
Essential Oils of Hyptis pectinata Chemotypes: Isolation, Binary Mixtures and Acute Toxicity on Leaf-Cutting Ants


 ,
,
Abstract
:1. Introduction
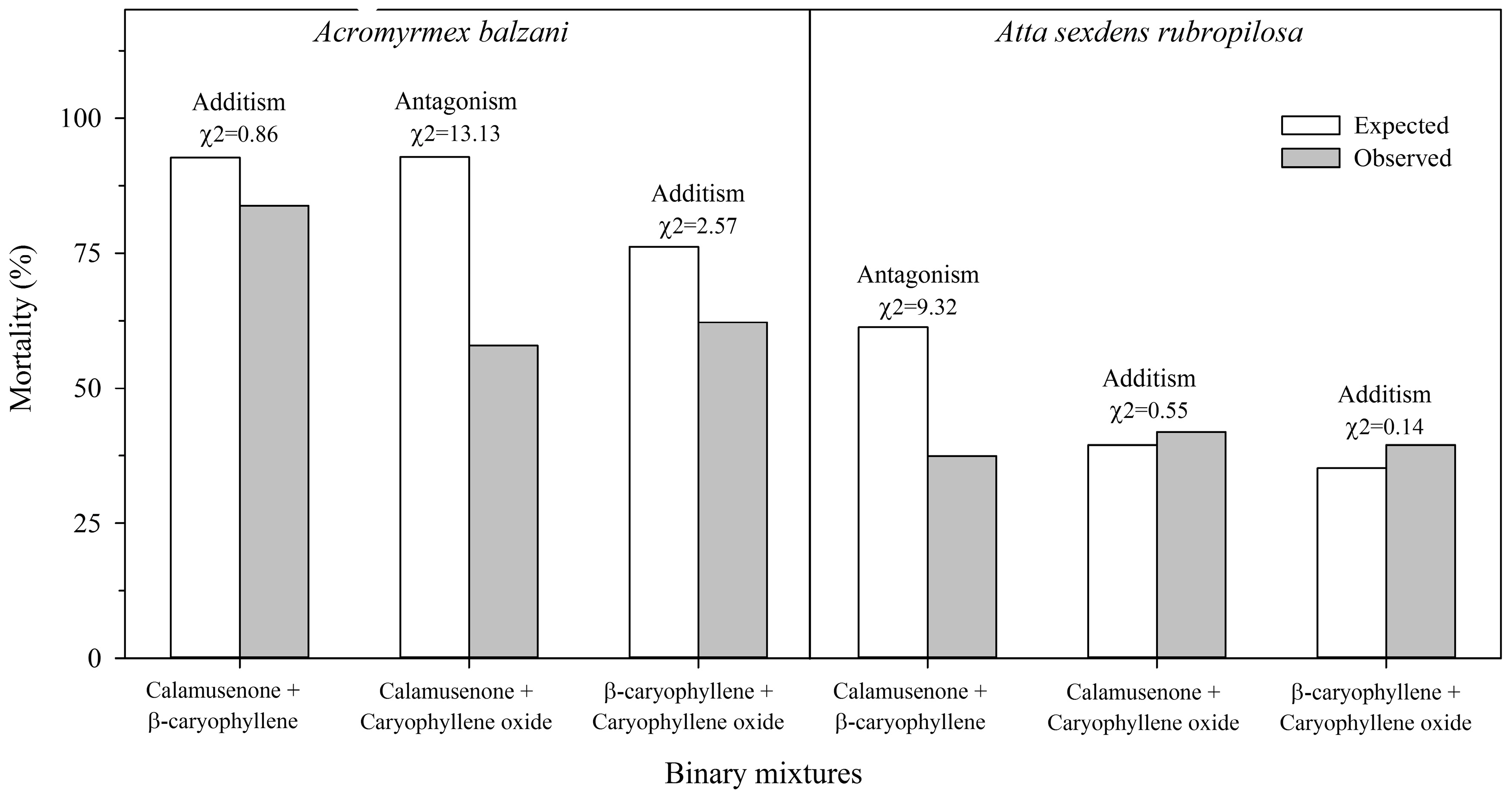
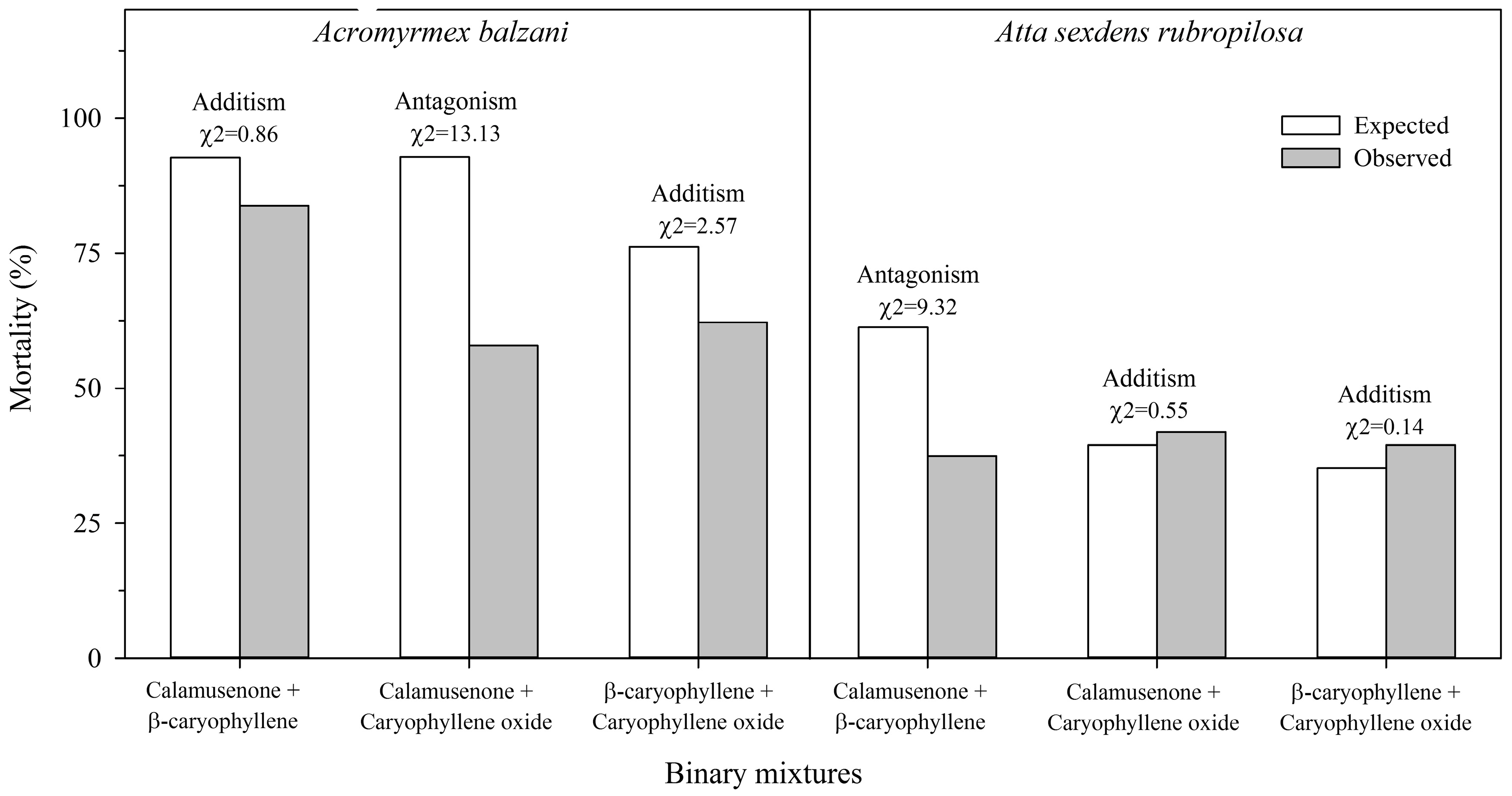
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material, Extraction and Chemical Analysis of Essential Oils
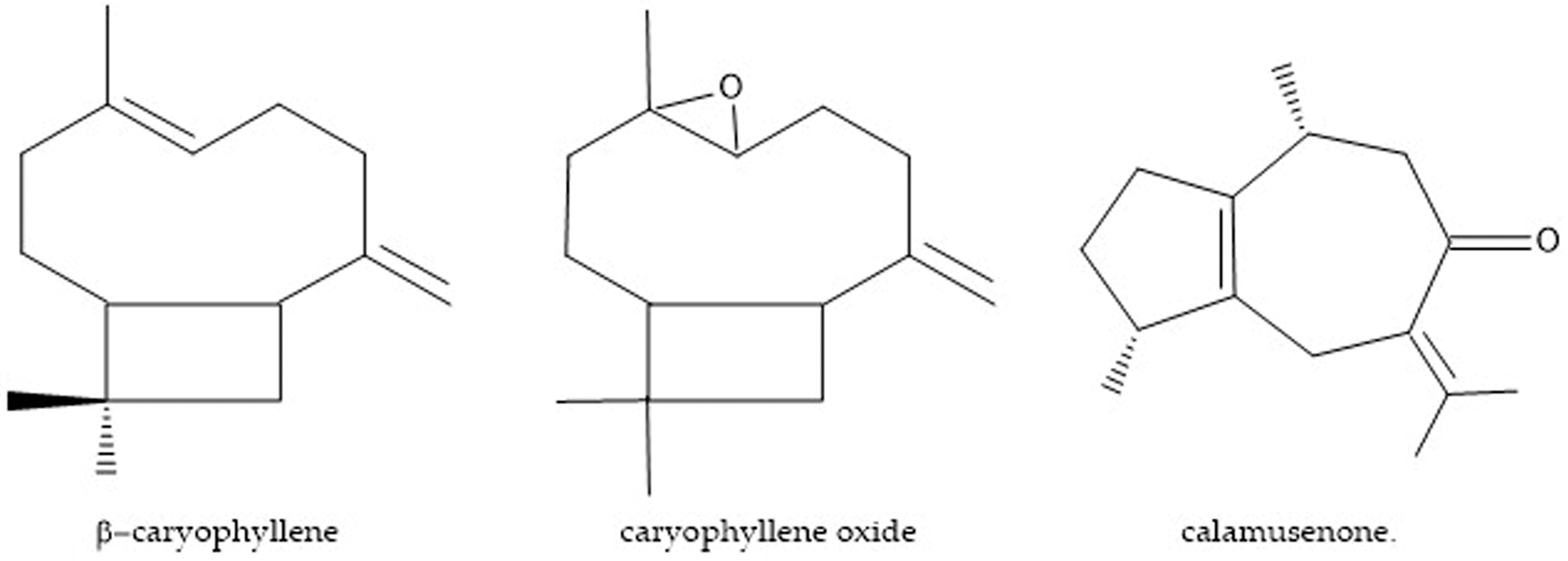
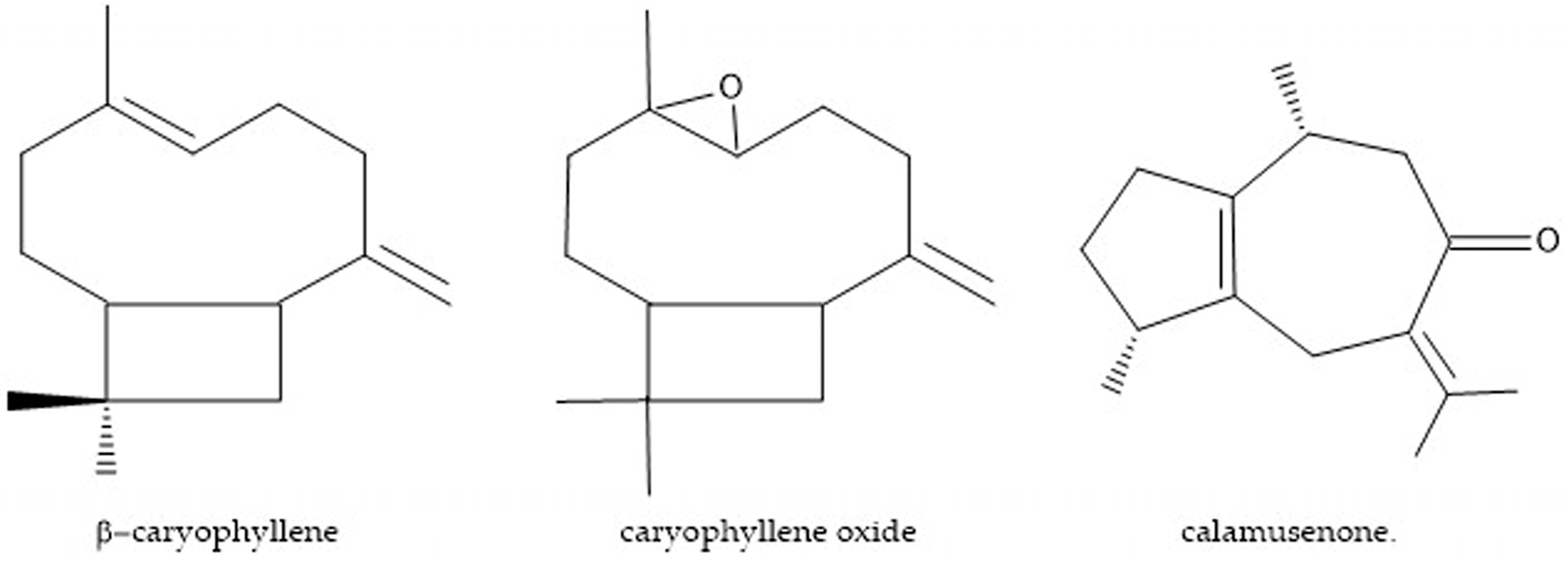
4.2. Obtainment of Major Compounds
4.3. Insects
4.4. Bioassays
4.4.1. Exposure Pathways
4.4.2. Binary Mixtures
4.5. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Della Lucia, T.M.C. Hormigas de importancia econômica en la región Neotropical. In Introducción a las Hormigas de la Región Neotropical; Fernández, F., Ed.; Instituto de Investigación de Recursos Bioloogicos Alexander Von Humboldt: Bogotá, Colombia, 2003; pp. 337–349. [Google Scholar]
- Loeck, A.E.; Grützmacher, D.D.; Coimbra, S.M. Ocorrência de formigas cortadeiras do gênero Acromyrmex nas principais regiões agropecuárias do Rio Grande do Sul. Rev. Bras. Agroc. 2003, 9, 129–133. [Google Scholar]
- Buratto, D.A.; Cardoso, J.T.; Rolim, F.A.; Filho, W.R. Avaliação dos danos causados por formigas cortadeiras do gênero Acromyrmex (Hymenoptera) aos plantios de Pinus taeda no planalto Sul-Catarinense. Floresta 2012, 42, 683–690. [Google Scholar] [CrossRef]
- Santana, D.A.Q.; Couto, L. Resistência intra-específica de eucaliptos a formigas cortadeiras. Bol. Pesqui. Florest. 1990, 20, 13–21. [Google Scholar]
- Castellani, M.A.; Forti, L.C.; Fenille, R.C.; Novcaes, Q.S. Isolation and growth of the symbiotic fungus of Atta capiguara (Hymenoptera: Formicidae). Sociobiology 2007, 50, 959–972. [Google Scholar]
- Moreira, M.D.; Picanço, M.C.; Barbosa, L.C. A. Compounds from Ageratum conyzoides: Isolation, structural elucidation and insecticide activity. Pest Manag. Sci. 2007, 63, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Kordali, S.; Yldirim, E.; Yazici, G.; Emsen, B.; Kabaagac, G.; Ercisli, S. Fumigant toxicity of essential oils of nine plant species from Asteraceae and Clusiaceae against Sitophilus granarius (L.) (Coleoptera: Curculionidae). Egypt J. Biol. Pest Control 2012, 22, 11–14. [Google Scholar]
- Jung, P.H.; Silveira, A.C.; Nieri, E.M.; Potrich, M.; Silva, E.R.L.; Refatti, M. Atividade Inseticida de Eugenia uniflora L. e Melia azedarach L. sobre Atta laevigata Smith. Floresta Ambient. 2013, 20, 191–196. [Google Scholar] [CrossRef]
- FSC-Forest Stewardship Council. Available online: https://br.fsc.org/pt-br (accessed on 9 December 2016).
- Marinho, C.G.S.; Della Lucia, T.M.C.; Guedes, R.N.C. β-Eudesmol-induced aggression in the leaf-cutting ant Atta sexdens rubropilosa. 2005, 117, 89–93. [Google Scholar] [CrossRef]
- Albuquerque, E.L.D.; Lima, J.K.A.; Souza, F.H.O.; Silva, I.M.A.; Santos, A.A.; Araújo, A.P.A.; Blank, A.F.; Lima, R.N.; Alves, P.B.; Bacci, L. Insecticidal and repellence activity of the essential oil of Pogostemon cablin against urban ants species. Acta Trop. 2013, 127, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.K.A.; Albuquerque, E.L.D.; Santos, A.C.C.; Oliveira, A.P.; Araújo, A.P.A.; Blank, A.F.; Arrigoni-Blank, M.F.; Alves, P.B.; Santos, D.A.; Bacci, L. Biotoxicity of some plant essential oils against the termite Nasutitermes corniger (Isoptera: Termitidae). Ind. Crops Prod. 2013, 47, 246–251. [Google Scholar] [CrossRef]
- Bacci, L.; Lima, J.K.A.; Araújo, A.P.A.; Blank, A.F.; Silva, I.M.A.; Santos, A.A.; Santos, A.C.C.; Alves, P.B.; Picanço, M.C. Toxicity, behavior impairment, and repellence of essential oils from pepper-rosmarin and patchouli to termites. Entomol. Exp. Appl. 2015, 156, 66–76. [Google Scholar] [CrossRef]
- Campos, R.S.; Lima, C.B.N.; Oliveira, A.P.; Albano, A.P.A.; Blank, A.F.; Alves, P.B.; Nascimento, L.R.; Araújo, V.A.; Santana, A.S.; Bacci, L. Acaricidal properties of vetiver essential oil from Chrysopogon zizanioides (Poaceae) against the tick species Amblyomma cajennense and Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Vet. Parasitol. 2015, 212, 1–7. [Google Scholar]
- Peixoto, M.G.; Bacci, L.; Blank, A.F.; Araújo, A.P.A.; Alves, P.B.; Silva, J.H.S.; Santos, A.A.; Oliveira, A.P.; Costa, A.S.; Arrigoni-Blank, M.F. Toxicity and repellency of essential oils of Lippia alba chemotypes and their major monoterpenes against stored grain insects. Ind. Crop. Prod. 2015, 71, 31–36. [Google Scholar] [CrossRef]
- Peixoto, M.G.; Costa-Júnior, L.M.; Blank, A.F.; Lima, A.S.; Menezes, T.S.A.; Santos, D.A.; Alves, P.B.; Cavalcanti, S.C.H.; Bacci, L.; Arrigoni-Blank, M.F. Acaricidal activity of essential oils from Lippia alba genotypes and its major components carvone, limonene, and citral against Rhipicephalus microplus. Vet. Parasitol. 2015, 210, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Tak, J.H.; Jovel, E.; Isman, M.B. Contact, fumigant, and cytotoxic activities of thyme and lemongrass essential oils against larvae and an ovarian cell line of the cabbage looper, Trichoplusia ni. J. Pest Sci. 2016, 89, 183–193. [Google Scholar] [CrossRef]
- Autran, E.S.; Neves, I.A.; Silva, C.S.B.; Santos, G.K.N.; Câmara, C.A.G.; Navarro, D.M.A.F. Chemical composition, oviposition deterrent and larvicidal activities against Aedes aegypti of essential oils from Piper marginatum Jacq. (Piperaceae). Bioresour. Technol. 2009, 100, 2284–2288. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Mancini, E.; Camele, I.; Martino, L.D.; de Feo, V. In vivo antifungal activity of two essential oils from Mediterranean plants against postharvest brown rot disease of peach fruit. Ind. Crops Prod. 2015, 66, 11–15. [Google Scholar] [CrossRef]
- Gonzalez, M.S.; Lima, B.G.; Oliveira, A.F.R.; Nunes, D.D.; Fernandes, C.P.; Santos, M.G.; Tietbohl, L.A.C.; Mello, C.B.; Rocha, L.; Feder, D. Effects of essential oil from leaves of Eugenia sulcataon the development of agricultural pest insects. Rev. Bras. Farmacogn. 2014, 24, 413–418. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Mancini, E.; Sakr, S.; de Martino, L.; Mattia, C.A.; de Feo, V.; Camele, I. Antifungal activity of some constituents of Origanum vulgare L. essential oil against postharvest disease of peach fruit. J. Med. Food 2015, 18, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Sakr, S.; Mang, S.M.; de Feo, V.; Camele, I. Antimicrobial activity and chemical composition of three essential oils extracted from Mediterranean aromatic plants. J. Med. Food 2016, 19, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, L.G.L.; Cardoso, M.G.; Zacaroni, L.M.; Lima, R.K. Influência da luz e da temperatura sobre a oxidação do óleo essencial de capim limão (Cymbopogon citratus (D.C.) Stapf). Quim. Nova 2008, 31, 1476–1480. [Google Scholar] [CrossRef]
- Arrigoni-Blank, M.F.; Antoniolli, A.R.; Caetano, L.C.; Campos, D.A.; Blank, A.F.; Alves, P.B. Antinociceptive activity of the volatile oils of Hyptis pectinata L. Poit. (Lamiaceae) genotypes. Phytomedicine 2008, 15, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, P.F.C.; Alviano, W.S.; Nascimento, A.L.C.; Santos, P.O.; Arrigoni-Blank, M.F.; Jesus, R.A.; Azevedo, V.G.; Alviano, D.S.; Bolognese, A.M.; Trindade, R.C. Hyptis pectinata essential oil: Chemical composition and anti-Streptococcus mutans activity. Oral Dis. 2008, 14, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.J.; Dória, G.A.A.; Maia, R.T.; Nunes, R.S.; Carvalho, G.A.; Blank, A.F.; Alves, P.B.; Marçal, R.M.; Cavalcanti, S.C.H. Effects of essential oils on Aedes aegypti larvae: Alternatives to environmentally safe insecticides. Bioresour. Technol. 2008, 99, 3251–3255. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, L.J.R.P.; Guilhon, C.C.; Alviano, D.S.; Matheus, M.E.; Antoniolli, A.R.; Cavalcanti, S.C.H.; Alves, P.B.; Alviano, C.S.; Fernandes, P.D. Characterisation of the anti-inflammatory and antinociceptive activities of the Hyptis pectinata (L.) Poit essential oil. J. Ethnopharmacol. 2011, 134, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Falcao, R.A.; Nascimento, P.L.A.; Souza, S. Antileishmanial Phenylpropanoids from the Leaves of Hyptis pectinata (L.) Poit. J. Evid. Based Complement. Altern. Med. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni-Blank, M.F.; Alcantara-Feitosa, R.B.; Blank, A.F.; Bacci, L.; Santos, A.A.; Silva, I.M.A.; Santos, A.C.C.; Santana, E.D.R.; Oliveria, A.P.; Lima, A.P.S.; et al. Formulação Formicida, Método Para Controlar ou Eliminar Formigas Cortadeiras à Base de Óleo Essencial de Genótipos de Hyptis pectinata. Br. Patent PI 1020160101514, 5 May 2016. [Google Scholar]
- Martins, F.T.; Santos, M.H.; Polo, M.; Barbosa, L.C.A. Variação química do óleo essencial de Hyptis suaveolens (L.) Poit., sob condições de cultivo. Quím. Nova 2006, 29, 1203–1209. [Google Scholar] [CrossRef]
- Nizio, D.A.C.; Brito, F.A.; Sampaio, T.S.; Melo, J.O.; Silva, F.L.S.; Gagliardi, P.L.; Arrigoni-Blank, M.F.; Anjos, C.S.; Alves, P.B.; Wisniewski, A.; et al. Chemical diversity of native populations of Varronia curassavica Jacq. and antifungal activity against Lasiodiplodia theobromae. Ind. Crops Prod. 2015, 76, 437–448. [Google Scholar] [CrossRef]
- Sampaio, T.S.; White, L.A.S.; Nizio, D.A.C.; Melo, J.O.; Almeida, C.S.; Alves, M.F.; Gagliardi, P.R.; Arrigoni-Blank, M.F.; Junior, A.W.; Sobral, M.E.G.; et al. Chemical diversity of a wild population of Myrcia ovata Cambessedes and antifungal activity against Fusarium solani. Ind. Crops Prod. 2016, 86, 196–209. [Google Scholar] [CrossRef]
- McNeil, M.; Facey, P.; Porter, R. Essential Oils from the Hyptis genus—A review (1909–2009). Nat. Prod. Commun. 2011, 6, 1775–1796. [Google Scholar] [PubMed]
- Oliveira, B.M.S.; Melo, C.R.; Alves, P.B.; Santos, A.A.; Santos, A.C.; Santana, A.D.; Araújo, A.P.; Nascimento, P.E.; Blank, A.F.; Bacci, L. Essential oil of Aristolochia trilobata: Synthesis, routes of exposure, acute toxicity, binary mixtures and behavioral effects on leaf-cutting ants. Molecules 2015, 22, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.G. Atividade Formicida de Pogostemon cablin e Sua Nanoformulação Sobre Formigas Cortadeiras. Master’s Thesis, Universidade Federal de Sergipe, São Cristóvão, Brazil, 2015. [Google Scholar]
- Oliveira, M.S.S.; Roel, A.R.; Arruda, E.J.; Marques, A.S. Eficiência de Produtos Vegetais no Controle da Lagarta-do-cartucho-do-milho Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae). Ciênc. Agrotec. 2007, 31, 326–331. [Google Scholar] [CrossRef]
- Bacci, L.; Crespo, A.L.; Galvan, T.L.; Pereira, E.J.; Picanço, M.C.; Silva, G.A.; Chediak, M. Toxicity of insecticides to the sweetpotato whitefly (Hemiptera: Aleyrodidae) and its natural enemies. Pest Manag. Sci. 2007, 63, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.M.G. Desarrollo y Validación de un Método Analítico Mediante LC-QTOF-MS para la Caracterización de Aguas Residuales y Estudio de su Aplicación en Agua de Riego Para Cultivos Hortícolas. Master’s Thesis, Universidad de Almería, Almería, Spain, 2012. [Google Scholar]
- Klaumann, P.R.; Wouk, A.F.P.F.; Sillas, T. Patofisiologia da dor. Arch. Vet. Sci. 2008, 13, 1–12. [Google Scholar] [CrossRef]
- Trindade, H.I.; Batista, M.C.S.; Silva, L.L.B. Dor: Mecanismos envolvidos na sua transmissão e recursos terapêuticos aplicados à sua inibição. Rev. Med. Vet. 2013, 7, 6–18. [Google Scholar]
- Corrêa, J.C.R.; Salgado, H.R.N. Atividade inseticida das plantas e aplicações: Revisão. Rev. Bras. Plantas Med. 2011, 3, 500–506. [Google Scholar] [CrossRef]
- Gomes, S.P.; Favero, S. Avaliação de óleos essenciais de plantas aromáticas com atividade inseticida em Triatoma infestans (Klug, 1834) (Hemiptera: Reduviidae). Acta Sci. Health Sci. 2011, 33, 147–151. [Google Scholar] [CrossRef]
- Delfino, R.T.; Ribeiro, T.S.; Figueroa-Villar, J.D. Organophosphorus compounds as chemical warfare agents: A review. J. Braz. Chem. Soc. 2009, 20, 407–428. [Google Scholar] [CrossRef]
- Pauliquevis, C.F.; Favero, S. Atividade insetistática de óleo essencial de Pothomorphe umbellata sobre Sitophilus zeamais. Rev. Bras. Eng. Agríc. Ambient. 2015, 19, 1192–1196. [Google Scholar] [CrossRef]
- Manna, S.; Bhattacharyya, D.; Mandal, T.K.; Dey, S. Neuropharmacological effects of deltamethrin in rats. J. Vet. Sci. 2006, 72, 133–136. [Google Scholar] [CrossRef]
- Lopez, M.D.; Contreras, J.; Pascual-Villalobos, M.J. Selection for tolerance to volatile monoterpenoids in Sitophilus oryzae (L.), Rhyzopertha dominica (F.) and Cryptolestes pusillus (Schonherr). J. Stored Prod. Res. 2008, 46, 52–58. [Google Scholar] [CrossRef]
- Zapata, N.; Smagghe, G. Repellency and toxicity of essential oils from the leaves and bark of Laurelia sempervirens and Drimys winteri against Tribolium castaneum. Ind. Crops Prod. 2010, 32, 405–410. [Google Scholar] [CrossRef]
- Gallardo, A.; Picollo, M.I.; Mougabure-Cueto, G. Lethal activity of individual and mixed monoterpenoids of geranium essential Oil on Musca domestica. Parasitol. Res. 2015, 114, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.J.; Howes, M.J.; Lee, C.C.; Steventon, G. Uses and abuses of in vitro tests in ethnopharmacology: Visualizing an elephant. J. Ethnopharmacol. 2007, 110, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. Insecticidal properties of several essential oils on the house fly (Musca domestica L.). Phytother. Res. 2008, 22, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.F.M.; Oliveira, L.F.G., Jr.; Santos, M.C.; Narain, N.; Leite Neta, M.T.S. Tempo de destilação e perfil volátil do óleo essencial de aroeira da praia (Schinus terebinthifolius) em Sergipe. Rev. Bras. Plantas Med. 2014, 16, 243–249. [Google Scholar] [CrossRef]
- Santos, E.R.; Wang, S.H.; Correia, F.C.; Costa, I.R.; Sonnenberg, V.; Burini, E.C., Jr.; Onmori, R.K. Influência de diferentes solventes utilizados na deposição de filme de poli (9-vinilcarbazol) em dispositivos OLEDs. Quím. Nova 2014, 37, 1–5. [Google Scholar] [CrossRef]
- Cheméo. Available online: https://www.chemeo.com/cid/56-264-7/Calamusenone (accessed on 20 December 2016).
- Pubchem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281515 (accessed on 20 December 2016).
- Terpene.info. Available online: http://terpene.info/terpene/caryophyllene-oxide/ (accessed on 20 December 2016).
- Froehner, S.; Martins, R.F. Avaliação do destino e bioacumulação de benzo(a)pireno através de simulação computacional. Quim. Nova 2008, 31, 1089–1093. [Google Scholar] [CrossRef]
- Pereira Netto, A.D.; Moreira, J.C.; Dias, A.E.X.O.; Arbilla, G.; Ferreira, L.F.V.; Oliveira, A.S.; Barek, J. Avaliação da contaminação humana por hidrocarbonetos policíclicos aromáticos (HPAs) e seus derivados nitrados (NHPAs): Uma revisão metodológica. Quim. Nova 2000, 23, 765–773. [Google Scholar] [CrossRef]
- Braga, I.A.; Valle, D. Aedes aegypti: Insecticides, mechanisms of action and resistance. Epidemiol. Serv. Saúde 2007, 16, 295–302. [Google Scholar]
- Mendes, S.S.; Bomfim, R.R.; Jesus, H.C.R.; Alves, P.B.; Blank, A.F.; Estevam, C.S.; Antoniolli, A.R.; Thomazzi, S.M. Evaluation of the analgesic and antiinflammatory effects of the essential oil of Lippia gracilis leaves. J. Ethnopharmacol. 2010, 129, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects—A review. Plant Protect. Sci. 2016, 52, 229–241. [Google Scholar]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Marinho, C.G.S.; Della Lucia, T.M.C.; Picanço, M.C. Fatores que dificultam o controle das formigas cortadeiras. Bahia Agric. 2006, 7, 18–21. [Google Scholar]
- Ehlert, P.A.D.; Blank, A.F.; Arrigoni-Blank, M.F.; Paula, J.W.A.; Campos, D.A.; Alviano, C.S. Tempo de hidrodestilação na extração de óleo essencial de sete espécies de plantas medicinais. Rev. Bras. Plantas Med. 2006, 8, 79–80. [Google Scholar]
- Van den Dool, H.; Dec Kratz, P. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatograpy/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Santos, P.O.; Costa, M.J.C.; Alves, J.A.B.; Nascimento, P.F.C.; Melo, D.L.F.M.; Barbosa, A.M., Jr.; Trindade, R.C.; Branco, A.F.; Arrigoni-Blank, M.F.; Alves, P.B.; et al. Chemical composition and antimicrobial activity of the essential oil of Hyptis pectinata (L.) Poit. Quím. Nova 2008, 31, 1648–1652. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT 9.1 User’s Guide; SAS Institute: Cary, NC, USA, 2004. [Google Scholar]
- Hummelbrunner, L.; Isman, M.B. Acute, Sublethal, Antifeedant, and Synergistic Effects of Monoterpenoid Essential Oil Compounds on the Tobacco Cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds β-caryophyllene and caryophyllene oxide are available from the authors. |

{kind=link}
{kind=link}
| Compound | RRI a | Concentration (%) b of the Compounds in H. pectinate Chemotypes | |
|---|---|---|---|
| Plant SAM-016 | Plant SAM-019 | ||
| β-pinene | 974 | 2.26 | - |
| p-cymene | 1020 | 1.32 | - |
| Limonene | 1024 | 1.65 | - |
| δ-elemene | 1335 | 3.92 | - |
| α-cubenene | 1345 | 1.56 | 0.31 |
| α-copaene | 1374 | 4.85 | 1.12 |
| β-bourbonene | 1387 | 1.02 | 0.47 |
| β-elemene | 1389 | 7.79 | 2.05 |
| β-caryophyllene | 1417 | 17.66 | 9.42 |
| γ-elemene | 1434 | - | 0.63 |
| (Z)-muurola-3,5-diene | 1448 | - | 0.56 |
| α-humulene | 1452 | 1.66 | 0.81 |
| γ-muurolene | 1478 | 5.66 | - |
| Germacrene D | 1484 | - | 1.87 |
| (Z)-β-guaiene | 1492 | - | 4.77 |
| Bicyclogermacrene | 1500 | 4.57 | - |
| Lepidozene | 1502 | - | 0.68 |
| γ-cadinene | 1513 | - | 2.72 |
| (E)-calamenene | 1521 | 5.76 | 1.81 |
| Spathulenol | 1577 | 10.09 | - |
| Caryophyllene oxide | 1582 | 11.52 | 22.89 |
| Globulol | 1590 | 1.82 | - |
| α-acorenol | 1632 | - | 0.15 |
| Epi-α-cadinol | 1638 | 4.88 | - |
| Cubenol | 1645 | - | 3.29 |
| α-cadinol | 1652 | 2.22 | 1.59 |
| Calamusenone | 1676 | - | 36.08 |
| Essential oil content (%) | 0.67 | 0.62 | |
| Treatment | N. Insects | LD50 a (95%CI) (μg mg−1) | LD90 (95%CI) (μg mg−1) | Slope b | Χ2 | p-Value |
|---|---|---|---|---|---|---|
| Acromyrmex balzani | ||||||
| Chemotype β-caryophyllene | 391 | 8.18 (7.09–8.96) | 31.16 (22.17–52.47) | 2.20 | 2.59 | 0.53 |
| Chemotype calamusenone | 406 | 3.48 (3.05–3.91) | 9.84 (8.32–12.42) | 2.84 | 2.97 | 0.22 |
| β-caryophyllene | 224 | 15.59 (13.87–17.68) | 47.31 (37.41–65.98) | 2.66 | 1.92 | 0.62 |
| Calamusenone | 811 | 13.21 (12.56–13.82) | 18.93 (17.87–20.40) | 8.19 | 1.90 | 0.61 |
| Caryophyllene oxide | 252 | 18.97 (17.00–21.08) | 49.53 (40.47–66.70) | 3.05 | 2.97 | 0.22 |
| Atta sexdens rubropilosa | ||||||
| Chemotype β-caryophyllene | 310 | 3.61 (3.13–4.20) | 11.67 (9.20–16.09) | 2.51 | 2.01 | 0.36 |
| Chemotype calamusenone | 406 | 4.65 (3.77–5.52) | 25.80 (18.05–47.17) | 1.72 | 3.60 | 0.16 |
| β-caryophyllene | 392 | 4.76 (4.14–5.46) | 14.74 (12.09–18.99) | 2.60 | 0.24 | 0.88 |
| Calamusenone | 335 | 5.96 (5.25–6.61) | 12.19 (10.88–14.11) | 4.12 | 5.18 | 0.07 |
| Caryophyllene oxide | 644 | 35.34 (29.47–40.99) | 158.92 (107.62–355.32) | 1.96 | 0.05 | 0.97 |
| Treatment | N. Insects | LC50 a (95%CI) (μL L−1) | LC90 (95%CI) (μL L−1) | Slope b | Χ2 | p-Value |
|---|---|---|---|---|---|---|
| Acromyrmex balzani | ||||||
| Chemotype β-caryophyllene | 294 | 1.82 (1.57–2.08) | 6.34 (5.14–8.37) | 2.36 | 1.86 | 0.60 |
| Chemotype calamusenone | 173 | 0.59 (0.53–0.65) | 1.35 (1.17–1.63) | 3.58 | 4.50 | 0.10 |
| β-caryophyllene | 497 | >100.00 * | - | - | - | - |
| Calamusenone | 441 | >100.00 * | - | - | - | - |
| Caryophyllene oxide | 495 | >100.00 * | - | - | - | - |
| Atta sexdens rubropilosa | ||||||
| Chemotype β-caryophyllene | 203 | 1.18 (0.95–1.41) | 6.15 (4.47–10.40) | 1.79 | 5.30 | 0.06 |
| Chemotype calamusenone | 171 | 2.15 (2.04–2.29) | 3.52 (3.18–4.05) | 6.01 | 1.52 | 0.52 |
| β-caryophyllene | 422 | >100.00 * | - | - | - | - |
| Calamusenone | 392 | >100.00 * | - | - | - | - |
| Caryophyllene oxide | 392 | >100.00 * | - | - | - | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feitosa-Alcantara, R.B.; Bacci, L.; Blank, A.F.; Alves, P.B.; Silva, I.M.d.A.; Soares, C.A.; Sampaio, T.S.; Nogueira, P.C.d.L.; Arrigoni-Blank, M.D.F. Essential Oils of Hyptis pectinata Chemotypes: Isolation, Binary Mixtures and Acute Toxicity on Leaf-Cutting Ants. Molecules 2017, 22, 621. https://doi.org/10.3390/molecules22040621
Feitosa-Alcantara RB, Bacci L, Blank AF, Alves PB, Silva IMdA, Soares CA, Sampaio TS, Nogueira PCdL, Arrigoni-Blank MDF. Essential Oils of Hyptis pectinata Chemotypes: Isolation, Binary Mixtures and Acute Toxicity on Leaf-Cutting Ants. Molecules. 2017; 22(4):621. https://doi.org/10.3390/molecules22040621
Chicago/Turabian StyleFeitosa-Alcantara, Rosana B., Leandro Bacci, Arie F. Blank, Péricles B. Alves, Indira Morgana de A. Silva, Caroline A. Soares, Taís S. Sampaio, Paulo Cesar de L. Nogueira, and Maria De Fátima Arrigoni-Blank. 2017. "Essential Oils of Hyptis pectinata Chemotypes: Isolation, Binary Mixtures and Acute Toxicity on Leaf-Cutting Ants" Molecules 22, no. 4: 621. https://doi.org/10.3390/molecules22040621
APA StyleFeitosa-Alcantara, R. B., Bacci, L., Blank, A. F., Alves, P. B., Silva, I. M. d. A., Soares, C. A., Sampaio, T. S., Nogueira, P. C. d. L., & Arrigoni-Blank, M. D. F. (2017). Essential Oils of Hyptis pectinata Chemotypes: Isolation, Binary Mixtures and Acute Toxicity on Leaf-Cutting Ants. Molecules, 22(4), 621. https://doi.org/10.3390/molecules22040621