
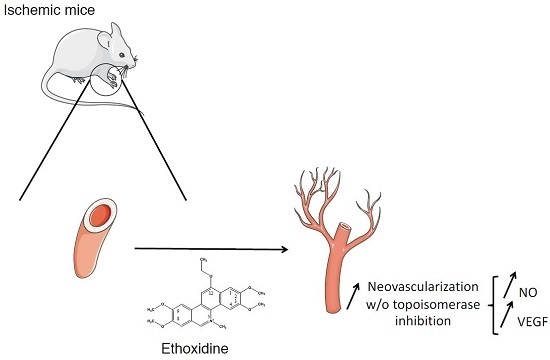
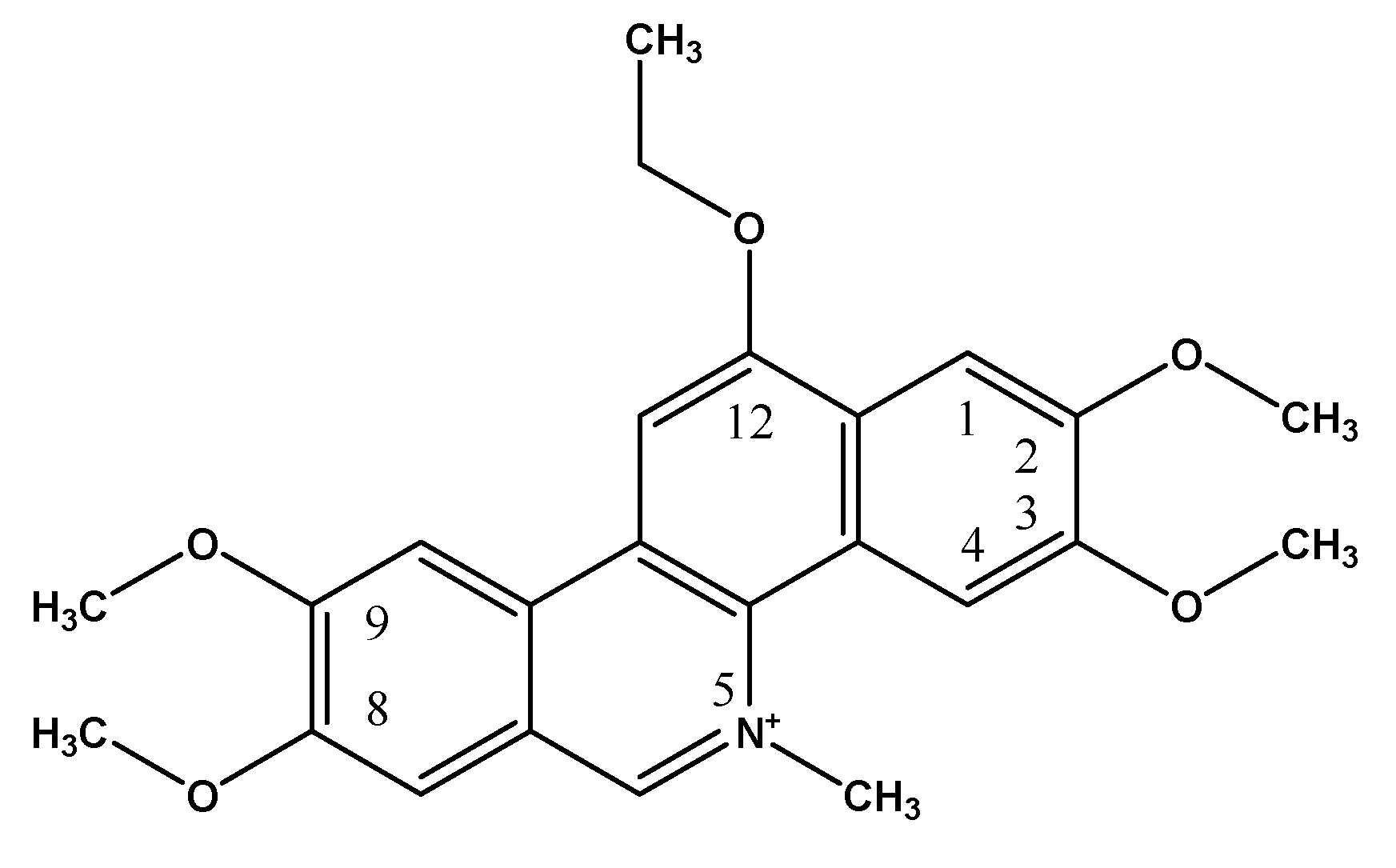
Pro-Angiogenic Effects of Low Dose Ethoxidine in a Murine Model of Ischemic Hindlimb: Correlation between Ethoxidine Levels and Increased Activation of the Nitric Oxide Pathway

 , ,
, ,  ,
, 
Abstract
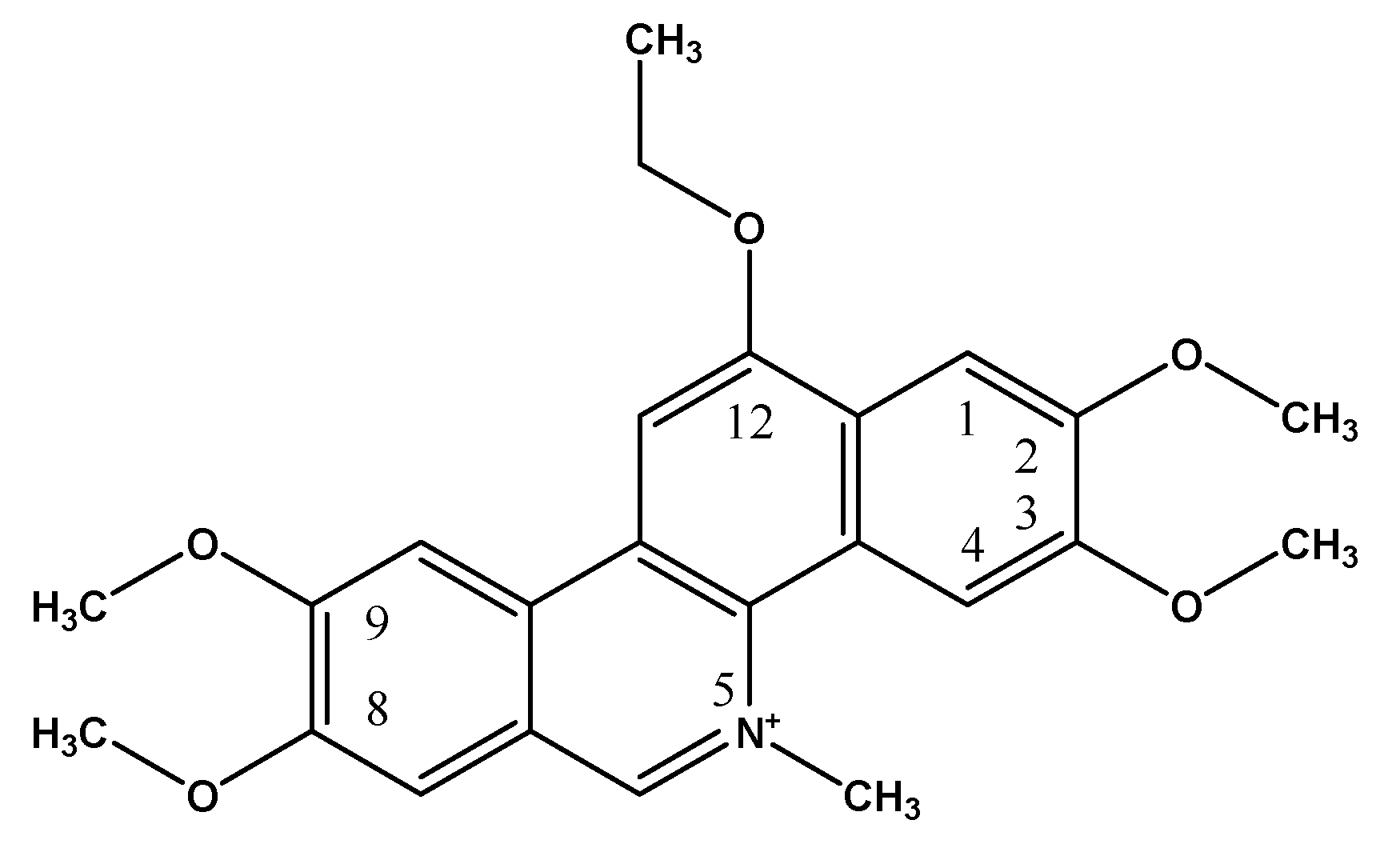
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Ethoxidine Does Not Modify Blood Pressure or Body Weight
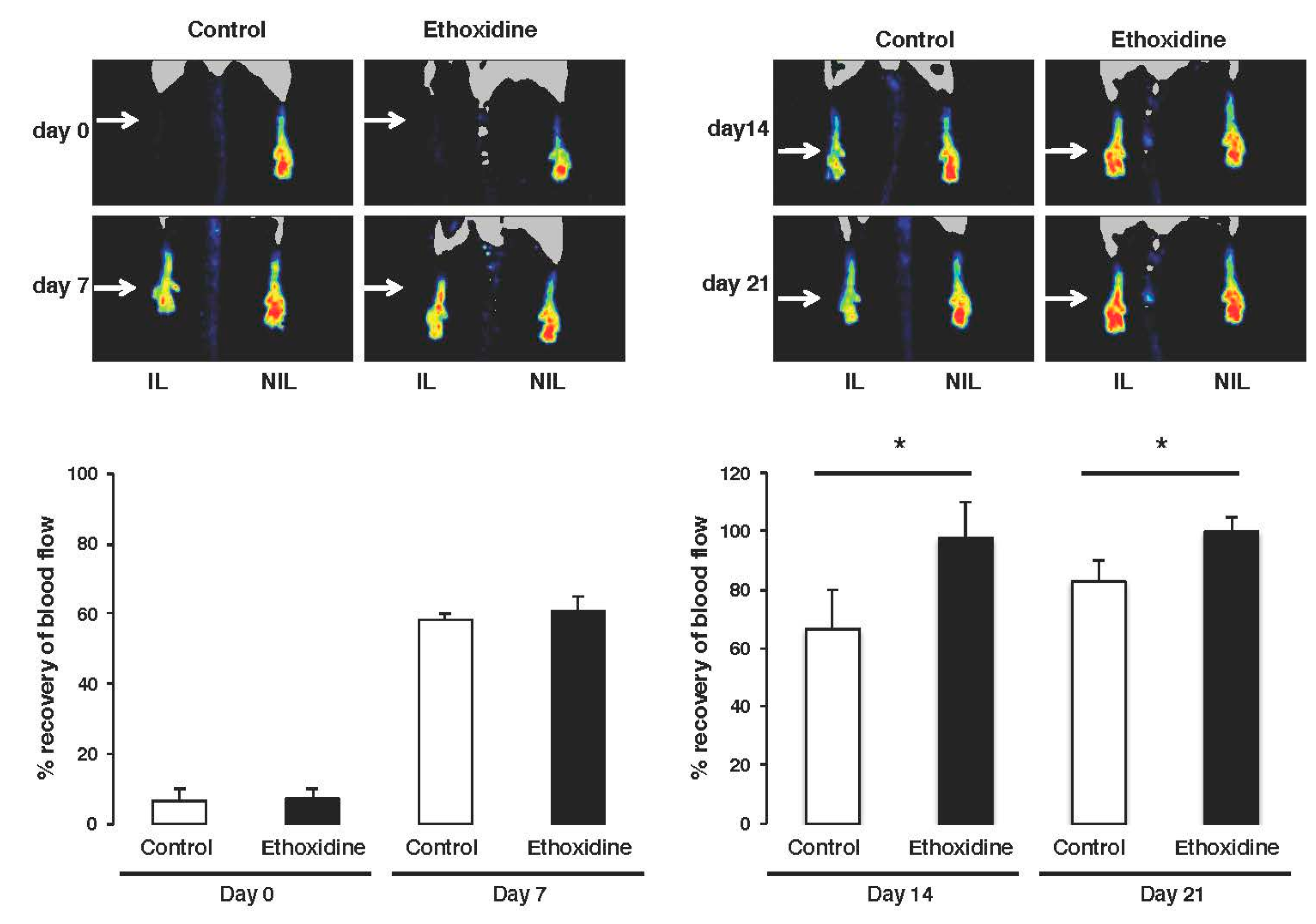
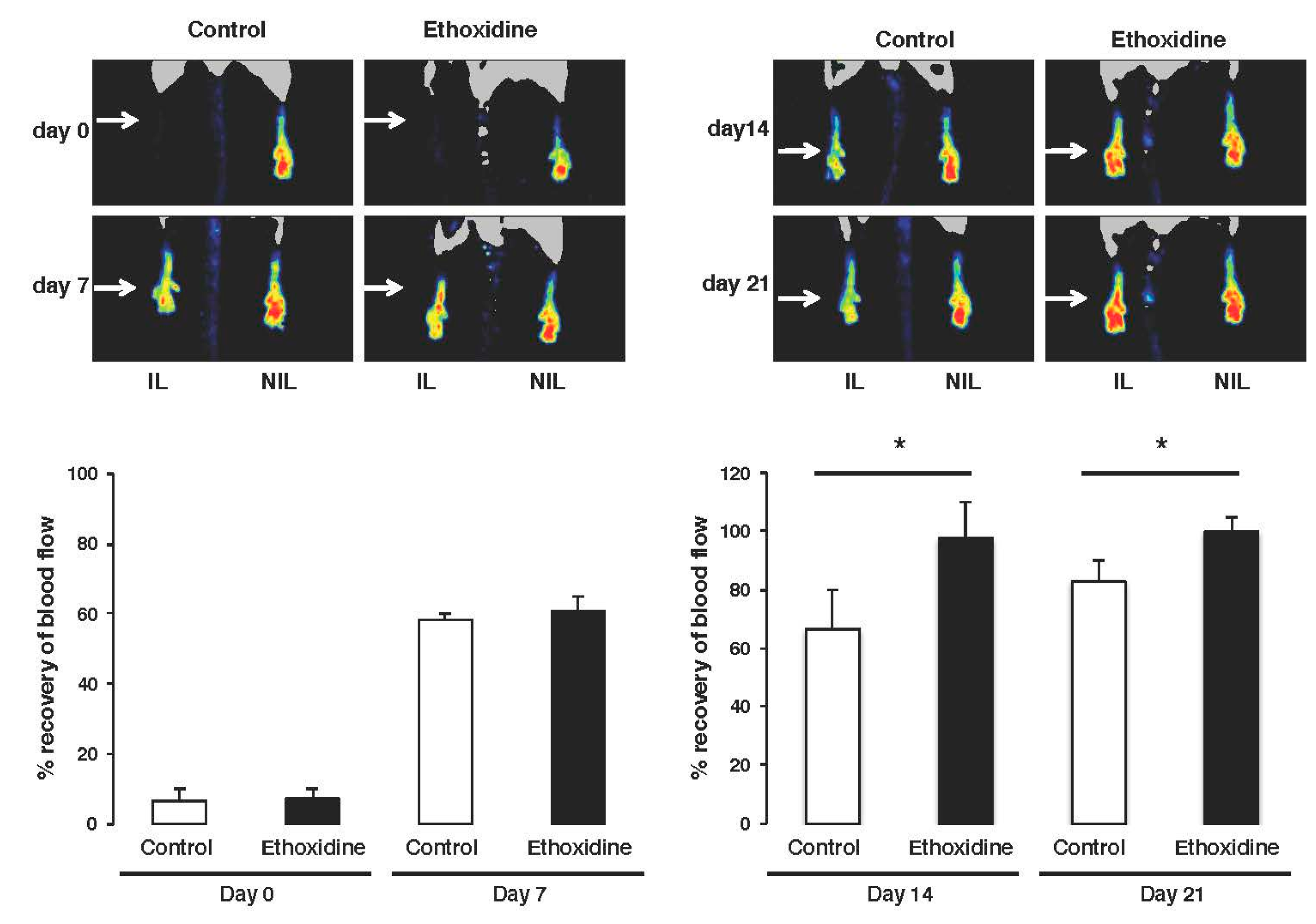
2.2. Ethoxidine Promotes Blood Flow Recovery after Induction of Hindlimb Ischemia
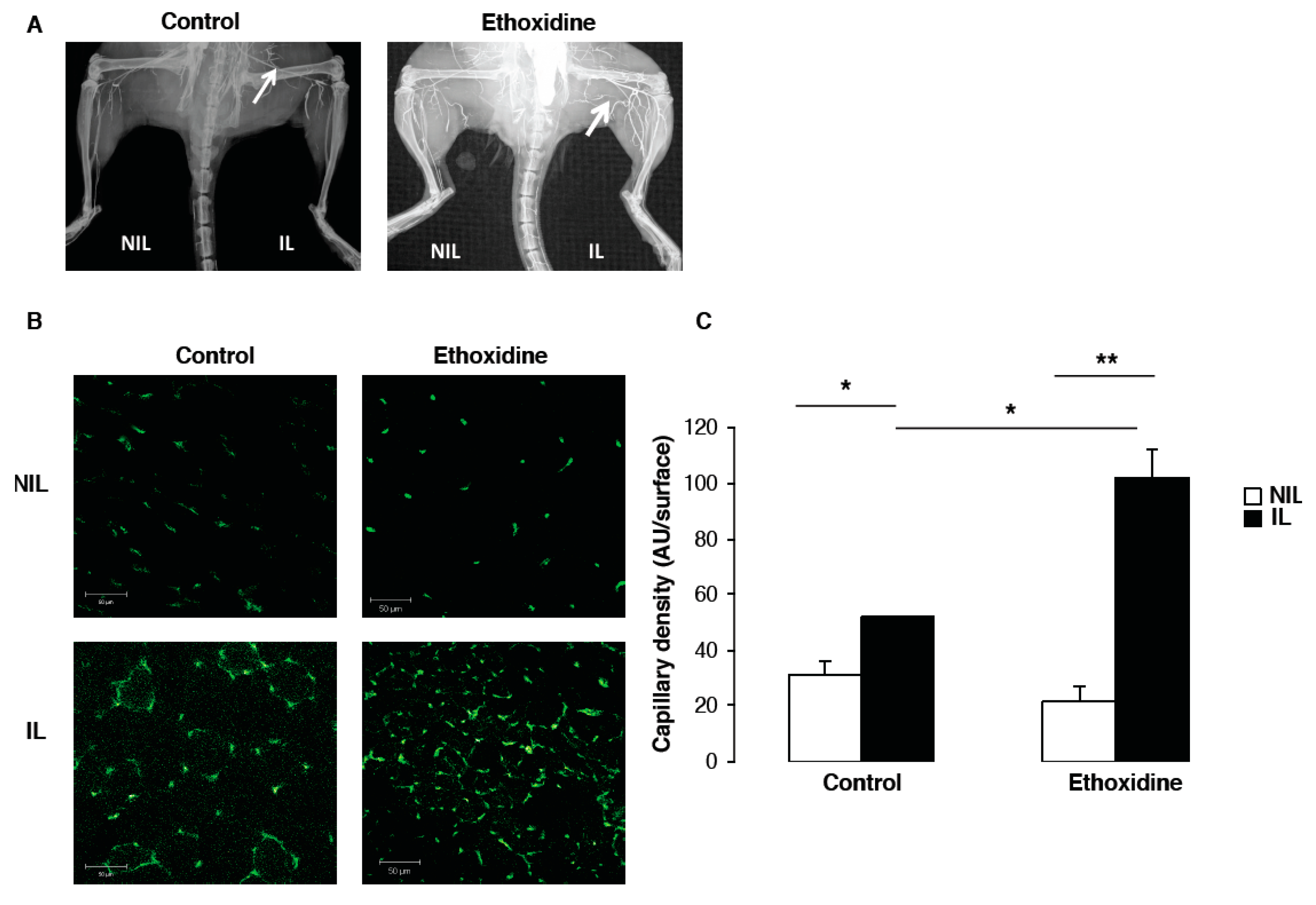
2.3. Ethoxidine Enhances Vascular Density after Induction of Hindlimb Ischemia
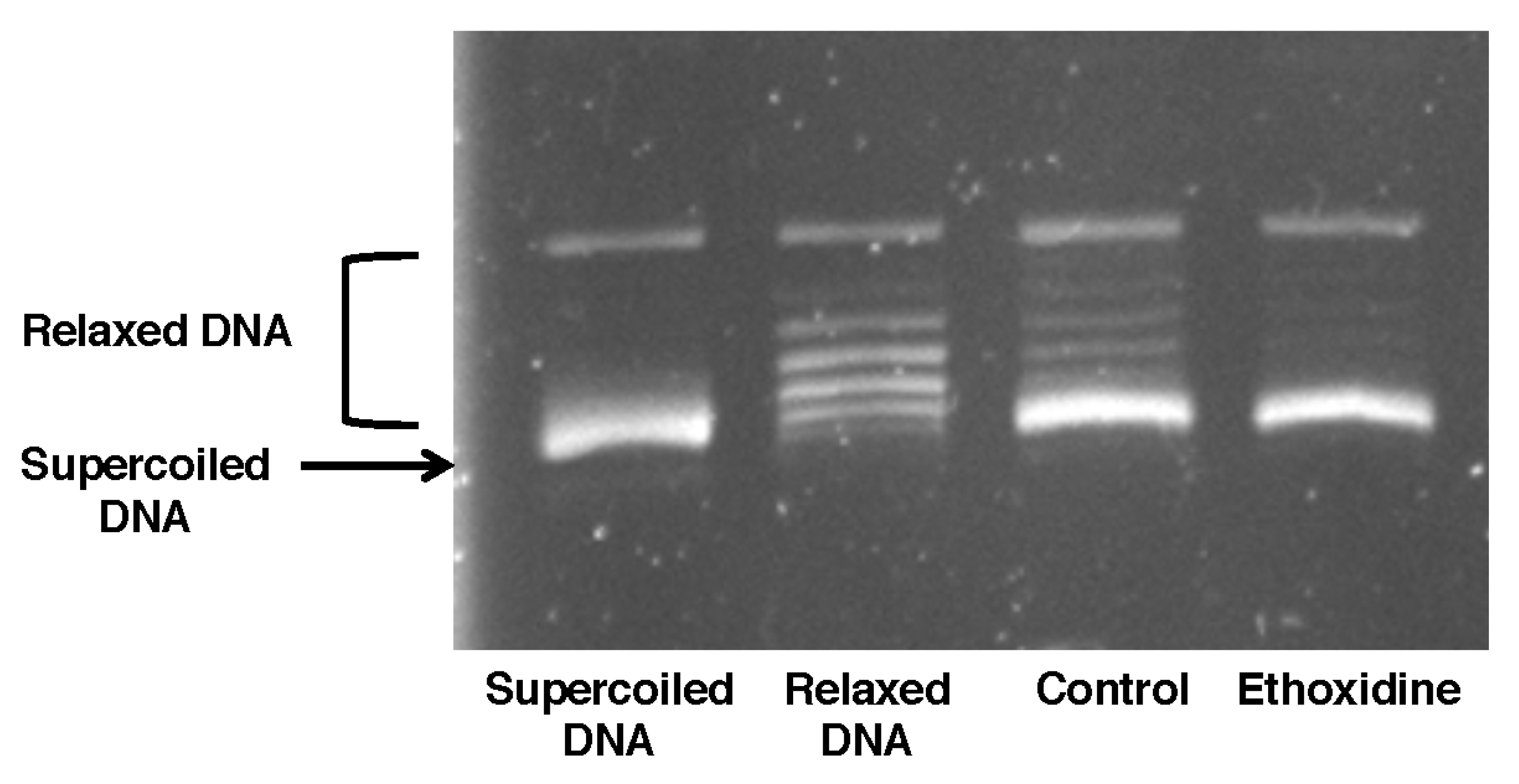
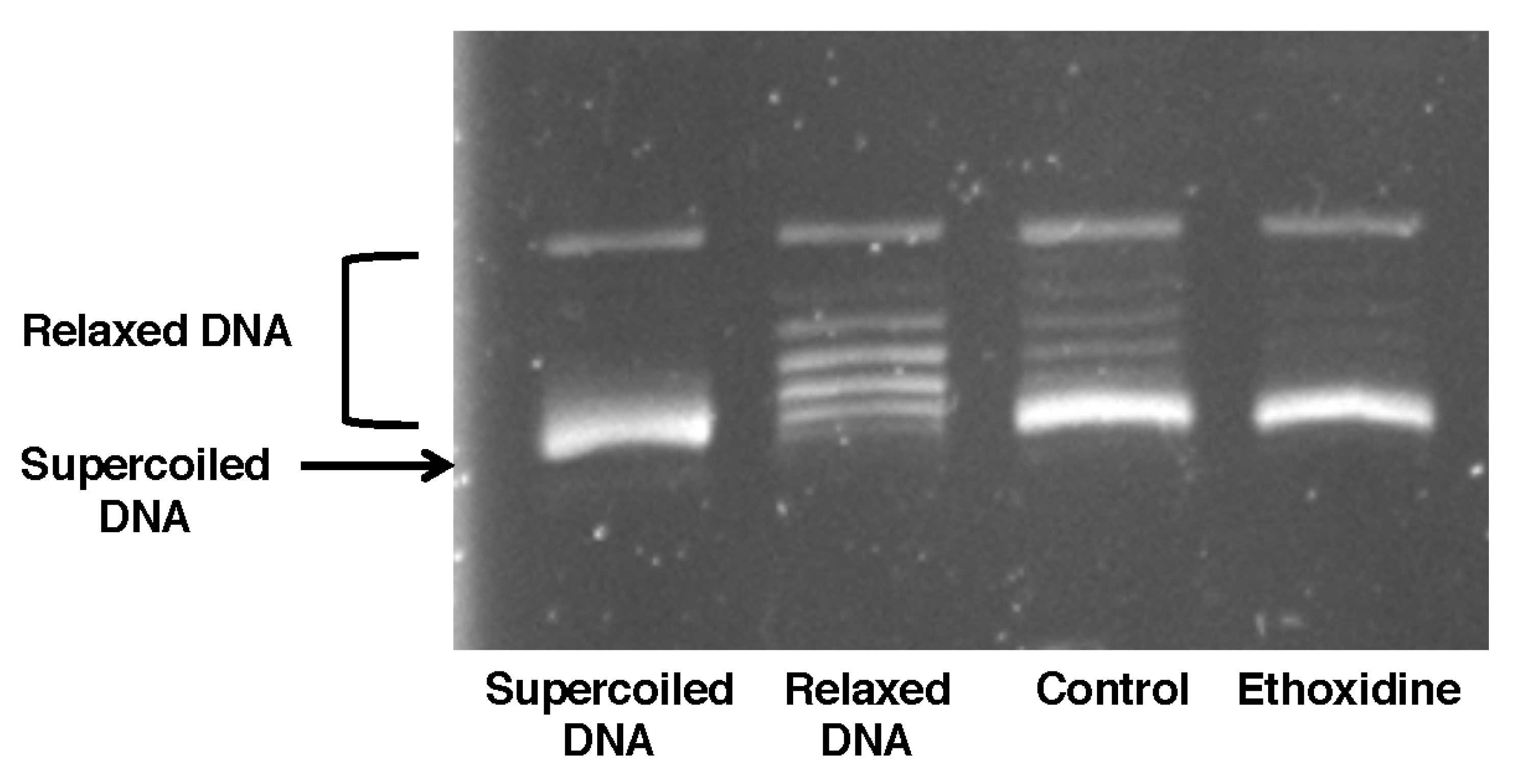
2.4. Ethoxidine Does Not Inhibit Topoisomerase I Activity in Ischemic Hindlimb
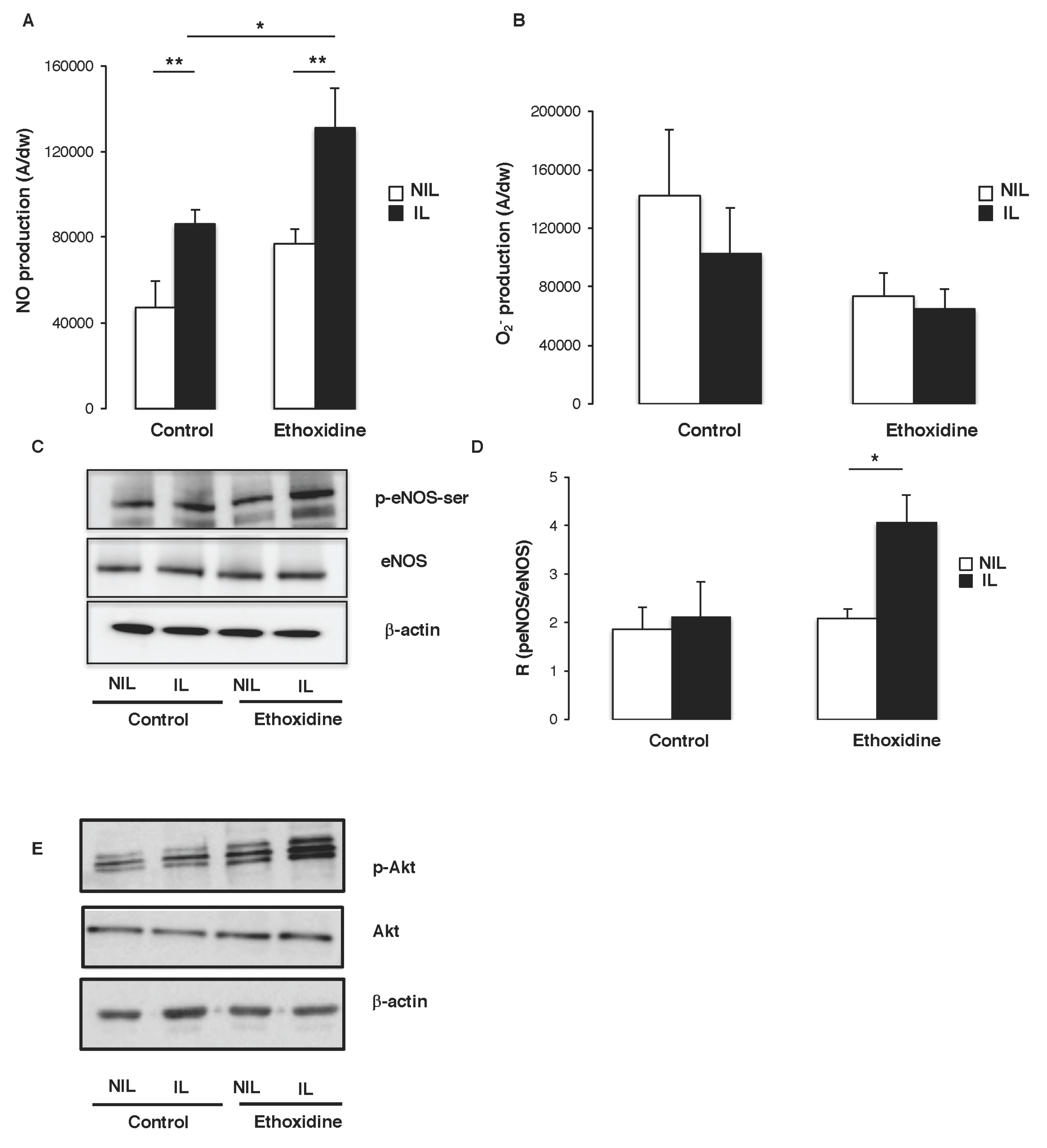
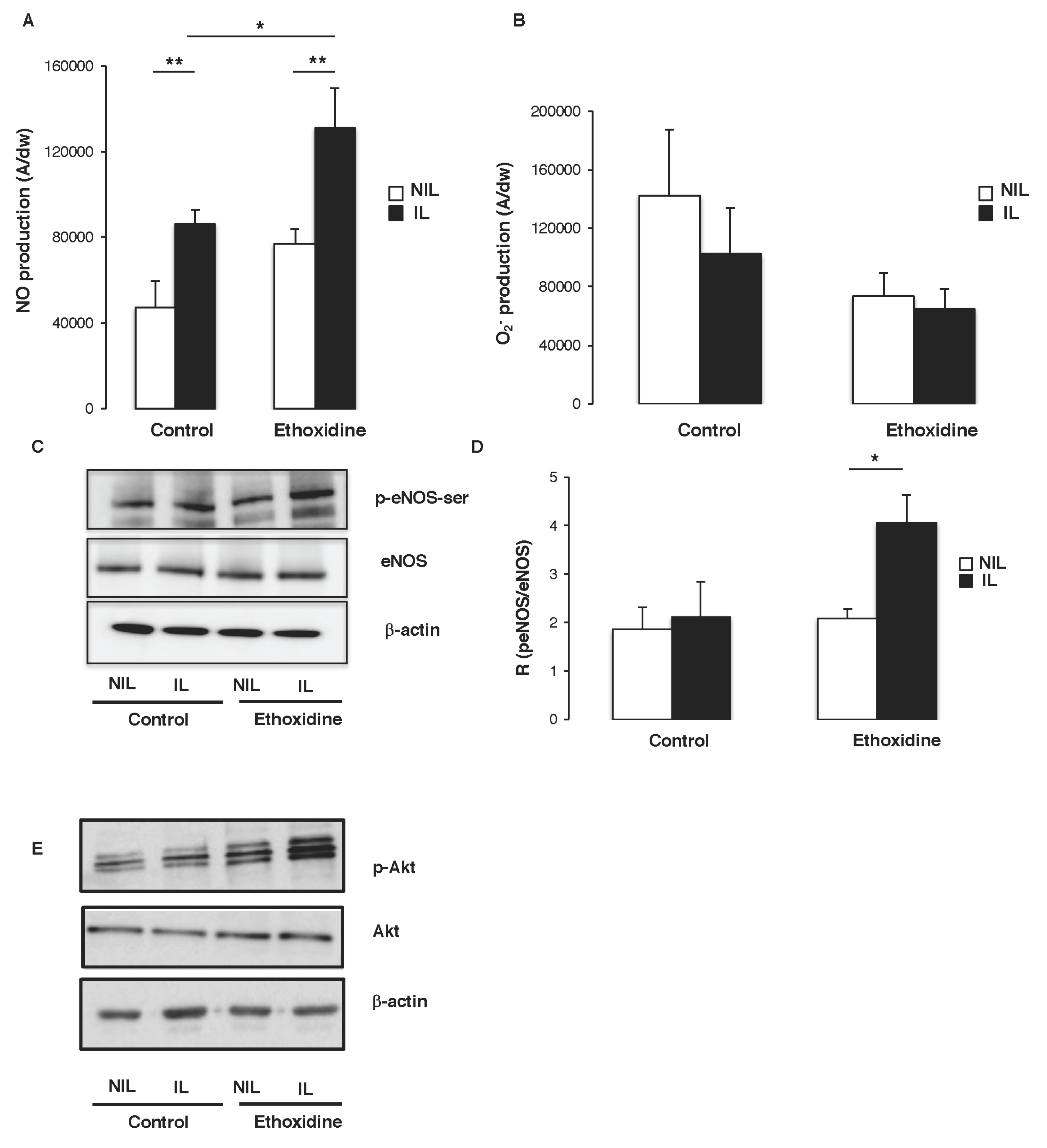
2.5. Ethoxidine Induces NO Production by Enhancing eNOS Activity in Skeletal Muscle
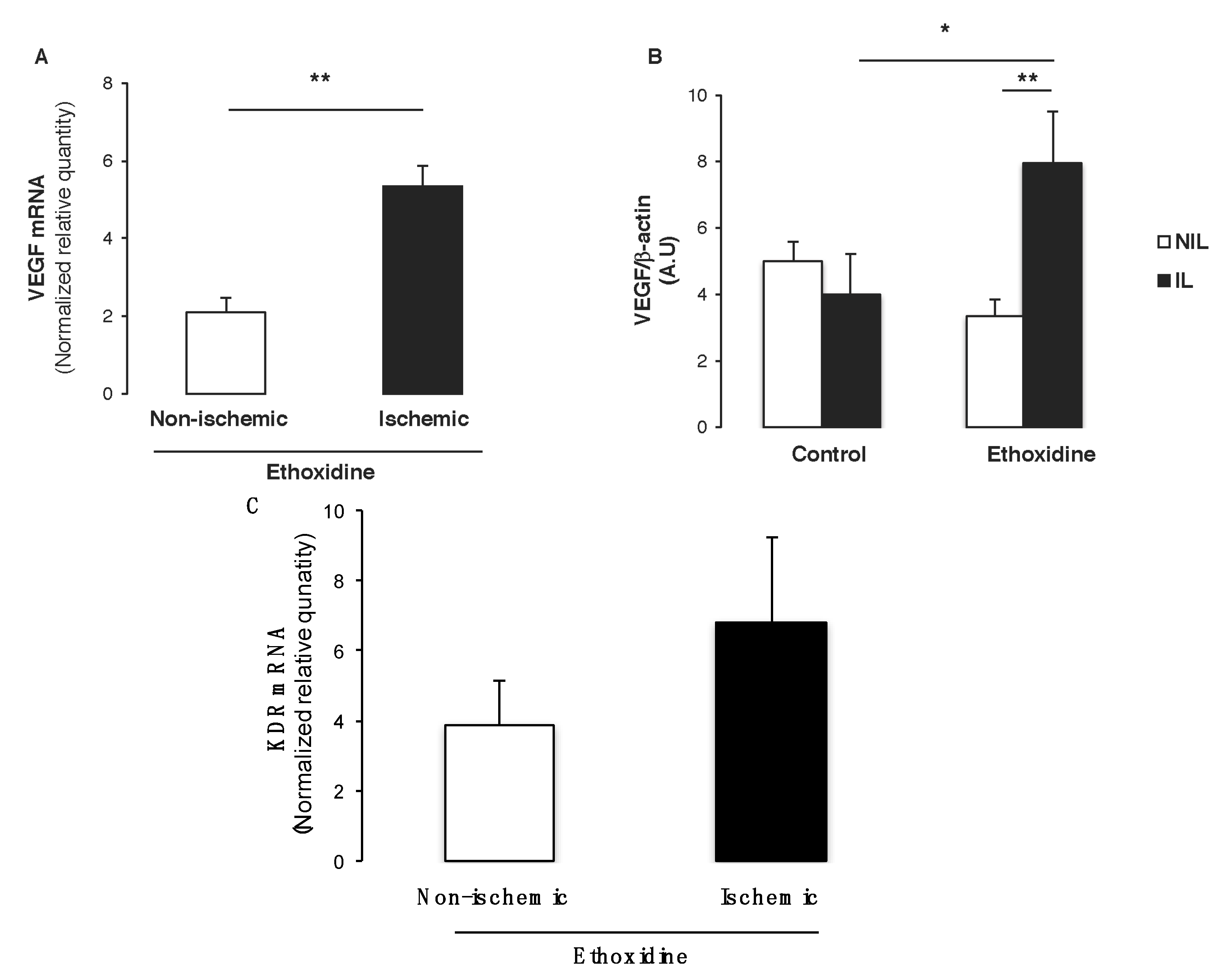
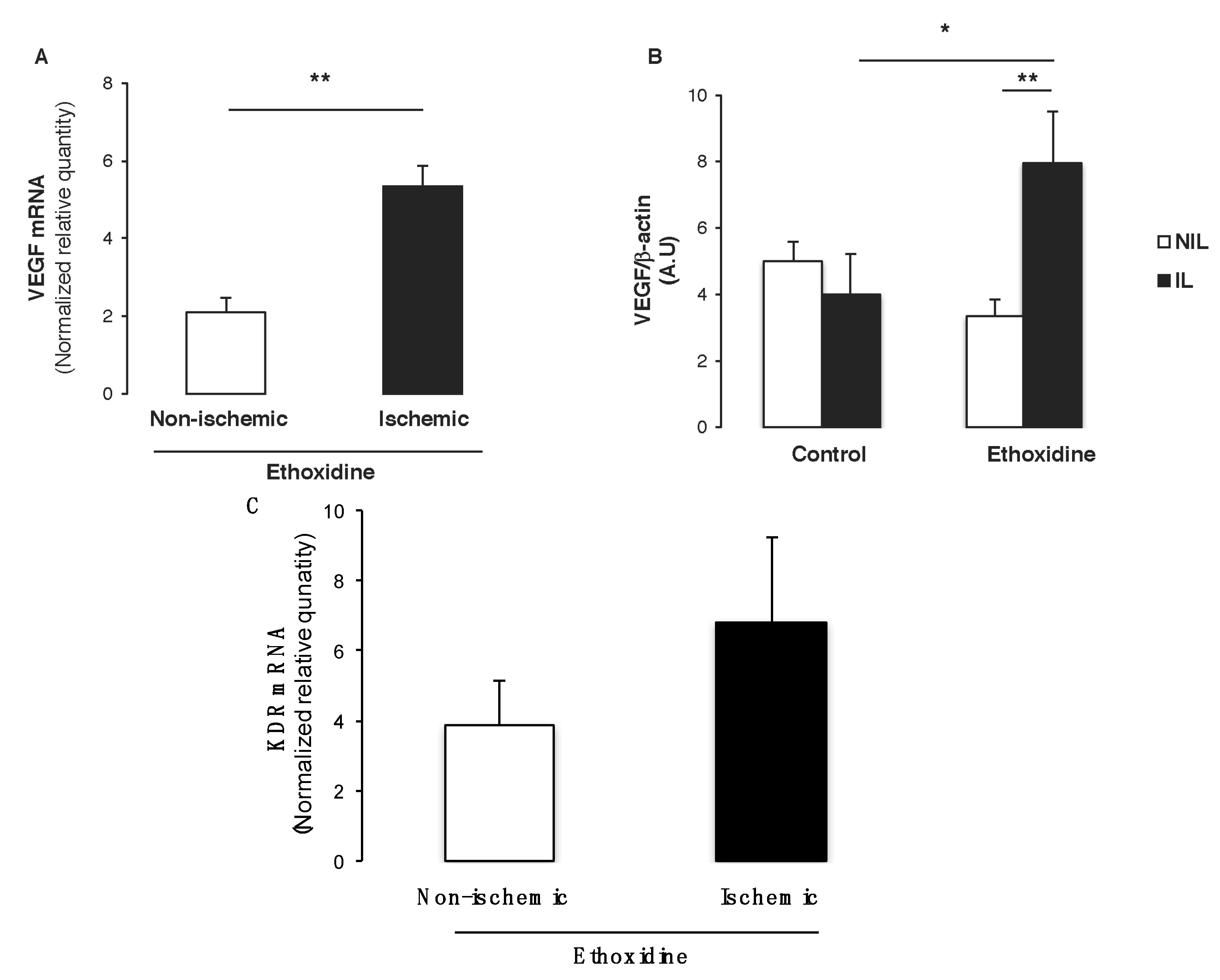
2.6. Ethoxidine Promotes VEGF Expression in Skeletal Muscle from Ischemic Leg
3. Material and Methods
3.1. Mouse Model of Hindlimb Ischemia
3.2. Blood Pressure Measurement
3.3. Evaluation of Plasma Concentration of Ethoxidine
3.4. DNA Topoisomerase I Activity Assays
3.5. Laser-Doppler Whole Body Imaging
3.6. Angiography
3.7. Arteriolar and Capillary Density
3.8. NO Determination by Electron Paramagnetic Resonance (EPR)
3.9. Quantitative Real-Time Reverse Transcription–Polymerase Chain Reaction Analysis
3.10. Western Blot
3.11. Statistical Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of interest
References
- Clark, R.L.; Deane, F.M.; Anthony, N.G.; Johnston, B.F.; McCarthy, F.O.; Mackay, S.P. Exploring DNA topoisomerase i inhibition by the benzo[c]phenanthridines fagaronine and ethoxidine using steered molecular dynamics. Bioorg. Med. Chem. 2007, 15, 4741–4752. [Google Scholar] [CrossRef] [PubMed]
- Mackay, S.; Comoe, L.; Desoize, B.; Duval, O.; Jardillier, J.; Waigh, R. The effect of 12-alkoxy modification on the in vitro antileukaemic activity of n-methyl 2,3,8,9-tetramethoxybenzo[c]phenanthridinium salts. Anti-Cancer Drug Des. 1998, 13, 797–813. [Google Scholar]
- Lynch, M.; Duval, O.; Sukhanova, A.; Devy, J.; MacKay, S.; Waigh, R.; Nabiev, I. Synthesis, biological activity and comparative analysis of dna binding affinities and human dna topoisomerase i inhibitory activities of novel 12-alkoxy-benzo[c]phenanthridinium salts. Bioorg. Med. Chem. Lett. 2001, 11, 2643–2646. [Google Scholar] [CrossRef]
- Fleury, F.; Sukhanova, A.; Ianoul, A.; Devy, J.; Kudelina, I.; Duval, O.; Alix, A.; Jardillier, J.; Nabiev, I. Molecular determinants of site-specific inhibition of human dna topoisomerase i by fagaronine and ethoxidine. Relation to dna binding. J. Biol. Chem. 2000, 275, 3501–3509. [Google Scholar] [CrossRef] [PubMed]
- Soret, J.; Gabut, M.; Dupon, C.; Kohlhagen, G.; Stévenin, J.; Pommier, Y.; Tazi, J. Altered serine/arginine-rich protein phosphorylation and exonic enhancer-dependent splicing in mammalian cells lacking topoisomerase i. Cancer Res. 2003, 63, 8203–8211. [Google Scholar] [PubMed]
- Eisenreich, A.; Bogdanov, V.; Zakrzewicz, A.; Pries, A.; Antoniak, S.; Poller, W.; Schultheiss, H.; Rauch, U. Cdc2-like kinases and dna topoisomerase i regulate alternative splicing of tissue factor in human endothelial cells. Circ. Res. 2009, 104, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, A.; Boltzen, U.; Poller, W.; Schultheiss, H.; Rauch, U. Effects of the cdc2-like kinase-family and DNA topoisomerase i on the alternative splicing of enos in tnf-alpha-stimulated human endothelial cells. Biol. Chem. 2008, 389, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, F.; Zhang, S.; Jia, J.; Li, Z.; Guo, X.; Yang, Y.; Sun, H.; You, Q. Indenoisoquinoline derivatives as topoisomerase i inhibitors that suppress angiogenesis by affecting the hif signaling pathway. Biomed. Pharmacother. 2013, 67, 715–722. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, J.; Shapiro, R.; Ren, C.; Chuang, N.; Cohen, H.; Potmesil, M. Antiangiogenic effects of camptothecin analogues 9-amino-20(s)-camptothecin, topotecan, and cpt-11 studied in the mouse cornea model. Clin. Cancer Res. 1999, 5, 181–187. [Google Scholar] [PubMed]
- Gaur, S.; Wang, Y.; Kretzner, L.; Chen, L.; Yen, T.; Wu, X.; Yuan, Y.C.; Davis, M.; Yen, Y. Pharmacodynamic and pharmacogenomic study of the nanoparticle conjugate of camptothecin crlx101 for the treatment of cancer. Nanomedicine 2014, 10, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.; Germain, G.; Harwood, F.; Schuetz, J.; Stewart, C.; Buchdunger, E.; Traxler, P. Imatinib mesylate is a potent inhibitor of the abcg2 (bcrp) transporter and reverses resistance to topotecan and sn-38 in vitro. Cancer Res. 2004, 64, 2333–2337. [Google Scholar] [CrossRef] [PubMed]
- Devy, J.; Ouchani, F.; Oudot, C.; Helesbeux, J.; Vanquelef, E.; Salesse, S.; Rabenoelina, F.; Al-Khara, S.; Letinois, I.; Duval, O.; et al. The anti-invasive activity of synthetic alkaloid ethoxyfagaronine on l1210 leukemia cells is mediated by down-regulation of plasminogen activators and mt1-mmp expression and activity. Invest. New Drugs 2010, 29, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Clere, N.; Faure, S.; Helesbeux, J.J.; Duval, O.; Andriantsitohaina, R. Paradoxical effects of ethoxidine, a topoisomerase i inhibitor, in the cellular processes leading to angiogenesis on endothelial cells. Carcinogenesis 2011, 32, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Limbourg, A.; Korff, T.; Napp, L.C.; Schaper, W.; Drexler, H.; Limbourg, F.P. Evaluation of postnatal arteriogenesis and angiogenesis in a mouse model of hind-limb ischemia. Nat. Protoc. 2009, 4, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Deroussent, A.; Ré, M.; Hoellinger, H.; Vanquelef, E.; Duval, O.; Sonnier, M.; Cresteil, T. In vitro metabolism of ethoxidine by human cyp1a1 and rat microsomes: Identification of metabolites by high-performance liquid chromatography combined with electrospray tandem mass spectrometry and accurate mass measurements by time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 474–482. [Google Scholar] [PubMed]
- Innocenti, F.; Undevia, S.D.; Iyer, L.; Chen, P.X.; Das, S.; Kocherginsky, M.; Karrison, T.; Janisch, L.; Ramírez, J.; Rudin, C.M.; et al. Genetic variants in the udp-glucuronosyltransferase 1a1 gene predict the risk of severe neutropenia of irinotecan. J. Clin. Oncol. 2004, 22, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Kümler, I.; Brünner, N.; Stenvang, J.; Balslev, E.; Nielsen, D.L. A systematic review on topoisomerase 1 inhibition in the treatment of metastatic breast cancer. Breast Cancer Res. Treat. 2013, 138, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Madeddu, P.; Emanueli, C.; Spillmann, F.; Meloni, M.; Bouby, N.; Richer, C.; Alhenc-Gelas, F.; Van Weel, V.; Eefting, D.; Quax, P.H.; et al. Murine models of myocardial and limb ischemia: Diagnostic end-points and relevance to clinical problems. Vascul. Pharmacol. 2006, 45, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Papapetropoulos, A.; García-Cardeña, G.; Madri, J.A.; Sessa, W.C. Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells. J. Clin. Invest. 1997, 100, 3131–3139. [Google Scholar] [CrossRef] [PubMed]
- Pfosser, A.; Thalgott, M.; Büttner, K.; Brouet, A.; Feron, O.; Boekstegers, P.; Kupatt, C. Liposomal hsp90 cDNA induces neovascularization via nitric oxide in chronic ischemia. Cardiovasc. Res. 2005, 65, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Couffinhal, T.; Silver, M.; Zheng, L.P.; Kearney, M.; Witzenbichler, B.; Isner, J.M. Mouse model of angiogenesis. Am. J. Pathol. 1998, 152, 1667–1679. [Google Scholar] [PubMed]
- Baron-Menguy, C.; Bocquet, A.; Guihot, A.L.; Chappard, D.; Amiot, M.J.; Andriantsitohaina, R.; Loufrani, L.; Henrion, D. Effects of red wine polyphenols on postischemic neovascularization model in rats: Low doses are proangiogenic, high doses anti-angiogenic. FASEB J. 2007, 21, 3511–3521. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Deindl, E.; Buschmann, I.; Hoefer, I.E.; Podzuweit, T.; Boengler, K.; Vogel, S.; van Royen, N.; Fernandez, B.; Schaper, W. Role of ischemia and of hypoxia-inducible genes in arteriogenesis after femoral artery occlusion in the rabbit. Circ. Res. 2001, 89, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Hershey, J.C.; Baskin, E.P.; Corcoran, H.A.; Bett, A.; Dougherty, N.M.; Gilberto, D.B.; Mao, X.; Thomas, K.A.; Cook, J.J. Vascular endothelial growth factor stimulates angiogenesis without improving collateral blood flow following hindlimb ischemia in rabbits. Heart Vessels 2003, 18, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Kazi, A.A.; Lee, W.S.; Wagner, E.; Becker, P.M. Vegf, fetal liver kinase-1, and permeability increase during unilateral lung ischemia. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2000, 279, L460–L467. [Google Scholar] [PubMed]
- Hazarika, S.; Dokun, A.O.; Li, Y.; Popel, A.S.; Kontos, C.D.; Annex, B.H. Impaired angiogenesis after hindlimb ischemia in type 2 diabetes mellitus: Differential regulation of vascular endothelial growth factor receptor 1 and soluble vascular endothelial growth factor receptor 1. Circ. Res. 2007, 101, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Sawano, A.; Takahashi, T.; Yamaguchi, S.; Aonuma, M.; Shibuya, M. Flt-1 but not kdr/flk-1 tyrosine kinase is a receptor for placenta growth factor, which is related to vascular endothelial growth factor. Cell Growth Differ. 1996, 7, 213–221. [Google Scholar] [PubMed]
- Hirst, D.G.; Robson, T. Nitric oxide physiology and pathology. Methods Mol. Biol. 2011, 704, 1–13. [Google Scholar] [PubMed]
- Ziche, M.; Morbidelli, L.; Choudhuri, R.; Zhang, H.T.; Donnini, S.; Granger, H.J.; Bicknell, R. Nitric oxide synthase lies downstream from vascular endothelial growth factor-induced but not basic fibroblast growth factor-induced angiogenesis. J. Clin. Invest. 1997, 99, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Murohara, T.; Asahara, T.; Silver, M.; Bauters, C.; Masuda, H.; Kalka, C.; Kearney, M.; Chen, D.; Symes, J.F.; Fishman, M.C.; et al. Nitric oxide synthase modulates angiogenesis in response to tissue ischemia. J. Clin. Invest. 1998, 101, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Benameur, T.; Soleti, R.; Porro, C.; Andriantsitohaina, R.; Martínez, M.C. Microparticles carrying sonic hedgehog favor neovascularization through the activation of nitric oxide pathway in mice. PLoS ONE 2010, 5, e12688. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Shibata, R.; Ouchi, N.; Sugimoto, M.; Murohara, T.; Komori, K. Metformin stimulates ischemia-induced revascularization through an enos dependent pathway in the ischemic hindlimb mice model. J. Vasc. Surg. 2015, 61, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, S.R.; Bae, S.K.; Choi, Y.K.; Bae, Y.H.; Kim, E.C.; Kim, W.J.; Jang, H.O.; Yun, I.; Kim, Y.M.; et al. Neuromedin b induces angiogenesis via activation of erk and akt in endothelial cells. Exp. Cell Res. 2009, 315, 3359–3369. [Google Scholar] [CrossRef] [PubMed]
- Daher, Z.; Boulay, P.L.; Desjardins, F.; Gratton, J.P.; Claing, A. Vascular endothelial growth factor receptor-2 activates adp-ribosylation factor 1 to promote endothelial nitric-oxide synthase activation and nitric oxide release from endothelial cells. J. Biol. Chem. 2010, 285, 24591–24599. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Zheng, J.; Aoki, M.; Vogt, P. Phosphatidylinositol 3-kinase signaling mediates angiogenesis and expression of vascular endothelial growth factor in endothelial cells. Proc. Natl. Acad. Sci. USA 2000, 97, 1749–1753. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clere, N.; To, K.H.T.; Legeay, S.; Bertrand, S.; Helesbeux, J.J.; Duval, O.; Faure, S. Pro-Angiogenic Effects of Low Dose Ethoxidine in a Murine Model of Ischemic Hindlimb: Correlation between Ethoxidine Levels and Increased Activation of the Nitric Oxide Pathway. Molecules 2017, 22, 627. https://doi.org/10.3390/molecules22040627
Clere N, To KHT, Legeay S, Bertrand S, Helesbeux JJ, Duval O, Faure S. Pro-Angiogenic Effects of Low Dose Ethoxidine in a Murine Model of Ischemic Hindlimb: Correlation between Ethoxidine Levels and Increased Activation of the Nitric Oxide Pathway. Molecules. 2017; 22(4):627. https://doi.org/10.3390/molecules22040627
Chicago/Turabian StyleClere, Nicolas, Kim Hung Thien To, Samuel Legeay, Samuel Bertrand, Jean Jacques Helesbeux, Olivier Duval, and Sébastien Faure. 2017. "Pro-Angiogenic Effects of Low Dose Ethoxidine in a Murine Model of Ischemic Hindlimb: Correlation between Ethoxidine Levels and Increased Activation of the Nitric Oxide Pathway" Molecules 22, no. 4: 627. https://doi.org/10.3390/molecules22040627
APA StyleClere, N., To, K. H. T., Legeay, S., Bertrand, S., Helesbeux, J. J., Duval, O., & Faure, S. (2017). Pro-Angiogenic Effects of Low Dose Ethoxidine in a Murine Model of Ischemic Hindlimb: Correlation between Ethoxidine Levels and Increased Activation of the Nitric Oxide Pathway. Molecules, 22(4), 627. https://doi.org/10.3390/molecules22040627