
In Vivo Metabolite Profiling of a Purified Ellagitannin Isolated from Polygonum capitatum in Rats

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
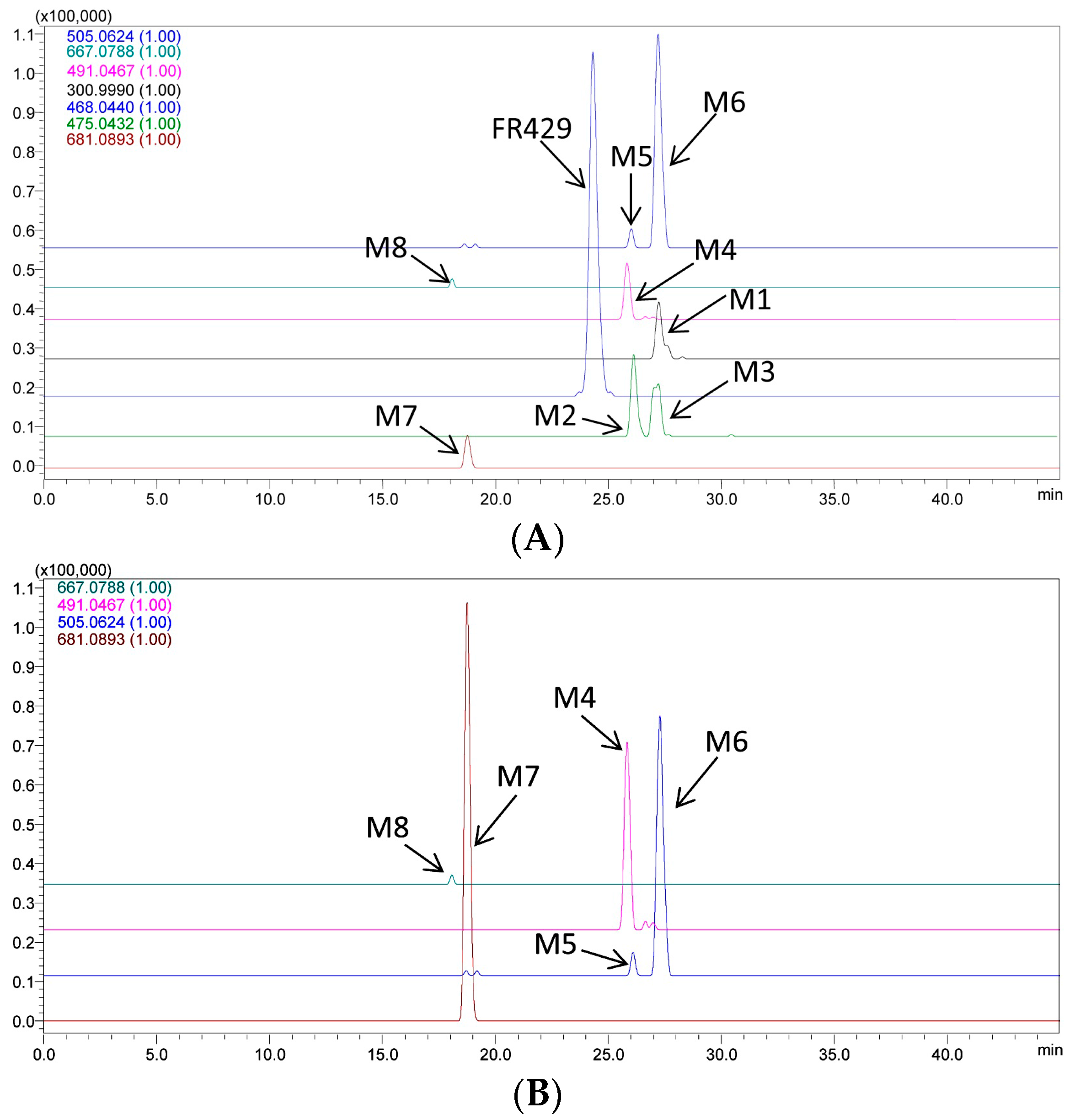
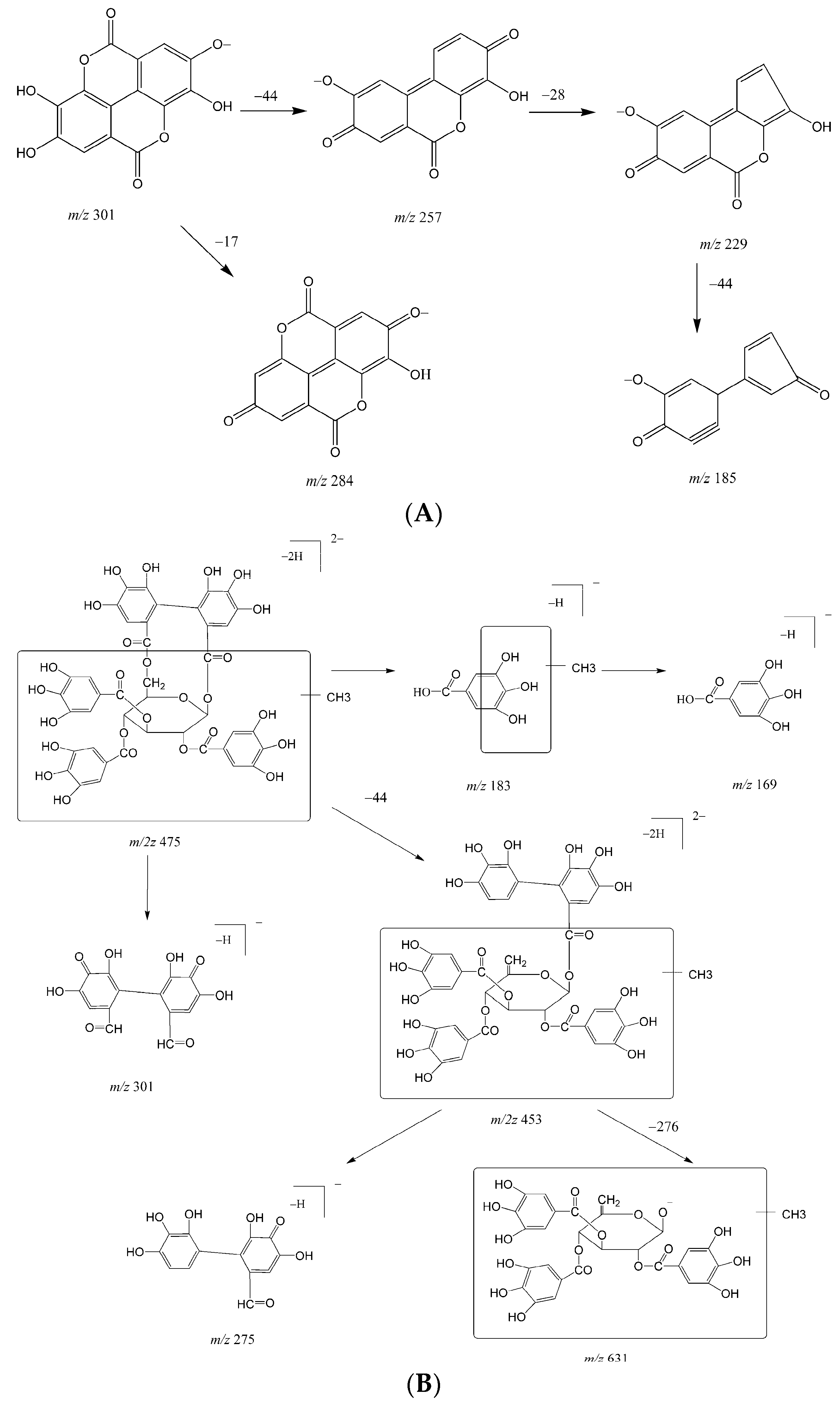
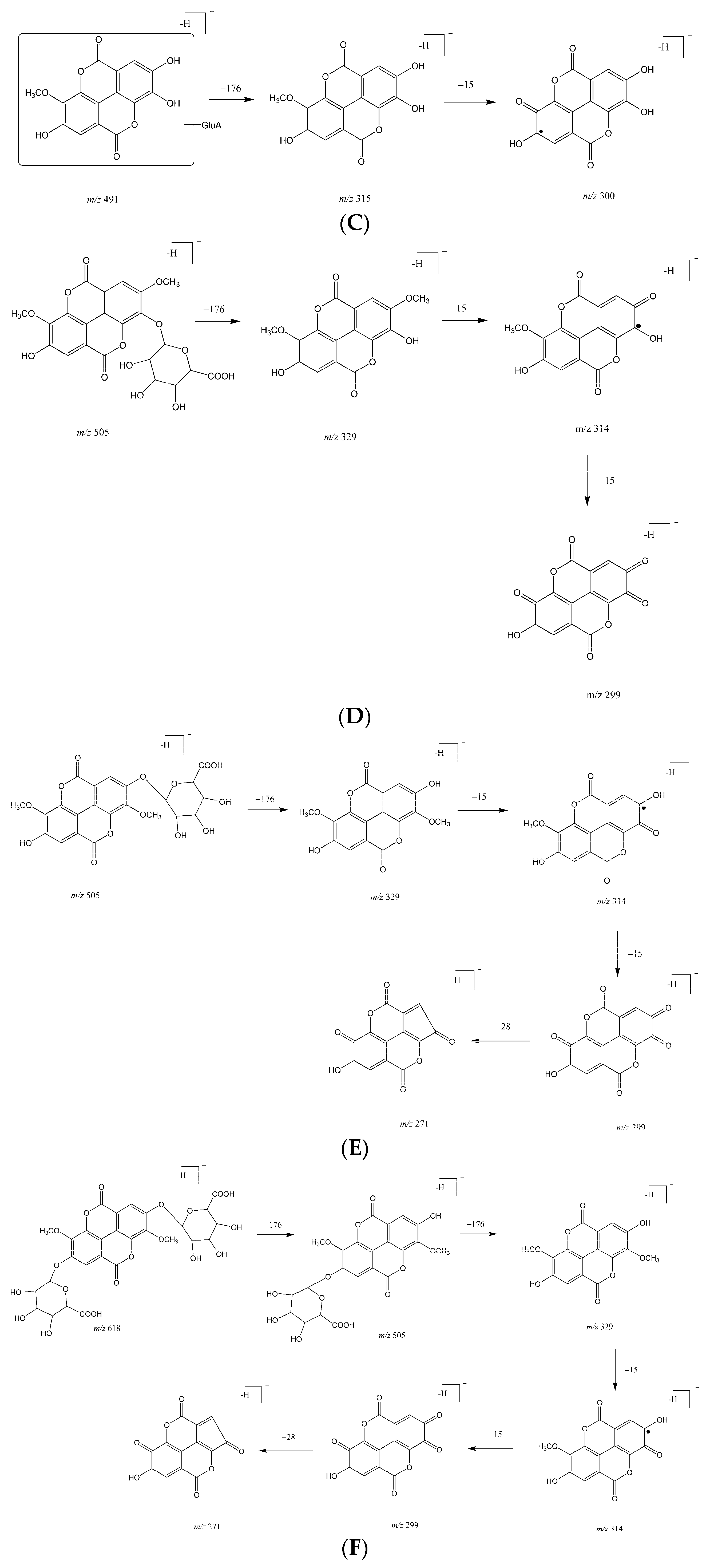
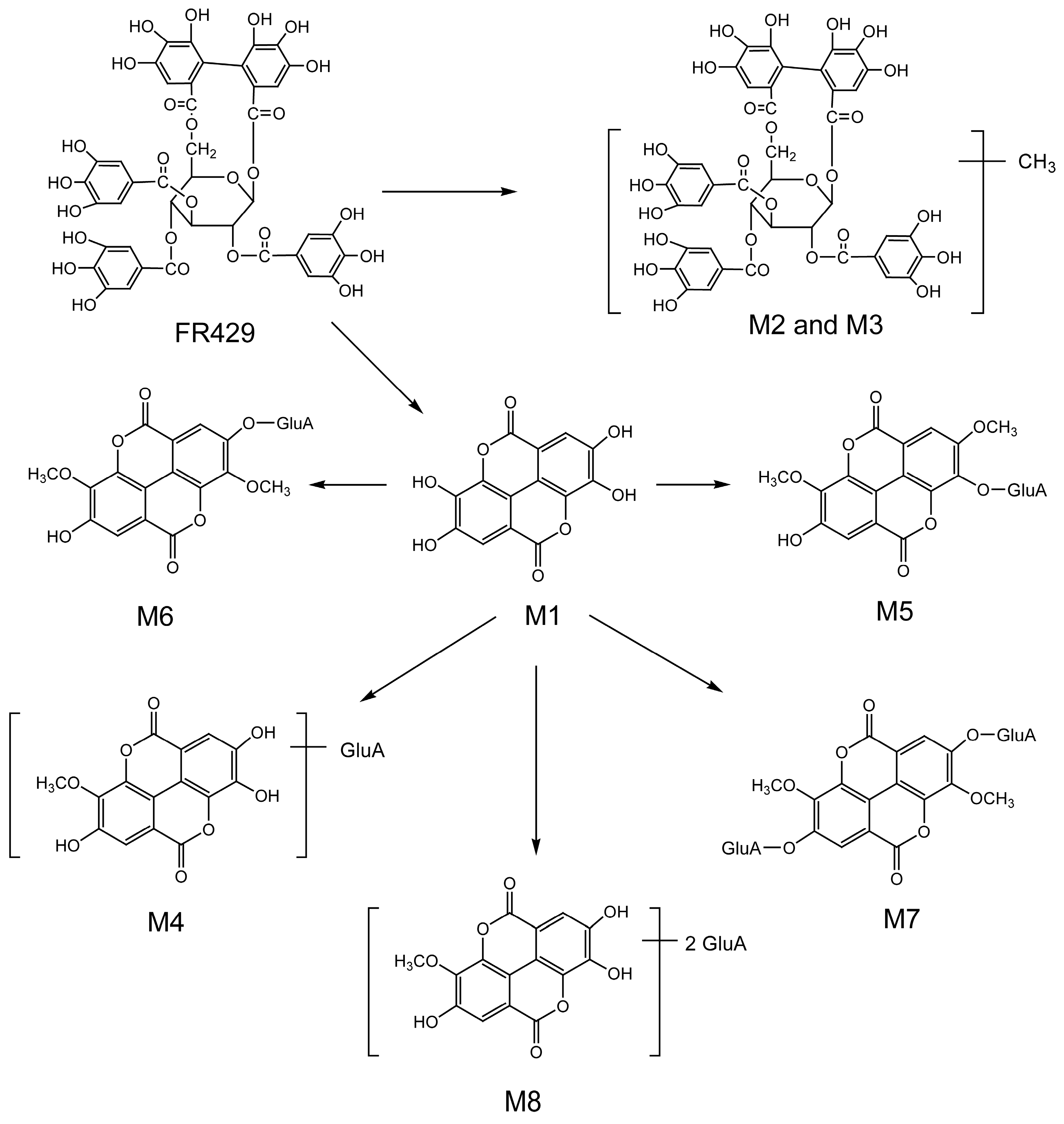
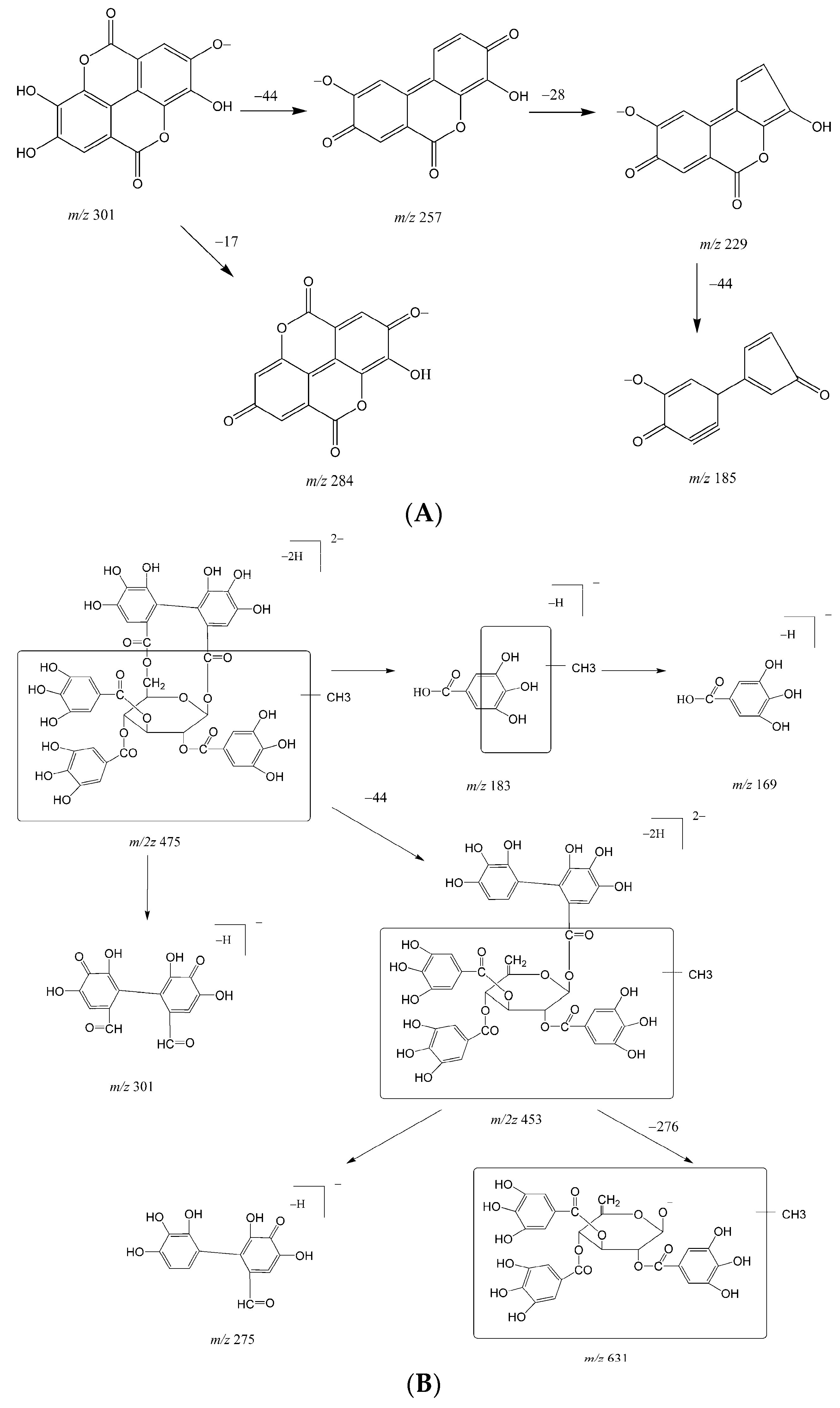
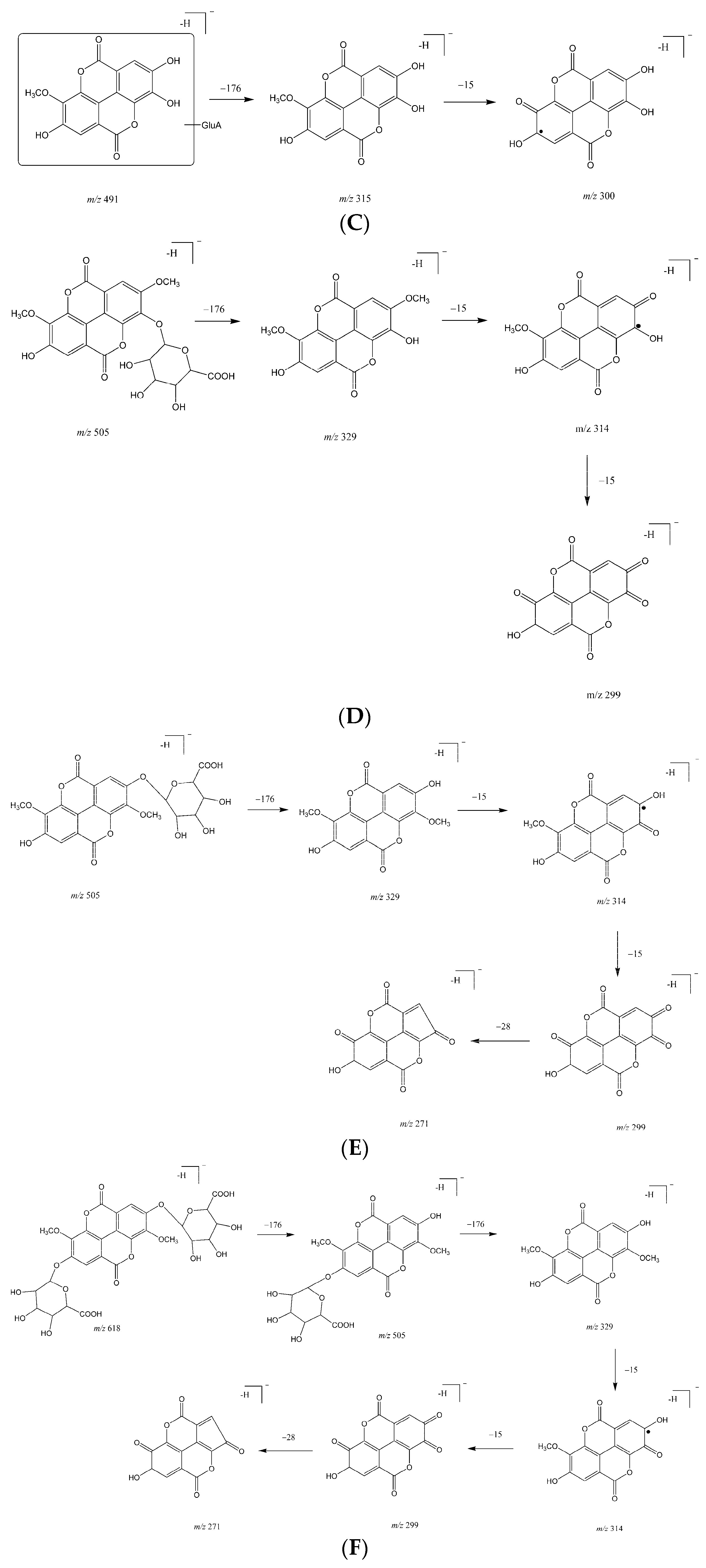
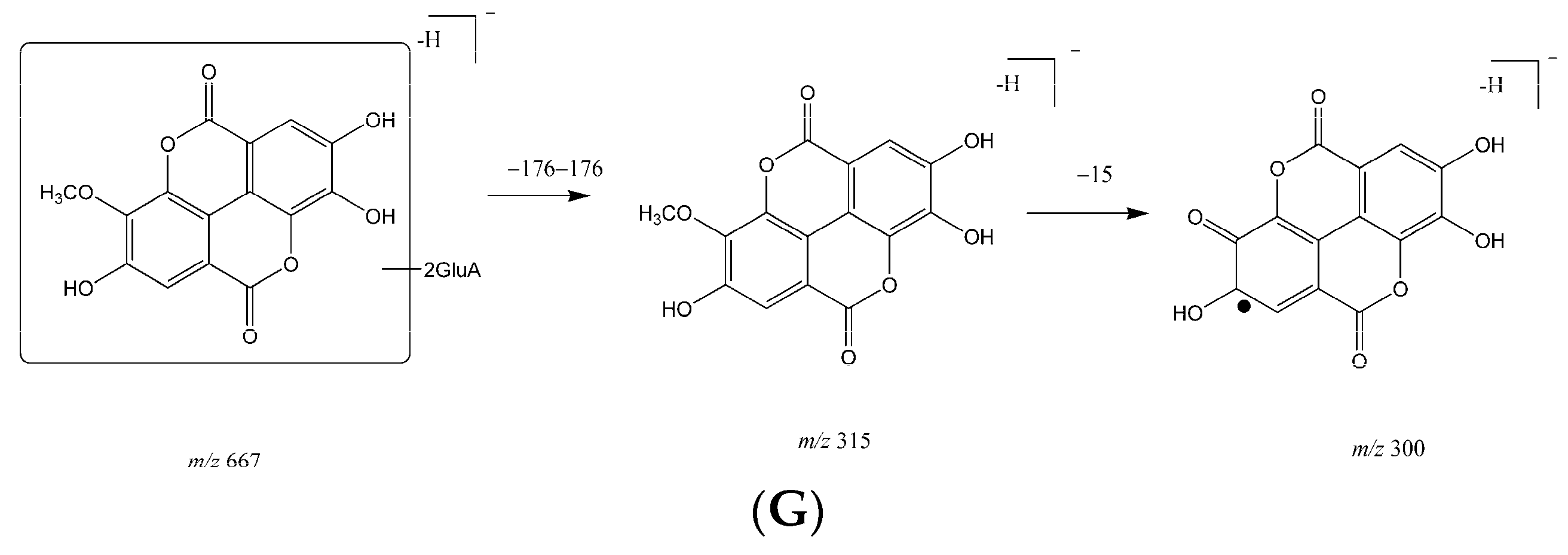
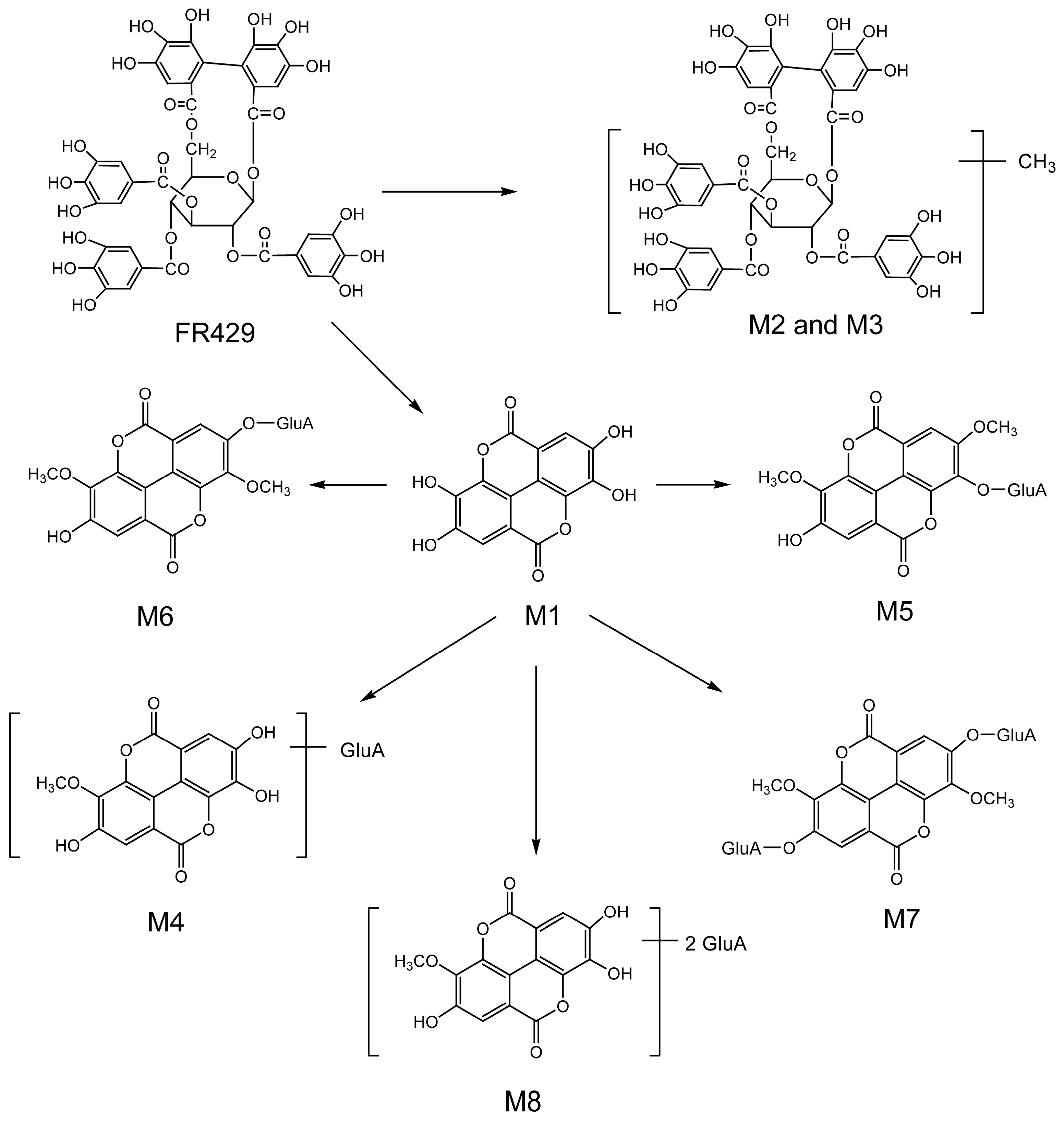
2.1. Metabolites in Plasma
2.2. Metabolites in Bile
2.3. Metabolites in Urine
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Animals
3.3. LC/MSn-IT-TOF Analysis Conditions
3.4. Metabolites Identification in Plasma, Urine and Bile in Vivo
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ito, H.; Iguchi, A.; Hatano, T. Identification of urinary and intestinal bacterial metabolites of ellagitannin Geraniin in rats. J. Agric. Food Chem. 2008, 56, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Bakkalbasi, E.; Mentes, O.; Artik, N. Food ellagitannins-occurrence, effects of processing and storage. Crit. Rev. Food Sci. Nutr. 2009, 49, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Okabe, S.; Suganuma, M.; Imayoshi, Y.; Taniguchi, S.; Yoshida, T.; Fujiki, H. New TNF-alpha releasing inhibitors, geraniin and corilagin, in leaves of Acer nikoense, Megusurino-ki. Biol. Pharm. Bull. 2001, 24, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, H.; Murakami, T.; Yamamoto, N.; Sakagami, H.; Tanuma, S.; Hatano, T.; Yoshida, T.; Okuda, T. Inhibition of Human Immunodeficiency Viral Replication by Tannins and Related-Compounds. Antivir. Res. 1992, 18, 91–103. [Google Scholar] [CrossRef]
- Boukharta, M.; Jalbert, G.; Castonguay, A. Biodistribution of ellagic acid and dose-related inhibition of lung tumorigenesis in A/J mice. Nutr. Cancer 1992, 18, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Cerda, B.; Llorach, R.; Ceron, J.J.; Espin, J.C.; Tomas-Barberan, F.A. Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice. Eur. J. Nutr. 2003, 42, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Zhang, Y.; McKeever, R.; Henning, S.M.; Lee, R.P.; Suchard, M.A.; Li, Z.; Chen, S.; Thames, G.; Zerlin, A.; et al. Pomegranate juice and extracts provide similar levels of plasma and urinary ellagitannin metabolites in human subjects. J. Med. Food 2008, 11, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Munoz, C.; Vaillant, F. Metabolic fate of ellagitannins: Implications for health, and research perspectives for innovative functional foods. Crit. Rev. Food Sci. Nutr. 2014, 54, 1584–1598. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Ma, J.Y.; Zhang, X.F.; Wang, Y.; Feng, R.; Chen, Y.C.; Tan, X.S.; Zhang, Y.Y.; Sun, Y.P.; Zhou, Y.; et al. Identification of metabolites of FR429, a potential antitumor ellagitannin, transformed by rat intestinal bacteria in vitro, based on liquid chromatography-ion trap-time of flight mass spectrometry analysis. J. Pharm. Biomed. Anal. 2012, 71, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Feng, R.; Wang, W.; Zhang, L.; Ye, X.; Wang, Y.; Wang, M. Quantitative analysis of three active constituents in Miao regional herb, Polygonum capitatum by HPLC/DAD/MS. Chin. J. Pharm. Anal. 2008, 28, 1793–1796. [Google Scholar]
- Ma, J.Y.; Zhou, X.; Fu, J.; Hu, T.; Or, P.M.Y.; Feng, R.; He, C.Y.; Chen, W.J.; Zhang, X.; Chen, Y.; et al. Metabolite profiling analysis of FR429, an ellagitannin purified from Polygonum capitatum, in rat and human liver microsomes, cytosol and rat primary hepatocytes in vitro. Chem. Biol. Interact. 2014, 220, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, J.; Chow, S.C.; Li, C.H.; Xiao, Z.; Feng, R.; Fu, J.; Chen, Y. A potential antitumor ellagitannin, davidiin, inhibited hepatocellular tumor growth by targeting EZH2. Tumour Biol. 2014, 35, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, Z.Y.; Mo, S.L.; Loo, T.Y.; Wang, D.M.; Luo, H.B.; Yang, D.P.; Chen, Y.L.; Shen, J.G.; Chen, J.P. Ellagic acid, a phenolic compound, exerts anti-angiogenesis effects via VEGFR-2 signaling pathway in breast cancer. Breast Cancer Res. Treat. 2012, 134, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Kasimsetty, S.G.; Khan, S.I.; Ferreira, D. Urolithins, intestinal microbial metabolites of Pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J. Agric. Food Chem. 2009, 57, 10181–10186. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Sarrias, A.; Gimenez-Bastida, J.A.; Garcia-Conesa, M.T.; Gomez-Sanchez, M.B.; Garcia-Talavera, N.V.; Gil-Izquierdo, A.; Sanchez-Alvarez, C.; Fontana-Compiano, L.O.; Morga-Egea, J.P.; Pastor-Quirante, F.A.; et al. Occurrence of urolithins, gut microbiota ellagic acid metabolites and proliferation markers expression response in the human prostate gland upon consumption of walnuts and pomegranate juice. Mol. Nutr. Food Res. 2010, 54, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Ngounou, F.N.; Choudhary, M.I.; Malik, S.; Makhmoor, T.; Nur, E.A.M.; Zareen, S.; Lontsi, D.; Ayafor, J.F.; Sondengam, B.L. New antioxidant and antimicrobial ellagic acid derivatives from Pteleopsis hylodendron. Planta Med. 2001, 67, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | tR (min) | MS1 [M − H]− | Diff (ppm) | Fragments | Structure | Biological Matrix |
|---|---|---|---|---|---|---|
| FR429 | 24.5 | 468.0397(2) | 7.99 | 370.0413(2), 275.0364, 300.9967, 169.0164 | FR429 | Plasma, bile |
| M1 | 27.0 | 300.9994 | 2.99 | 229.0147, 185.0256, 257.0099, 283.9998 | ellagic acid | bile |
| M2 | 26.3 | 475.0526(2) | 1.68 | 300.9961, 275.0175, 169.0181, 183.0300, 631.0838 | FR429 methyl ether | bile |
| M3 | 27.4 | 475.0498(2) | 4.21 | 300.9956, 275.0174, 169.0186, 453.0572(2), 631.0849 | FR429 methyl ether | bile |
| M4 | 25.8 | 491.0469 | 0.41 | 315.0138, 299.9911 | ellagic acid methyl ether glucuronide | bile, urine |
| M5 | 26.0 | 505.0661 | 7.33 | 329.0211, 314.0071, 298.9756 | ellagic acid dimethyl ether glucuronide | bile, urine |
| M6 | 27.3 | 505.0635 | 2.18 | 329.0288, 314.0041, 298.9793, 270.9909 | ellagic acid dimethyl ether glucuronide (isomer) | bile, urine |
| M7 | 18.4 | 681.0947 | 0.29 | 505.0564, 329.0250, 314.0029, 298.9759, 270.9918 | ellagic acid dimethyl ether diglucuronide | bile, urine |
| M8 | 17.8 | 667.0827 | 5.85 | 315.0170, 299.9915 | ellagic acid methyl ether diglucuronide | bile, urine |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.-Y.; Zhou, X.; Fu, J.; He, C.-Y.; Feng, R.; Huang, M.; Shou, J.-W.; Zhao, Z.-X.; Li, X.-Y.; Zhang, L.; et al. In Vivo Metabolite Profiling of a Purified Ellagitannin Isolated from Polygonum capitatum in Rats. Molecules 2016, 21, 1110. https://doi.org/10.3390/molecules21091110
Ma J-Y, Zhou X, Fu J, He C-Y, Feng R, Huang M, Shou J-W, Zhao Z-X, Li X-Y, Zhang L, et al. In Vivo Metabolite Profiling of a Purified Ellagitannin Isolated from Polygonum capitatum in Rats. Molecules. 2016; 21(9):1110. https://doi.org/10.3390/molecules21091110
Chicago/Turabian StyleMa, Jing-Yi, Xuelin Zhou, Jie Fu, Chi-Yu He, Ru Feng, Min Huang, Jia-Wen Shou, Zhen-Xiong Zhao, Xiao-Yang Li, Luye Zhang, and et al. 2016. "In Vivo Metabolite Profiling of a Purified Ellagitannin Isolated from Polygonum capitatum in Rats" Molecules 21, no. 9: 1110. https://doi.org/10.3390/molecules21091110
APA StyleMa, J.-Y., Zhou, X., Fu, J., He, C.-Y., Feng, R., Huang, M., Shou, J.-W., Zhao, Z.-X., Li, X.-Y., Zhang, L., Chen, Y.-C., & Wang, Y. (2016). In Vivo Metabolite Profiling of a Purified Ellagitannin Isolated from Polygonum capitatum in Rats. Molecules, 21(9), 1110. https://doi.org/10.3390/molecules21091110