
Catechins Variously Affect Activities of Conjugation Enzymes in Proliferating and Differentiated Caco-2 Cells

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Activities and Expression of Conjugation Enzymes in P and D Caco-2 Cells
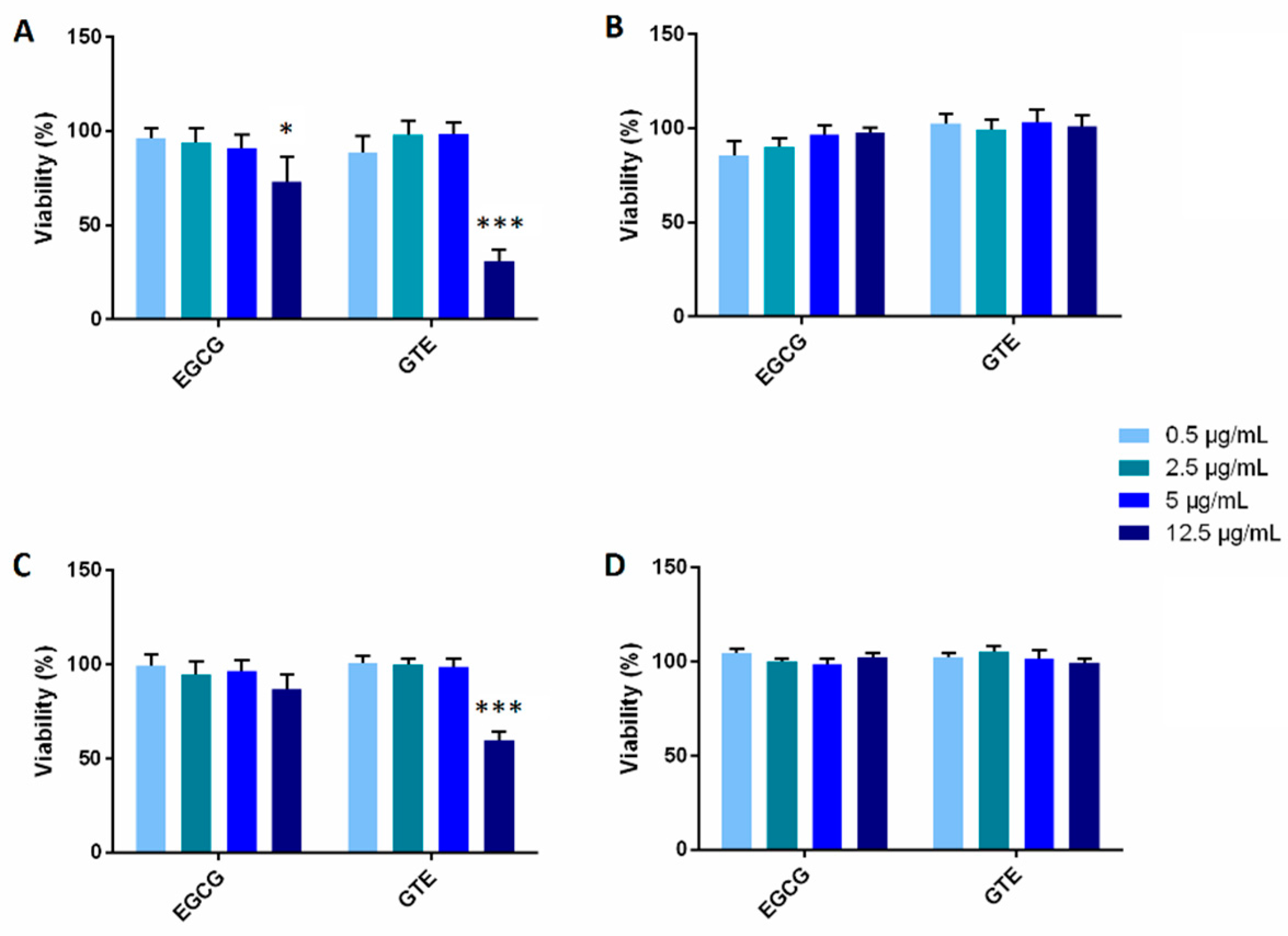
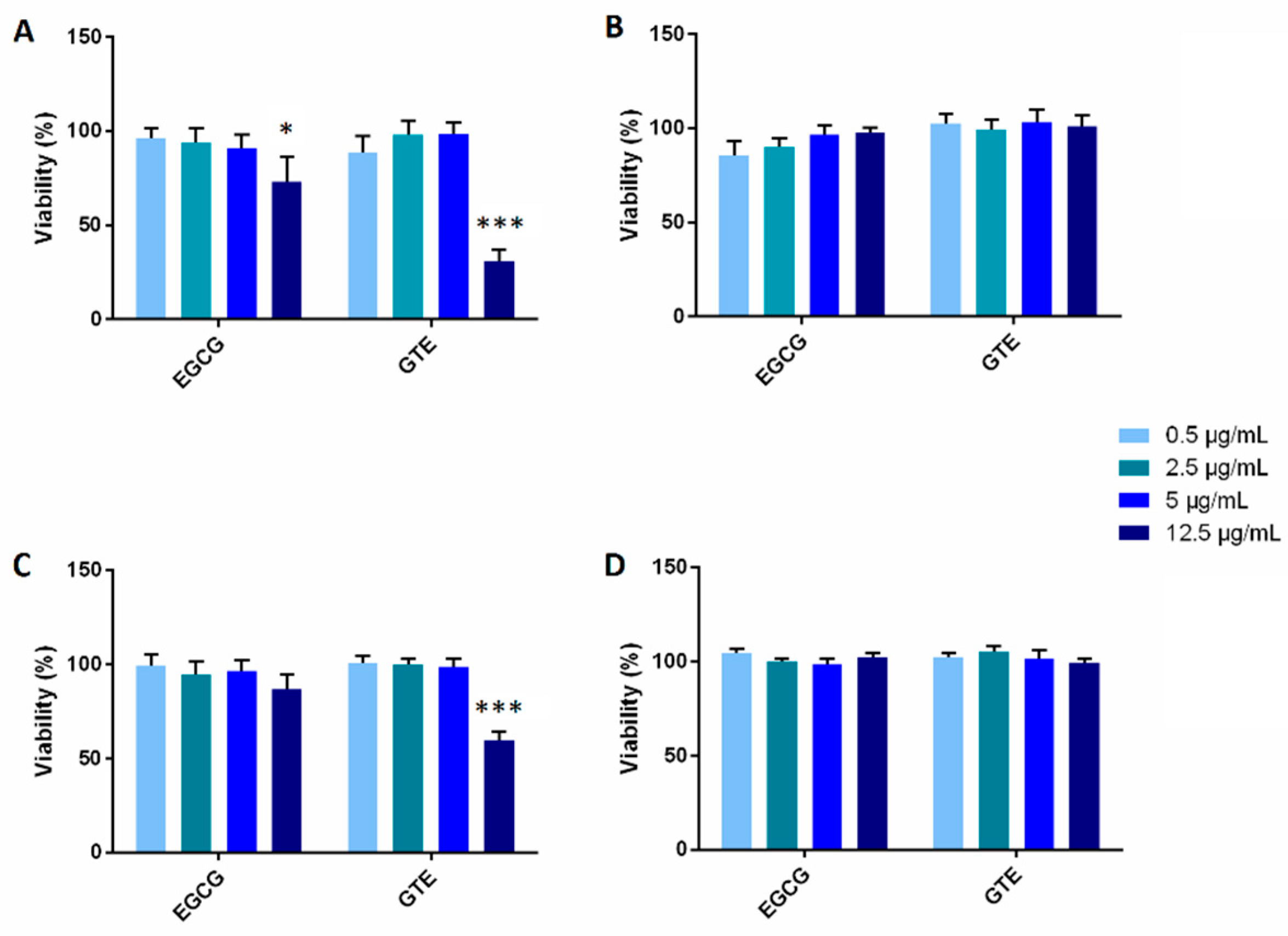
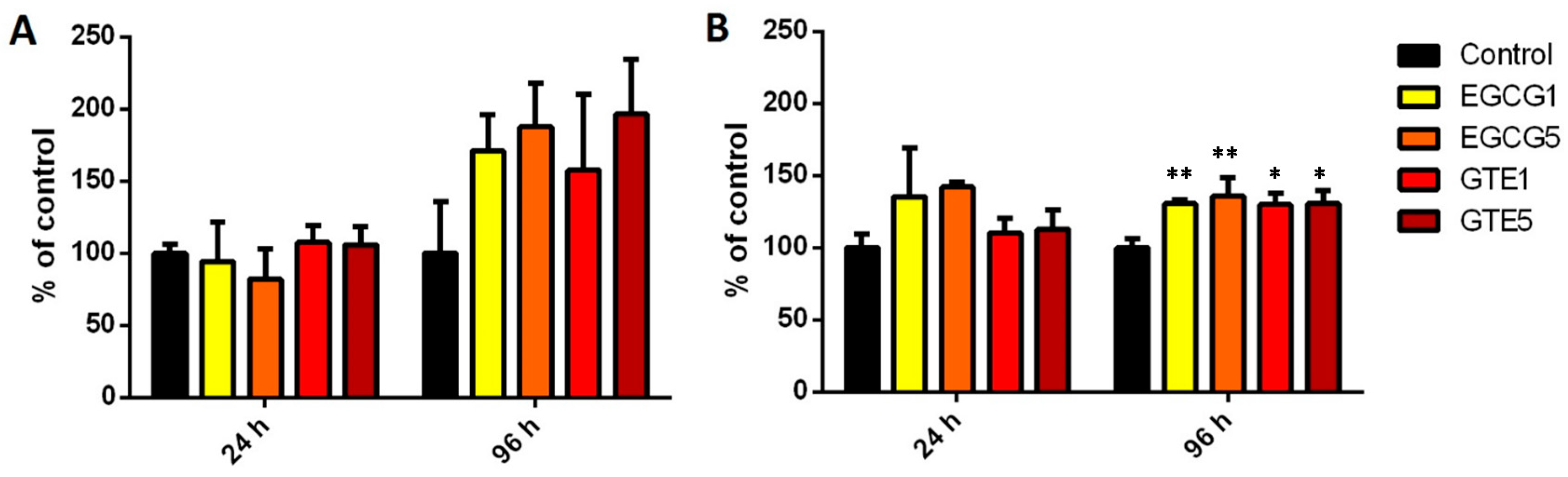
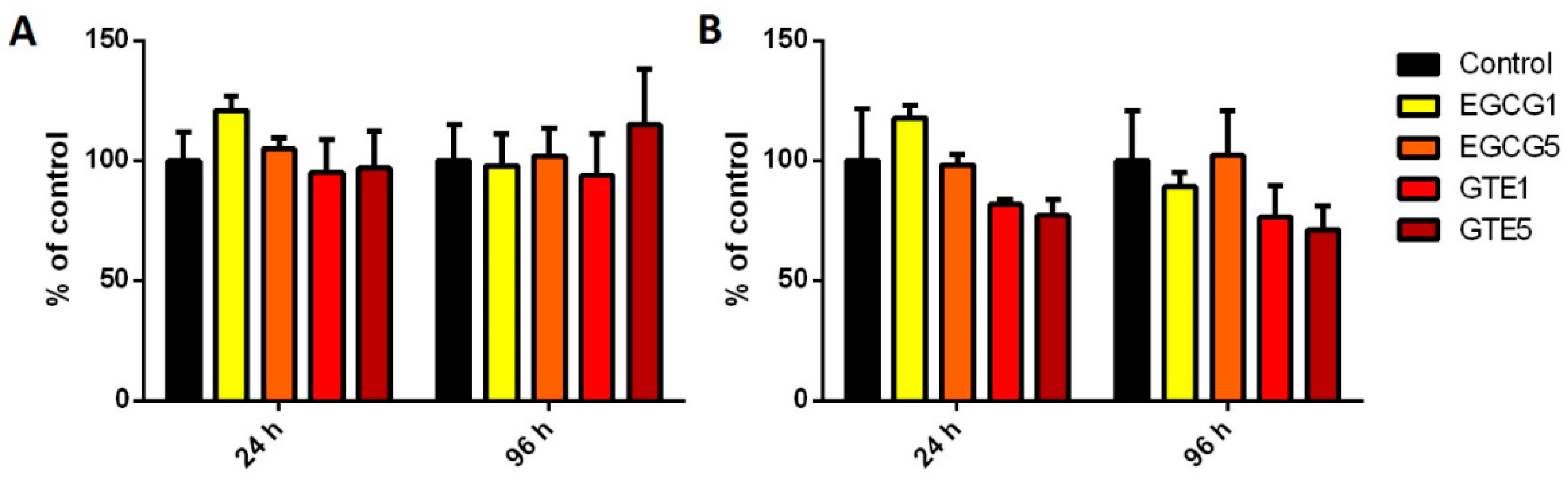
2.2. The Effect of GTE and EGCG on Cell Viability/Proliferation
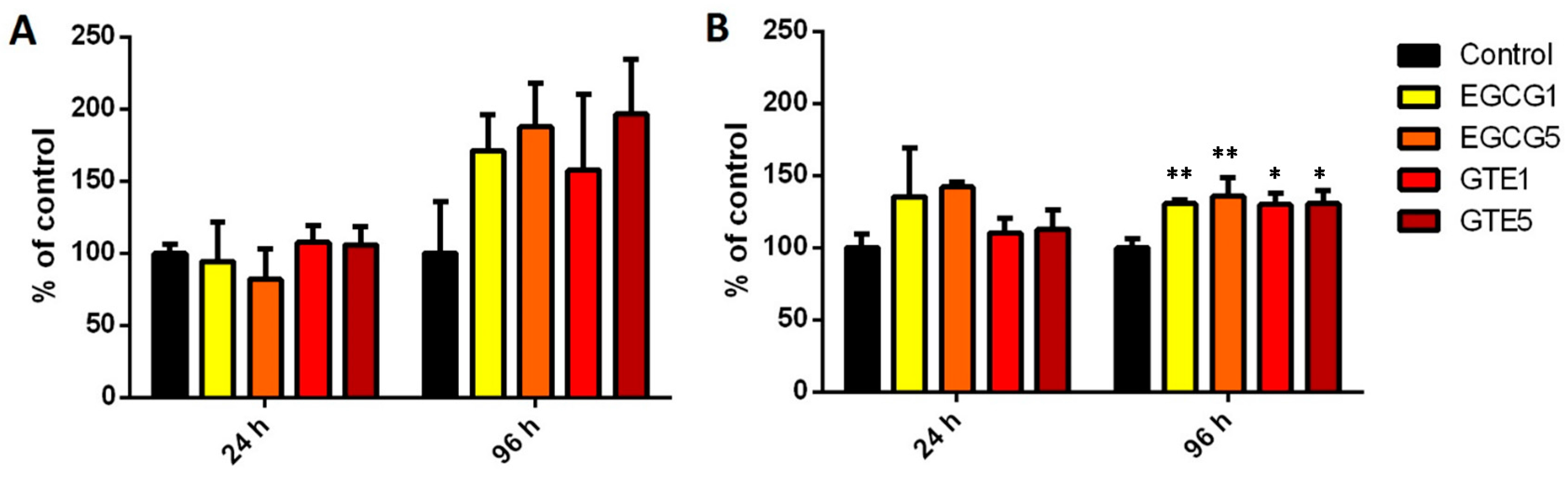
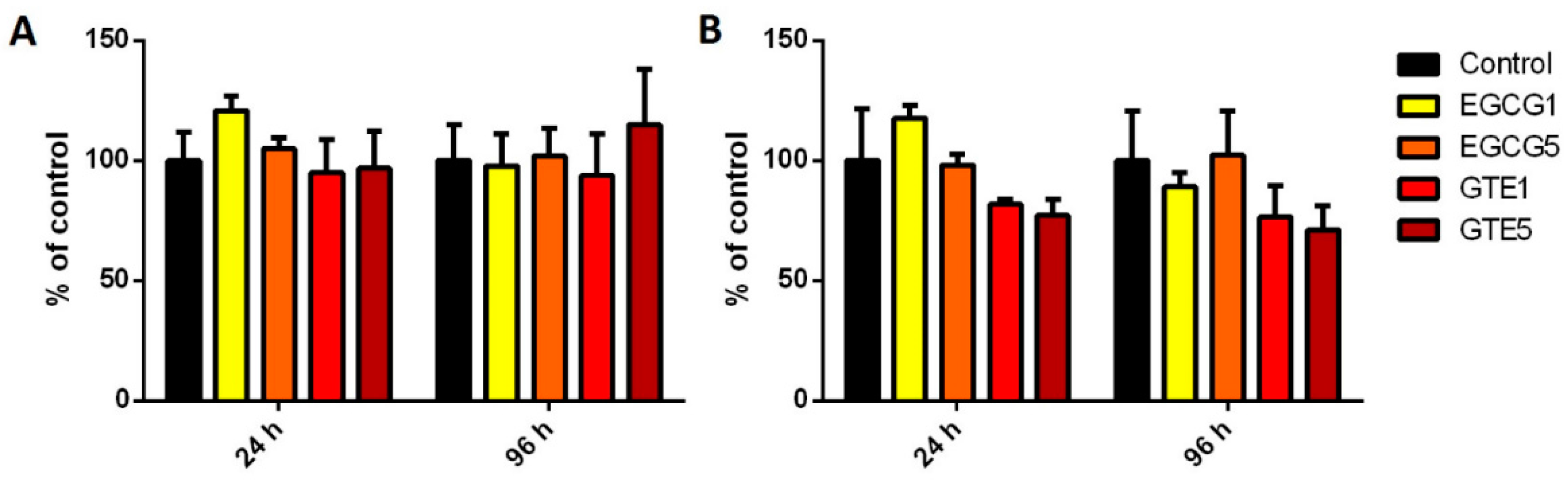
2.3. The Effect of GTE and EGCG on Conjugation Enzymes Activities and Expressions in P and D Caco-2 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cultivation of the Caco-2 Cell Line
4.3. Tests of Cell Viability
4.3.1. NRU Test
4.3.2. MTT Assay
4.4. Treatment of Caco-2 Cells with GTE and EGCG
4.5. Preparation of Subcellular Fractions
4.6. Enzyme Assays
4.7. RNA Isolation and Quantitative Real-Time PCR
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GTE | Green tea extract, Polyphenon 60 |
| EGCG | (−)-Epigallocatechin gallate |
| GST | Glutathione S-transferases |
| UGT | UDP-glucuronosyltransferases |
| SULT | Sulfotransferases |
| COMT | Catechol-O-methyltransferases |
| P | Proliferating Caco-2 cells |
| D | Differentiated Caco-2 cells |
| NRU | Neutral red uptake |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| EMEM | Eagle’s minimum essential medium |
| SAM | S-(5′-Adenosyl)-l-methionine |
| GSH | l-Glutathione |
| CDNB | 1-Chloro-2,4-dinitrobenzene |
| PAP | 3′-Phosphoadenosine 5′-phosphate |
| DMSO | Dimethyl sulfoxide |
| PBS | Phosphate buffered saline |
References
- Tran, C.D.; Timmins, P.; Conway, B.R.; Irwin, W.J. Investigation of the coordinated functional activities of cytochrome P450 3A4 and P-glycoprotein in limiting the absorption of xenobiotics in Caco-2 cells. J. Pharm. Sci. 2002, 91, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Buhrke, T.; Lengler, I.; Lampen, A. Analysis of proteomic changes induced upon cellular differentiation of the human intestinal cell line Caco-2. Dev. Growth Differ. 2011, 53, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Mariadason, J.M.; Arango, D.; Corner, G.A.; Aranes, M.J.; Hotchkiss, K.A.; Yang, W.; Augenlicht, L.H. A gene expression profile that defines colon cell maturation in vitro. Cancer Res. 2002, 62, 4791–4804. [Google Scholar] [PubMed]
- Scharmach, E.; Hessel, S.; Niemann, B.; Lampen, A. Glutathione S-transferase expression and isoenzyme composition during cell differentiation of Caco-2 cells. Toxicology 2009, 265, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, E.; Auclair, J.; Delvin, E.; Levy, E.; Menard, D.; Pshezhetsky, A.V.; Rivard, N.; Seidman, E.G.; Sinnett, D.; Vachon, P.H.; et al. Gene expression profiles of normal proliferating and differentiating human intestinal epithelial cells: A comparison with the Caco-2 cell model. J. Cell Biochem. 2006, 99, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Jancova, P.; Anzenbacher, P.; Anzenbacherova, E. Phase II drug metabolizing enzymes. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech. Repub. 2010, 154, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Kramer, S.D. The biochemistry of drug metabolism-an introduction: Part 4. reactions of conjugation and their enzymes. Chem. Biodivers. 2008, 5, 2171–2336. [Google Scholar] [CrossRef] [PubMed]
- Boušová, I.; Skálová, L. Inhibition and induction of glutathione S-transferases by flavonoids: Possible pharmacological and toxicological consequences. Drug Metab. Rev. 2012, 44, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Kramer, S.D. The biochemistry of drug metabolism-an introduction: Part 1. Principles and overview. Chem. Biodivers. 2006, 3, 1053–1101. [Google Scholar] [CrossRef] [PubMed]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef] [PubMed]
- Misaka, S.; Kawabe, K.; Onoue, S.; Werba, J.P.; Giroli, M.; Tamaki, S.; Kan, T.; Kimura, J.; Watanabe, H.; Yamada, S. Effects of green tea catechins on cytochrome P450 2B6, 2C8, 2C19, 2D6 and 3A activities in human liver and intestinal microsomes. Drug Metab. Pharmacokinet. 2013, 28, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Pan, E. The effects of green tea polyphenols on drug metabolism. Expert Opin. Drug Metab. Toxicol. 2012, 8, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Meinl, W.; Ebert, B.; Glatt, H.; Lampen, A. Sulfotransferase forms expressed in human intestinal Caco-2 and TC7 cells at varying stages of differentiation and role in benzo[a]pyrene metabolism. Drug Metab. Dispos. 2008, 36, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Du, G.J.; Zhang, Z.; Wen, X.D.; Yu, C.; Calway, T.; Yuan, C.S.; Wang, C.Z. Epigallocatechin Gallate (EGCG) is the most effective cancer chemopreventive polyphenol in green tea. Nutrients 2012, 4, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Thangapazham, R.L.; Singh, A.K.; Sharma, A.; Warren, J.; Gaddipati, J.P.; Maheshwari, R.K. Green tea polyphenols and its constituent epigallocatechin gallate inhibits proliferation of human breast cancer cells in vitro and in vivo. Cancer Lett. 2007, 245, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Liao, J.; Kim, K.; Yurkow, E.J.; Yang, C.S. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 1998, 19, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Sang, S.; Yang, C.S. Biotransformation of green tea polyphenols and the biological activities of those metabolites. Mol. Pharm. 2007, 4, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, U.; Szewczyk, K.; Hrabec, E.; Janecka, A.; Gorlach, S. Overview of metabolism and bioavailability enhancement of polyphenols. J. Agric. Food Chem. 2013, 61, 12183–12199. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mukhtar, H. Tea and health: Studies in humans. Curr. Pharm. Des. 2013, 19, 6141–6147. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Meng, X.; Yang, C.S. Enzymology of methylation of tea catechins and inhibition of catechol-O-methyltransferase by (−)-epigallocatechin gallate. Drug Metab. Dispos. 2003, 31, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Greenhaw, J.; Salminen, W.F.; Shi, Q. Mechanisms for epigallocatechin gallate induced inhibition of drug metabolizing enzymes in rat liver microsomes. Toxicol. Lett. 2012, 214, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.Y. Metabolism of green tea catechins: An overview. Curr. Drug Metab. 2006, 7, 755–809. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.H.; Hakim, I.A.; Vining, D.R.; Crowell, J.A.; Tome, M.E.; Ranger-Moore, J.; Cordova, C.A.; Mikhael, D.M.; Briehl, M.M.; Alberts, D.S. Modulation of human glutathione s-transferases by polyphenon e intervention. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1662–1666. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Pandey, M.; Bhaskaran, N.; Maclennan, G.T.; Fu, P.; Ponsky, L.E.; Gupta, S. Protection against oxidative DNA damage and stress in human prostate by glutathione S-transferase P1. Mol. Carcinog. 2014, 53, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Maliakal, P.P.; Coville, P.F.; Wanwimolruk, S. Tea consumption modulates hepatic drug metabolizing enzymes in Wistar rats. J. Pharm. Pharmacol. 2001, 53, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Matoušková, P.; Bártíková, H.; Boušová, I.; Szotáková, B.; Martin, J.; Skorkovská, J.; Hanušová, V.; Tománková, V.; Anzenbacherová, E.; Lišková, B.; et al. Effect of defined green tea extract in various dosage schemes on drug-metabolizing enzymes in mice in vivo. J. Funct. Foods 2014, 10, 327–335. [Google Scholar] [CrossRef]
- Pandey, M.; Shukla, S.; Gupta, S. Promoter demethylation and chromatin remodeling by green tea polyphenols leads to re-expression of GSTP1 in human prostate cancer cells. Int. J. Cancer 2010, 126, 2520–2533. [Google Scholar] [CrossRef] [PubMed]
- Nishimuta, H.; Ohtani, H.; Tsujimoto, M.; Ogura, K.; Hiratsuka, A.; Sawada, Y. Inhibitory effects of various beverages on human recombinant sulfotransferase isoforms SULT1A1 and SULT1A3. Biopharm. Drug Dispos. 2007, 28, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Matsui, M. Inhibitory effects of green tea and grape juice on the phenol sulfotransferase activity of mouse intestines and human colon carcinoma cell line, Caco-2. Biol. Pharm. Bull. 2000, 23, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Paul, F.; Moobed, M.; Baumann, G.; Zimmermann, B.F.; Stangl, K.; Stangl, V. The activity of catechol-O-methyltransferase (COMT) is not impaired by high doses of epigallocatechin-3-gallate (EGCG) in vivo. Eur. J. Pharmacol. 2014, 740, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.T.; Shim, J.Y.; Nagai, M.; Bai, H.W. Molecular modelling study of the mechanism of high-potency inhibition of human catechol-O-methyltransferase by (−)-epigallocatechin-3-O-gallate. Xenobiotica 2008, 38, 130–146. [Google Scholar] [CrossRef] [PubMed]
- Weisz, J.; Fritz-Wolz, G.; Clawson, G.A.; Benedict, C.M.; Abendroth, C.; Creveling, C.R. Induction of nuclear catechol-O-methyltransferase by estrogens in hamster kidney: Implications for estrogen-induced renal cancer. Carcinogenesis 1998, 19, 1307–1312. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, N.; Tsunoda, M.; Imai, K. Improved assay for catechol-O-methyltransferase activity utilizing norepinephrine as an enzymatic substrate and reversed-phase high-performance liquid chromatography with fluorescence detection. J. Chromatogr. A 2005, 1074, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Y. Total intracellular accumulation levels of dietary isothiocyanates determine their activity in elevation of cellular glutathione and induction of Phase 2 detoxification enzymes. Carcinogenesis 2001, 22, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Frame, L.T.; Ozawa, S.; Nowell, S.A.; Chou, H.C.; DeLongchamp, R.R.; Doerge, D.R.; Lang, N.P.; Kadlubar, F.F. A simple colorimetric assay for phenotyping the major human thermostable phenol sulfotransferase (SULT1A1) using platelet cytosols. Drug Metab. Dispos. 2000, 28, 1063–1068. [Google Scholar] [PubMed]
- Letelier, M.E.; Pimentel, A.; Pino, P.; Lepe, A.M.; Faundez, M.; Aracena, P.; Speisky, H. Microsomal UDP-glucuronyltransferase in rat liver: Oxidative activation. Basic Clin. Pharmacol. Toxicol. 2005, 96, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | P | D |
|---|---|---|
| GST | ||
| Activity | 249 ± 13 | 268 ± 20 |
| mRNA GSTA1/2 | 1.00 ± 0.04 | *** 9.15 ± 0.45 |
| mRNA GSTP1 | 1.00 ± 0.19 | *** 2.60 ± 0.14 |
| SULT | ||
| Activity | 20.7 ± 1.3 | ** 14.1 ± 1.6 |
| mRNA SULT1A1/3 | 1.00 ± 0.03 | *** 7.65 ± 0.33 |
| COMT | ||
| Activity | 10.5 ± 0.9 | 12.9 ± 0.3 |
| mRNA COMT | 1.00 ± 0.09 | *** 3.17 ± 0.14 |
| UGT | ||
| Activity | n.d. | n.d. |
| mRNA UGT1A | 1.00 ± 0.09 | *** 2.17 ± 0.08 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lněničková, K.; Procházková, E.; Skálová, L.; Matoušková, P.; Bártíková, H.; Souček, P.; Szotáková, B. Catechins Variously Affect Activities of Conjugation Enzymes in Proliferating and Differentiated Caco-2 Cells. Molecules 2016, 21, 1186. https://doi.org/10.3390/molecules21091186
Lněničková K, Procházková E, Skálová L, Matoušková P, Bártíková H, Souček P, Szotáková B. Catechins Variously Affect Activities of Conjugation Enzymes in Proliferating and Differentiated Caco-2 Cells. Molecules. 2016; 21(9):1186. https://doi.org/10.3390/molecules21091186
Chicago/Turabian StyleLněničková, Kateřina, Eliška Procházková, Lenka Skálová, Petra Matoušková, Hana Bártíková, Pavel Souček, and Barbora Szotáková. 2016. "Catechins Variously Affect Activities of Conjugation Enzymes in Proliferating and Differentiated Caco-2 Cells" Molecules 21, no. 9: 1186. https://doi.org/10.3390/molecules21091186
APA StyleLněničková, K., Procházková, E., Skálová, L., Matoušková, P., Bártíková, H., Souček, P., & Szotáková, B. (2016). Catechins Variously Affect Activities of Conjugation Enzymes in Proliferating and Differentiated Caco-2 Cells. Molecules, 21(9), 1186. https://doi.org/10.3390/molecules21091186