
In Vivo Monitoring of the Growth of Fertilized Eggs of Medaka Fish (Oryzias latipes) by Near-Infrared Spectroscopy and Near-Infrared Imaging—A Marked Change in the Relative Content of Weakly Hydrogen-Bonded Water in Egg Yolk Just before Hatching

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Breeding of Japanese Medaka (Oryzias Latipes)
2.2. Instrumentation and NIR Measurements
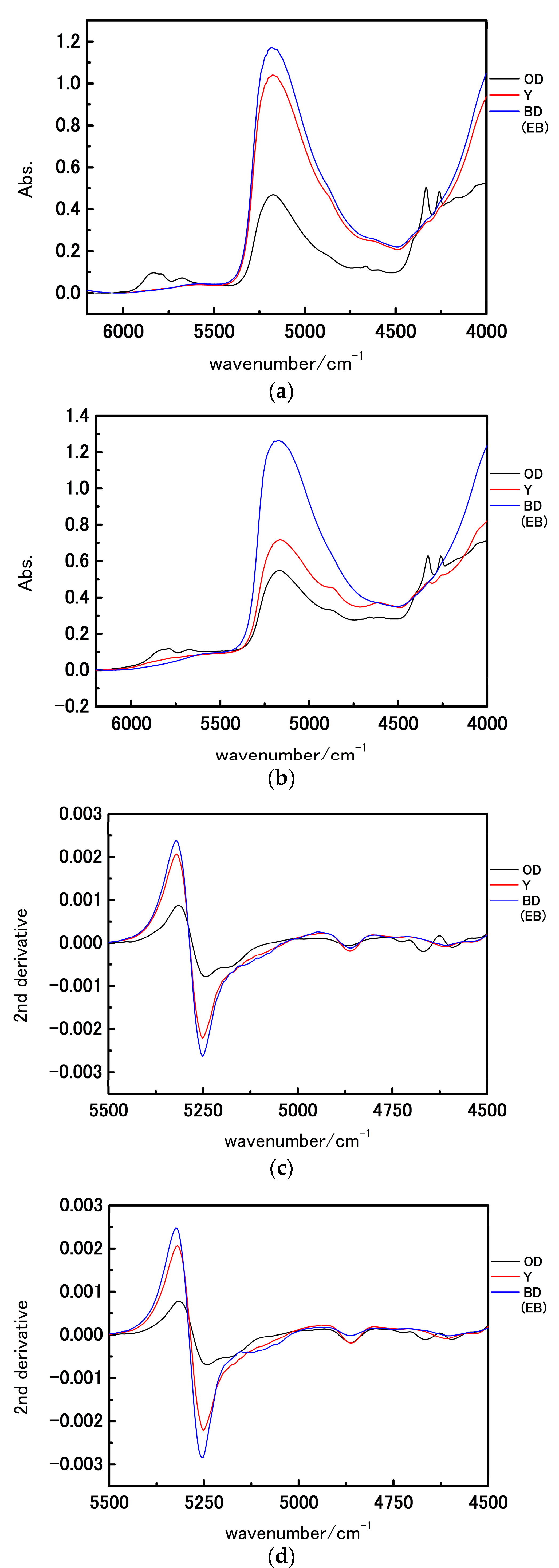
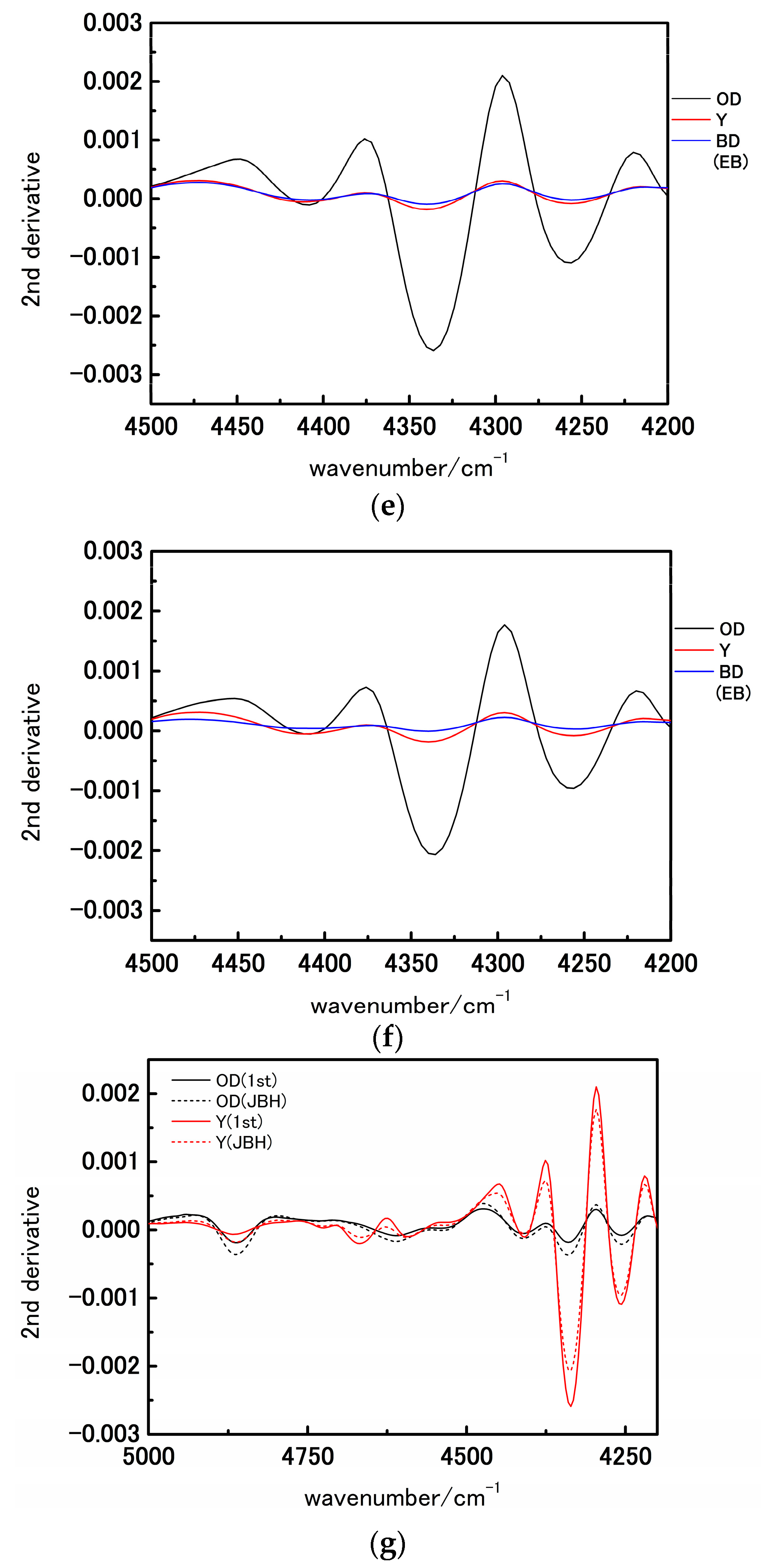
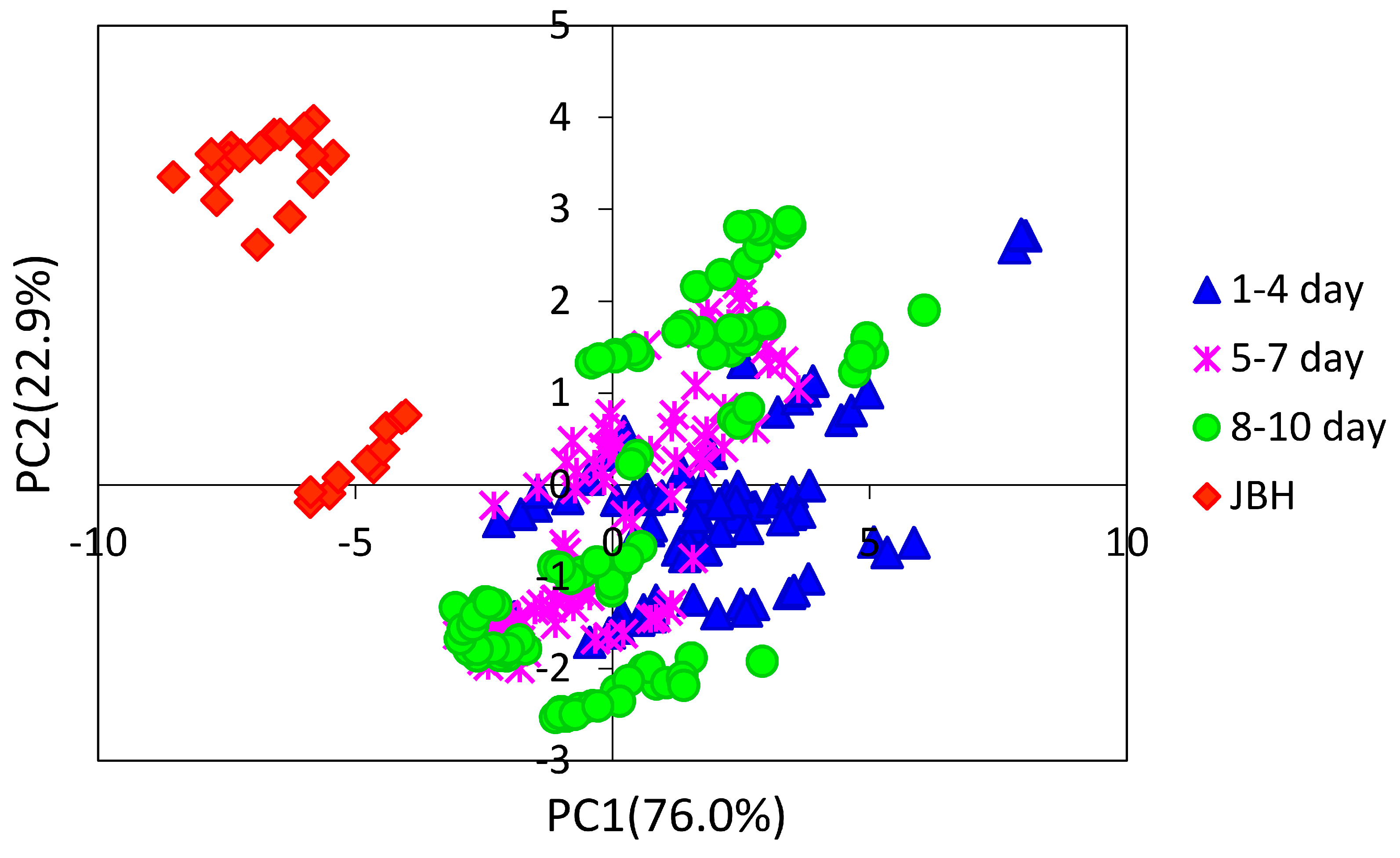
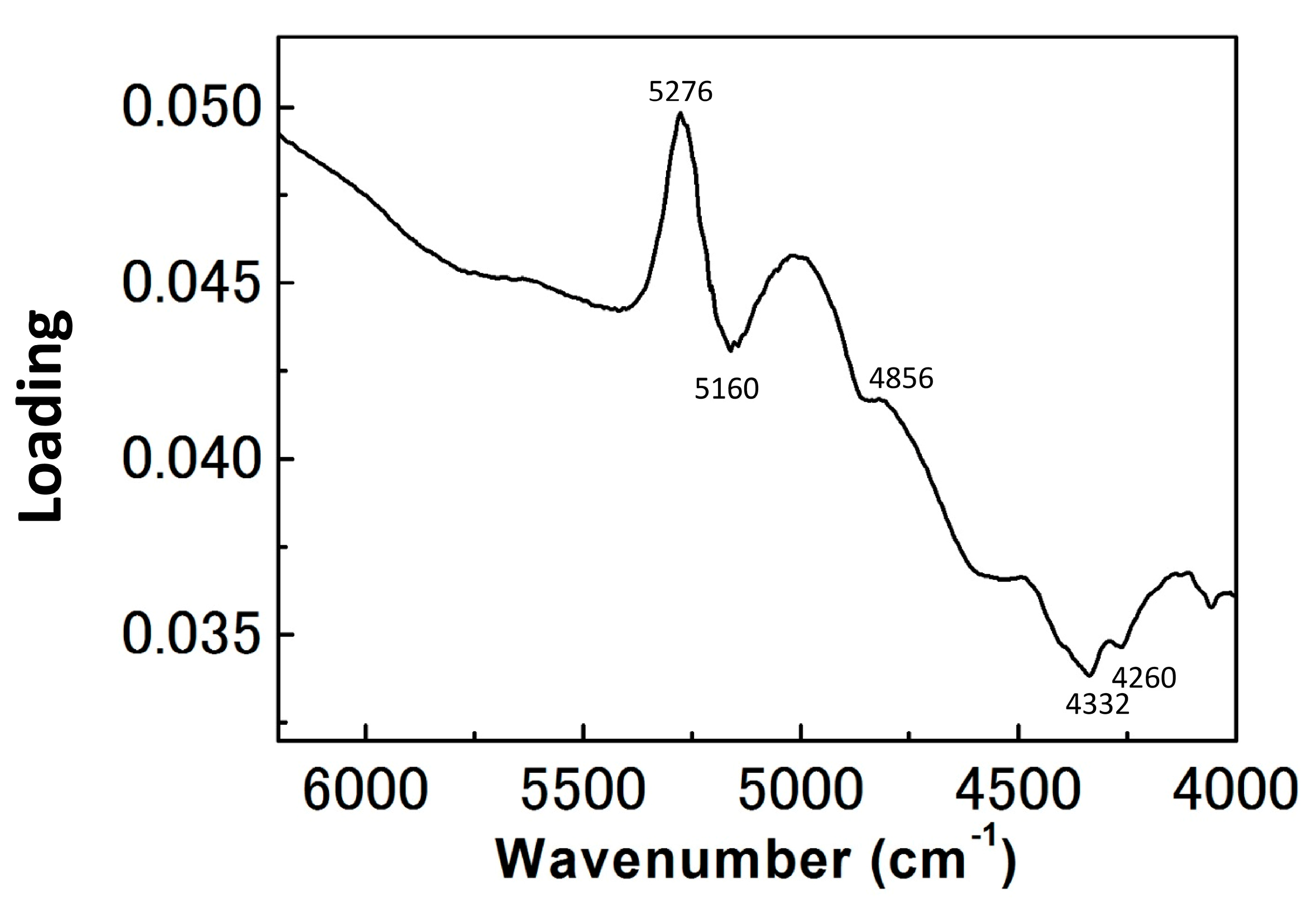
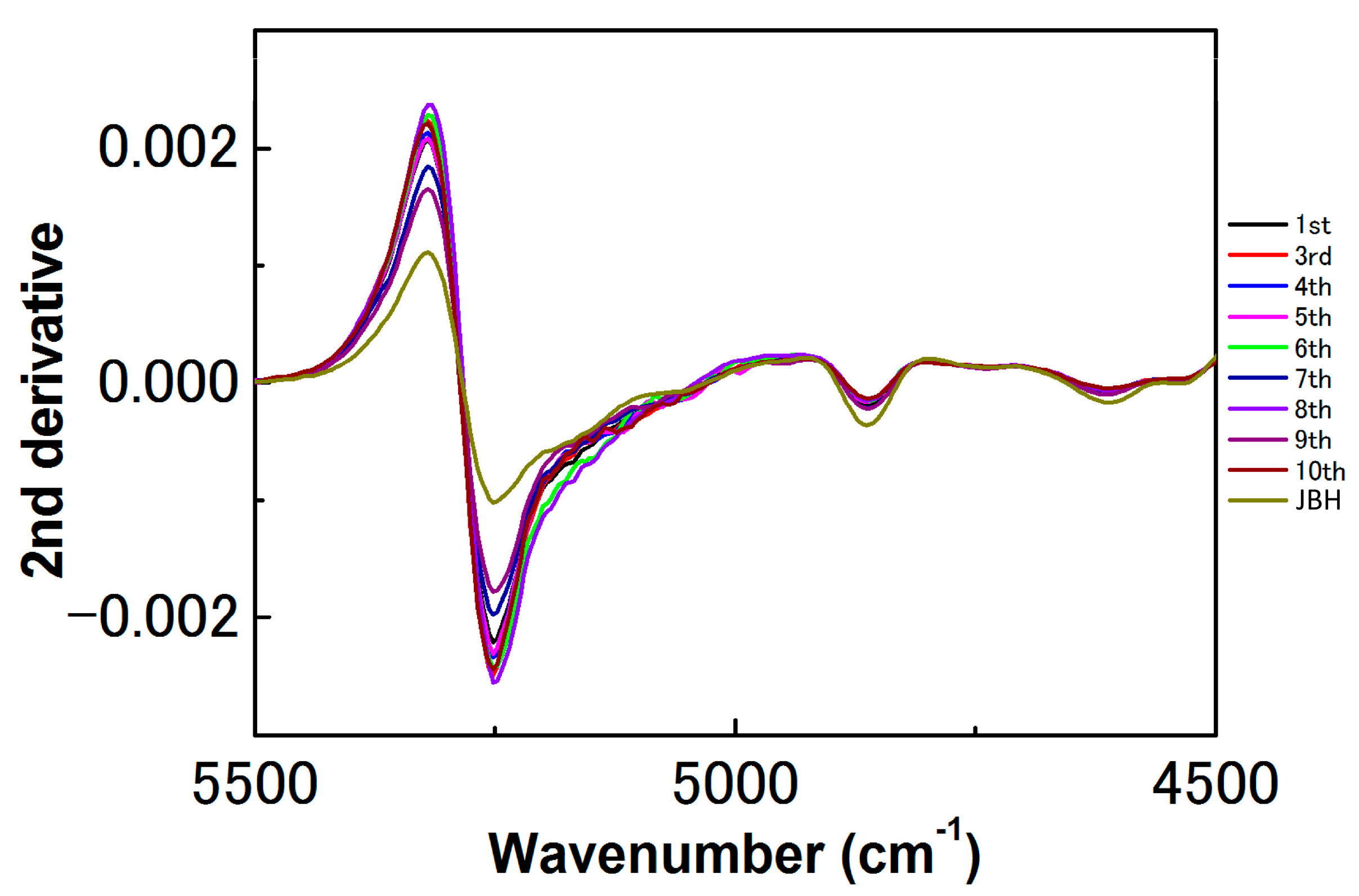
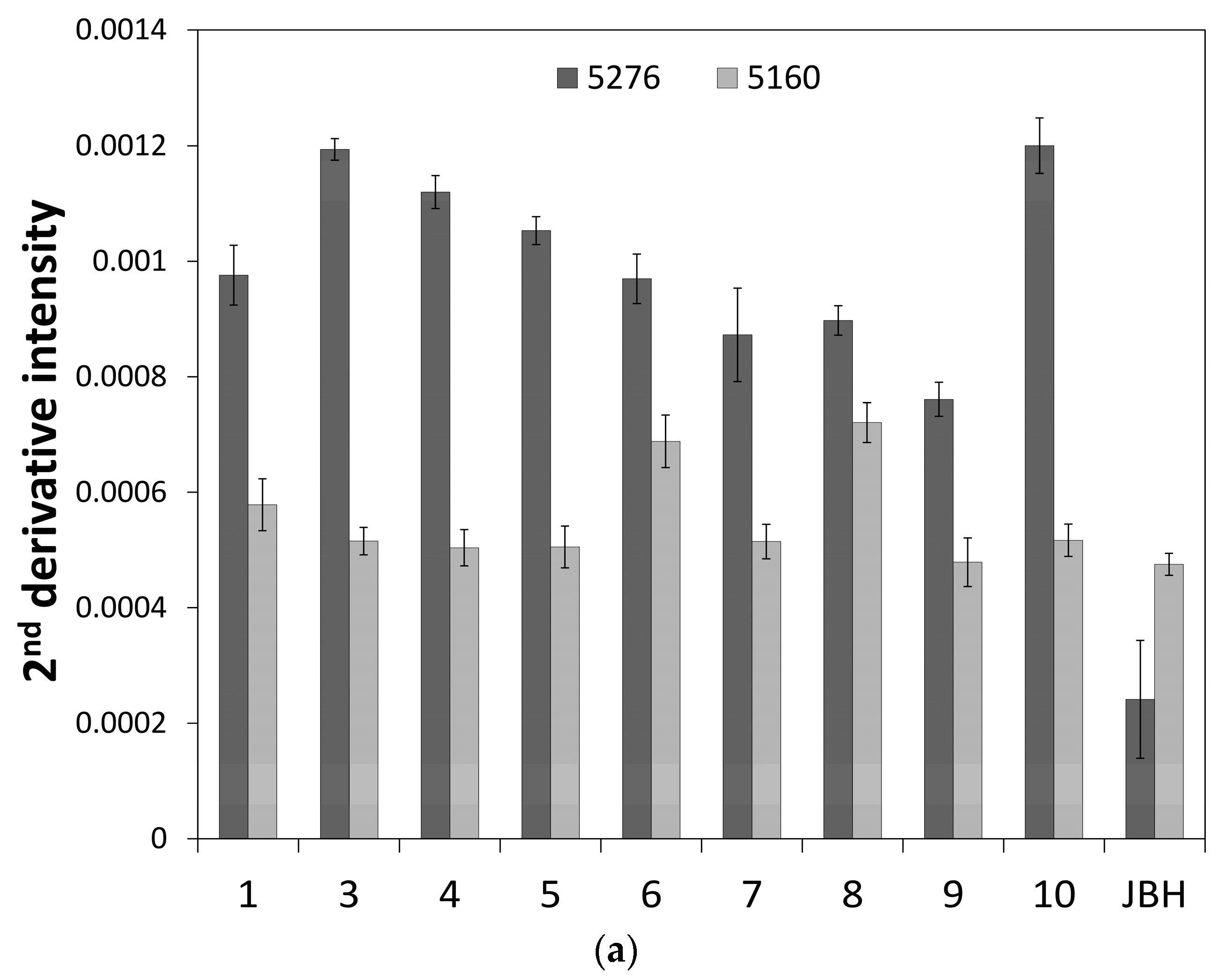
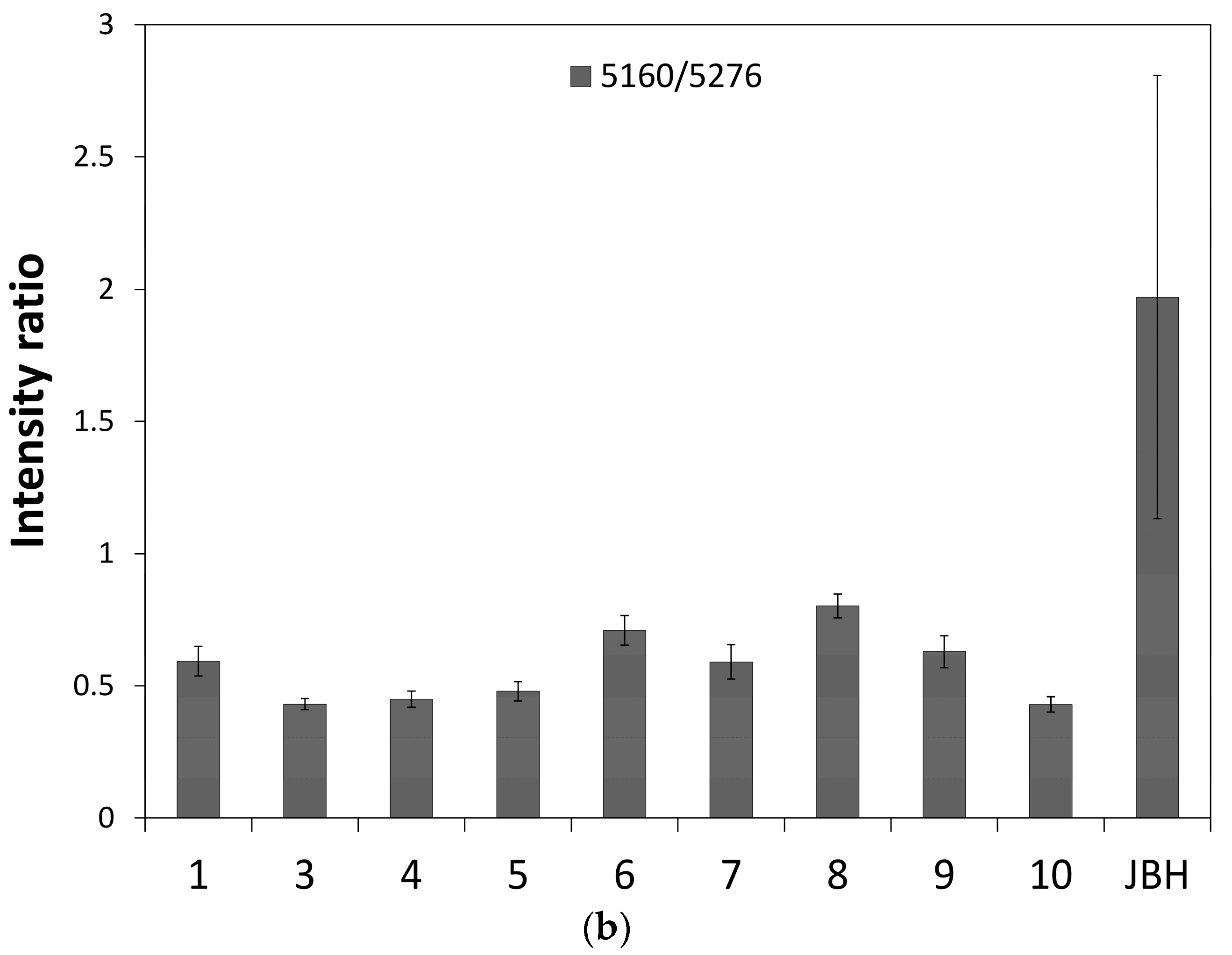
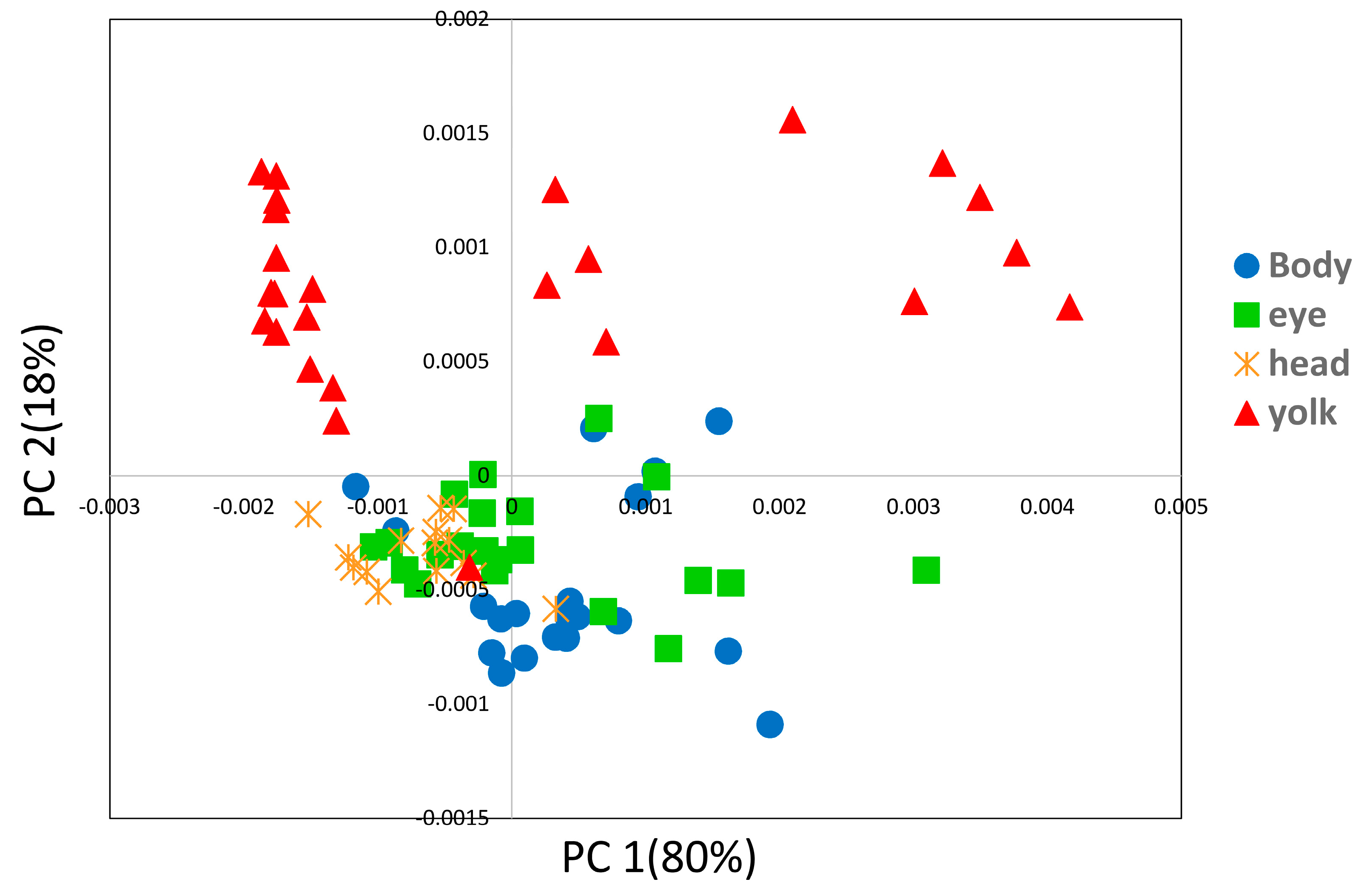
3. Results and Discussion
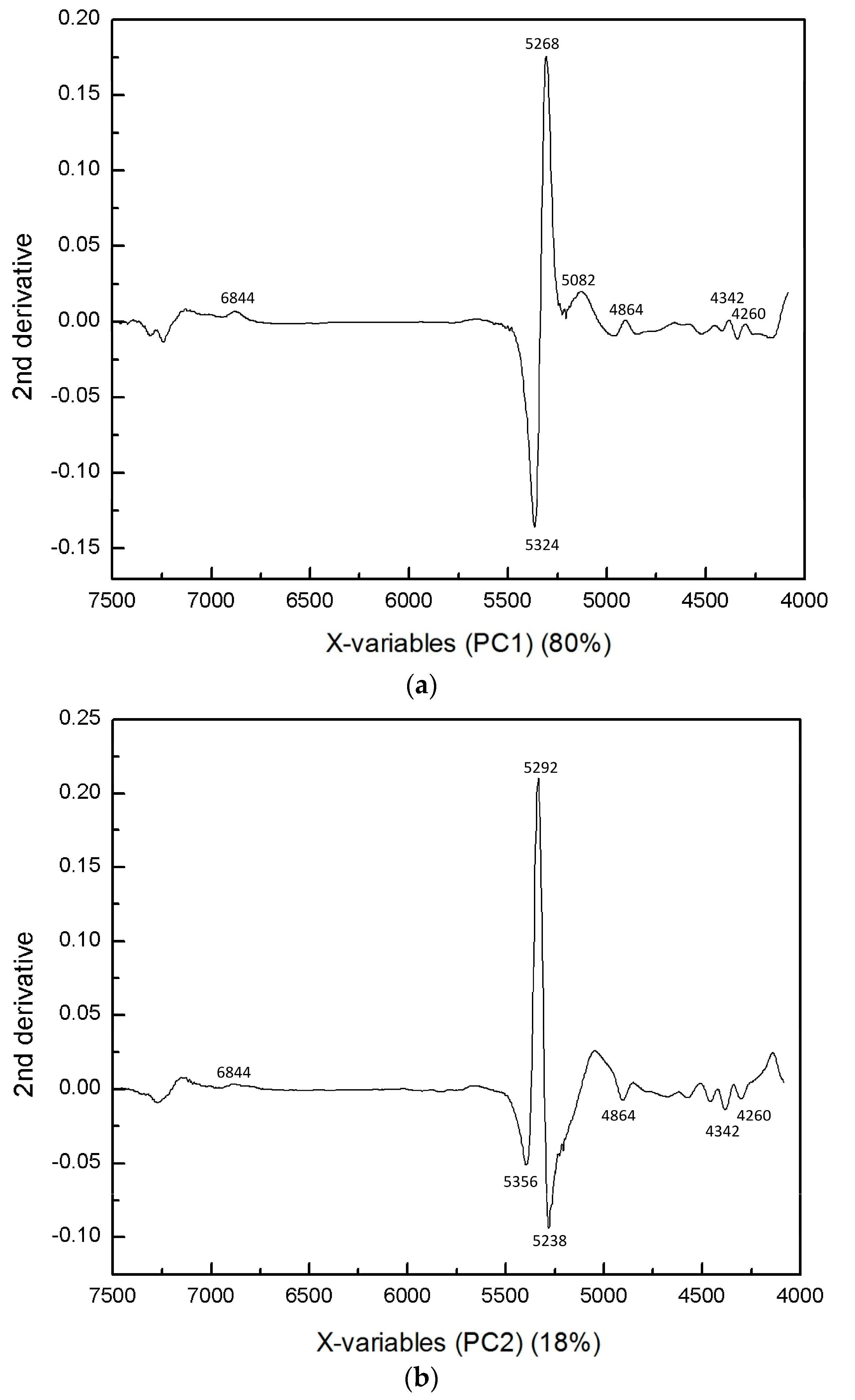
3.1. NIR Spectroscopy Study
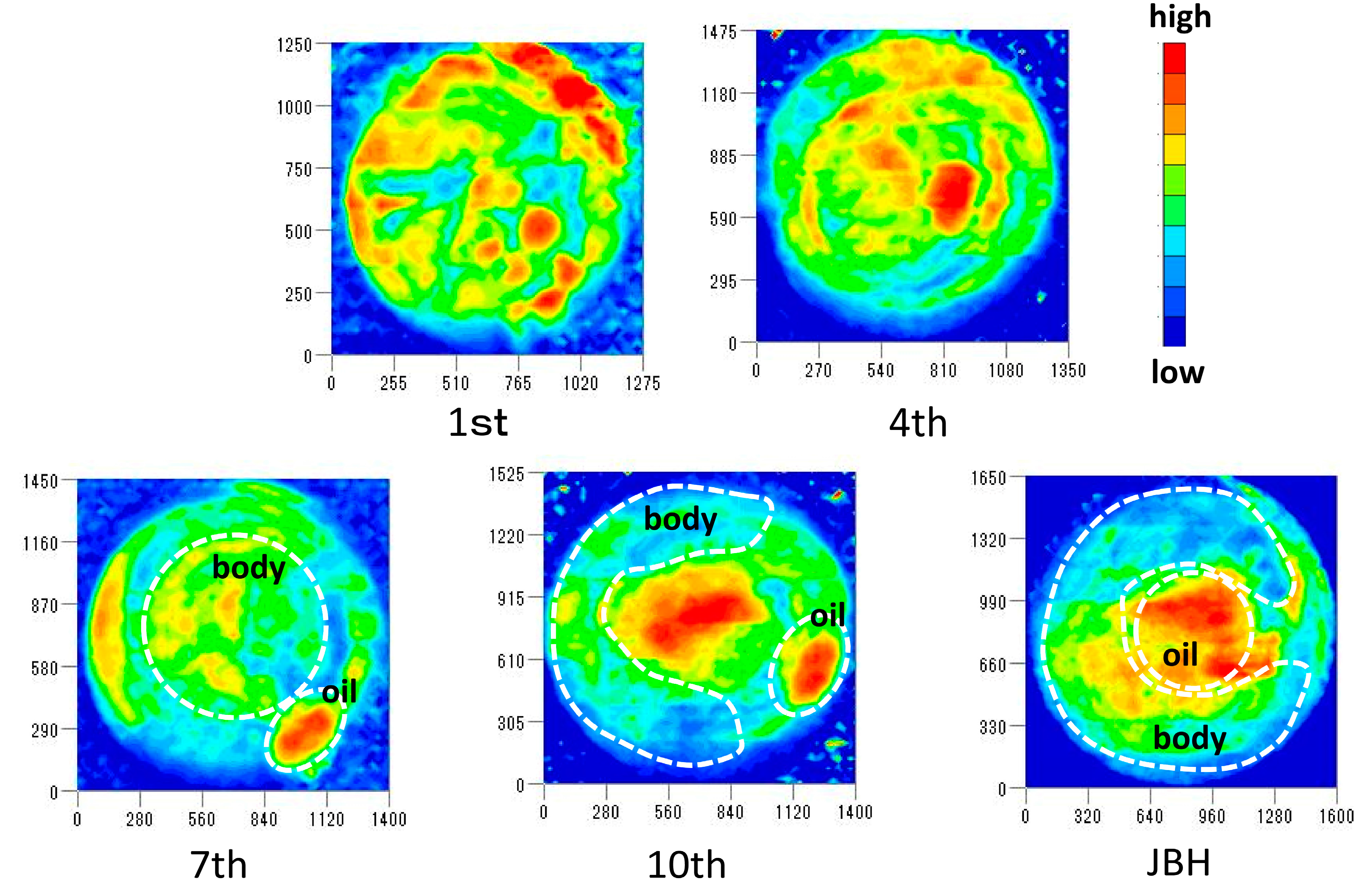
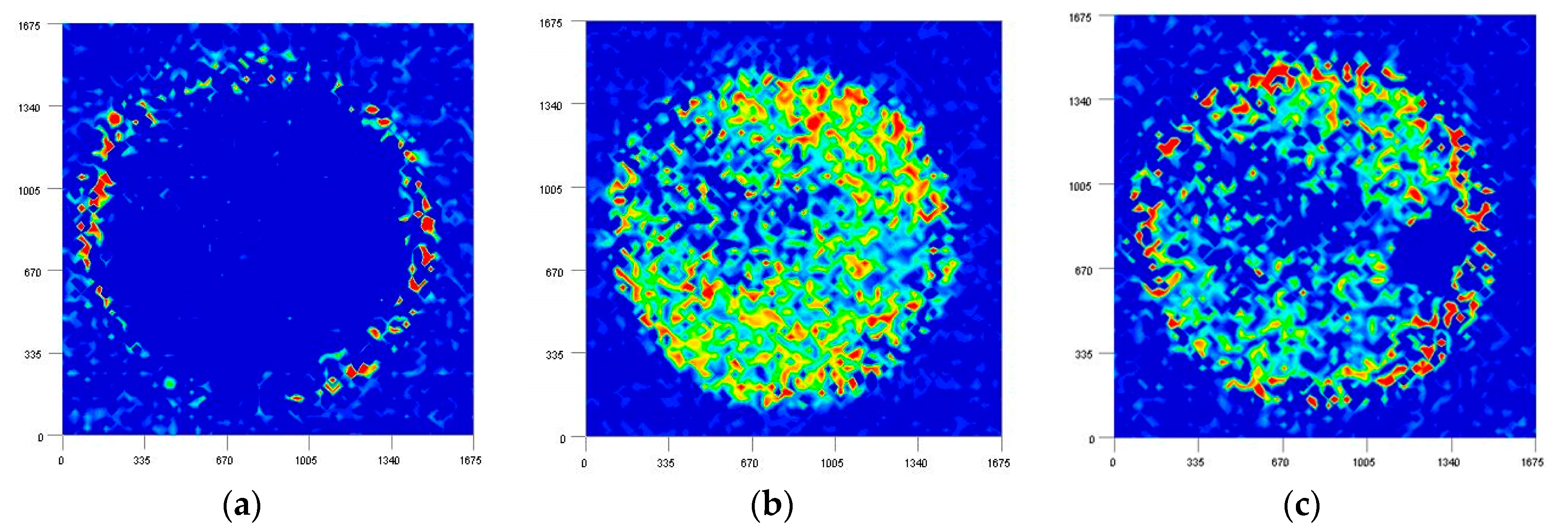
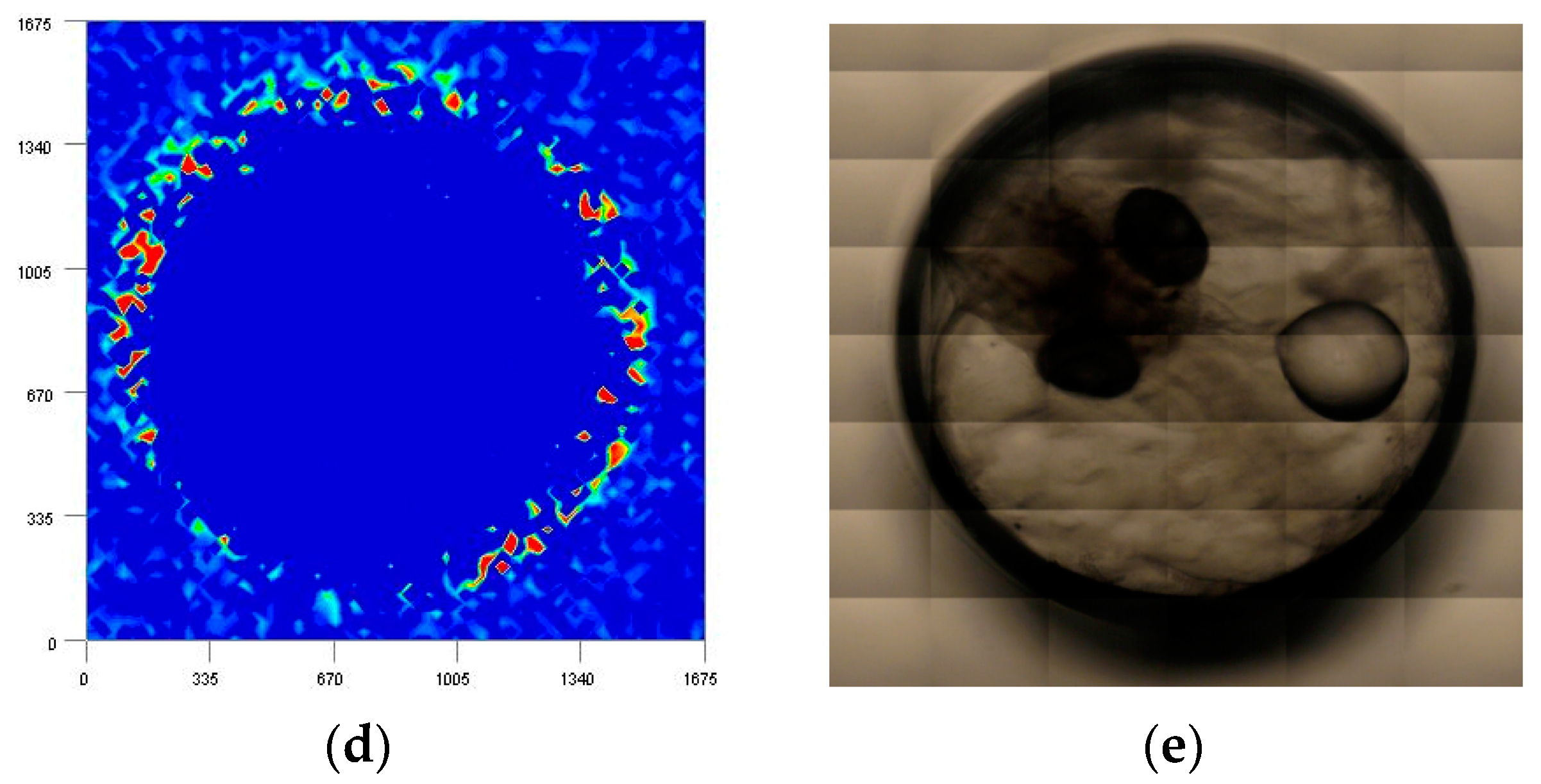
3.2. Imaging Study
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Watanabe, T.; Thayil, A.; Jesacher, A.; Grieve, K.; Debarre, D.; Wilson, T.; Martin, B.; Srinivas, S. Characterisation of the dynamic behaviour of lipid droplets in the early mouse embryo using adaptive harmonic generation microscopy. BMC Cell Biol. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Cashman, K.; Russell, D.L.; Thompson, J.G.; Norman, R.J.; Robker, R.L. Beta-oxidation is essential for mouse oocyte developmental competence and early embryo development. Biol. Reprod. 2010, 83, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Haggarty, P.; Wood, M.; Ferguson, E.; Hoad, G.; Srikantharajah, A.; Milne, E.; Hamilton, M.; Bhattacharya, S. Fatty acid metabolism in human preimplantation embryos. Hum. Reprod. 2006, 21, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, M.D. Composition, accumulation and utilization of yolk lipids in teleost fish. Rev. Fish Biol. Fish. 1996, 6, 259–286. [Google Scholar] [CrossRef]
- Bromage, N.; Jones, J.; Randall, C.; Thrush, M.; Davies, B.; Springate, J.; Duston, J.; Barker, G. Broodstock management, fecundity, egg quality and the timing of egg production in the rainbow trout (Oncorhynchus mykiss). Aquaculture 1992, 100, 141–166. [Google Scholar] [CrossRef]
- Latham, K.E.; Garrels, J.I.; Chang, C.; Solter, D. Quantitative analysis of protein synthesis in mouse embryos. I. Extensive reprogramming at the one-and two-cell stages. Development 1991, 112, 921–932. [Google Scholar] [PubMed]
- Giebelhaus, D.H.; Heikkila, J.J.; Schultz, G.A. Changes in the quantity of histone and actin messenger RNA during the development of preimplantation mouse embryos. Dev. Biol. 1983, 98, 148–154. [Google Scholar] [CrossRef]
- Scott, L.A.; Samuel, S. The successful use of pronuclear embryo transfers the day following oocyte retrieval. Hum. Reprod. 1998, 13, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Van Royen, E.; Mangelschots, K.; de Neubourg, D.; Valkenburg, M.; van de Meerssche, M.; Ryckaert, G.; Eestermans, W.; Gerris, J. Characterization of a top quality embryo, a step towards single-embryo transfer. Hum. Reprod. 1999, 14, 2345–2349. [Google Scholar] [CrossRef] [PubMed]
- Gerris, J.; de Neubourg, D.; Mangelschots, K.; Van Royen, E.; Van de Meerssche, M.; Valkenburg, M. Prevention of twin pregnancy after in vitro fertilization or intracytoplasmic sperm injection based on strict embryo criteria (a prospective randomized clinical trial). Hum. Reprod. 1999, 14, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schlenker, T.; Schoolcraft, W.B. Blastocyst score affects implantation and pregnancy outcome: Towards a single blastocyst transfer. Fertil. Steril. 2000, 73, 1155–1158. [Google Scholar] [CrossRef]
- Cummins, J.M.; Breen, T.M.; Harrison, K.L.; Shaw, J.M.; Wilson, L.M.; Hennessey, J.F. A formula for scoring human embryo growth rates in in vitro fertilization: Its value in predicting pregnancy and in comParison with visual estimates of embryo quality. J. Vitro Fertil. Embryo Transf. 1997, 3, 284–295. [Google Scholar] [CrossRef]
- Bromage, N.R.; Bruce, M.; Basavaraja, N.; Rana, K.; Shields, R.; Young, C.; Dye, J.; Smith, P.; Gamble, J. Egg quality determinants in finfish the role of overripening with special reference to the timing of stripping in the Atlantic halibut Hippoglossus hippoglossus. J. World Aquac. Soc. 1994, 25, 13–21. [Google Scholar] [CrossRef]
- Shields, R.J.; Brown, N.P.; Bromage, N.R. Blastomere morphology as a predictive measure of fish egg viability. Aquaculture 1997, 155, 1–12. [Google Scholar] [CrossRef]
- Ishigaki, M.; Kawasaki, S.; Ishikawa, D.; Ozaki, Y. Near-Infrared Spectroscopy and Imaging Studies of Fertilized Fish Eggs: In Vivo Monitoring of Egg Growth at the Molecular Level. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, D.; Shinzawa, H.; Genkawa, T.; Kazarian, S.G.; Ozaki, Y. Recent Progress of Near-Infrared (NIR) Imaging—Development of Novel Instruments and Their Applicability for Practical Situations. Anal. Sci. 2014, 30, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Shinzawa, H.; Nishida, M.; Tanaka, T.; Kanematsu, W. Accelerated weathering-induced degradation of poly (lactic acid) fiber studied by near-infrared (NIR) hyperspectral imaging. Appl. Spectrosc. 2012, 66, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, D.; Murayama, K.; Genkawa, T.; Awa, K.; Komiyama, M.; Ozaki, Y. Development of compact near infrared imaging device with high-speed and portability for pharmaceutical process monitoring. NIR News 2012, 23, 14–17. [Google Scholar] [CrossRef]
- Ishikawa, D.; Murayama, K.; Awa, K.; Genkawa, T.; Komiyama, M.; Kazarian, S.G.; Ozaki, Y. Application of a newly developed portable NIR imaging device to monitor the dissolution process of tablets. Anal. Bioanal. Chem. 2013, 405, 9401–9409. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, D.; Nishii, T.; Mizuno, F.; Kazarian, S.G.; Ozaki, Y. Development of a high-speed monitoring near infrared hyperspectral camera (Compovision) for wide area imaging and its applications. NIR News 2013, 24, 6–11. [Google Scholar] [CrossRef]
- Ishikawa, D.; Nishii, T.; Mizuno, F.; Sato, H.; Kazarian, S.G.; Ozaki, Y. Potential of a newly developed high-speed near-infrared (NIR) camera (Compovision) in polymer industrial analyses: Monitoring crystallinity and crystal evolution of polylactic acid (PLA) and concentration of PLA in PLA/Poly-(R)-3hydroxybutyrate (PHB) blends. Appl. Spectrosc. 2013, 67, 1441–1446. [Google Scholar] [PubMed]
- FurUKawa, T.; Sato, H.; Shinzawa, H.; Noda, I.; Ochiai, S. Evaluation of homogeneity of binary blends of poly (3-hydroxybutyrate) and poly (l-lactic acid) studied by near infrared chemical imaging (NIRCI). Anal. Sci. 2007, 23, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Siesler, H.W. Infrared and Raman Spectroscopic Imaging; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Lasch, P.; Haensch, W.; Naumann, D.; Diem, M. Imaging of colorectal adenocarcinoma using FT-IR microspectroscopy and cluster analysis. Biochim. Biophys. Acta Mol. Basis Dis. 2004, 1688, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.; Camacho, N.P. FT-IR imaging of native and tissue-engineered bone and cartilage. Biomaterials 2007, 28, 2465–2478. [Google Scholar] [CrossRef] [PubMed]
- Pallua, J.D.; Pezzei, C.; Zelger, B.; Schaefer, G.; Bittner, L.K.; Huck-Pezzei, V.A.; Schoenbichler, S.A.; Hahn, H.; Kloss-Brandstaetter, A.; Kloss, F.; et al. Fourier transform infrared imaging analysis in discrimination studies of squamous cell carcinoma. Analyst 2012, 137, 3965–3974. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, W.E.; Overstreet, R.M.; Fournie, J.W.; Walker, W.W. Development of aquarium fish models for environmental carcinogenesis: Tumor induction in seven species. J. Appl. Toxicol. 1985, 5, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, W.E.; Walker, W.W.; Fournie, J.W.; Manning, C.S.; Krol, R.M. Use of the Japanese medaka (Oryzias latipes) and guppy (Poecilia reticulata) in carcinogenesis testing under national toxicology program protocols. Toxicol. Pathol. 2003, 31, 88–91. [Google Scholar] [PubMed]
- Shima, A.; Mitani, H. Medaka as a research organism: Past, present and future. Mech. Dev. 2004, 121, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Ishigaki, M.; Taketani, A.; Sato, H. Discrimination of fish egg quality and viability by Raman spectroscopy. Anal. Methods 2014, 6, 9206–9211. [Google Scholar] [CrossRef]
- Holliday, F.G.T.; Jones, M.P. Some effects of salinity on the developing eggs and larvae of the plaice (Pleuronectes platessa). J. Mar. Biol. Assoc. UK 1967, 47, 39–48. [Google Scholar] [CrossRef]
- Vetter, R.D.; Hodson, R.E.; Arnold, C. Energy metabolism in a rapidly developing marine fish egg, the red drum (Sciaenops ocellata). Can. J. Fish. Aquat. Sci. 1983, 40, 627–634. [Google Scholar] [CrossRef]
- Siesler, H.W.; Ozaki, Y.; Kawata, S.; Heise, H.M. Near-Infrared Spectroscopy: Principles, Instruments, Applications; John Wiley & Sons: New York, NY, USA, 2008. [Google Scholar]
- Ozaki, Y.; McClure, W.F.; Christy, A.A. Near-Infrared Spectroscopy in Food Science and Technology; John Wiley & Sons: New York, NY, USA, 2006. [Google Scholar]
- Šašic, S.; Segtnan, V.H.; Ozaki, Y. Self-modeling curve resolution study of temperature-dependent near-infrared spectra of water and the investigation of water structure. J. Phys. Chem. A 2002, 106, 760–766. [Google Scholar] [CrossRef]
- Czarnik-Matusewicz, B.; Pilorz, S.; Hawranek, J.P. Temperature-dependent water structural transitions examined by near-IR and mid-IR spectra analyzed by multivariate curve resolution and two-dimensional correlation spectroscopy. Anal. Chim. Acta 2005, 544, 15–25. [Google Scholar] [CrossRef]
- Izutsu, K.I.; Fujimaki, Y.; Kuwabara, A.; Hiyama, Y.; Yomota, C.; Aoyagi, N. Near-infrared analysis of protein secondary structure in aqueous solutions and freeze-dried solids. J. Pharm. Sci. 2006, 95, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Workman, J., Jr.; Weyer, L. Practical Guide and Spectral Atlas for Interpretive Near-Infrared Spectroscopy; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Grigorieff, N.; Ceska, T.A.; Downing, K.H.; Baldwin, J.M.; Henderson, R. Electron-crystallographic refinement of the structure of bacteriorhodopsin. J. Mol. Biol. 1996, 2593, 393–421. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.S.; Schulz, G.E. Structure of porin refined at 1.8 Å resolution. J. Mol. Biol. 1992, 227, 493–509. [Google Scholar] [CrossRef]
- Sample Availability: Not Available.

© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishigaki, M.; Yasui, Y.; Puangchit, P.; Kawasaki, S.; Ozaki, Y. In Vivo Monitoring of the Growth of Fertilized Eggs of Medaka Fish (Oryzias latipes) by Near-Infrared Spectroscopy and Near-Infrared Imaging—A Marked Change in the Relative Content of Weakly Hydrogen-Bonded Water in Egg Yolk Just before Hatching. Molecules 2016, 21, 1003. https://doi.org/10.3390/molecules21081003
Ishigaki M, Yasui Y, Puangchit P, Kawasaki S, Ozaki Y. In Vivo Monitoring of the Growth of Fertilized Eggs of Medaka Fish (Oryzias latipes) by Near-Infrared Spectroscopy and Near-Infrared Imaging—A Marked Change in the Relative Content of Weakly Hydrogen-Bonded Water in Egg Yolk Just before Hatching. Molecules. 2016; 21(8):1003. https://doi.org/10.3390/molecules21081003
Chicago/Turabian StyleIshigaki, Mika, Yui Yasui, Paralee Puangchit, Shoya Kawasaki, and Yukihiro Ozaki. 2016. "In Vivo Monitoring of the Growth of Fertilized Eggs of Medaka Fish (Oryzias latipes) by Near-Infrared Spectroscopy and Near-Infrared Imaging—A Marked Change in the Relative Content of Weakly Hydrogen-Bonded Water in Egg Yolk Just before Hatching" Molecules 21, no. 8: 1003. https://doi.org/10.3390/molecules21081003
APA StyleIshigaki, M., Yasui, Y., Puangchit, P., Kawasaki, S., & Ozaki, Y. (2016). In Vivo Monitoring of the Growth of Fertilized Eggs of Medaka Fish (Oryzias latipes) by Near-Infrared Spectroscopy and Near-Infrared Imaging—A Marked Change in the Relative Content of Weakly Hydrogen-Bonded Water in Egg Yolk Just before Hatching. Molecules, 21(8), 1003. https://doi.org/10.3390/molecules21081003