Anti-Inflammatory Potential of 1-Nitro-2-Phenylethylene

 ,
, 

Abstract
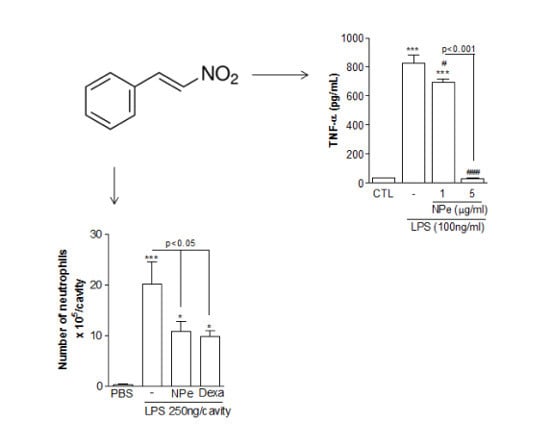
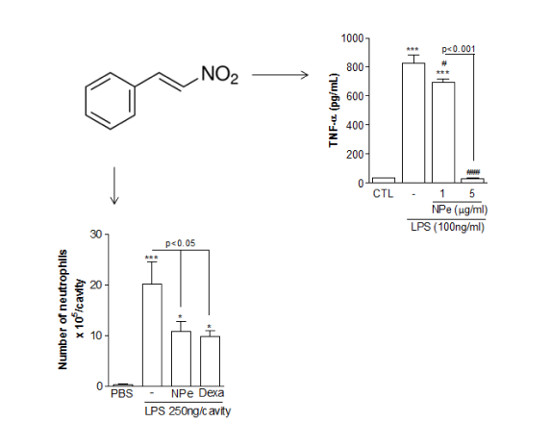
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
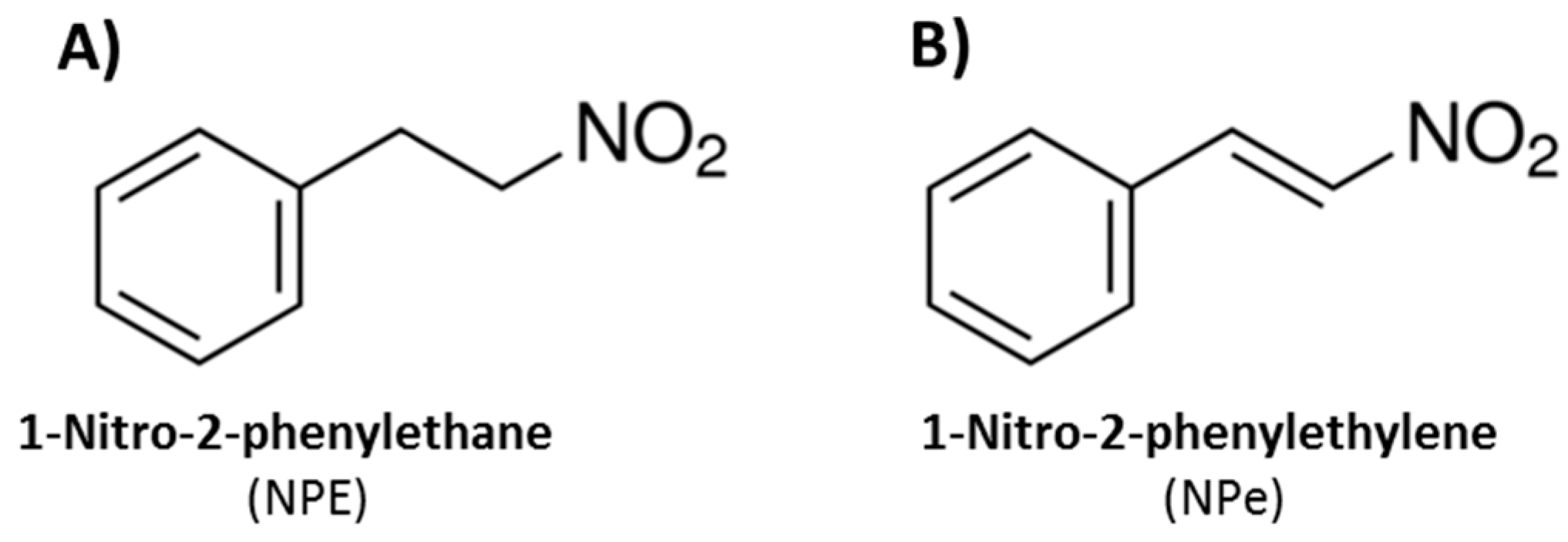
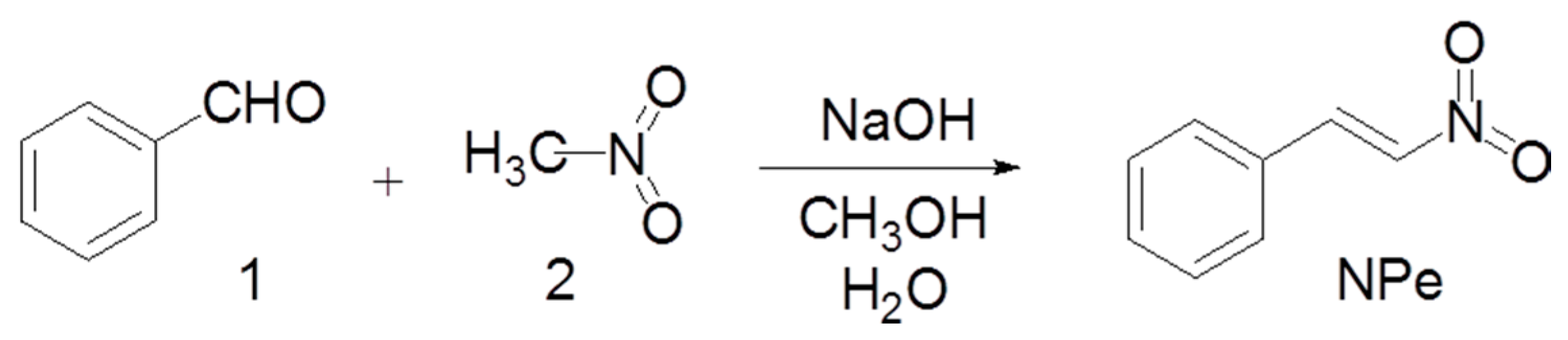
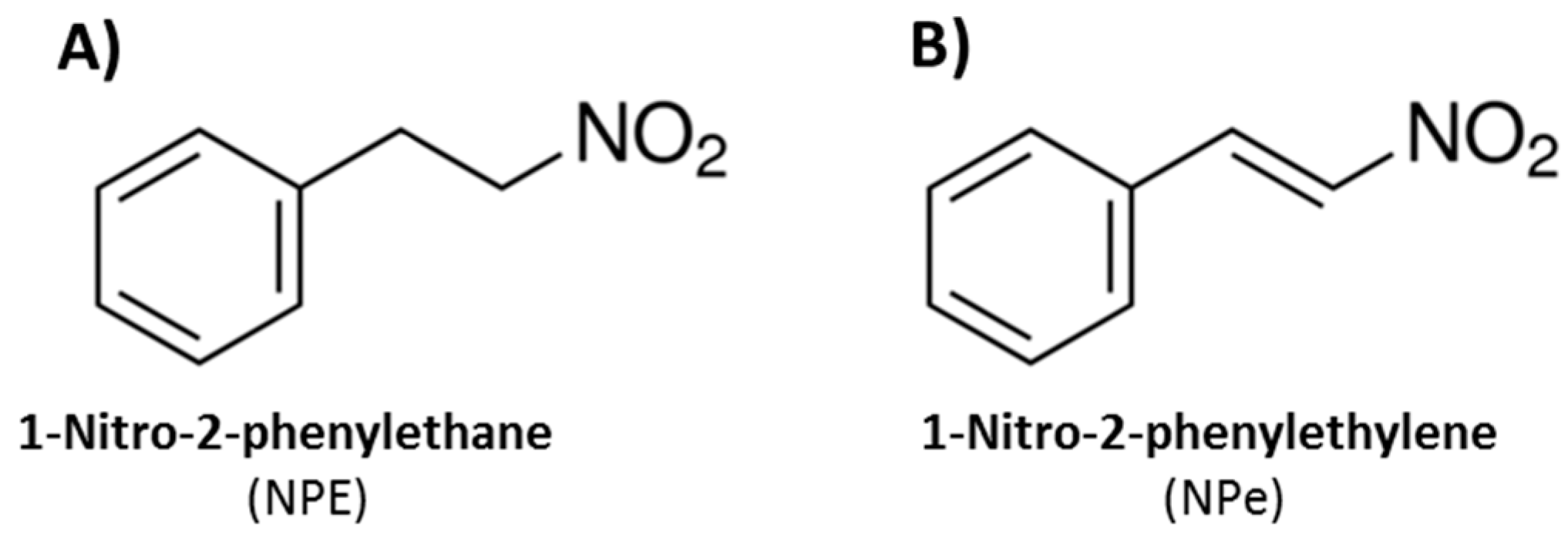
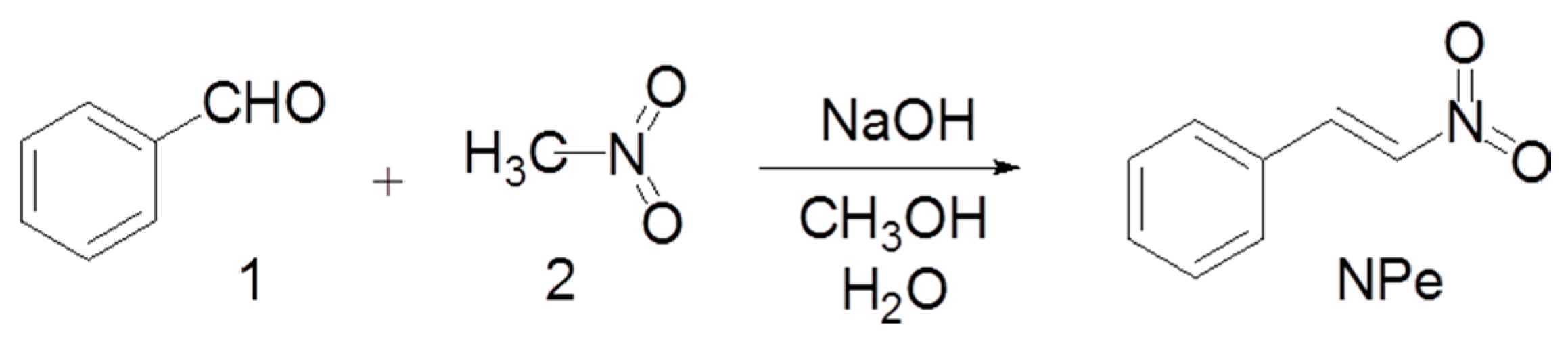
2.1. Synthesis of NPe
2.2. Mice and Ethics
2.3. Drugs, Reagents and Antibodies
2.4. Cell Culture and In Vitro Assays
2.5. LPS-Induced Pleurisy Model
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
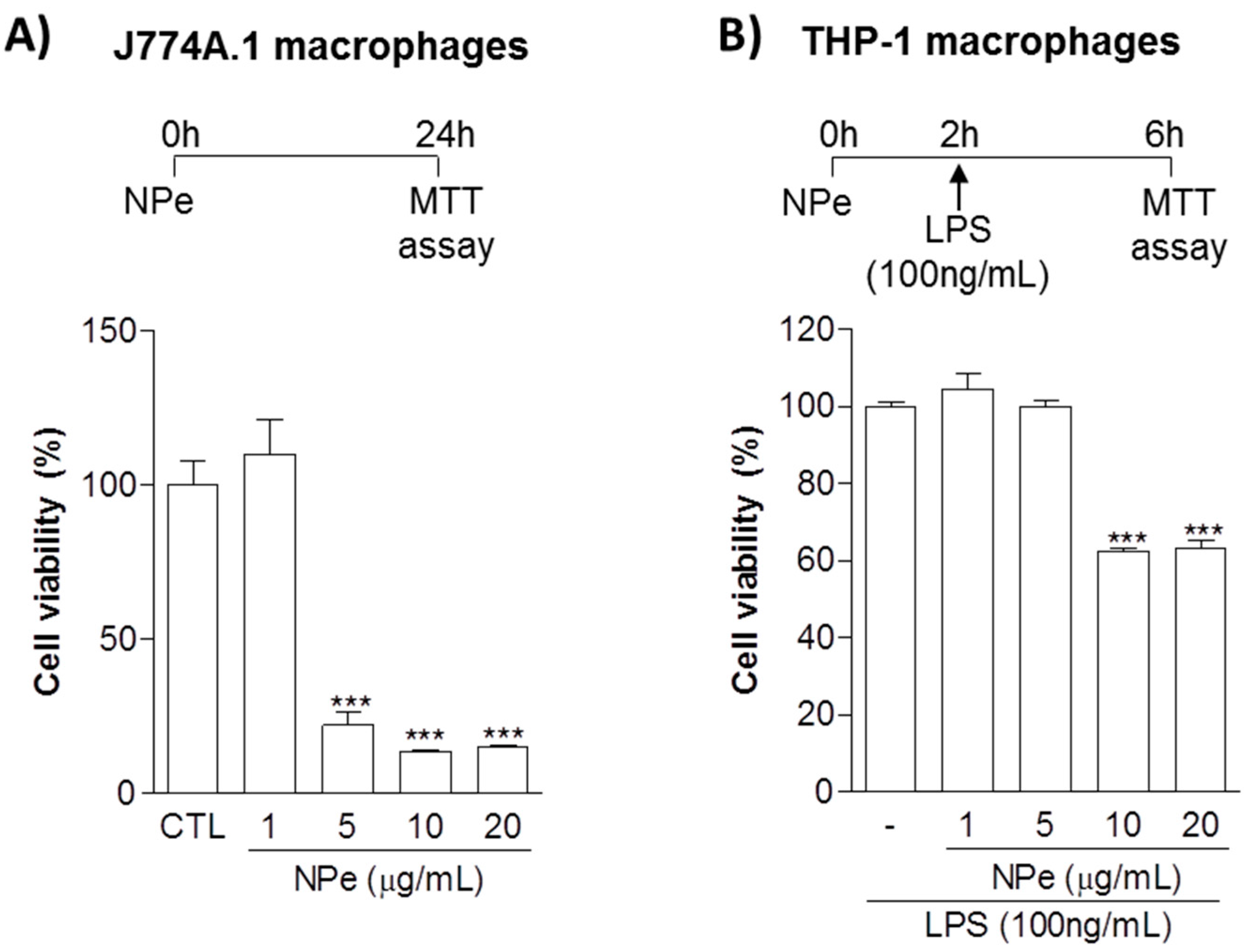
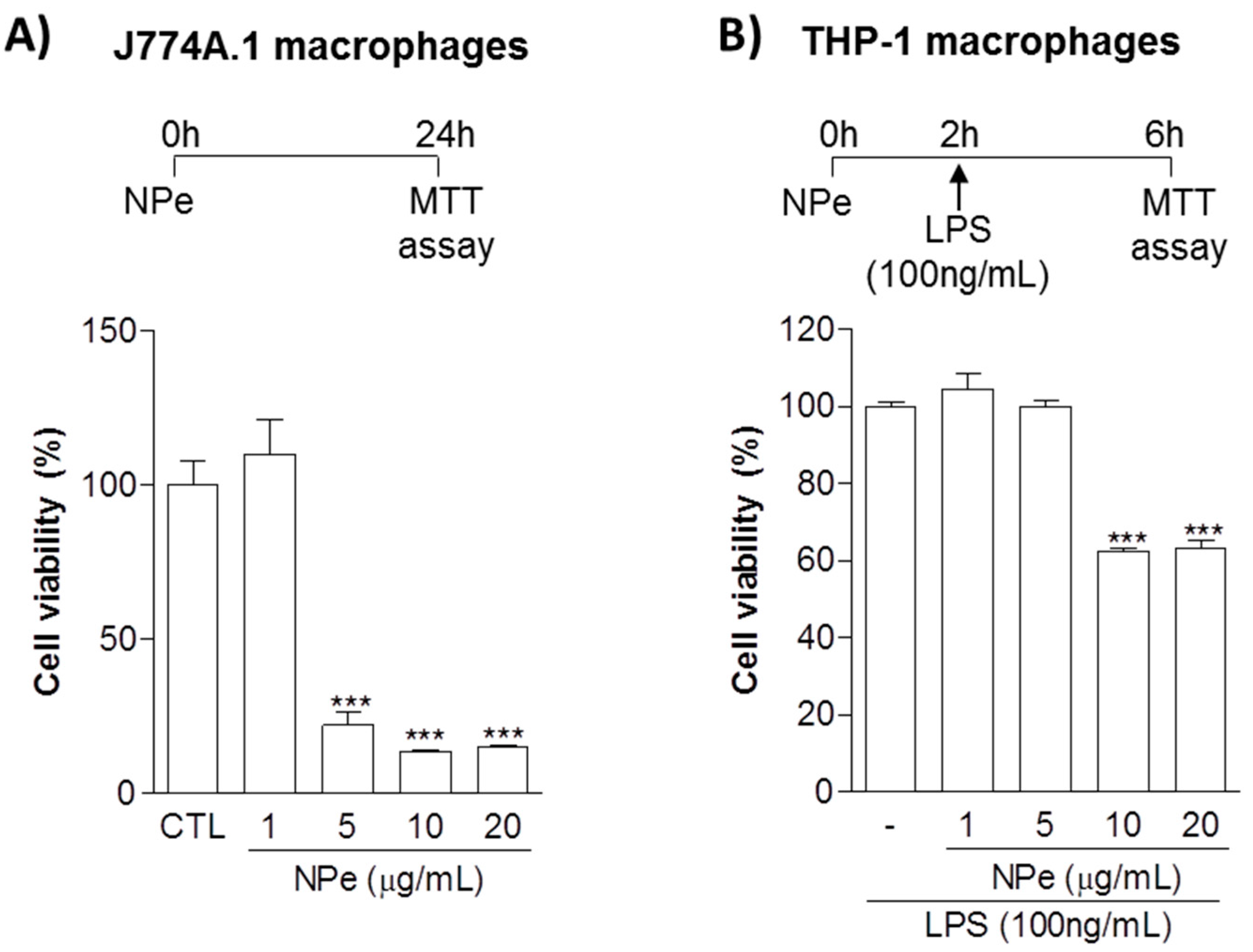
3.1. Definition of Toxic and Non-Toxic Doses of NPe on Murine and Human Macrophages
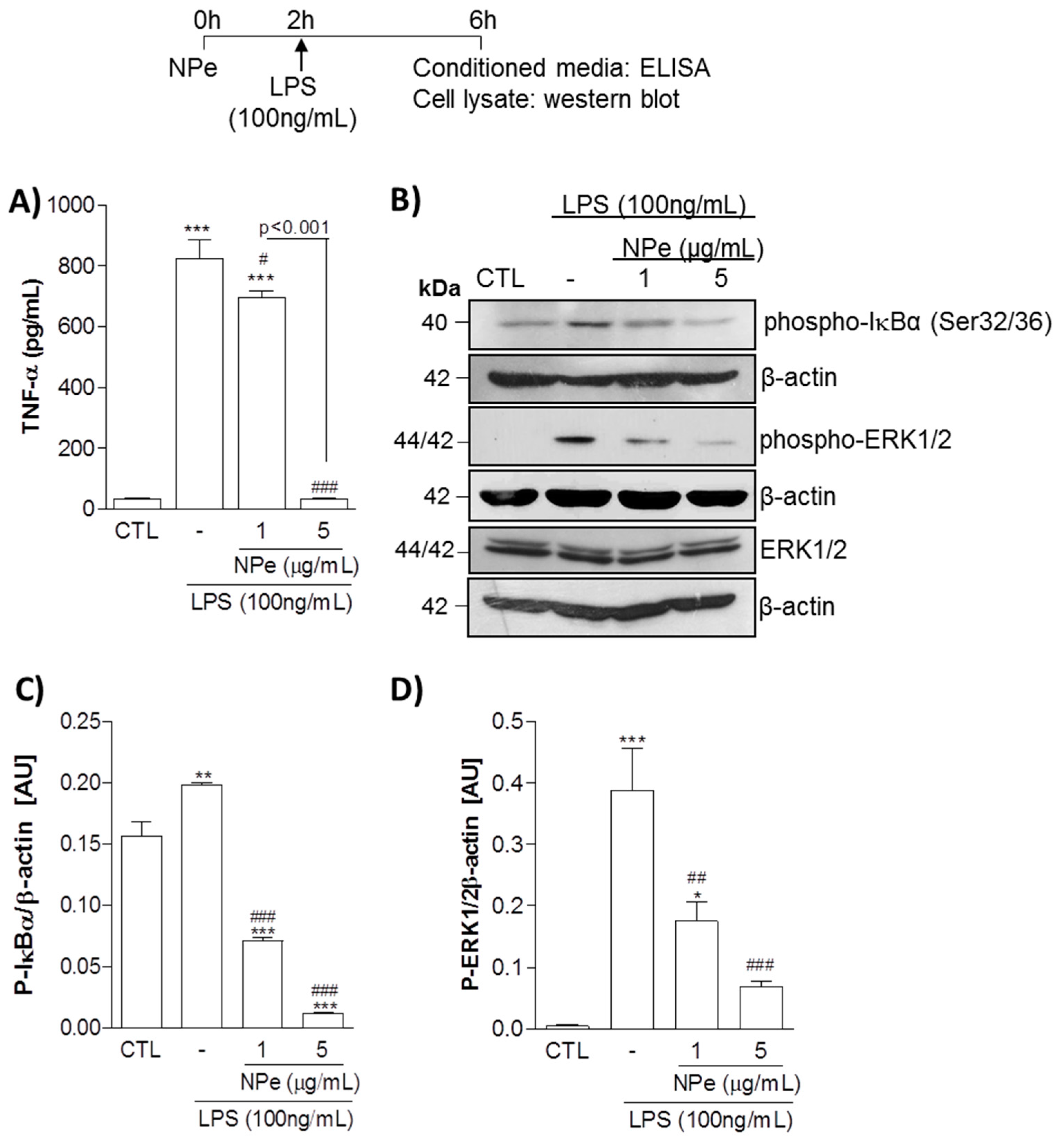
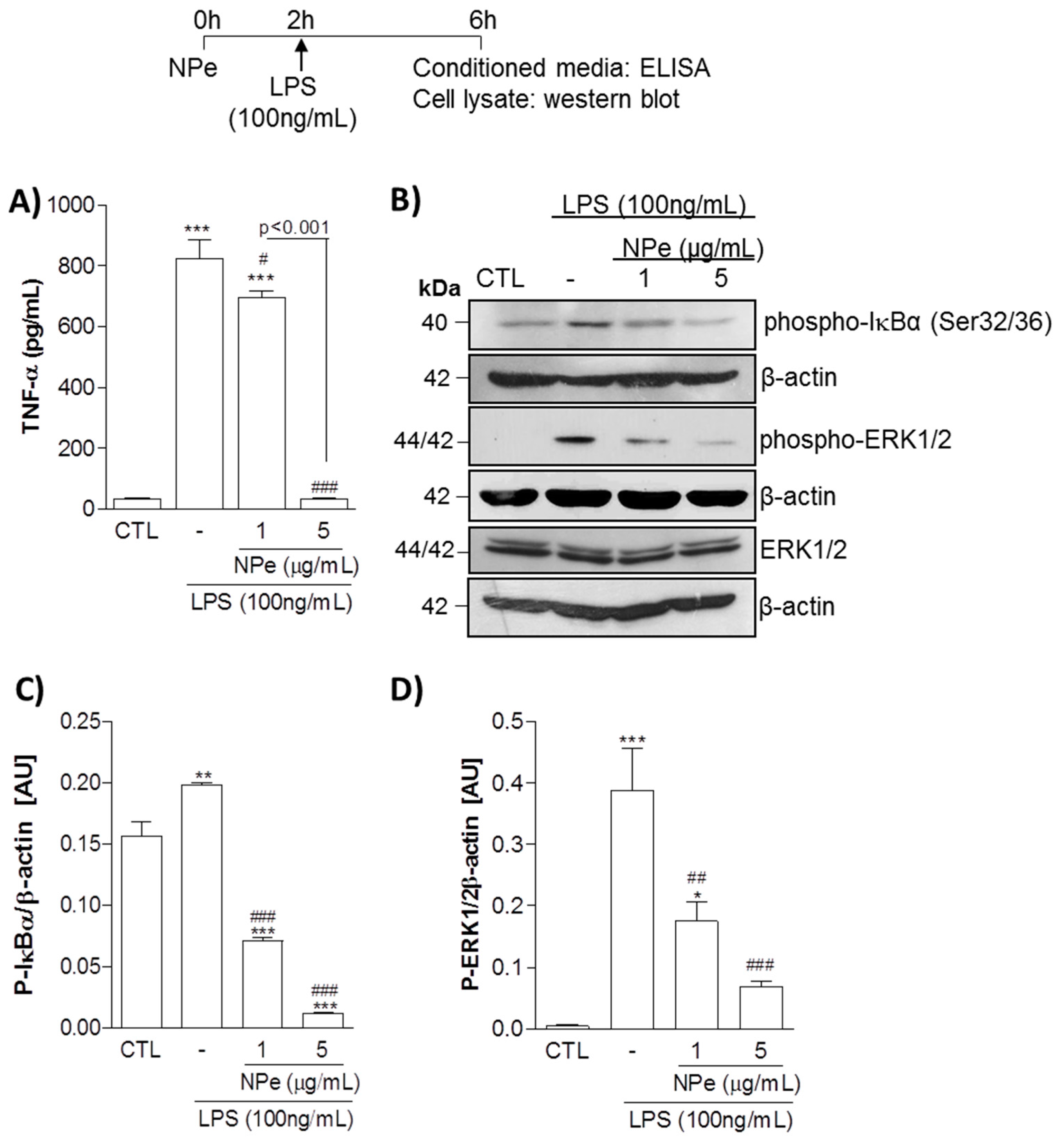
3.2. Pre-Treatment of THP-1 Differentiated Macrophages with Non-Toxic Doses of NPe Has Anti-Inflammatory Effects
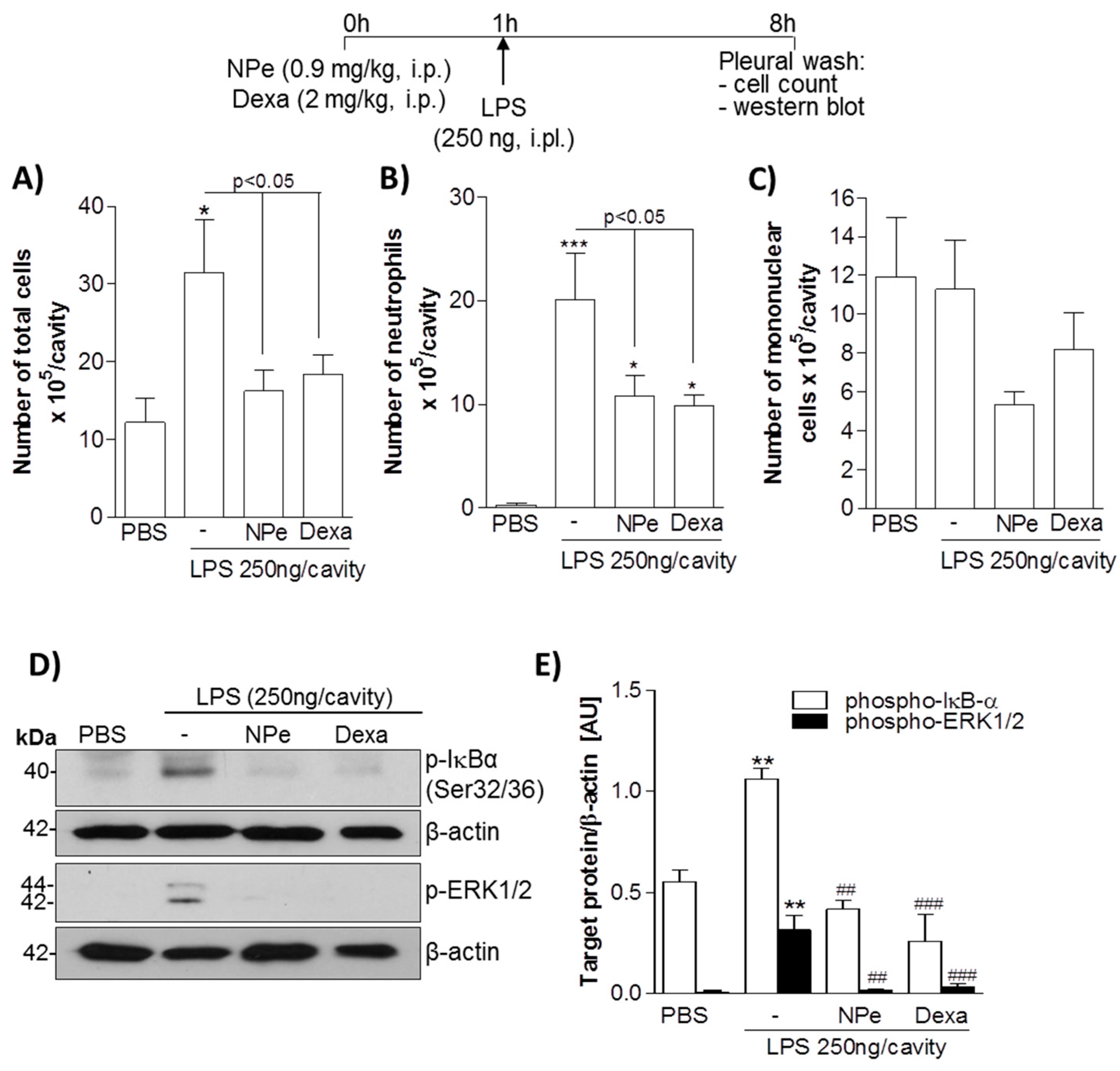
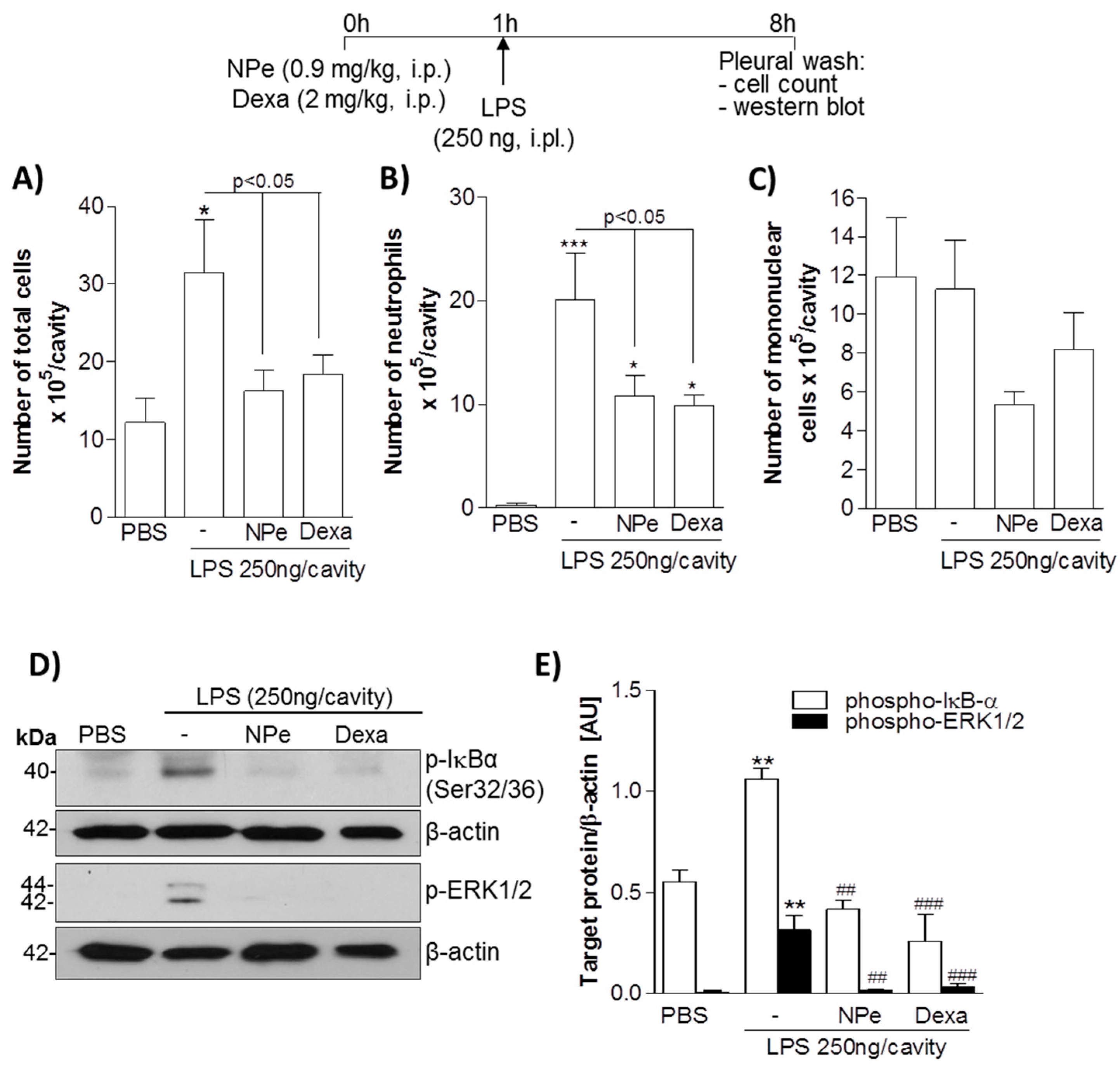
3.3. Pre-Treatment of LPS-Inflamed Mice with NPe Reduces Neutrophil Accumulation and Inflammatory Markers
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pimentel, A.A.M.P. Cultivo de Plantas Medicinais na Amazonia; FCAP: Belém, Brazil, 1994. [Google Scholar]
- Vieira, L.S. Manual da Medicina Popular: A Fitoterapia da Amazonia; FCAP: Belém, Brazil, 1991. [Google Scholar]
- Estrella, E. Plantas Medicinales Amazonicas: Realidad y Perspectivas; GEF/PNUD: Lima, Peru, 1995. [Google Scholar]
- Lahlou, S.; Magalhaes, P.J.; de Siqueira, R.J.; Figueiredo, A.F.; Interaminense, L.F.; Maia, J.G.; Sousa, P.J. Cardiovascular effects of the essential oil of Aniba canelilla bark in normotensive rats. J. Cardiovasc. Pharmacol. 2005, 46, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.K.; Sousa, P.J.; Andrade, E.H.; Maia, J.G. Antioxidant capacity and cytotoxicity of essential oil and methanol extract of Aniba canelilla (H.B.K.) Mez. J. Agric. Food Chem. 2007, 55, 9422–9426. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, O.R.; Magalhães, M.T. Essential oil of the bark and wood of Aniba canelilla. Perf. Essent. Oil Rec. 1960, 51, 69–70. [Google Scholar]
- Gottlieb, O.R.; Magalhães, M.T. Occurrence of 1-nitro-2-phenylethane in Ocotea pretiosa and Aniba canelilla. J. Org. Chem. 1959, 24, 2070–2071. [Google Scholar] [CrossRef]
- Taveira, F.S.N.; de Lima, W.N.; Andrade, E.H.A.; Maia, J.G.S. Seasonal essential oil variation of Aniba canelilla. Biochem. Syst. Ecol. 2003, 31, 69–75. [Google Scholar] [CrossRef]
- De Lima, A.B.; Santana, M.B.; Cardoso, A.S.; da Silva, J.K.; Maia, J.G.; Carvalho, J.C.; Sousa, P.J. Antinociceptive activity of 1-nitro-2-phenylethane, the main component of Aniba canelilla essential oil. Phytomedicine 2009, 16, 555–559. [Google Scholar] [CrossRef] [PubMed]
- De Siqueira, R.J.; Macedo, F.I.; Interaminense Lde, F.; Duarte, G.P.; Magalhães, P.J.; Brito, T.S.; da Silva, J.K.; Maia, J.G.; Sousa, P.J.; Leal-Cardoso, J.H.; et al. 1-Nitro-2-phenylethane, the main constituent of the essential oil of Aniba canelilla, elicits a vago-vagal bradycardiac and depressor reflex in normotensive rats. Eur. J. Pharmacol. 2010, 638, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Interaminense Lde, F.; de Siqueira, R.J.; Xavier, F.E.; Duarte, G.P.; Magalhães, P.J.; da Silva, J.K.; Maia, J.G.; Sousa, P.J.; Leal-Cardoso, J.H.; Lahlou, S. Cardiovascular effects of 1-nitro-2-phenylethane, the main constituent of the essential oil of Aniba canelilla, in spontaneously hypertensive rats. Fundam. Clin. Pharmacol. 2011, 25, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Vale, J.K.; Lima, A.B.; Pinheiro, B.G.; Cardoso, A.S.; Silva, J.K.; Maia, J.G.; de Sousa, G.E.; da Silva, A.B.; Sousa, P.J.; Borges, R.S. Evaluation and theoretical study on the anti-inflammatory mechanism of 1-nitro-2-phenylethane. Planta Med. 2013, 79, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Arruda-Barbosa, L.; Rodrigues, K.M.; Souza-Neto, F.; Duarte, G.P.; Borges, R.S.; Magalhães, P.J.; Lahlou, S. Vasorelaxant effects of 1-nitro-2-phenylethene in rat isolated aortic rings. Vascul. Pharmacol. 2014, 63, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Song, Y.; Zhan, P.; Zhang, Q.; Liu, X. Conformational restriction: An effective tactic in ‘follow-on‘-based drug discovery. Future Med. Chem. 2014, 6, 885–901. [Google Scholar] [CrossRef] [PubMed]
- Chaires, J.B. Calorimetry and thermodynamics in drug design. Annu. Rev. Biophys. 2008, 37, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of Inflammation: What Controls Its Onset? Front. Immunol. 2016, 7, 160. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Ding, A. Nonresolving inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, A.L.; Sousa, L.P.; Lucas, C.D.; Rossi, A.G.; Pinho, V.; Teixeira, M.M. Resolution of inflammation: Mechanisms and opportunity for drug development. Pharmacol. Ther. 2013, 139, 189–212. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C.A., Jr. Decoding the patterns of self and nonself by the innate immune system. Science 2002, 296, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Yamamoto, Y.; Wang, Q.M. The IKK NF-kappa B system: A treasure trove for drug development. Nat. Rev. Drug Discov. 2004, 3, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional profiling of the LPS induced NF-kappaB response in macrophages. BMC Immunol. 2007, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.S., Jr. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [PubMed]
- Huxford, T.; Malek, S.; Ghosh, G. Structure and mechanism in NF-kappa B/I kappa B signaling. Cold Spring Harb. Symp. Quant. Biol. 1999, 64, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Bonizzi, G.; Karin, M. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004, 25, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Leung, T.H.; Baltimore, D. Genetic analysis of NF-kappaB/Rel transcription factors defines functional specificities. EMBO J. 2003, 22, 5530–5539. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, C.D.; Ceci, J.D.; Tsatsanis, C.; Kontoyiannis, D.; Stamatakis, K.; Lin, J.H.; Patriotis, C.; Jenkins, N.A.; Copeland, N.G.; Kollias, G.; et al. TNF-alpha induction by LPS is regulated posttranscriptionally via a Tpl2/ERK-dependent pathway. Cell 2000, 103, 1071–1083. [Google Scholar] [CrossRef]
- Vogel, A.I. Practical Organic Chemistry, 5th ed.; Longman Scientific and Technical: New York, NY, USA, 1989. [Google Scholar]
- Ford, P.W.; Narbut, M.R.; Belli, J.; Davidson, B.S. Synthesis and Structural Properties of the Benzopentathiepins Varacin and Isolissoclinotoxin A. J. Org. Chem. 1994, 59, 5955–5960. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S. The Rapid Synthesis of β-Nitrostyrenes under Microwave Irradiation without Solvent. Synth. Commun. 2009, 32, 3481–3486. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Sousa, L.P.; Lopes, F.; Silva, D.M.; Tavares, L.P.; Vieira, A.T.; Rezende, B.M.; Carmo, A.F.; Russo, R.C.; Garcia, C.C.; Bonjardim, C.A.; et al. PDE4 inhibition drives resolution of neutrophilic inflammation by inducing apoptosis in a PKA-PI3K/Akt-dependent and NF-kappaB-independent manner. J. Leukoc. Biol. 2010, 87, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Vago, J.P.; Nogueira, C.R.; Tavares, L.P.; Soriani, F.M.; Lopes, F.; Russo, R.C.; Pinho, V.; Teixeira, M.M.; Sousa, L.P. Annexin A1 modulates natural and glucocorticoid-induced resolution of inflammation by enhancing neutrophil apoptosis. J. Leukoc. Biol. 2012, 92, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.A.; Ribeiro, A.L.C.; Costa, B.R.C.; Vago, J.P.; Lima, K.M.; Carneiro, F.S.; Ortiz, M.M.O.; Lima, G.L.N.; Carmo, A.A.F.; Rocha, R.M.; et al. Plasmin and plasminogen induce macrophage reprogramming and regulate key steps of inflammation resolution via annexin A1. Blood 2017, 129, 2896–2907. [Google Scholar] [CrossRef] [PubMed]
- Villar, J.A.; Lima, F.T.; Veber, C.L.; Oliveira, A.R.; Calgarotto, A.K.; Marangoni, S.; da Silva, S.L. Synthesis and evaluation of nitrostyrene derivative compounds, new snake venom phospholipase A2 inhibitors. Toxicon 2008, 51, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Y.; Hsieh, P.W.; Wu, Y.C.; Wu, C.C. Synthesis and pharmacological evaluation of novel beta-nitrostyrene derivatives as tyrosine kinase inhibitors with potent antiplatelet activity. Biochem. Pharmacol. 2007, 74, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.W.; Chang, Y.T.; Chuang, W.Y.; Shih, H.C.; Chiang, S.Z.; Wu, C.C. The synthesis and biologic evaluation of anti-platelet and cytotoxic beta-nitrostyrenes. Bioorg. Med. Chem. 2010, 18, 7621–7627. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Varadarajan, S.; Munoz-Planillo, R.; Burberry, A.; Nakamura, Y.; Nunez, G. 3,4-methylenedioxy-beta-nitrostyrene inhibits NLRP3 inflammasome activation by blocking assembly of the inflammasome. J. Biol. Chem. 2014, 289, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.H.; Chang, F.R.; Wu, Y.C.; Kung, P.H.; Wu, C.C. 3,4-Methylenedioxy-beta-nitrostyrene inhibits adhesion and migration of human triple-negative breast cancer cells by suppressing beta1 integrin function and surface protein disulfide isomerase. Biochimie 2015, 110, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Hung, A.C.; Chen, Y.Y.; Chiu, Y.W.; Hsieh, P.W.; Lee, Y.C.; Su, Y.H.; Chang, P.C.; Hu, S.C.; Yuan, S.F. 3′-hydroxy-4′-methoxy-beta-methyl-beta-nitrostyrene inhibits tumorigenesis in colorectal cancer cells through ROS-mediated DNA damage and mitochondrial dysfunction. Oncotarget 2017, 8, 18106–18117. [Google Scholar] [PubMed]
- Rahmani-Nezhad, S.; Safavi, M.; Pordeli, M.; Ardestani, S.K.; Khosravani, L.; Pourshojaei, Y.; Mahdavi, M.; Emami, S.; Foroumadi, A.; Shafiee, A. Synthesis, in vitro cytotoxicity and apoptosis inducing study of 2-aryl-3-nitro-2H-chromene derivatives as potent anti-breast cancer agents. Eur. J. Med. Chem. 2014, 86, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Calgarotto, A.K.; da Silva Pereira, G.J.; Bechara, A.; Paredes-Gamero, E.J.; Barbosa, C.M.; Hirata, H.; de Souza Queiroz, M.L.; Smaili, S.S.; Bincoletto, C. Autophagy inhibited Ehrlich ascitic tumor cells apoptosis induced by the nitrostyrene derivative compounds: Relationship with cytosolic calcium mobilization. Eur. J. Pharmacol. 2012, 678, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Mitroulis, I.; Alexaki, V.I.; Kourtzelis, I.; Ziogas, A.; Hajishengallis, G.; Chavakis, T. Leukocyte integrins: Role in leukocyte recruitment and as therapeutic targets in inflammatory disease. Pharmacol. Ther. 2015, 147, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Sun, Z.; Huang, M.; Zhang, W.; Liu, J.; Chen, L.; Chen, F.; Zhou, Y.; Lin, J.; Huang, F.; et al. Nitrostyrene Derivatives Act as RXRα Ligands to Inhibit TNFα Activation of NF-κB. Cancer Res. 2015, 75, 2049–2060. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compound NPe are available from the authors. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugimoto, M.A.; De Jesus Amazonas da Silva, M.; Froede Brito, L.; Dos Santos Borges, R.; Amaral, F.A.; De Araujo Boleti, A.P.; Ordoñez, M.E.; Carlos Tavares, J.; Pires Sousa, L.; Lima, E.S. Anti-Inflammatory Potential of 1-Nitro-2-Phenylethylene. Molecules 2017, 22, 1977. https://doi.org/10.3390/molecules22111977
Sugimoto MA, De Jesus Amazonas da Silva M, Froede Brito L, Dos Santos Borges R, Amaral FA, De Araujo Boleti AP, Ordoñez ME, Carlos Tavares J, Pires Sousa L, Lima ES. Anti-Inflammatory Potential of 1-Nitro-2-Phenylethylene. Molecules. 2017; 22(11):1977. https://doi.org/10.3390/molecules22111977
Chicago/Turabian StyleSugimoto, Michelle A., Márcia De Jesus Amazonas da Silva, Larissa Froede Brito, Rosivaldo Dos Santos Borges, Flávio Almeida Amaral, Ana Paula De Araujo Boleti, Maritza Echevarria Ordoñez, Jose Carlos Tavares, Lirlandia Pires Sousa, and Emerson Silva Lima. 2017. "Anti-Inflammatory Potential of 1-Nitro-2-Phenylethylene" Molecules 22, no. 11: 1977. https://doi.org/10.3390/molecules22111977
APA StyleSugimoto, M. A., De Jesus Amazonas da Silva, M., Froede Brito, L., Dos Santos Borges, R., Amaral, F. A., De Araujo Boleti, A. P., Ordoñez, M. E., Carlos Tavares, J., Pires Sousa, L., & Lima, E. S. (2017). Anti-Inflammatory Potential of 1-Nitro-2-Phenylethylene. Molecules, 22(11), 1977. https://doi.org/10.3390/molecules22111977