Molecular Insights Reveal Psy1, SGR, and SlMYB12 Genes are Associated with Diverse Fruit Color Pigments in Tomato (Solanum lycopersicum L.)

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Genomic DNA Extraction
2.2. PCR Amplification
2.3. RNA Isolation and cDNA Synthesis
2.4. Cloning and Sequencing
2.5. Detection of SNPs Using Hybprobe
2.6. HPLC Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Mutation of the RIN Gene and Its Effect on Tomato Color
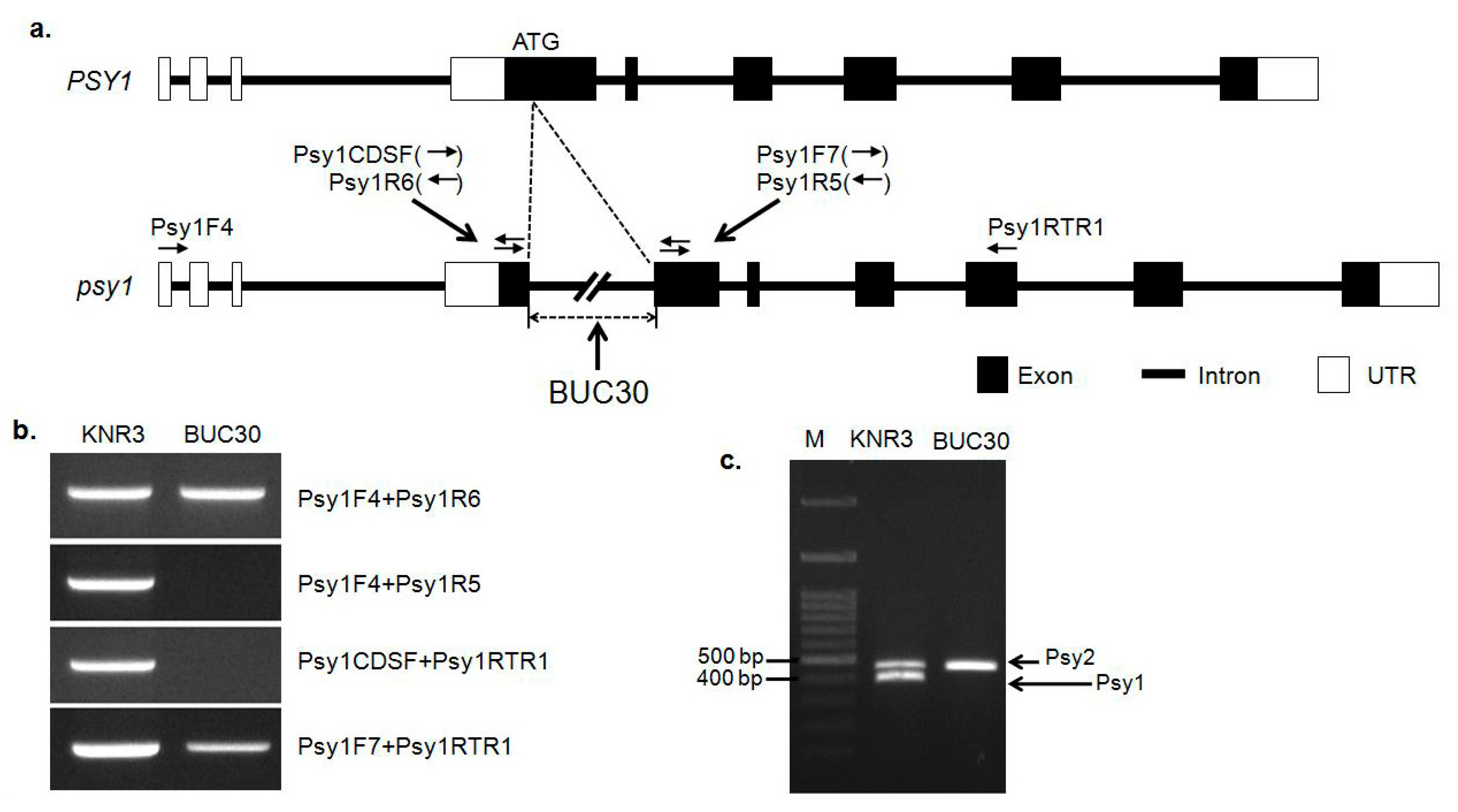
3.2. Mutation of the Psy1 Gene and Its Effect on Tomato Color
3.3. Mutation in the SGR Gene and Its Effect on Tomato Color
3.4. Mutation of the SlMYB12 Gene and Its Effect on Tomato Fruit Color
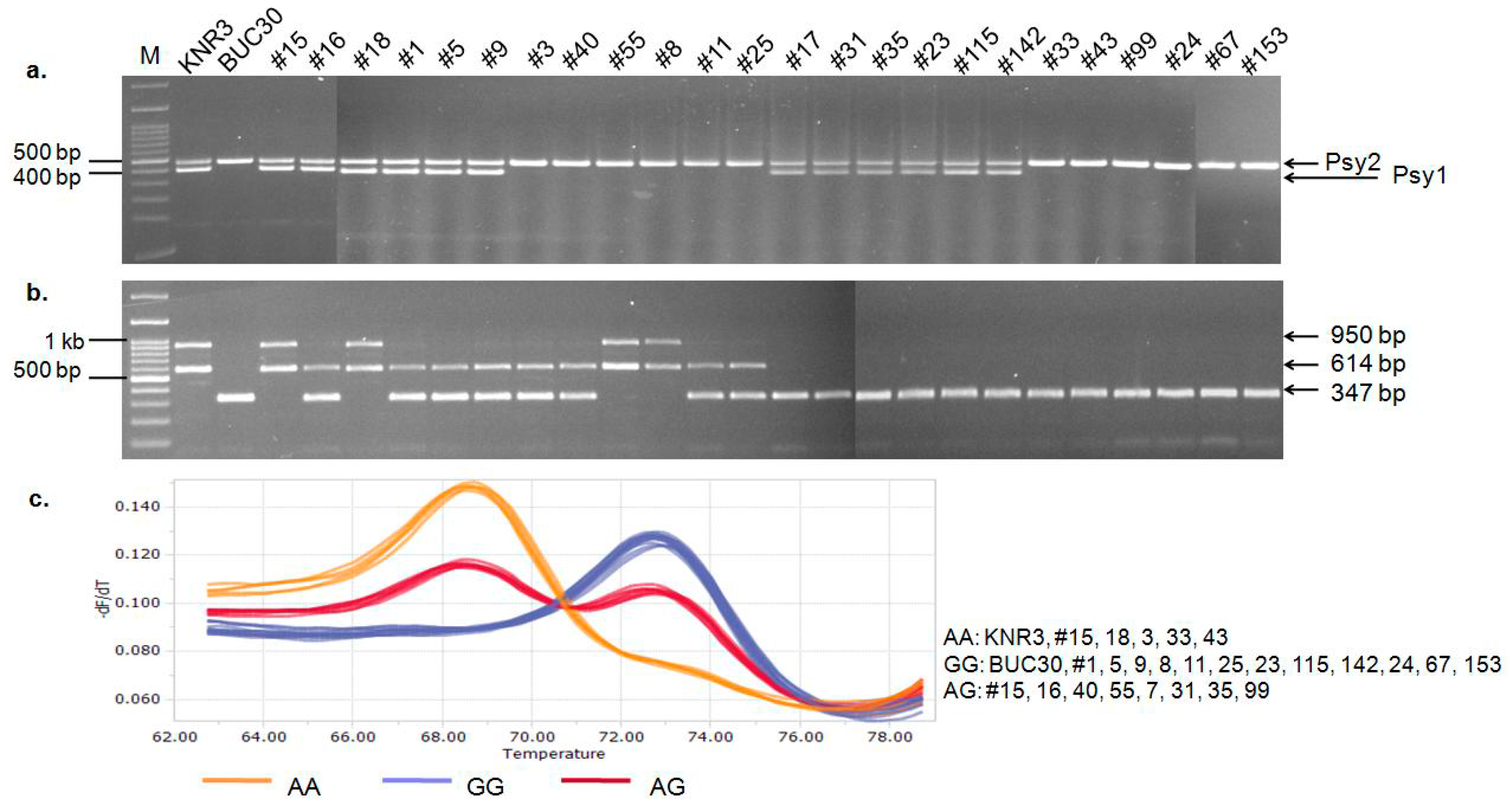
3.5. Characterization of the Psy1, SGR, and SlMYB12 Mutations in the F2Segregants
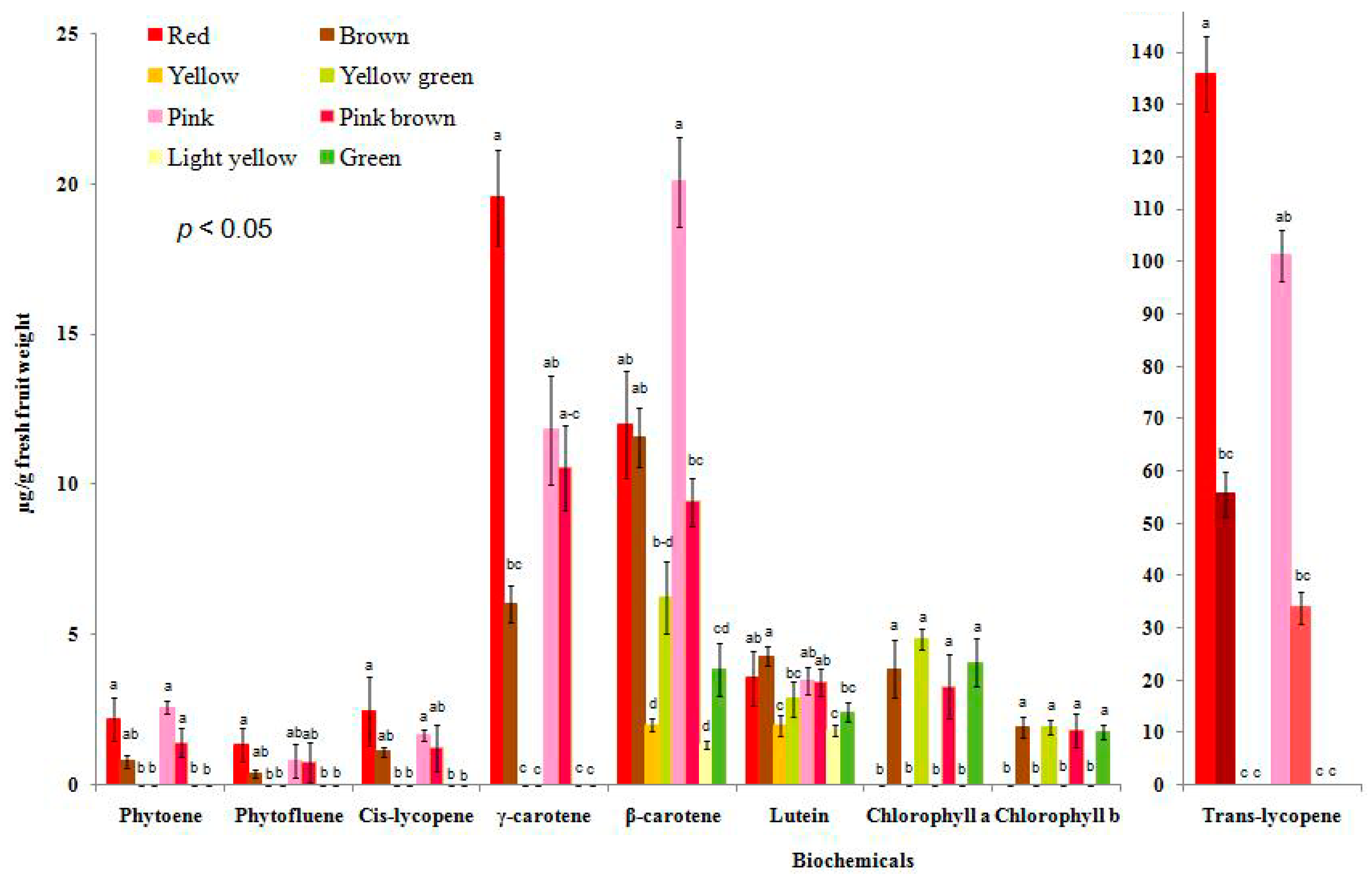
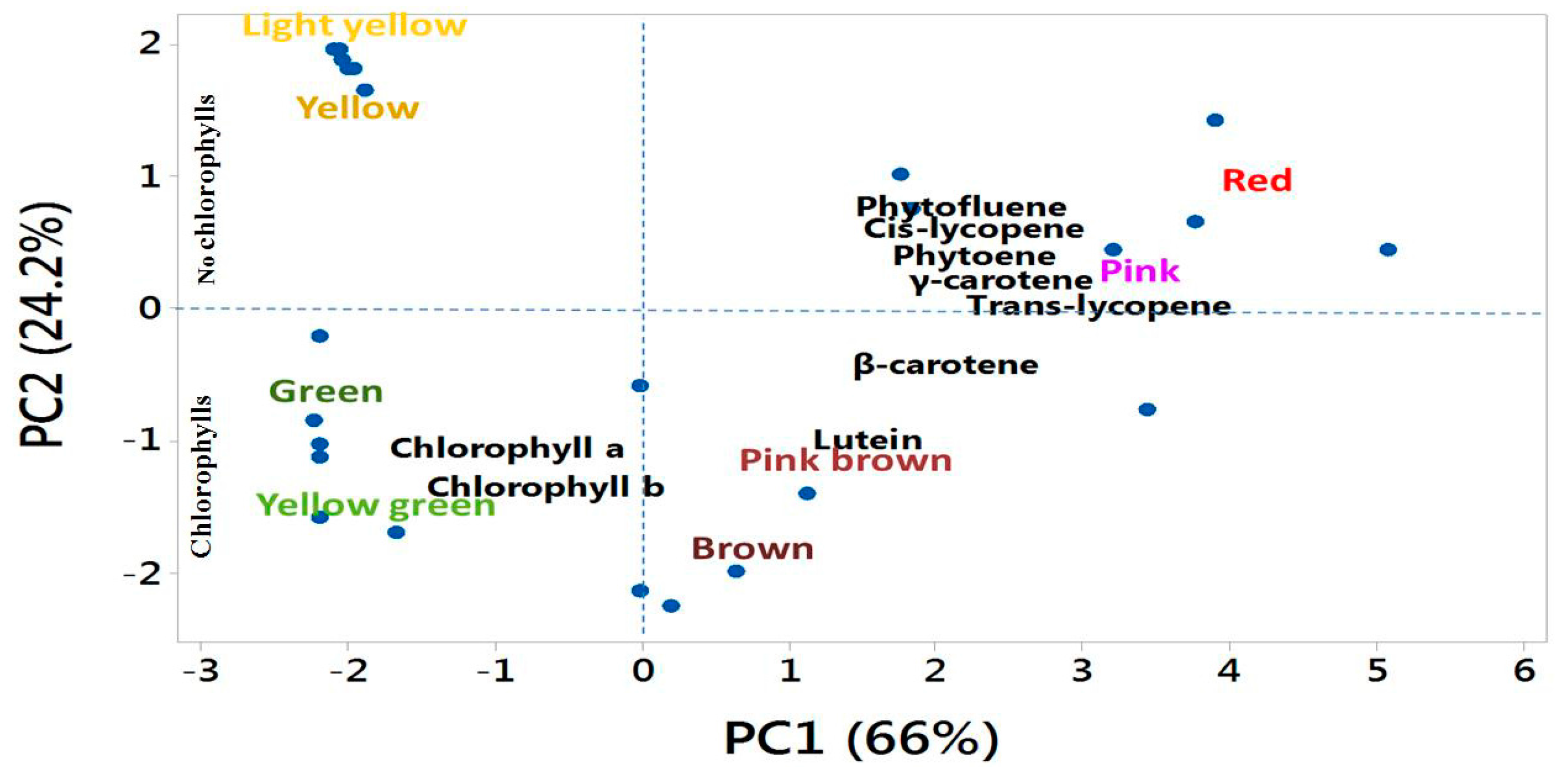
3.6. Association among Biochemical Accumulation, Gene Mutation, and Different Fruit Colors
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meissner, R.; Jacobson, Y.; Melamed, S.; Levyatuv, S.; Shalev, G.; Ashri, A.; Elkind, Y.; Levy, A. A new model System for tomato genetics. Plant J. 1997, 12, 1465–1472. [Google Scholar] [CrossRef]
- Ballester, A.R.; Molthoff, J.; de Vos, R.; Hekkert, B.; Orzaez, D.; Fernández-Moreno, J.P.; Tripodi, P.; Grandillo, S.; Martin, C.; Heldens, J.; et al. Biochemical and molecular analysis of pink tomatoes: Deregulated expression of the gene encoding transcription factor SlMYB12 leads to pink tomato fruit colour. Plant Physiol. 2010, 152, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, E.; Sitrit, Y.; Bar, E.; Azulay, Y.; Ibdah, M.; Meir, A.; Yosef, E.; Zamir, D.; Tadmor, Y. Not just colours: Carotenoid degradation as a link between pigmentation and aroma in tomato and watermelon fruit. Trends. Food Sci. Technol. 2005, 16, 407–415. [Google Scholar] [CrossRef]
- Hunt, G.M.; Baker, E.A. Phenolic constituents of tomato fruit cuticles. Phytochemistry 1980, 19, 1415–1419. [Google Scholar] [CrossRef]
- Harborne, J.B. Nature, distribution and function of plant flavonoids. Prog. Clin. Biol. Res. 1986, 213, 15–24. [Google Scholar] [PubMed]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Arts, I.C.W.; Hollman, P.C.H. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81, 317S–325S. [Google Scholar] [PubMed]
- Hooper, L.; Cassidy, A. A review of the health care potential of bioactive compounds. J. Sci. Food Agric. 2006, 86, 1805–1813. [Google Scholar] [CrossRef]
- Lotito, S.B.; Frei, B. Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: Cause, consequence, or epiphenomenon? Free Radic. Biol. Med. 2006, 41, 1727–1746. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G. Plant polyphenols: How to translate their in vitro antioxidant actions to in vivo conditions. IUBMB Life 2007, 59, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Tapas, A.R.; Sakarkar, D.M.; Kakde, R.B. Flavonoids as nutraceuticals: A review. Trop. J. Pharm. Res. 2008, 7, 1089–1099. [Google Scholar] [CrossRef]
- Davies, K.M. An introduction to plant pigments in biology and commerce. In Plant Pigments and Their Manipulation. Annual Plant Reviews; Davies, K.M., Ed.; Blackwell Publishing: Oxford, FL, USA; CRC Press: Boca Raton, FL, USA, 2004; Volume 14, pp. 1–22. [Google Scholar]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; De Groot, J.; Van Beek, T.A.; Vervoort, J.; De Vos, C.H.R. A liquid chromatography-mass spectrometry-based metabolome database for tomato. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Nakamura, Y.; Ogata, Y.; Tanaka, K.; Sakurai, N.; Suda, K.; Suzuki, T.; Suzuki, H.; Okazaki, K.; Kitayama, M.; et al. Metabolite annotations based on the integration of mass spectral information. Plant J. 2008, 54, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.J.; Veerappan, K.; Hwang, I.; Goswami, G.; Chung, M.Y.; Nou, I.S. New SNPs and InDel Variations in SlMYB12 Associated with Regulation of Pink Colour in Tomato. Trop. Plant Biol. 2017, 10, 126–133. [Google Scholar] [CrossRef]
- Liu, L.; Shao, Z.; Zhang, M.; Wang, Q. Regulation of carotenoid metabolism in tomato. Mol. Plant 2015, 8, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Nath, P.; Bouzayen, M.; Mattoo, A.; Pech, J.C. Fruit Ripening: Physiology, Signalling and Genomics; CAB International: Wallingford, UK, 2014. [Google Scholar]
- Gady, A.L.; Vriezen, W.H.; Van de Wal, M.H.; Huang, P.; Bovy, A.G.; Visser, R.G.; Bachem, C.W. Induced point mutations in the phytoene synthase 1 gene cause differences in carotenoid content during tomato fruit ripening. Mol. Breed. 2012, 29, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.; Gu, Q.; Tian, P.; Xiao, L.; Cao, H.; Yang, W. A chimeric transcript containing Psy1 and a potential mRNA is associated with yellow flesh colour in tomato accession PI 114490. Planta 2014, 240, 1011–1021. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of beta-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, T.; Ohad, I.; Beyer, P.; Hirschberg, J. Analysis in vitro of the enzyme CRTISO establishes a poly-cis-carotenoid biosynthesis pathway in plants. Plant Physiol. 2004, 136, 4246–4255. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Ghisla, S.; Hirschberg, J.; Mann, V.; Beyer, P. Plant carotene cis-trans isomerase CRTISO: A new member of the FAD (RED)-dependent flavoproteins catalyzing non-redox reactions. J. Biol. Chem. 2011, 286, 8666–8676. [Google Scholar] [CrossRef] [PubMed]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to beta-carotene formation in plant chromoplasts discovered by map-based cloning of beta and old-gold colour mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Kim, Y.; Han, J.; Nou, I.S. Orange colour is associated with CYC-B expression in tomato fleshy fruit. Mol. Breed. 2016, 36, 1–10. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Goldschmidt, E.E.; John, I.; Rodoni, S.; Matile, P.; Grierson, D. Altered patterns of senescence and ripening in gf, a stay-green mutant of tomato (Lycopersiconesculentum Mill.). J. Exp. Bot. 1999, 50, 1115–1122. [Google Scholar] [CrossRef]
- Barry, C.S.; Mc Quinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-flesh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol. 2008, 147, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S. Stay-green regulates chlorophyll and chlorophyll-binding protein degradation during senescence. Trends Plant Sci. 2009, 14, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Nakano, T.; Shima, Y.; Ito, Y. A large-scale identification of direct targets of the tomato MADS box transcription factor RIPENING INHIBITOR reveals the regulation of fruit ripening. Plant Cell 2013, 25, 371–386. [Google Scholar] [CrossRef] [PubMed]
- Tomato Genomic Database. Available online: http://solgenomics.net (accessed on 26 October 2016).
- Yoo, H.J.; Park, W.J.; Lee, G.M.; Oh, C.S.; Yeam, I.; Won, D.C.; Kim, C.K.; Lee, J.M. Inferring the genetic determinants of fruit colors in tomato by carotenoid profiling. Molecules 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.W.; Tomes, M.L. Ripening inhibitor a gene with multiple effects on ripening. Rep. Tomato Genet. Coop. 1968, 18, 36–37. [Google Scholar]
- Sink, K.C.; Herner, R.C.; Knowlton, L.L. Chlorophyll and carotenoids of the “rin” tomato mutant. Can. J. Bot. 1974, 52, 1657–1660. [Google Scholar] [CrossRef]
- McGlasson, W.B.; Last, J.H.; Shaw, K.J.; Meldrum, S.K. Influence of the non-ripening mutants rin and nor on the aroma of tomato fruit. Hortic. Sci. 1987, 22, 632–634. [Google Scholar]
- Tang, W.; Zheng, Y.; Dong, J.; Yu, J.; Yue, J.; Liu, F.; Guo, X.; Huang, S.; Wisniewski, M.; Sun, J.; et al. Comprehensive Transcriptome Profiling Reveals Long Noncoding RNA Expression and Alternative Splicing Regulation during Fruit Development and Ripening in Kiwifruit (Actinidia chinensis). Front. Plant Sci. 2016, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Suárez, C.; Atienza, S.G.; Pistón, F. Allelic variation, alternative splicing and expression analysis of Psy1 gene in Hordeum chilense Roem. et Schult. PLoS ONE 2011, 6, e19885. [Google Scholar] [CrossRef] [PubMed]
- Veerappan, K.; Jung, H.J.; Hwang, I.; Kang, H.K.; Chung, M.Y.; Nou, I.S. Sequence variation in SlMYB12 is associated with fruit peel colour in pink tomato cultivars. Hortic. Environ. Biotechnol. 2016, 57, 274–279. [Google Scholar] [CrossRef]
- Lin, T.; Zhu, G.; Zheng, Z.; Xu, X.; Yu, Q.; Zheng, Z.; Zhang, Z.; Lun, Y.; Li, S.; Wang, X.; et al. Genomic analyses provide insights into the history of tomato breeding. Nat. Genet. 2014, 46, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Fray, R.G.; Grierson, D. Identification and genetic-analysis of normal and mutant phytoene synthase genes of tomato by sequencing, complementation and co-suppression. Plant Mol. Biol. 1993, 22, 589–602. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not Available. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruit Phenotypes | Genetic Make up | ||
|---|---|---|---|
| Psy1 | SGR | SlMYB12 | |
| Red | N | N | N |
| Yellow | M | N | N |
| Brown | N | M | N |
| Pink | N | N | M |
| Light yellow | M | N | M |
| Pink brown | N | M | M |
| Yellow green | M | M | N |
| Green | M | M | M |
| Gene | Primer Name | Sequence (5′-3′) | Fruit Phenotypes with Exact PCR Amplicon Size (bp)/SNP at bp | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Red | Yellow | Brown | Pink | Light Yellow | Pink Brown | Yellow Green | Green | |||
| Psy1 | Psy1CDSF | ATGTCTGTTGCCTTGTTATGGGTTGTTTC | 396 | 468 | 396 | 396 | 468 | 396 | 468 | 468 |
| Psy1R5 | TGCATACTCTGCACATACTTCACCAC | |||||||||
| SGR | SGR4-aR2 | AGGATCTGACACAGGACCAATAACA | A at 1679 | A at 1679 | G at 1679 | A at 1679 | A at 1679 | G at 1679 | G at 1679 | G at 1679 |
| SGR4-aF2 | GAAGATGTCACTTCATGTCCATTG | |||||||||
| SlMYB12 | aF1 | GTGACGAACAACCGACCTAGAATAA | 950 (aF1 + aR6) 614 (aF1 + aR5) | 950 (aF1 + aR6) 614 (aF1 + aR5) | 950 (aF1 + aR6) 614 (aF1 + aR5) | 347 (aF1 + aR6) | 347 (aF1 + aR6) | 347 (aF1 + aR6) | 950 (aF1 + aR6) 614 (aF1 + aR5) | 347 (aF1 + aR6) |
| aR6 | GCGGACAAAGTTAATTGGTCACTCA | |||||||||
| aR5 | ATTCTAGCGTTATCAGTCGGCATACA | |||||||||
| Variable | PC1 | PC2 |
|---|---|---|
| Phytoene | 0.385 | 0.002 |
| Phytofluene | 0.374 | 0.044 |
| cis-lycopene | 0.396 | −0.002 |
| trans-lycopene | 0.408 | 0.000 |
| γ-carotene | 0.401 | −0.010 |
| β-carotene | 0.353 | −0.172 |
| Lutein | 0.267 | −0.418 |
| Chlorophyll a | −0.140 | −0.622 |
| Chlorophyll b | −0.115 | −0.638 |
| % variation explained | 0.660 | 0.242 |
| p-value | <0.01 | <0.01 |
| Fruit Colour | Mean PC Sores | |
| Red | 3.59 a | 0.86 a |
| Yellow | −1.97 cd | 1.80 a |
| Brown | 0.65 bc | −1.89 b |
| Pink | 2.90 ab | 0.69 a |
| Light yellow | −2.051 cd | 1.88 a |
| Pink brown | 1.12 ab | −1.17 b |
| Yellow green | −2.04 cd | −1.38 b |
| Green | −2.20 d | −0.79 b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.-I.; Hwang, I.; Goswami, G.; Jung, H.-J.; Nath, U.K.; Yoo, H.-J.; Lee, J.M.; Nou, I.S. Molecular Insights Reveal Psy1, SGR, and SlMYB12 Genes are Associated with Diverse Fruit Color Pigments in Tomato (Solanum lycopersicum L.). Molecules 2017, 22, 2180. https://doi.org/10.3390/molecules22122180
Kang S-I, Hwang I, Goswami G, Jung H-J, Nath UK, Yoo H-J, Lee JM, Nou IS. Molecular Insights Reveal Psy1, SGR, and SlMYB12 Genes are Associated with Diverse Fruit Color Pigments in Tomato (Solanum lycopersicum L.). Molecules. 2017; 22(12):2180. https://doi.org/10.3390/molecules22122180
Chicago/Turabian StyleKang, Song-I., Indeok Hwang, Gayatri Goswami, Hee-Jeong Jung, Ujjal Kumar Nath, Hee-Ju Yoo, Je Min Lee, and Ill Sup Nou. 2017. "Molecular Insights Reveal Psy1, SGR, and SlMYB12 Genes are Associated with Diverse Fruit Color Pigments in Tomato (Solanum lycopersicum L.)" Molecules 22, no. 12: 2180. https://doi.org/10.3390/molecules22122180
APA StyleKang, S.-I., Hwang, I., Goswami, G., Jung, H.-J., Nath, U. K., Yoo, H.-J., Lee, J. M., & Nou, I. S. (2017). Molecular Insights Reveal Psy1, SGR, and SlMYB12 Genes are Associated with Diverse Fruit Color Pigments in Tomato (Solanum lycopersicum L.). Molecules, 22(12), 2180. https://doi.org/10.3390/molecules22122180