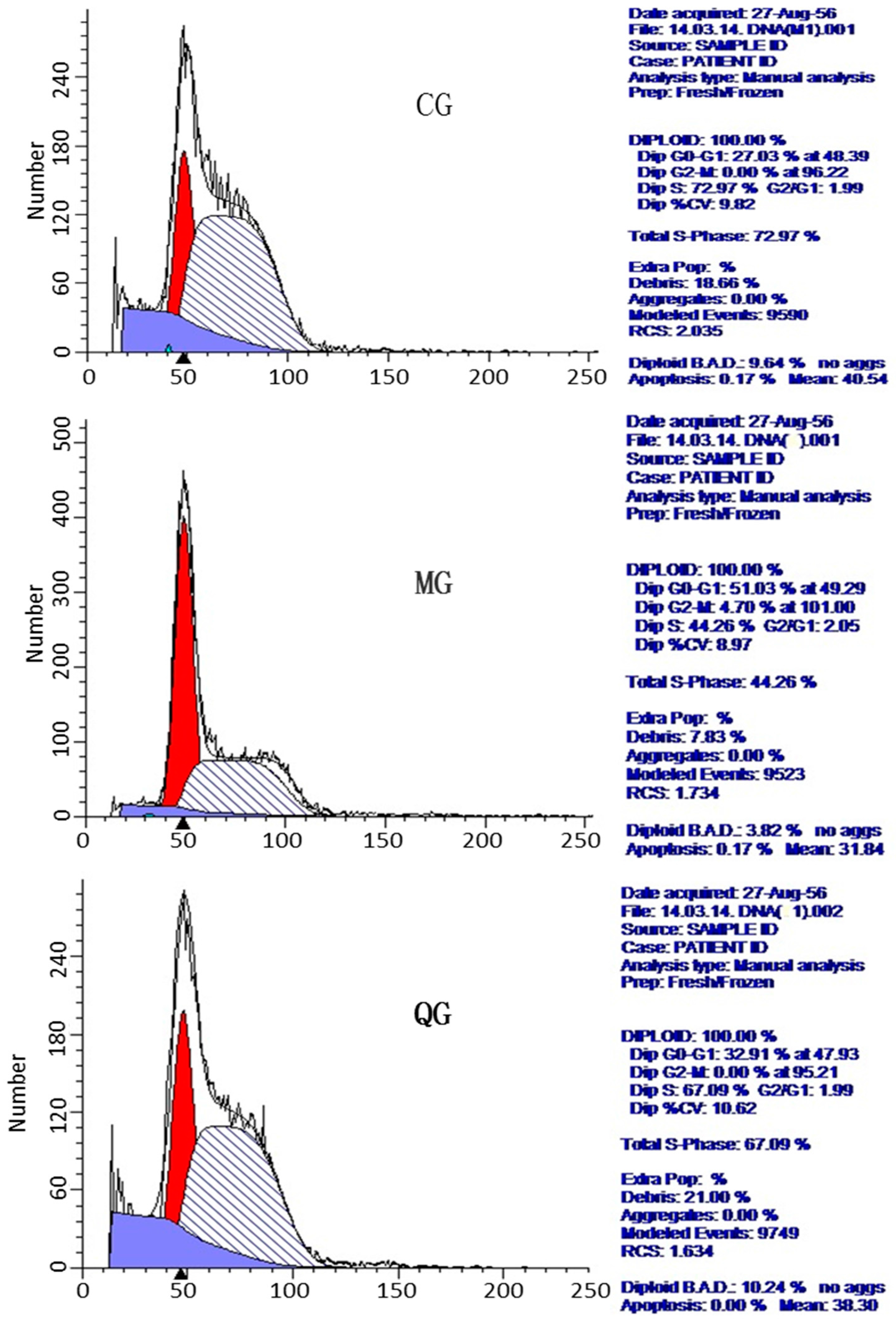
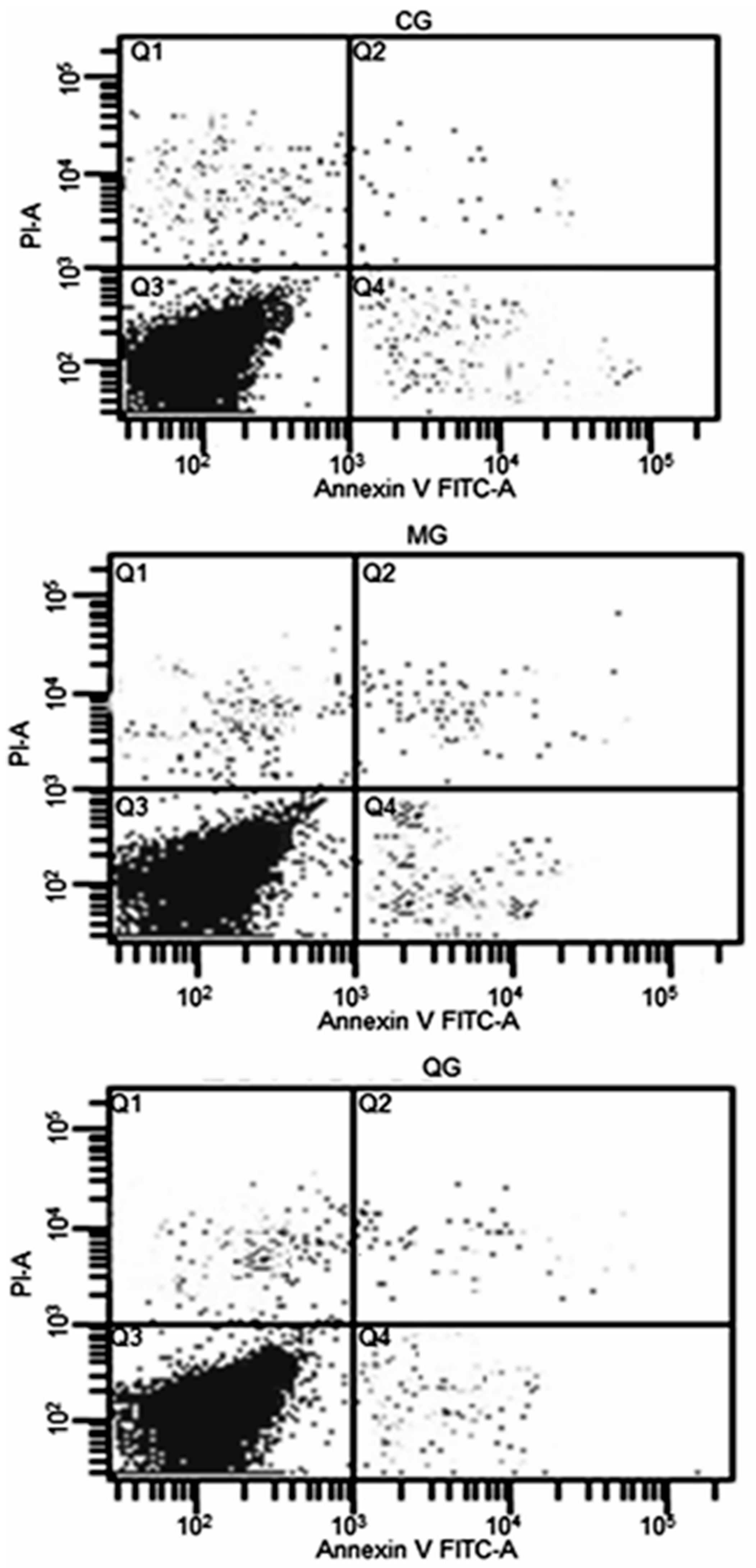
2.6. The Effects of Quercetin on Cell Cycle and Apoptosis
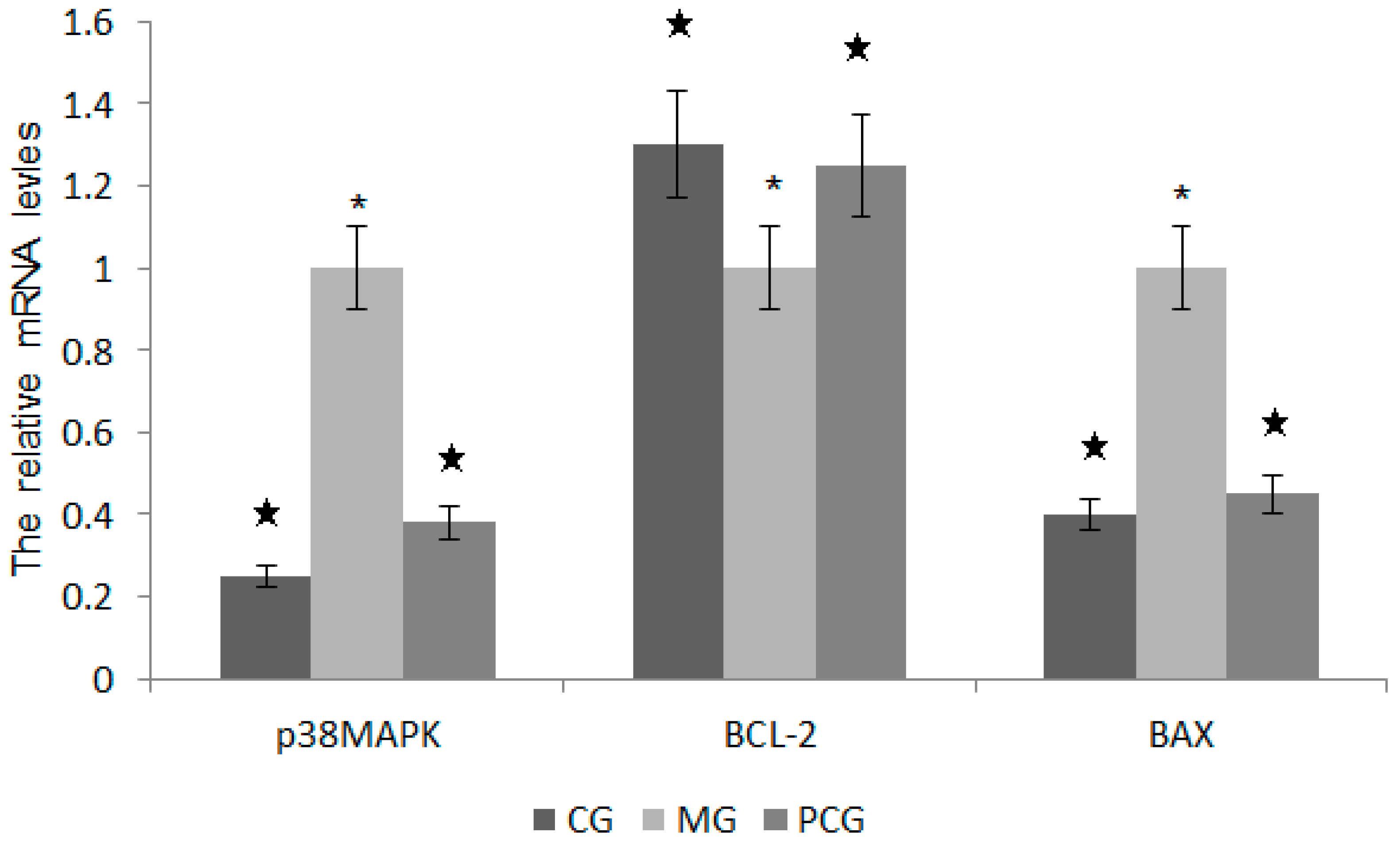
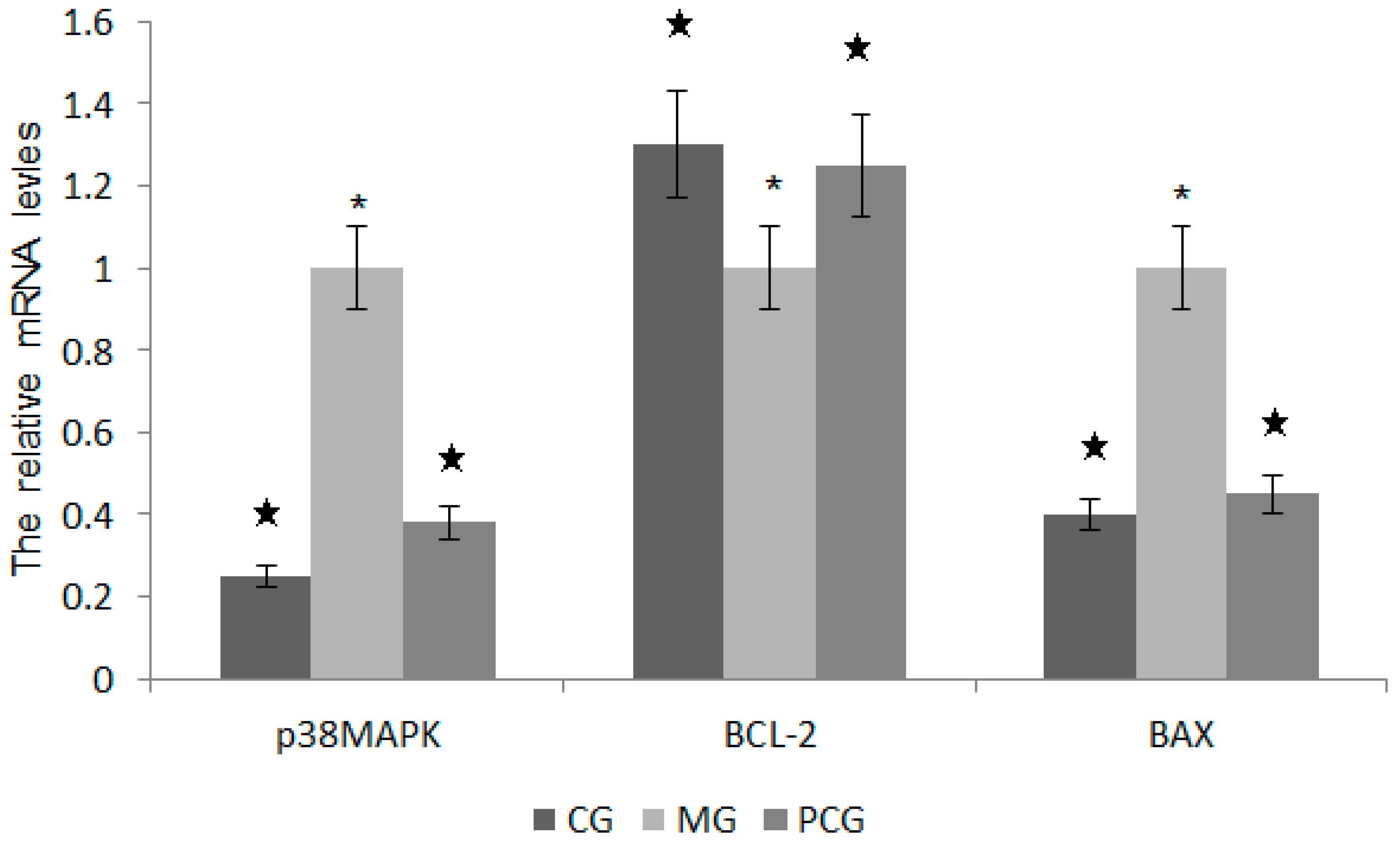
2.7. The Effects of Quercetin on the mRNA Levels of p38MAPK, BCL-2 and BAX
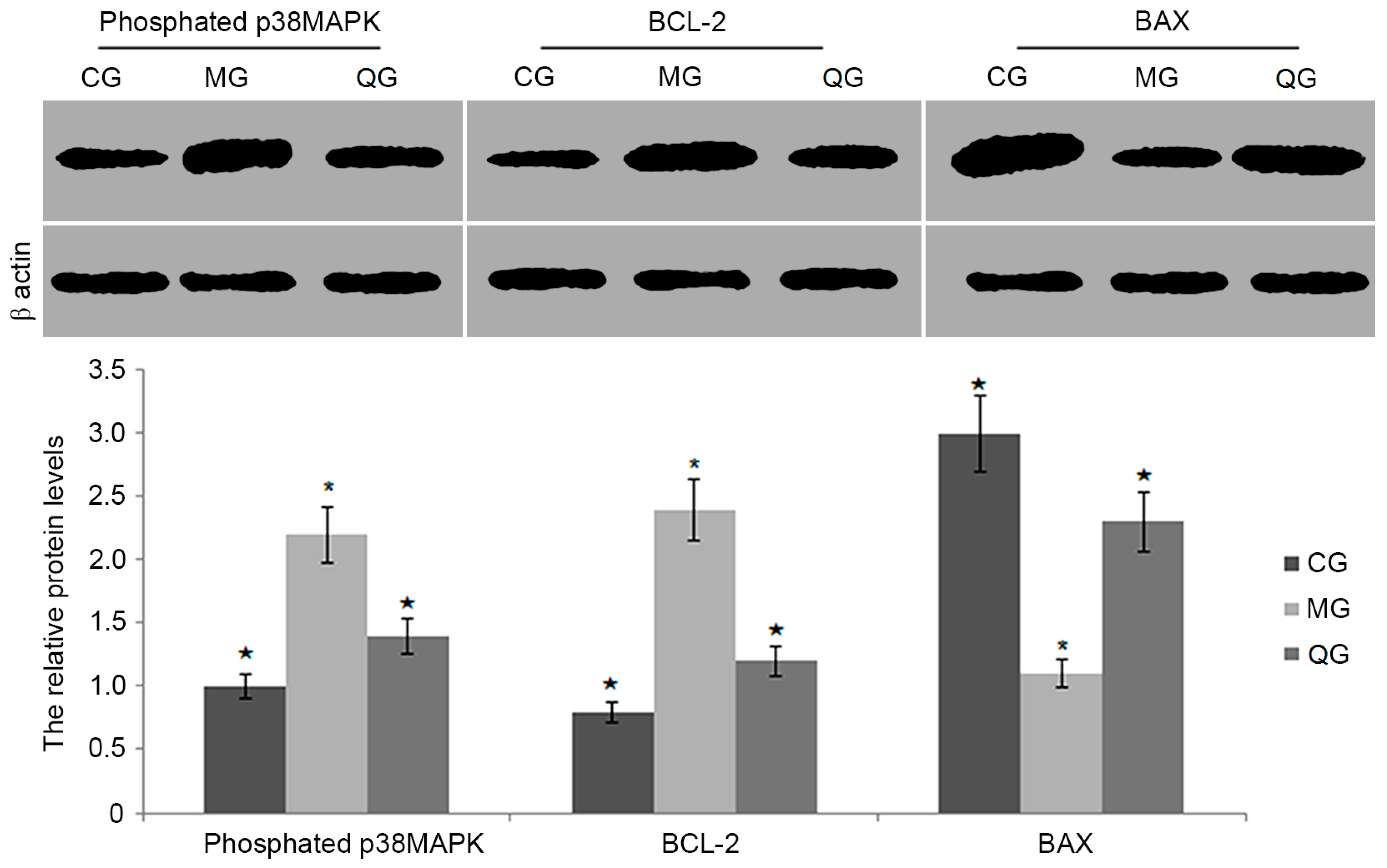
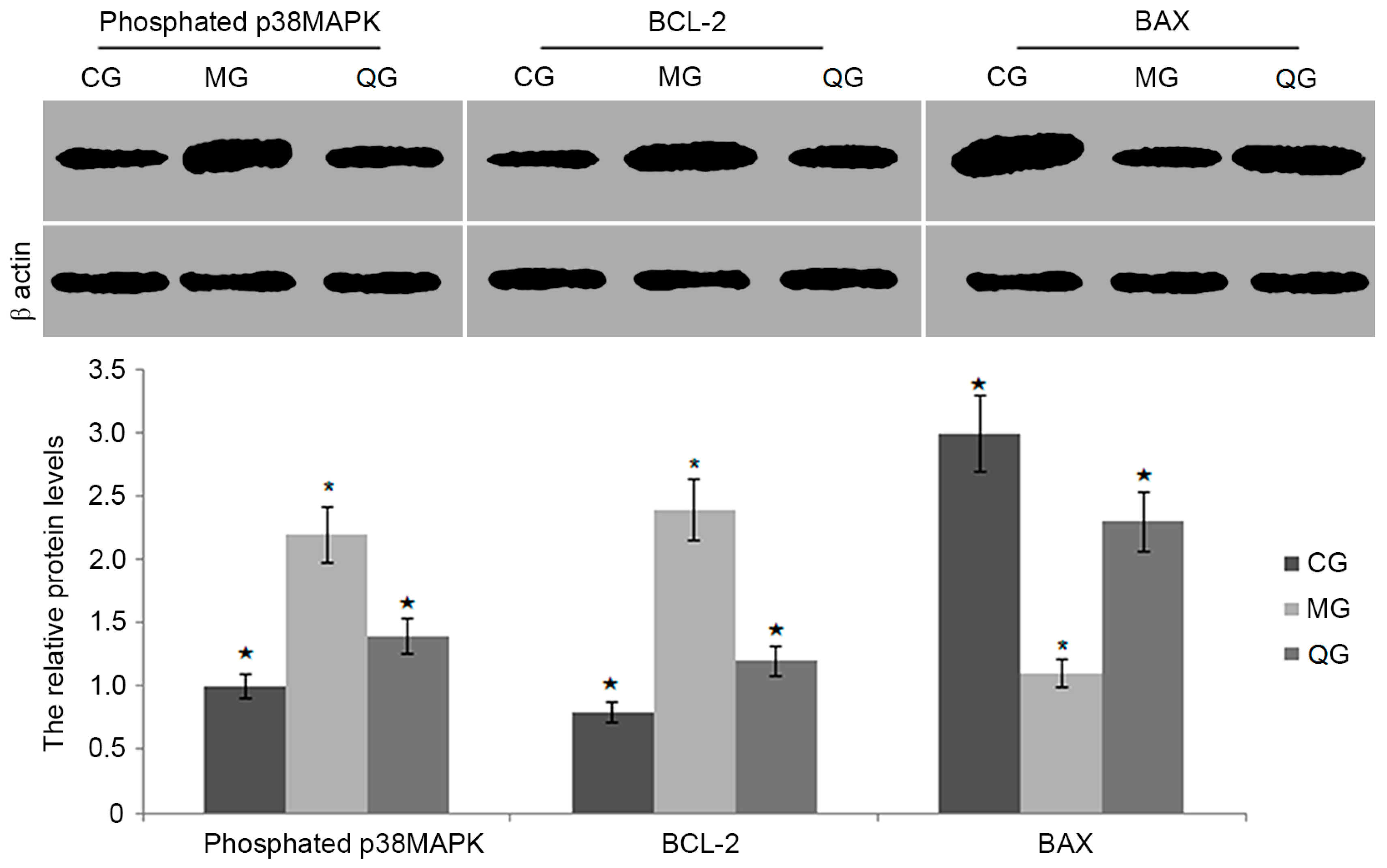
2.8. The Effects of Quercetin on the Protein Levels of BCL-2, BAX and Phosphated p38MAPK
3. Materials and Methods
3.1. Materials
3.2. Polyphenol and Ascorbic Acid Extracts
3.3. Bioactive Component Isolation by Preparative HPLC
3.4. Mass Spectrometry
3.5. H. pylori Seed Culture
3.6. Subculture of GES-1 Cells
3.7. Toxicity Test
3.8. H. pylori-Infected Mouse Model
3.9. Identification of H. pylori
3.10. Grouping
3.11. Flow Cytometry Detection of Cell Cycle and Apoptosis
3.12. Real Time-PCR Assay for p38MAPK, BCL-2 and BAX
3.13. Western Blot
3.14. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nguyen, T.T.; Kim, S.J.; Park, J.M.; Hahm, K.B.; Lee, H.J. Repressed TGF-beta signaling through CagA-Smad3 interaction as pathogenic mechanisms of Helicobacter pylori-associated gastritis. J. Clin. Biochem. Nutr. 2015, 57, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.; Genta, R.M. Inverse Association between Helicobacter pylori Gastritis and Microscopic Colitis. Inflamm. Bowel. Dis. 2016, 22, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Gao, Q.Y.; Fang, J.Y. Meta-analysis of the efficacy and safety of Lactobacillus-containing and Bifidobacterium-containing probiotic compound preparation in Helicobacter pylori eradication therapy. J. Clin. Gastroenterol. 2013, 47, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Omata, F.; Shimbo, T.; Ohde, S.; Deshpande, G.A.; Fukui, T. Cost-Effectiveness Analysis of Helicobacter pylori Diagnostic Methods in Patients with Atrophic Gastritis. Gastroenterol. Res. Pract. 2017, 2017, 2453254. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.C.; Tsai, Y.C.; Yang, H.B.; Yeh, Y.C.; Chang, W.L.; Kuo, H.Y.; Lu, C.C.; Sheu, B.S. The corpus-predominant gastritis index can be an early and reversible marker to identify the gastric cancer risk of Helicobacter pylori-infected nonulcer dyspepsia. Helicobacter 2017. [Google Scholar] [CrossRef] [PubMed]
- Chmiela, M.; Karwowska, Z.; Gonciarz, W.; Allushi, B.; Staczek, P. Host pathogen interactions in Helicobacter pylori related gastric cancer. World J. Gastroenterol. 2017, 23, 1521–1540. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, S.; Nunn, L. Helicobacter pylori and gastric cancer: A state of the art review. Gastroenterol. Hepatol. Bed Bench 2015, 8, S6–S14. [Google Scholar] [PubMed]
- Burkitt, M.D.; Duckworth, C.A.; Williams, J.M.; Pritchard, D.M. Helicobacter pylori-induced gastric pathology: Insights from in vivo and ex vivo models. Dis. Model. Mech. 2017, 10, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Moriya, K.; Tamura, H.; Nakamura, K.; Hosone, M.; Inokuchi, K. A primary esophageal MALT lymphoma patient with Helicobacter pylori infection achieved complete remission after H. pylori eradication without anti-lymphoma treatment. Leuk. Res. Rep. 2017, 7, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Cekin, A.H.; Sahinturk, Y.; Akbay Harmandar, F.; Uyar, S.; Yolcular, B.O.; Cekin, Y. Use of probiotics as an adjuvant to sequential H. pylori eradication therapy: Impact on eradication rates, treatment resistance, treatment-related side effects, and patient compliance. Turk. J. Gastroenterol. 2017, 28, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.B.; Lee, K.L.; Kim, J.S.; Lee, J.K.; Kim, W.; Jung, Y.J.; Jeong, J.B.; Kim, J.W.; Kim, B.G. Antibiotics-associated diarrhea and other gastrointestinal abnormal responses regarding Helicobacter pylori eradication. Korean J. Gastroenterol. 2010, 56, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, E.; Montes, M.; Fernandez-Reyes, M.; Lizasoain, J.; Ibarra, B.; Mendarte, U.; Zapata, E.; Mendiola, J.; Perez-Trallero, E. Clarithromycin resistance in Helicobacter pylori and its molecular determinants in northern Spain, 2013–2015. J. Glob. Antimicrob. Resist. 2017, 9, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, K.F.; Valdivieso, M.; Bussalleu, A.; Sexton, R.; Thompson, K.C.; Osorio, S.; Reyes, I.N.; Crowley, J.J.; Baker, L.H.; Xi, C. Antibiotic resistance among Helicobacter pylori clinical isolates in Lima, Peru. Infect. Drug Resist. 2017, 10, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Yang, S.; Wang, X.; Chen, T.; Wang, X.; Tang, X.; Zhang, R.; Shen, L. Reclamation of Chinese herb residues using probiotics and evaluation of their beneficial effect on pathogen infection. J. Infect. Public Health 2017, in press. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Li, P.; Liu, X.; Luo, L. Chinese herb medicine against Sortase A catalyzed transformations, a key role in gram-positive bacterial infection progress. J. Enzym. Inhib. Med. Chem. 2016, 31, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, K.; Watanabe, T.; Tanigawa, T.; Tominaga, K.; Fujiwara, Y.; Arakawa, T. Sofalcone, a gastroprotective drug, promotes gastric ulcer healing following eradication therapy for Helicobacter pylori: A randomized controlled comparative trial with cimetidine, an H2-receptor antagonist. J. Gastroenterol. Hepatol. 2010, 25, S155–S160. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, K.; Tanigawa, T.; Hamaguchi, M.; Takashima, T.; Sasaki, E.; Shiba, M.; Tominaga, K.; Fujiwara, Y.; Oshitani, N.; Matsumoto, T.; et al. Comparison of the effects of rebamipide with those of cimetidine on chronic gastritis associated with Helicobacter pylori in Mongolian gerbils. Aliment. Pharmacol. Ther. 2003, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Previti, F.W.; Holt, R.W. Failure of cimetidine prophylaxis in the critically ill patient: Two fatal cases of duodenal perforation. J. Med. Soc. N. J. 1980, 77, 827–828. [Google Scholar] [PubMed]
- Zhang, K.; Zhang, J.; Wei, S.; Jing, W.; Wang, Y.; Liu, A. Development and validation of HPLC coupled with triple quadrupole MS for the simultaneous determination of six phenolic acids, six flavonoids, and a lignan in Polygonum capitatum. J. Sep. Sci. 2013, 36, 2407–2413. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yu, M.; Meng, D.; Li, Z.; Zhang, L. A new chromone glycoside from Polygonum capitatum. Fitoterapia 2007, 78, 506–509. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.G.; Zhang, L.J.; Sun, F.; Zhang, J.J.; Chen, A.Y.; Lan, Y.Y.; Li, Y.J.; Wang, A.M.; He, X.; Xiong, Y.; et al. Antibacterial and anti-inflammatory effects of extracts and fractions from Polygonum capitatum. J. Ethnopharmacol. 2011, 134, 1006–1009. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Fu, J.; Yin, X.; Li, X.; Wang, B.; Cao, S.; Zhang, J.; Zhang, H.; Zhao, Y.; Ni, J. Pharmacological and other Bioactivities of the Genus Polygonum—A Review. Trop. J. Pharm. Res. 2014, 13, 1749–1759. [Google Scholar]
- Liu, Z.J.; Qi, J.; Zhu, D.N.; Yu, B.Y. Chemical constituents from Polygonum capitatum and their antioxidation activities in vitro. Zhong Yao Cai 2008, 31, 995–998. [Google Scholar] [PubMed]
- Wang, H.; Sun, Y.; Liu, S.; Yu, H.; Li, W.; Zeng, J.; Chen, C.; Jia, J. Upregulation of progranulin by Helicobacter pylori in human gastric epithelial cells via p38MAPK and MEK1/2 signaling pathway: Role in epithelial cell proliferation and migration. FEMS Immunol. Med. Microbiol. 2011, 63, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Ryszczuk, E.; Roszko-Kirpsza, I.; Guzinska-Ustymowicz, K.; Olejnik, B.J.; Kaczmarski, M.G.; Maciorkowska, E. EGFR and Bcl-2 in gastric mucosa of children infected with Helicobacter pylori. Postepy Hig. Med. Dosw. 2016, 70, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Bartchewsky, W., Jr.; Martini, M.R.; Squassoni, A.C.; Alvarez, M.C.; Ladeira, M.S.; Salvatore, D.M.; Trevisan, M.A.; Pedrazzoli, J., Jr.; Ribeiro, M.L. Effects of Helicobacter pylori infection on the expressions of Bax and Bcl-2 in patients with chronic gastritis and gastric cancer. Dig. Dis. Sci. 2010, 55, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Sun, H.-Y.; Qin, X.-L.; Li, Y.-J.; Liao, S.-G.; Gong, Z.-P.; Lu, Y.; Wang, Y.-L.; Wang, A.-M.; Lan, Y.-Y. A UPLC-MS/MS Method for Simultaneous Determination of Free and Total Forms of a Phenolic Acid and Two Flavonoids in Rat Plasma and Its Application to Comparative Pharmacokinetic Studies of Polygonum capitatum Extract in Rats. Molecules 2017, 22, 353. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Gogal, R.M.; Walsh, J.E. A new rapid and simple non-radioactive assay to monitor and determine the proliferation of lymphocytes: An alternative to [3H] thymidine incorporation assay. J. Immunol. Methods 1994, 170, 211–224. [Google Scholar] [CrossRef]
- Naumann, M.; Crabtree, J.E. Helicobacter pylori-induced epithelial cell signalling in gastric carcinogenesis. Trends. Microbiol. 2004, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Boyanova, L.; Evstatiev, I.; Gergova, G.; Yaneva, P.; Mitov, I. Linezolid susceptibility in Helicobacter pylori, including strains with multidrug resistance. Int. J. Antimicrob. Agents 2015, 46, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Ghotaslou, R.; Leylabadlo, H.E.; Asl, Y.M. Prevalence of antibiotic resistance in Helicobacter pylori: A recent literature review. World J. Methodol. 2015, 5, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.W.; Jia, M.; Salchow, R.; Kentsch, M.; Cui, X.J.; Deng, H.Y.; Sun, Z.J.; Kluwe, L. Efficacy and side effects of chinese herbal medicine for menopausal symptoms: A critical review. Evid. Based Complement. Altern. Med. 2012, 2012, 568106. [Google Scholar] [CrossRef] [PubMed]
- Ran, Z.H.; Liu, J.; Feng, Y.; Zou, J.; Xiao, S.D. Investigation of the sensitivities of distinct gastric cancer cells to parvovirus H-1 induced cytotoxicity. Chin. J. Dig. Dis. 2004, 5, 93–97. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, K.; Keiner, B.; Zhou, J.; Czudai, V.; Li, T.; Chen, Z.; Liu, J.; Klenk, H.D.; Shu, Y.L.; Sun, B. Influenza A virus replication induces cell cycle arrest in G0/G1 phase. J. Virol. 2010, 84, 12832–12840. [Google Scholar] [CrossRef] [PubMed]
- Belair, C.; Baud, J.; Chabas, S.; Sharma, C.M.; Vogel, J.; Staedel, C.; Darfeuille, F. Helicobacter pylori interferes with an embryonic stem cell micro RNA cluster to block cell cycle progression. Silence 2011, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Saito, K.; Reed, J.C. Structure-function analysis of the Bcl-2 oncoprotein. Addition of a heterologous transmembrane domain to portions of the Bcl-2 beta protein restores function as a regulator of cell survival. J. Biol. Chem. 1993, 268, 10920–10926. [Google Scholar] [PubMed]
- Chang, H.; Chen, D.; Ni, B.; Zuo, Q.; Wang, C.; Han, R.; Lan, C. Cortactin Mediates Apoptosis of Gastric Epithelial Cells Induced by VacA Protein of Helicobacter pylori. Dig. Dis. Sci. 2016, 61, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhu, H.; Lin, Z.; Lin, G.; Lv, G. Compound 13, an alpha1-selective small molecule activator of AMPK, inhibits Helicobacter pylori-induced oxidative stresses and gastric epithelial cell apoptosis. Biochem. Biophys. Res. Commun. 2015, 463, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Leri, A.; Claudio, P.P.; Li, Q.; Wang, X.; Reiss, K.; Wang, S.; Malhotra, A.; Kajstura, J.; Anversa, P. Stretch-mediated release of angiotensin II induces myocyte apoptosis by activating p53 that enhances the local renin-angiotensin system and decreases the Bcl-2-to-Bax protein ratio in the cell. J. Clin. Investig. 1998, 101, 1326. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y. Antioxidant activity of extract from Polygonum aviculare L. Biol. Res. 2006, 39, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.P.; Hu, L.J.; Li, X.L.; Xiao, H.; Tao, D.D.; Hu, J.B.; Gong, J.P. Preparation of suspension of gastric mucous membrane single cell and expression of cyclins in cells. Zhonghua Wei Chang Wai Ke Za Zhi 2008, 11, 253–255. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds gallic acid, quercitrin and quercetin are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | 24 h |
|---|---|
| CG | 150.33 ± 43.13 * |
| MG | 1200.25 ± 16.43 ★ |
| QG | 655.12 ± 31.45 ★,* |
| Groups | Concentration of Quercetin | 24 h | 48 h | 72 h |
|---|---|---|---|---|
| H. pylori-infected mice | 0 μg/mL | 1200.25 ± 16.43 | 1153.26 ± 30.25 | 1103.14 ± 21.32 |
| 8 μg/mL | 1099.42 ± 21.34 * | 1047.08 ± 38.19 * | 990.13 ± 19.63 * | |
| 16 μg/mL | 959.47 ± 23.16 * | 900.92 ± 26.02 * | 860.08 ± 21.36 * | |
| 32 μg/mL | 830.36 ± 32.17 * | 787.92 ± 27.32 * | 743.25 ± 12.50 * | |
| 64 μg/mL | 760.34 ± 21.65 * | 656.58 ± 13.96 * | 590.63 ± 19.75 * | |
| Uninfected mice | 0 μg/mL | 150.33 ± 43.13 | 142.39 ± 37.44 | 160.26 ± 30.18 |
| 8 μg/mL | 149.32 ± 32.17 | 152.32 ± 39.76 | 140.64 ± 31.73 | |
| 16 μg/mL | 139.56 ± 21.87 * | 134.72 ± 25.98 * | 136.45 ± 23.76 * | |
| 32 μg/mL | 130.12 ± 21.35 * | 138.43 ± 21.79 | 132.46 ± 18.73 * | |
| 64 μg/mL | 121.17 ± 15.88 * | 126.34 ± 16.24 * | 128.42 ± 18.37 * |
| Groups | DipG0–G1% | DipG2-M% | DipS% |
|---|---|---|---|
| CG | 29.48 ± 2.18 * | 0 * | 70.52 ± 3.31 * |
| MG | 50.35 ± 1.38 ★ | 4.82 ± 0.40 ★ | 44.83 ± 3.02 ★ |
| PCG | 31.07 ± 1.60 * | 2.14 ± 0.72 * | 66.79 ± 0.38 * |
| wt % | |
|---|---|
| Casein | 23.1 ± 2.1 |
| Corn starch | 43.9 ± 4.1 |
| Sucrose | 5.1 ± 0.5 |
| Maltodextrin | 5.3 ± 0.5 |
| Soy oil | 5.1 ± 0.4 |
| Palm oil | – |
| Lard | – |
| Cellulose | 5.1 ± 0.6 |
| Mineral mixture | 6.8 ± 0.7 |
| Vitamin mixture | 1.3 ± 0.1 |
| Kj % | |
| Protein | 23.2 ± 2.3 |
| Fat | 12.5 ± 1.2 |
| Carbohydrates | 65.3 ± 3.4 |
| Energy content [kJ/g] | 15.3 ± 1.6 |
| Mean sterol contents, mg/100 g | |
| Cholesterol | 0.52 ± 0.08 |
| Campesterol | 1.21 ± 0.16 |
| Stigmasterol | 2.13 ± 0.31 |
| Sitosterol | 3.04 ± 0.52 |
| ∆5-Avenasterol | 0.18±0.02 |
| Sitostanol | 1.30 ± 0.32 |
| 24-Methylene cycloartenol | 0.31 ± 0.04 |
| Cycloartenol | Not detectable |
| Campestanol | 0.41 ± 0.10 |
| Total sterols | 8.50 ± 0.79 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Huang, J.; Xie, X.; He, Y.; Mo, F.; Luo, Z. Quercetin from Polygonum capitatum Protects against Gastric Inflammation and Apoptosis Associated with Helicobacter pylori Infection by Affecting the Levels of p38MAPK, BCL-2 and BAX. Molecules 2017, 22, 744. https://doi.org/10.3390/molecules22050744
Zhang S, Huang J, Xie X, He Y, Mo F, Luo Z. Quercetin from Polygonum capitatum Protects against Gastric Inflammation and Apoptosis Associated with Helicobacter pylori Infection by Affecting the Levels of p38MAPK, BCL-2 and BAX. Molecules. 2017; 22(5):744. https://doi.org/10.3390/molecules22050744
Chicago/Turabian StyleZhang, Shu, Jian Huang, Xiaoqin Xie, Yun He, Fei Mo, and Zhaoxun Luo. 2017. "Quercetin from Polygonum capitatum Protects against Gastric Inflammation and Apoptosis Associated with Helicobacter pylori Infection by Affecting the Levels of p38MAPK, BCL-2 and BAX" Molecules 22, no. 5: 744. https://doi.org/10.3390/molecules22050744
APA StyleZhang, S., Huang, J., Xie, X., He, Y., Mo, F., & Luo, Z. (2017). Quercetin from Polygonum capitatum Protects against Gastric Inflammation and Apoptosis Associated with Helicobacter pylori Infection by Affecting the Levels of p38MAPK, BCL-2 and BAX. Molecules, 22(5), 744. https://doi.org/10.3390/molecules22050744