
Diversity Analysis and Bioresource Characterization of Halophilic Bacteria Isolated from a South African Saltpan

 ,
, 


Abstract
:1. Introduction
2. Results
2.1. Physicochemical Variables
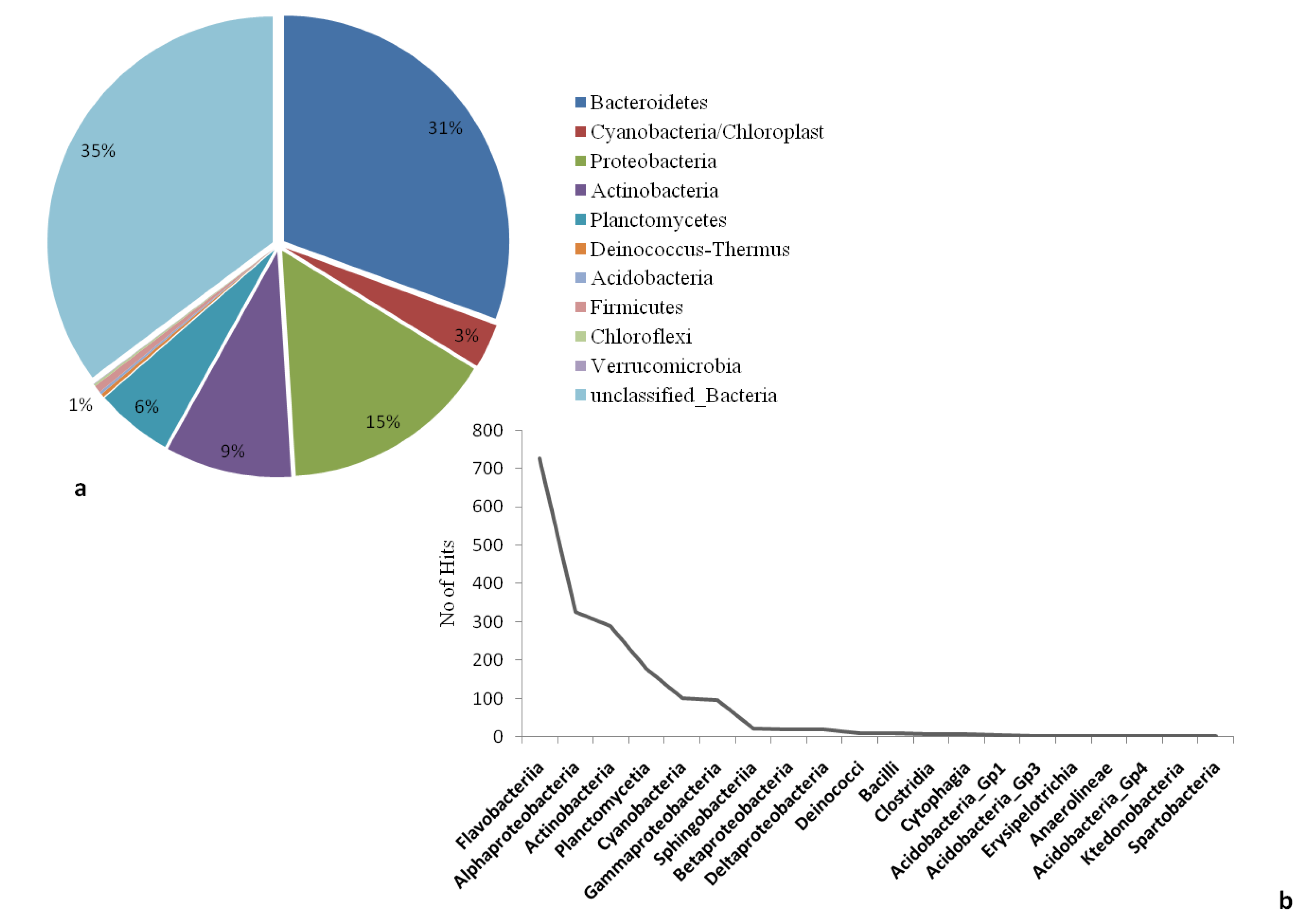
2.2. Culture Independent Analysis
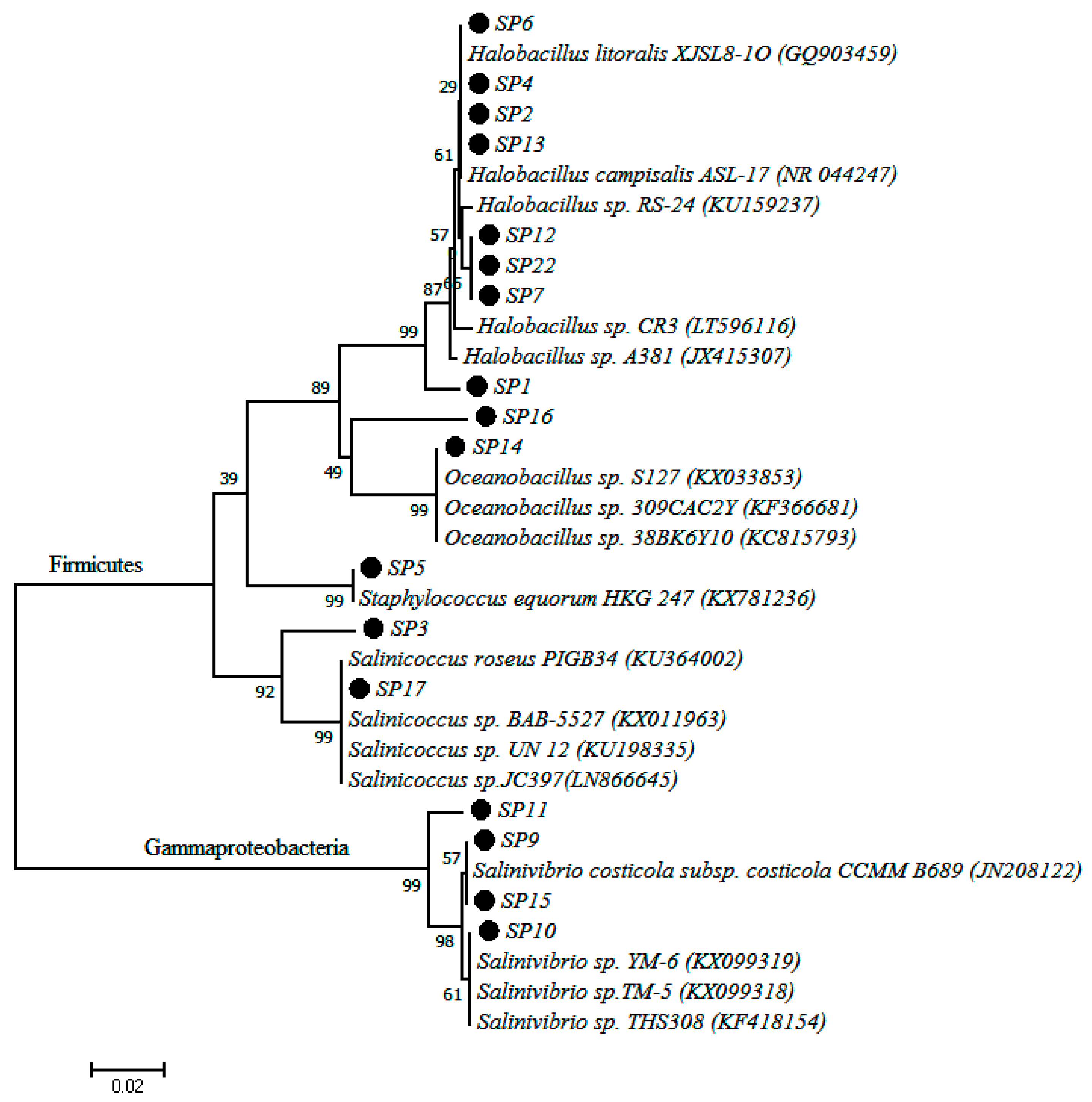
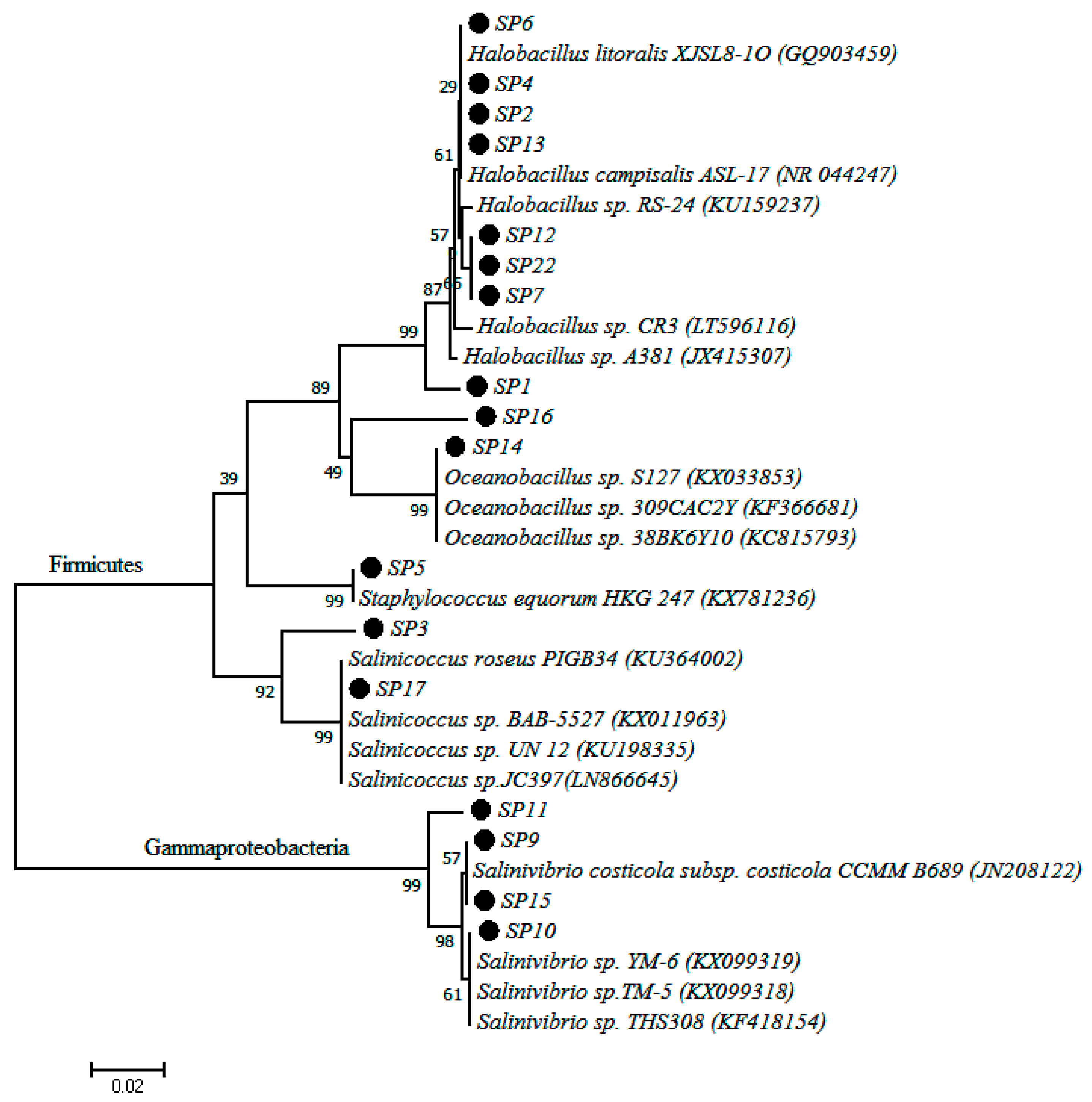
2.3. Culture Dependent Analysis
2.4. Enzyme Assay
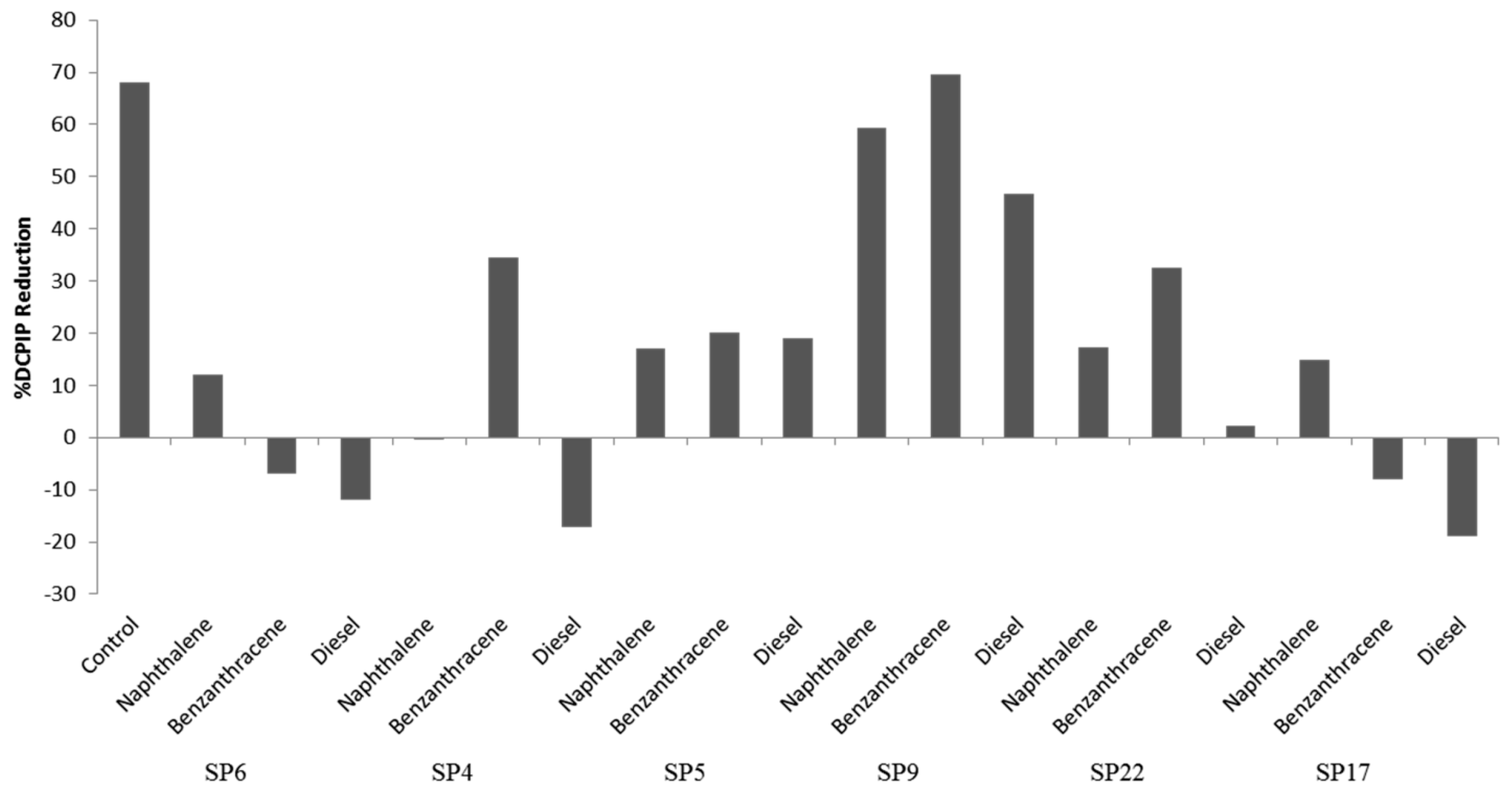
2.5. Hydrocarbon Degradation Assay
2.6. Secondary Metabolites of Halophilic Bacteria
3. Discussion
4. Materials and Methods
4.1. Description of Study Area, Sample Collection and Bacterial Isolation
4.2. DNA Extraction and Sequence Analysis
4.3. Lipase Assay
4.4. Cellulase Assay
4.5. Hydrocarbon Degradation Assay
4.6. Analysis of Bacterial Secondary Metabolites
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ramganesh, S.; Maredza, A.; Tekere, M. Microbial exploration in extreme conditions: Metagenomic analysis and future perspectives. In Metagenomics-Methods, Applications and Perspectives; Benedetti, C., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2014; Chapter 8; pp. 157–181. [Google Scholar]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Rampelotto, P.H. Extremophiles and extreme environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Azua-Bustos, A.; González-Silva, C. Biotechnological applications derived from microorganisms of the Atacama Desert. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Fenchel, T.; King, G.M.; Blackburn, T.H. Microbial biogeochemistry and extreme environments. In Bacterial Biogeochemistry, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; Chapter 8; pp. 143–161. [Google Scholar]
- Brumm, P.; Allen, L.; Xie, D.; Hermanson, S.; Hochstein, B.; Boyum, J.; Deneke, J.; Schoenfeld, T.; Gowda, K.; Mead, D. Thermophilic ellulolytic nzymes. In Advanced Products for Molecular Biology; Lucigen: Middleton, WI, USA, 2011; p. 12155. Available online: https://www.lucigen.com/docs/posters/Thermophilic-Cellulolytic-Enzymes-Poster-Thermophiles-2011.pdf (accessed on 19 April 2017).
- Bertoldo, C.; Antranikian, G. Starch-Hydrolyzing enzymes from thermophilic archaea and bacteria. Curr. Opin. Chem. Biol. 2002, 6, 151–160. [Google Scholar] [CrossRef]
- Zuridah, H.; Norazwin, N.; Siti Aisyah, M.; Fakhruzzaman, M.N.A.; Zeenathul, N.A. Identification of lipase producing thermophilic bacteria from Malaysian hot springs. Afr. J. Microbiol. Res. 2011, 5, 3569–3573. [Google Scholar] [CrossRef]
- Dönmez, G. Microbial oil production from thermophile cyanobacteria for biodiesel production. Appl. Energy 2011, 88, 3632–3635. [Google Scholar]
- De Lourdes Moreno, M.; Pérez, D.; García, M.T.; Mellado, E. Halophilic bacteria as a source of novel hydrolytic enzymes. Life 2013, 3, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Waditee-Sirisattha, R.; Kageyama, H.; Takabe, T. Halophilic microorganism resources and their applications in industrial and environmental biotechnology. AIMS Microbiol. 2016, 2, 42–54. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P. Halophiles and their enzymes: Negativity put to good use. Curr. Opin. Microbiol. 2015, 25, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Chen, J.C.; Wu, Q.; Chen, G.Q. Halophiles, coming stars for industrial biotechnology. Biotechnol. Adv. 2015, 33, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Birbir, M.; Ogan, A.; Calli, B.; Mertoglu, B. Enzyme characteristics of extremely halophilic archaeal community in Tuzkoy Salt Mine, Turkey. World J. Microbiol. Biotechnol. 2004, 20, 613–621. [Google Scholar] [CrossRef]
- Rohban, R.; Amoozegar, M.A.; Ventosa, A. Screening and isolation of halophilic bacteria producing extracellular hydrolyses from Howz Soltan Lake, Iran. J. Ind. Microbiol. Biotechnol. 2009, 36, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Porro, C.; Martín, S.; Mellado, E.; Ventosa, A. Diversity of moderately halophilic bacteria producing extracellular hydrolytic enzymes. J. Appl. Microbiol. 2003, 94, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Ara, I.; Daram, D.; Baljinova, T.; Yamamura, H.; Bakir, M.A.; Suto, M.; Ando, K. Isolation, classification, phylogenetic analysis and scanning electron microscopy of halophilic, halotolerant and alkaliphilic actinomycetes isolated from hypersaline soil. Afr. J. Microbiol. Res. 2013, 7, 298–308. [Google Scholar]
- Hamdache, A.; Lamarti, A.; Aleu, J.; Collado, I.G. Non-peptide metabolites from the genus bacillus. J. Nat. Prod. 2011, 74, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Younis, K.M.; Usup, G.; Ahmad, A. Secondary metabolites produced by marine streptomyces as antibiofilm and quorum-sensing inhibitor of uropathogen Proteus mirabilis. Environ. Sci. Pollut. Res. Int. 2016, 23, 4756–4767. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Hewavitharana, A.K.; Ng, Y.K.; Shaw, P.N.; Fuerst, J.A.; odson, M.P. LC-MS-Based metabolomics study of marine bacterial secondary metabolite and antibiotic production in salinispora arenicola. Mar. Drugs 2015, 13, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Temperton, B.; Giovannoni, S.J. Metagenomics: Microbial diversity through a scratched lens. Curr. Opin. Microbiol. 2012, 15, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Pace, N.R. A molecular view of microbial diversity and the biosphere. Science 1997, 276, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sharma, P.; Dev, K.; Srivastava, M.; Sourirajan, A. A diverse group of halophilic bacteria exist in Lunsu, a natural salt water body of Himachal Pradesh, India. SpringerPlus 2015, 4, 274. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.T.; Ashton, P.J.; Williams, W.D. Inland salt waters of southern Africa. Hydrobiologia 1991, 210, 75–91. [Google Scholar] [CrossRef]
- Casanueva, A.; Galada, N.; Baker, G.C.; Grant, W.D.; Heaphy, S.; Jones, B.; Cowan, D.A. Nanoarchaeal 16S rRNA gene sequences are widely dispersed in hyperthermophilic and mesophilic halophilic environments. Extremophiles 2008, 12, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, A.N.; Cowan, D.; Porter, D.; Valverde, A. Dissimilatory sulphate reduction in hypersaline coastal pans: An integrated microbiological and geochemical study. Geobiology 2013, 11, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9–18. [Google Scholar]
- Köpke, M.; Mihalcea, C.; Liew, F.; Tizard, J.H.; Ali, M.S.; Conolly, J.J.; Simpson, S.D. 2,3-Butanediol production by acetogenic bacteria, an alternative route to chemical synthesis, using industrial waste gas. Appl. Environ. Microbiol. 2011, 77, 5467–5475. [Google Scholar] [CrossRef] [PubMed]
- Soltys, K.A.; Batta, A.K.; Koneru, B. Successful nonfreezing, subzero preservation of rat liver with 2,3-butanediol and type I antifreeze protein. J. Surg. Res. 2001, 96, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Celińska, E.; Grajek, W. Biotechnological production of 2,3-butanediol-Current state and prospects. Biotechnol. Adv. 2009, 27, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.J.; Akama, T.; Zhang, Y.K.; Sauro, V.; Pandit, C.; Singh, R.; Maples, K.R. Identification of a novel boron-containing antibacterial agent (AN0128) with anti-inflammatory activity, for the potential treatment of cutaneous diseases. Bioorg. Med. Chem. Lett. 2006, 16, 5963–5967. [Google Scholar] [CrossRef] [PubMed]
- Trippier, P.C.; McGuigan, C. Boronic acids in medicinal chemistry: Anticancer, antibacterial and antiviral applications. MedChemComm 2010, 1, 183. [Google Scholar] [CrossRef]
- Mohan, G.; Thangappanpillai, A.K.; Ramasamy, B. Antimicrobial activities of secondary metabolites and phylogenetic study of sponge endosymbiotic bacteria, Bacillus sp. at Agatti Island, Lakshadweep Archipelago. Biotechnol. Rep. 2016, 11, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Rabha, A.J.; Sharma, G.D.; Naglot, A.; Gogoi, H.K. GC-MS analysis of secondary metabolites of endophytic Colletotrichum Gloeosporioides isolated from Camellia Sinensis (L) O. Kuntze. Int. J. Innov. Res. Sci. Eng. 2015, 3, 373–379. [Google Scholar]
- Antonucci, J.M.; Dickens, S.H.; Fowler, B.O.; Xu, H.H.K. Chemistry of silanes: Interfaces in dental polymers and composites. J. Res. Natl. Inst. Stand. Technol. 2005, 110, 541–558. [Google Scholar] [CrossRef] [PubMed]
- Tossavainen, A. Styrene use and occupational exposure in the plastics industry. Scand. J. Work Environ. Health 1978, 4, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Thao, H.T.B.; Yamakawa, T. Phosphite (phosphorous acid): Fungicide, fertilizer or bio-stimulator? Soil Sci. Plant Nutr. 2009, 55, 228–234. [Google Scholar] [CrossRef]
- Alcocer, M.J.C.; Dillon, P.P.; Manning, B.M.; Doyen, C.; Lee, H.A.; Daly, S.J.; Morgan, M.R.A. Use of phosphonic acid as a generic hapten in the production of broad specificity anti-organophosphate pesticide antibody. J. Agric. Food. Chem. 2000, 48, 2228–2233. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J. Methylglyoxal-A potential risk factor of manuka honey in healing of diabetic ulcers. Evid.-Based Complement. Altern. Med. 2011, 2011, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Fukushima, A.; Hayashi-Nishino, M.; Nishino, K. Effect of methylglyoxal on multidrug-resistant Pseudomonas aeruginosa. Front. Microbiol. 2014, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ismail, F.M.D.; Levitsky, D.O.; Dembitsky, V.M. Aziridine alkaloids as potential therapeutic agents. Eur. J. Med. Chem. 2009, 44, 3373–3387. [Google Scholar] [CrossRef] [PubMed]
- Roose, P. Methylamines. In Ullmann’s Encyclopedia Industrial Chemistry; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–10. [Google Scholar]
- Longo, M.A.; Sanromán, M.A. Production of food aroma compounds: Microbial and enzymatic methodologies. Food technol biotechnol production of food aroma compounds: Microbial and enzymatic methodologies. Food Technol. Biotechnol. 2006, 3, 335–353. [Google Scholar]
- Kamika, I.; Azizi, S.; Tekere, M. Microbial profiling of South African acid mine water samples using next generation sequencing platform. Appl. Microbiol. Biotechnol. 2016, 100, 6069–6079. [Google Scholar] [CrossRef] [PubMed]
- Rappé, M.S.; Giovannoni, S.J. The uncultured microbial majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.F.; Peixoto, R.S. Biodegradation of petroleum hydrocarbons in hypersaline environments. Braz. J. Microbiol. 2012, 43, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Ollivier, B.; Caumette, P.; Garcia, J.L.; Mah, R.A. Anaerobic bacteria from hypersaline environments. Microbiol. Rev. 1994, 58, 27–38. [Google Scholar] [PubMed]
- Oremland, R.S.; King, G.M. Methanogenesis in hypersaline environments. In Microbial Mats: Physiological Ecology of Benthic Microbial Communities; Cohen, Y., Rosenberg, E., Eds.; American Society for Microbiology: Washington, DC, USA, 1989; pp. 180–190. [Google Scholar]
- Ventosa, A.; Arahal, D.R. Physico-Chemical characteristics of hipersaline environments and their biodiversity. Extremophiles 2009, 2, 1–6. [Google Scholar]
- Maturrano, L.; Santos, F.; Rosselló-Mora, R.; Antón, J. Microbial diversity in Maras salterns, a hypersaline environment in the Peruvian Andes. Appl. Environ. Microbiol. 2006, 72, 3887–3895. [Google Scholar] [CrossRef] [PubMed]
- Dave, B.P.; Soni, A. Diversity of halophilic archaea at salt pans around Bhavnagar coast, Gujarat. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2013, 83, 225–232. [Google Scholar] [CrossRef]
- Stan-Lotter, H.; Fendrihan, S.; Baross, J.A.; Bains, W. Halophilic Archaea: Life with desiccation, radiation and oligotrophy over geological times. Life 2015, 5, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Fendrihan, S.; Legat, A.; Pfaffenhuemer, M.; Gruber, C.; Weidler, G.; Gerbl, F.; Stan-Lotter, H. Extremely halophilic archaea and the issue of long-term microbial survival. Life Extreme Environ. 2007, 5, 125–140. [Google Scholar]
- Simachew, A.; Lanzén, A.; Gessesse, A.; Øvreås, L. Prokaryotic community diversity along an increasing salt gradient in a soda ash concentration pond. Microb. Ecol. 2016, 71, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Nedumaran, T.; Perumal, P. Biodiversity of cyanobacteria from Uppanar estuary, south-east coast of India. Emir. J. Food Agric. 2012, 24, 248–254. [Google Scholar]
- Luo, W.; Chen, H.; Lei, A.; Lu, J.; Hu, Z. Estimating cyanobacteria community dynamics and its relationship with environmental factors. Int. J. Environ. Res. Public Health 2014, 11, 1141–1160. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. The order Halanaerobiales, and the families Halanaerobiaceae and Halobacteroidaceae. In The Prokaryotes; Rosenberg, E., DeLong, F.E., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 12, pp. 153–177. [Google Scholar]
- Vahed, S.Z.; Forouhandeh, H.; Hassanzadeh, S.; Klenk, H.-P.; Mohammad Amin Hejazi, M.S.H. Isolation and identification of halophilic bacteria from Urmia Lake in Iran. Microbioloy 2011, 80, 834–841. [Google Scholar] [CrossRef]
- Antón, J.; Oren, A.; Benlloch, S.; Rodríguez-Valera, F.; Amann, R.; Rosselló-Mora, R. Salinibacter ruber gen. nov., sp. nov., a novel, extremely halophilic member of the Bacteria from saltern crystallizer ponds. Int. J. Syst. Evol. Microbiol. 2002, 52, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Govender, L.; Naidoo, L.; Setati, M.E. Isolation of hydrolase producing bacteria from Sua pan solar salterns and the production of endo-1, 4-bbbb-xylanase from a newly isolated haloalkaliphilic Nesterenkonia sp. Afr. J. Biotechnol. 2009, 8, 5458–5466. [Google Scholar]
- Babavalian, H.; Amoozegar, M.A.; Pourbabaee, A.A.; Moghaddam, M.M.; Shakeri, F. Isolation and identification of moderately halophilic bacteria producing hydrolytic enzymes from the largest hypersaline playa in Iran. Microbiology 2013, 82, 466–474. [Google Scholar] [CrossRef]
- Wang, C.Y.; Hsieh, Y.R.; Ng, C.C.; Chan, H.; Lin, H.T.; Tzeng, W.S.; Shyu, Y.T. Purification and characterization of a novel halostable cellulase from Salinivibrio sp. strain NTU-05. Enzyme Microb. Technol. 2009, 44, 373–379. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Rasoul-Amini, S.; Kazemi, A.; Zarrinic, G.; Morowvat, M.H.; Kargar, M. Isolation and characterization of some moderately halophilic bacteria with lipase activity. Microbiology 2011, 80, 483–487. [Google Scholar] [CrossRef]
- Andualema, B.; Gessesse, A. Microbial lipase and their industrial applications: Review. Biotechnology 2012, 11, 100–118. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Potential of halotolerant and halophilic microorganisms for biotechnology. Extremophiles 2001, 5, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Fathepure, B.Z. Recent studies in microbial degradation of petroleum hydrocarbons in hypersaline environments. Front. Microbiol. 2014, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bonfá, M.R.L.; Grossman, M.J.; Mellado, E.; Durrant, L.R. Biodegradation of aromatic hydrocarbons by Haloarchaea and their use for the reduction of the chemical oxygen demand of hypersaline petroleum produced water. Chemosphere 2011, 84, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Koma, D.; Matsumiya, Y.; Chung, S.Y.; Kubo, M. Phylogenetic analysis of long-chain hydrocarbon-degrading bacteria and evaluation of their hydrocarbon-degradation by the 2,6-DCPIP assay. Biodegradation 2008, 19, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kunihiro, M.; Ozeki, Y.; Nogi, Y.; Hamamura, N.; Kanaly, R.A. Benz[a]anthracene biotransformation and production of ring fission products by Sphingobium sp. strain KK22. Appl. Environ. Microbiol. 2013, 79, 4410–4420. [Google Scholar] [CrossRef] [PubMed]
- Mujahid, T.Y.; Wahab, A.; Padhiar, S.H.; Subhan, S.A.; Baloch, M.N.; Pirzada, Z.A. Isolation and characterization of hydrocarbon degrading bacteria from petrol contaminated soil. Aust. J. Basic Appl. Sci. 2015, 11, 223–231. [Google Scholar]
- Pacher, P.; Nivorozhkin, A.; Szabó, C. Therapeutic effects of xanthine oxidase inhibitors: Renaissance half a century after the discovery of allopurinol. Pharmacol. Rev. 2006, 58, 87–114. [Google Scholar] [CrossRef] [PubMed]
- Bordone, M.P.; Lanzani, M.F.; Lopez-Costa, J.J.; Chianelli, M.S.; Franco, P.; Saenz, D.A.; Rosenstein, R.E. Bacterial lipopolysaccharide protects the retina from light-induced damagae. J. Neurochem. 2012, 122, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Donald, G.; Hertzer, K.; Eibl, G. Baicalein-an intriguing therapeutic phytochemical in pancreatic cancer. Curr. Drug Targets 2012, 13, 1772–1776. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, G.; Bharti, R.; Jha, P.K.; Selvakumar, M.; Dey, G.; Jha, R.; Sivaramakrishnan, S. Dietary flavone chrysin (5,7-dihydroxyflavone ChR) functionalized highly-stable metal nanoformulations for improved anticancer applications. RSC Adv. 2015, 5, 89869–89878. [Google Scholar] [CrossRef]
- Wickrama Arachchilage, A.P.; Wang, F.; Feyer, V.; Plekan, O.; Prince, K.C. Photoelectron spectra and structures of three cyclic dipeptides: PhePhe, tyrPro, and hisGly. J. Chem. Phys. 2012, 136, 124301–124307. [Google Scholar] [CrossRef] [PubMed]
- Ström, K.; Sjögren, J.; Broberg, A.; Schnürer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides Cyclo(l-Phe-l-Pro) and Cyclo(l-Phe-trans-4-OH-l-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Bin Jantan, I.; Karim Moharam, B.A.; Santhanam, J.; Jamal, J.A. Correlation between chemical composition and antifungal activity of the essential oils of eight Cinnamomum. species. Pharm. Biol. 2008, 46, 406–412. [Google Scholar] [CrossRef]
- Day, J.A. The major ion chemistry of some southern African saline systems. Hydrobiologia 1993, 267, 37–59. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Native Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, L.; Nagar, S.; Raina, C.; Parshad, R.; Gupta, V.K. Screening, isolation and production of lipase/esterase producing Bacillus sp. strain DVL2 and its potential evaluation in esterification and resolution reactions. Arch. Appl. Sci. Res. 2012, 4, 1763–1770. [Google Scholar]
- Abd-elhakeem, M.A.; Elsayed, A.M.; Alkhulaqi, T.A. New colorimetric method for lipases activity assay in microbial media. Am. J. Anal. Chem. 2013, 4, 442–444. [Google Scholar] [CrossRef]
- Worthington, C.E. Worthington Enzyme Manual; Worthington Biochemical Corporation: Freehold, NJ, USA, 1988; Available online: http://openwetware.org/images/5/5a/Cellulase_Assay.pdf (accessed on 19 April 2017).
- Um, Y.; Chang, M.W.; Holoman, T.P. A simple and effective plating method to screen polycyclic aromatic hydrocarbon-degrading bacteria under various redox conditions. Appl. Microbiol. Biotechnol. 2010, 88, 291–297. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, N.C.; Rodrigues, A.A.; Alves, M.I.R.; Filho, N.R.A.; Sadoyama, G.; Vieira, J.D.G. Endophytic bacteria with potential for bioremediation of petroleum hydrocarbons and derivatives. Afr. J. Biotechnol. 2012, 11, 2977–2984. [Google Scholar]
Sample Availability: Not available. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Units | Saltpan (Soutpan) | |
|---|---|---|---|
| Physicochemical | Temperature | °C | 26.9 |
| pH | - | 8.84 | |
| Dissolved oxygen | mg/L | 1.58 | |
| Conductivity | S/m | 15.96 | |
| Salinity | % | 12.8 | |
| TDS | g/L | 102.6 | |
| ORP | mV | 96.5 | |
| Majority ions Nutrients | Total Iron | mg/L | 1.51 |
| Silicon | mg/L | 5.35 | |
| Calcium | mg/L | 1028 | |
| Potassium | mg/L | 210 | |
| Magnesium | mg/L | 206 | |
| Sodium | g/L | 42.6 | |
| Chloride | g/L | 75.8 | |
| Trace Elements | Silver | mg/L | Trace |
| Aluminum | mg/L | Trace | |
| Arsenic | mg/L | Trace | |
| Boron | mg/L | 0.22 | |
| Barium | mg/L | Trace | |
| Beryllium | mg/L | Trace | |
| Bismuth | mg/L | Trace | |
| Molybdenum | mg/L | Trace | |
| Nickel | mg/L | Trace | |
| Lead | mg/L | Trace | |
| Selenium | mg/L | 0.58 | |
| Strontium | mg/L | 31.0 | |
| Lead | mg/L | Trace | |
| Tellurium | mg/L | Trace | |
| Vanadium | mg/L | Trace | |
| Zinc | mg/L | Trace | |
| Other Parameters | Total Sulphur | mg/L | 1925 |
| Isolate Code | Sequence Length (nt) | Closest Similarity | % Similarity | Accession Number |
|---|---|---|---|---|
| SP1 | 859 | Thalassobacillusdevorans strain HME8790 | 99 | KX885465 |
| SP2 | 888 | Halobacillus sp. K13 | 99 | KX885460 |
| SP3 | 744 | Salinicoccushispanicus strain muz4C4 | 99 | KX885467 |
| SP4 | 972 | Halobacillus sp. K13 | 99 | KX885456 |
| SP5 | 768 | Staphylococcus equorum NIIST B-509 | 99 | KX885459 |
| SP6 | 879 | Halobacillus sp. A381 | 99 | KX885454 |
| SP7 | 929 | Halobacillusalkaliphilus strain MGR92 | 100 | KX885457 |
| SP8 | 876 | Halobacillusalkaliphilus strain MGR92 | 100 | KX885463 |
| SP9 | 935 | Salinivibrio sp. 89Y | 100 | KX885462 |
| SP10 | 860 | Salinivibrio sp. YH4 | 99 | KX885455 |
| SP11 | 612 | Salinivibriocosticola partial | 99 | KX885471 |
| SP12 | 880 | Halobacillus sp. K13 | 99 | KX885461 |
| SP13 | 831 | Halobacillus sp. A381 | 99 | KX885469 |
| SP14 | 920 | Oceanobacillus sp. 309CAC2Y12 | 100 | KX885470 |
| SP15 | 899 | Salinivibrio sp. 89Y | 100 | KX885466 |
| SP16 | 922 | Bacillus toyonensis strain L38 | 100 | KX885458 |
| SP17 | 621 | Salinicoccus sp. UN 12 | 99 | KX885468 |
| SP22 | 876 | Halobacillusalkaliphilus strain MGR92 | 100 | KX885464 |
| Isolate | Substrate | ||
|---|---|---|---|
| Hydrocarbon | CMC Salt (Cellulase) | Olive Oil (Lipase) | |
| SP1 | − | − | − |
| SP2 | − | − | − |
| SP3 | − | − | − |
| SP4 | + | − | + |
| SP5 | + | − | − |
| SP6 | + | − | − |
| SP7 | − | + | − |
| SP8 | − | − | + |
| SP9 | + | + | − |
| SP10 | − | − | − |
| SP11 | − | − | − |
| SP12 | − | − | − |
| SP13 | − | − | − |
| SP14 | − | − | − |
| SP15 | − | − | − |
| SP16 | − | − | − |
| SP17 | + | − | − |
| SP22 | + | − | + |
| Isolate | Compound Name and Chemical Structure | Potential Known Applications | References |
|---|---|---|---|
| SP7 SP9 | 2,3 Butane diol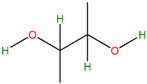 | Precursor in the manufacture of a range of chemical products. Antifreeze agent. Synthetic rubber production | [28,29,30] |
| SP8 | Borinic acid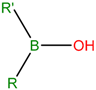 | Used in treatment of dermatological diseases including acne and atopic dermatitis | [31,32] |
| SP22 SP9 | Hexahydro-3-(2-methylpropyl)-pyrrolo[1,2a]pyrazine-1,4-dione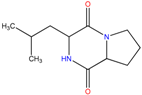 | Antibacterial, antifungal, nematicidal and anti-cancer properties. Also, commonly used as broad spectrum antibiotics | [33,34] |
| SP7 | Cyclohexyl(dimethoxy)methylsilane,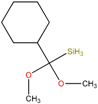 | Used to mediate interfacial bonding in mineral reinforced dental polymeric composites. | [35] |
| SP7 | Styrene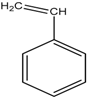 | used in the production of plastics and resins | [36] |
| SP7 | Phosphonic acid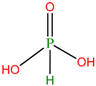 | Used as fertilizer and fungicide. Also, used in the production of broad specific antibody for pesticides | [37,38] |
| SP8 | Methylglyoxal | Used in healing diabetic ulcer and anti-bacterial activity against multi drug resistant bacteria. | [39,40] |
| SP9 | Aziridine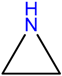 | potential therapeutic agents | [41] |
| SP9 | Dimethylamine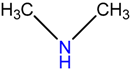 | precursor to several industrially significant compounds like DMF, CC etc. | [42] |
| SP22 SP9 | Ethyl acetate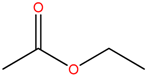 | Used as flavoring compounds in wine industry | [43] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvarajan, R.; Sibanda, T.; Tekere, M.; Nyoni, H.; Meddows-Taylor, S. Diversity Analysis and Bioresource Characterization of Halophilic Bacteria Isolated from a South African Saltpan. Molecules 2017, 22, 657. https://doi.org/10.3390/molecules22040657
Selvarajan R, Sibanda T, Tekere M, Nyoni H, Meddows-Taylor S. Diversity Analysis and Bioresource Characterization of Halophilic Bacteria Isolated from a South African Saltpan. Molecules. 2017; 22(4):657. https://doi.org/10.3390/molecules22040657
Chicago/Turabian StyleSelvarajan, Ramganesh, Timothy Sibanda, Memory Tekere, Hlengilizwe Nyoni, and Stephen Meddows-Taylor. 2017. "Diversity Analysis and Bioresource Characterization of Halophilic Bacteria Isolated from a South African Saltpan" Molecules 22, no. 4: 657. https://doi.org/10.3390/molecules22040657
APA StyleSelvarajan, R., Sibanda, T., Tekere, M., Nyoni, H., & Meddows-Taylor, S. (2017). Diversity Analysis and Bioresource Characterization of Halophilic Bacteria Isolated from a South African Saltpan. Molecules, 22(4), 657. https://doi.org/10.3390/molecules22040657