
A High-Throughput Automated Microfluidic Platform for Calcium Imaging of Taste Sensing



Abstract
:1. Introduction
2. Results
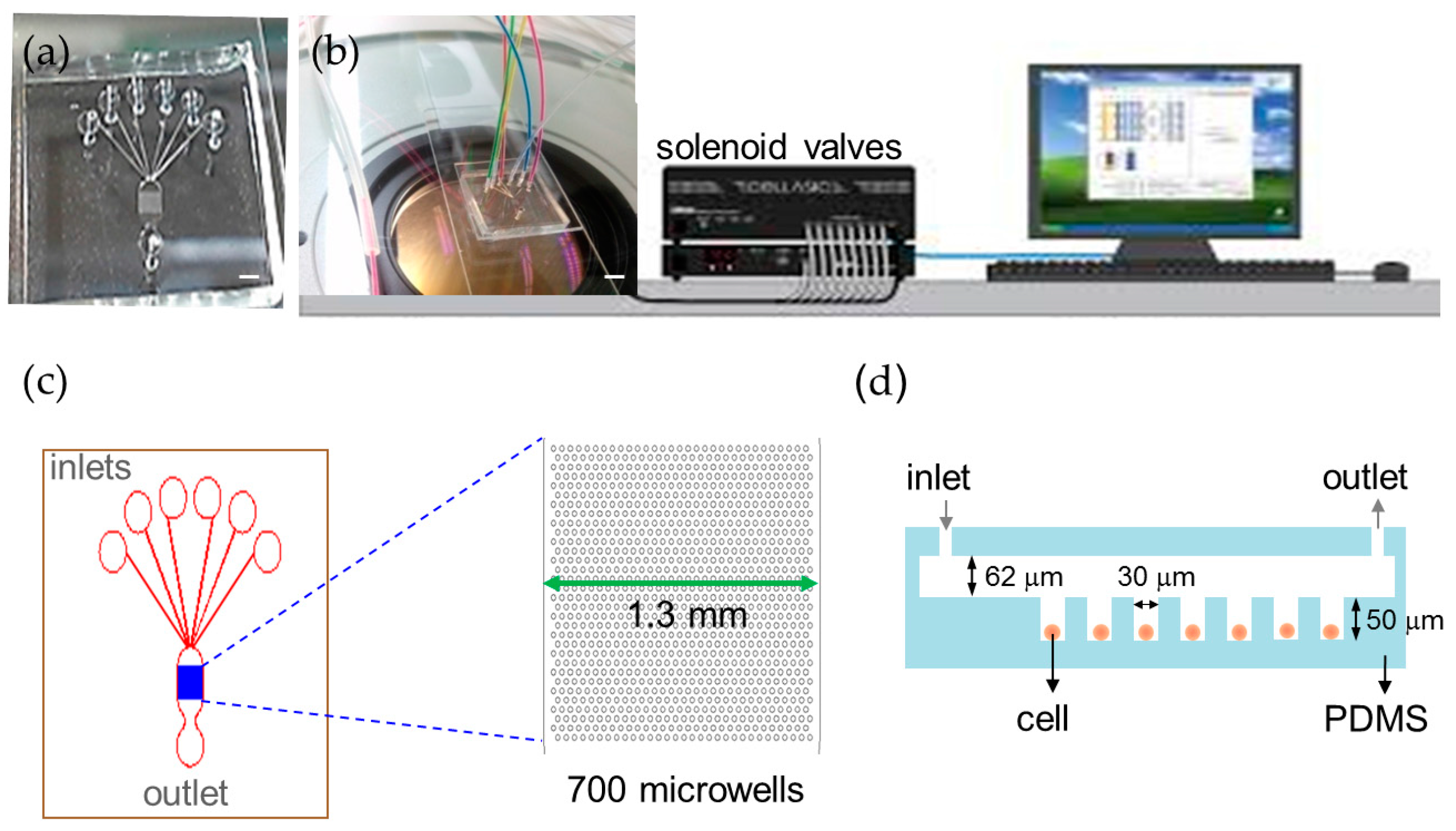
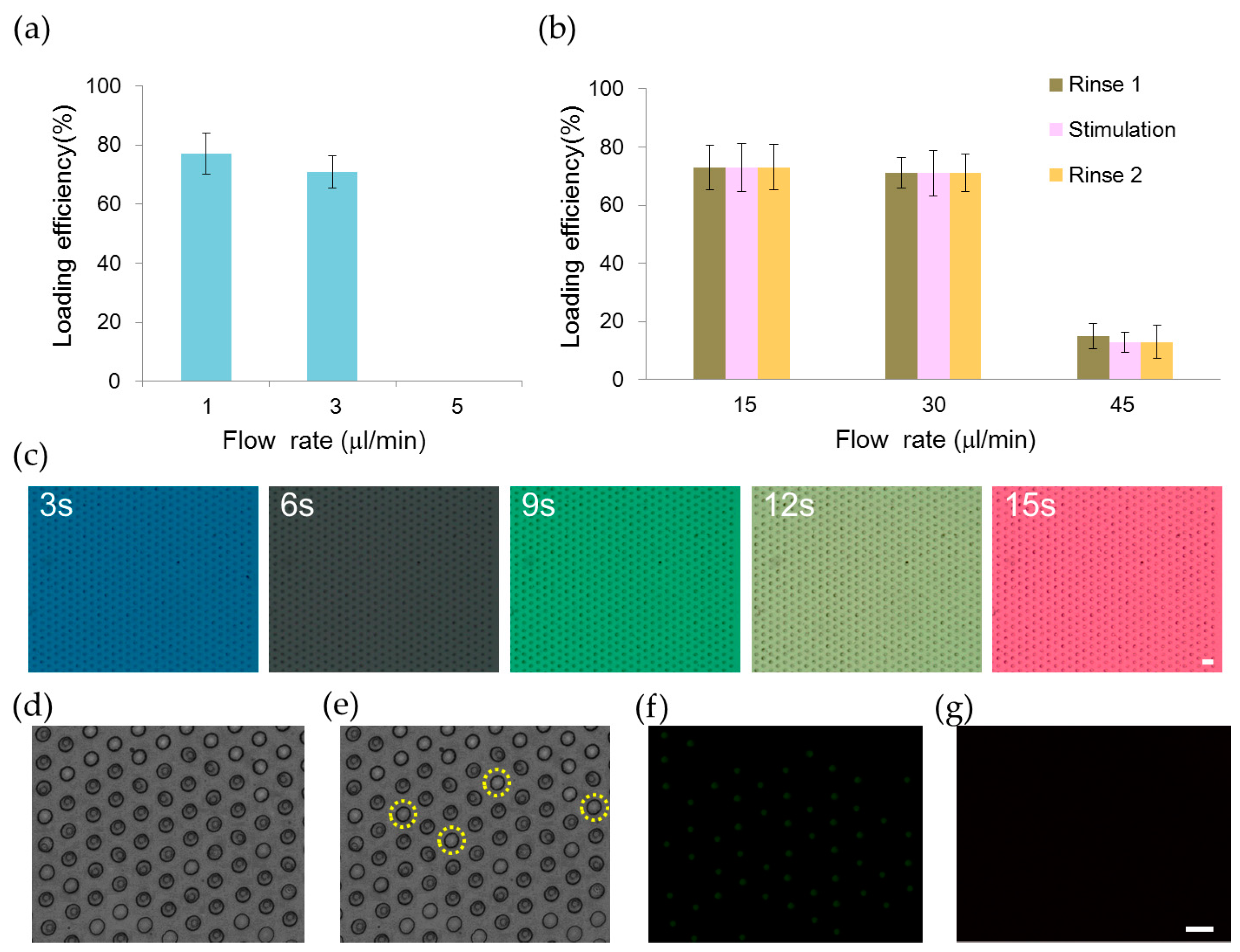
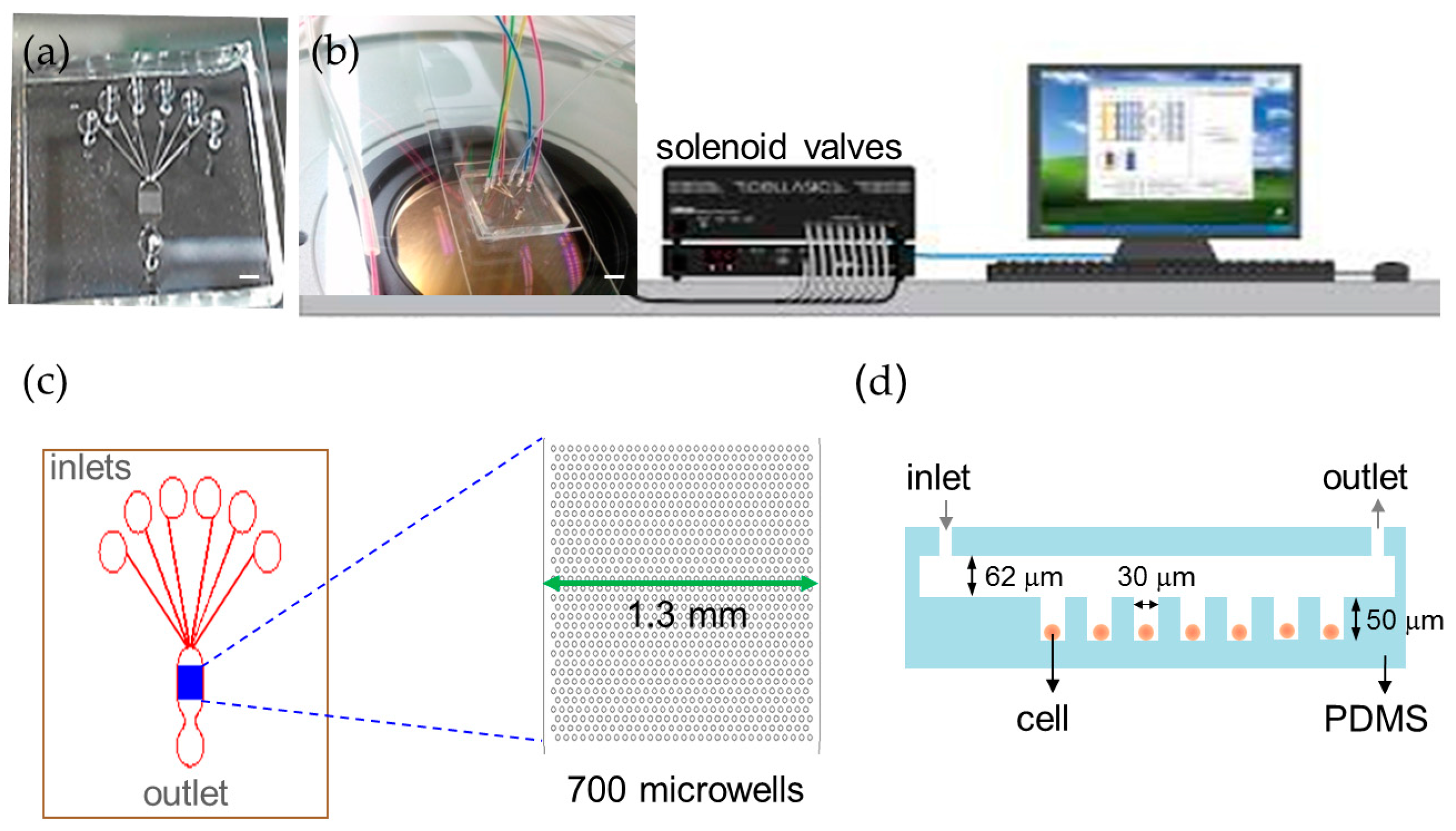
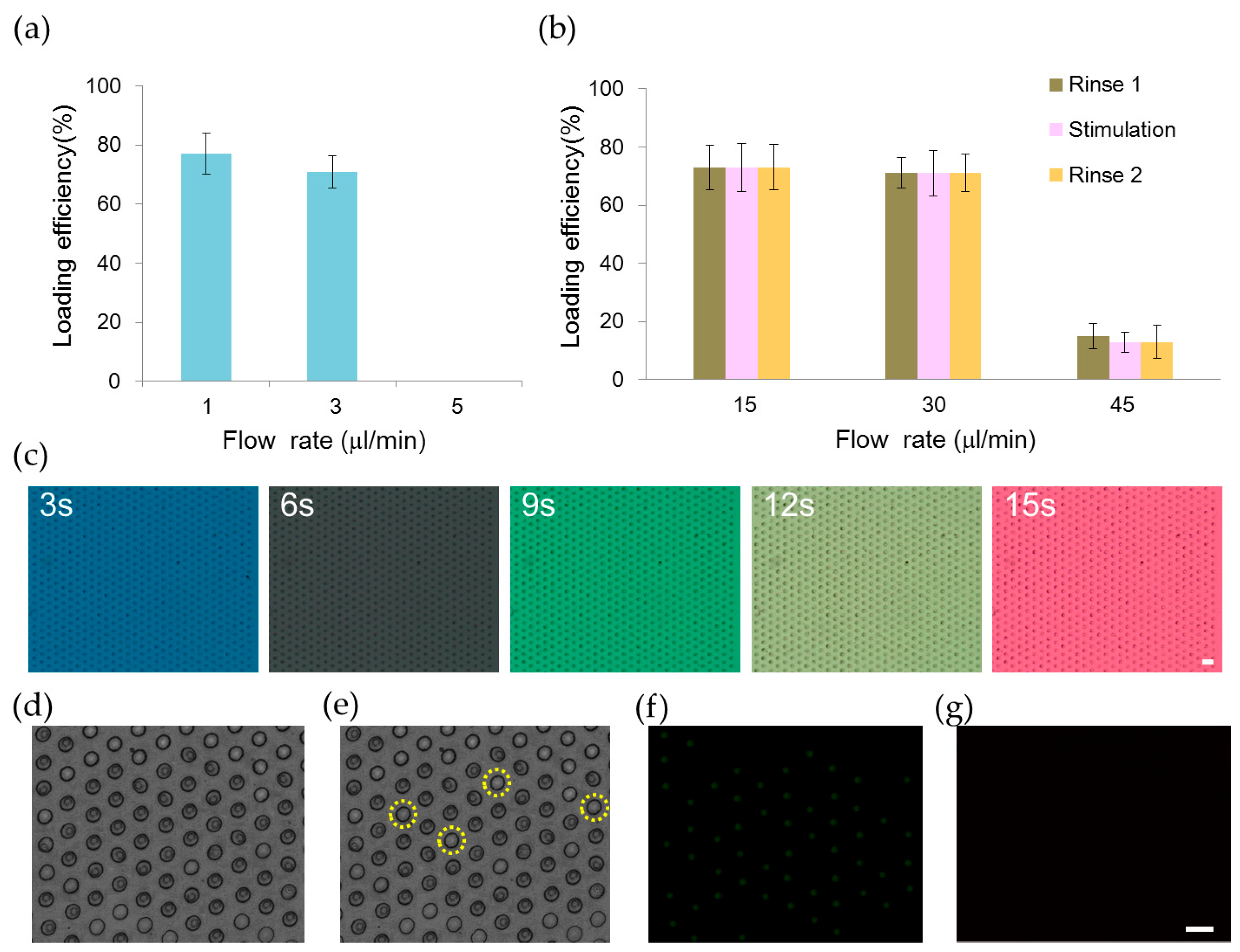
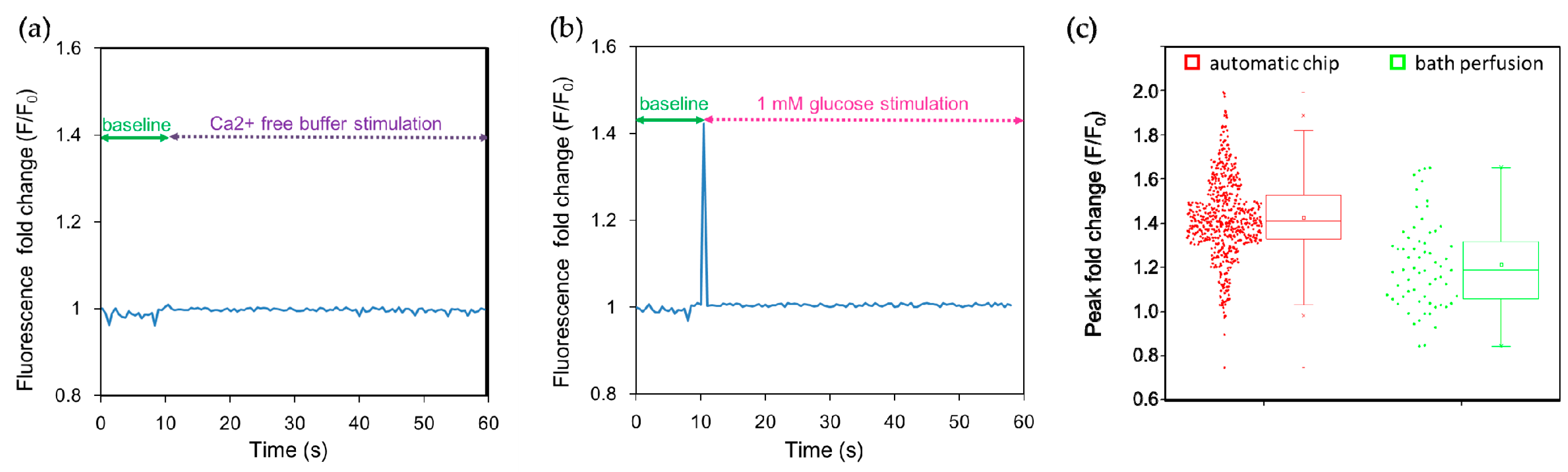
2.1. Trapping and Perfusion of NCI-H716 Cells
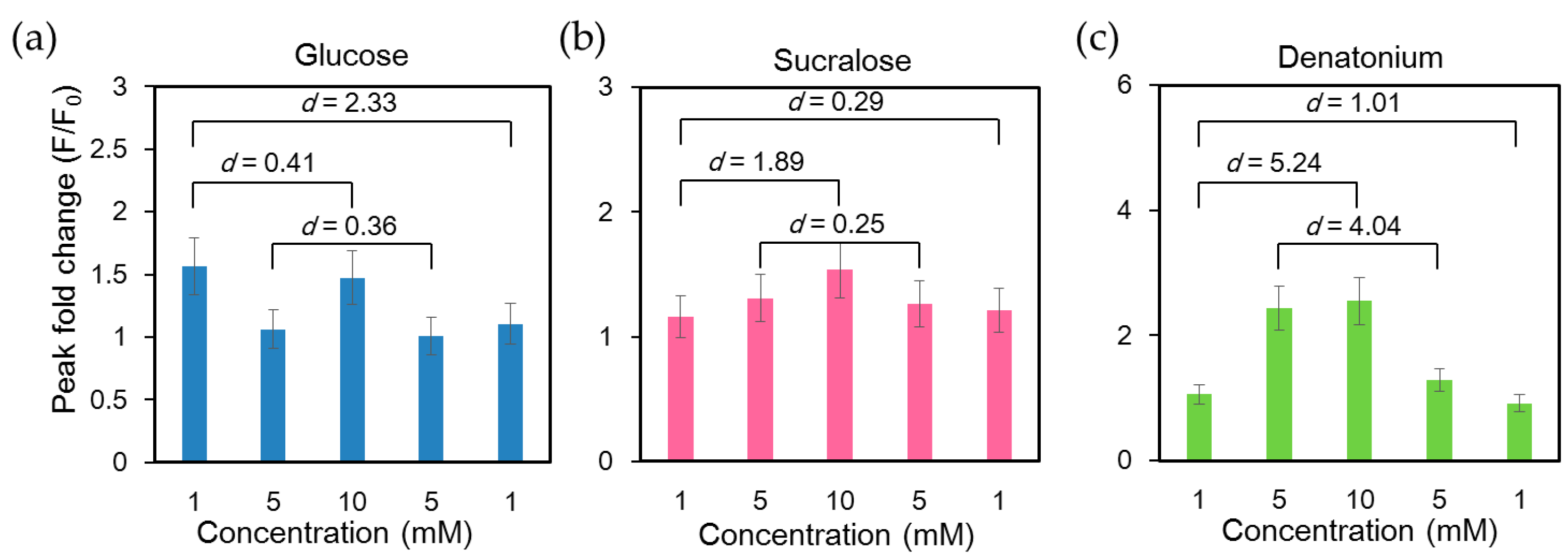
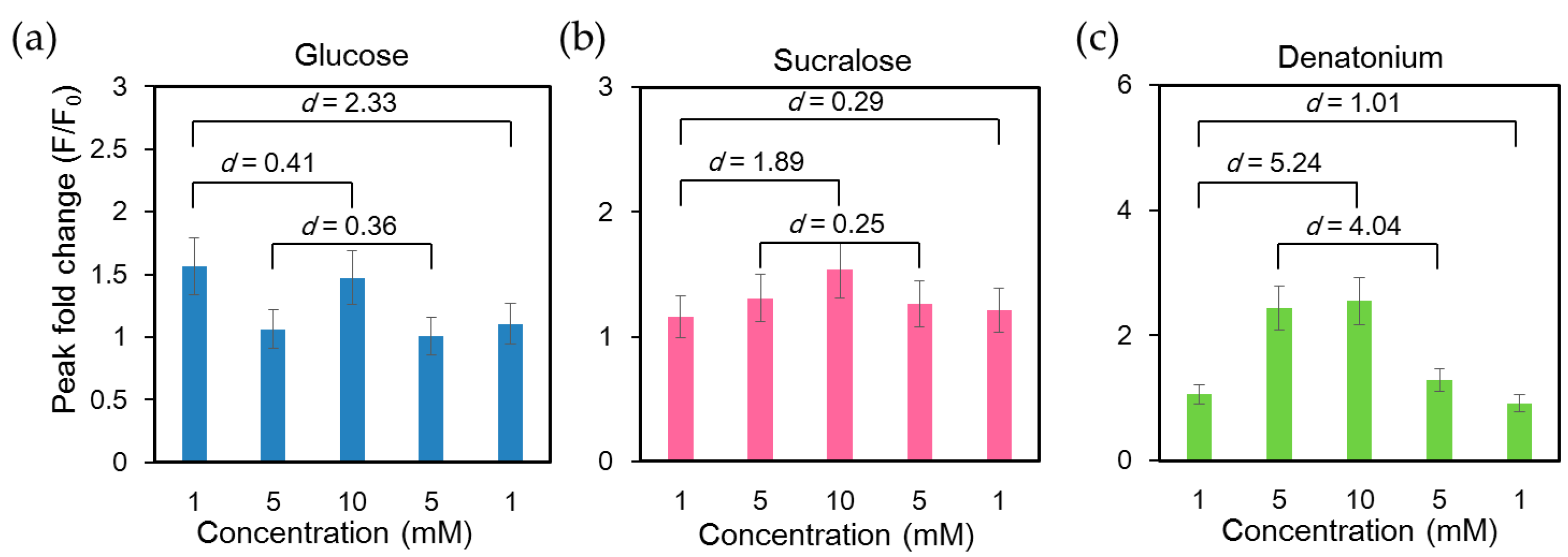
2.2 Stimulation of NCI-H716 Cells with Sweet Tastants
2.3 Order of Application of Tastant Stimuli
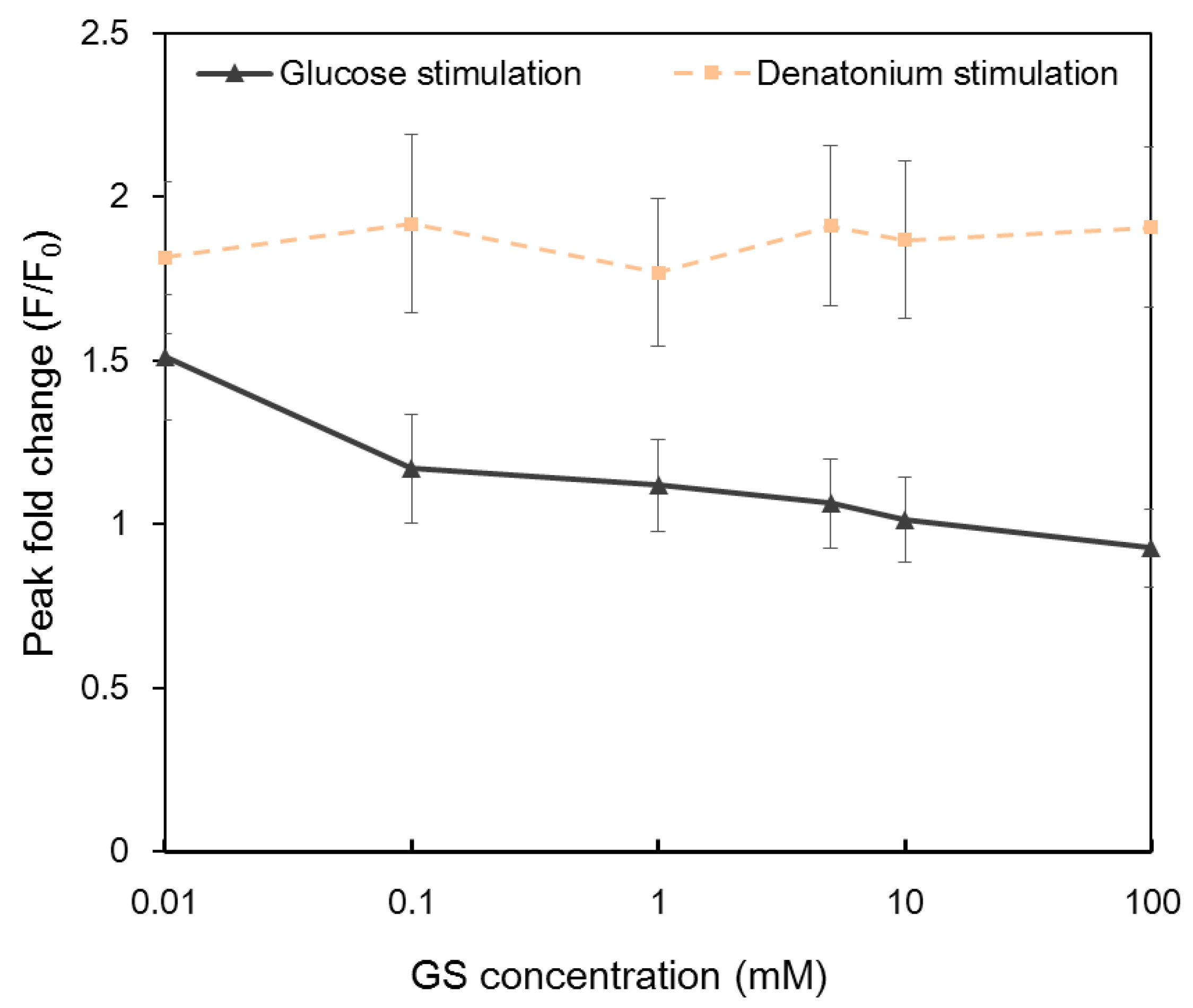
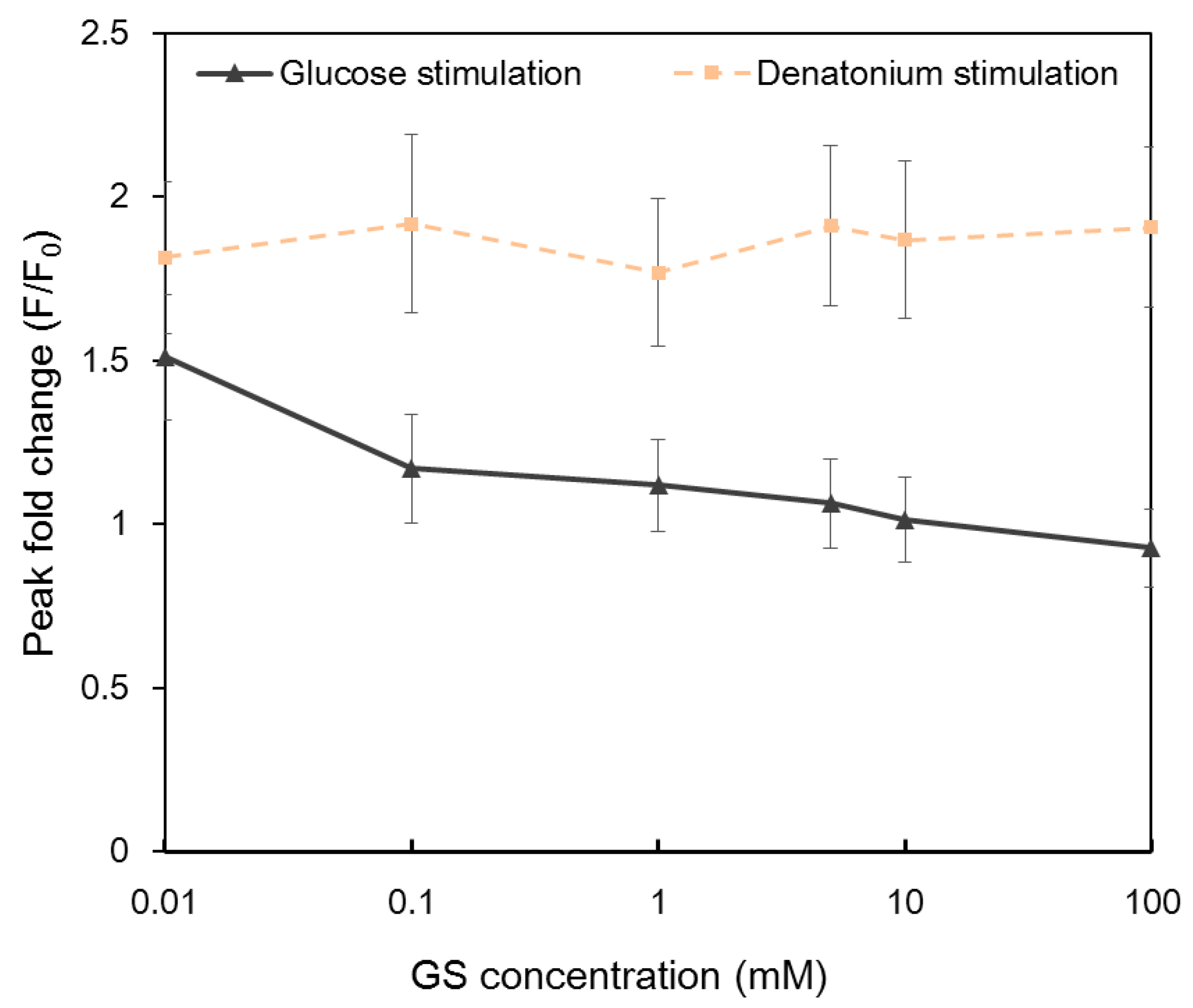
2.4 Calcium Responses to Glucose and Denatonium Benzoate after Gymnema Sylvestre (GS) Treatment
3. Discussion
4. Materials and Methods
4.1 Device Fabrication and Operation
4.2 Culture of NCI-H716 Cells
4.3 Stimulation of Tastants and Calcium Imaging on NCI-H716 Cells
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Trivedi, B.P. Neuroscience: Hardwired for taste. Nature 2012, 486, S7–S9. [Google Scholar] [CrossRef] [PubMed]
- Finger, T.E.; Kinnamon, S.C. Taste isn’t just for taste buds anymore. F1000 Biol. Rep. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Sarr, M.G. Effect of the artificial sweetener, acesulfame potassium, a sweet taste receptor agonist, on glucose uptake in small intestinal cell lines. J. Gastrointest. Surg. 2013, 17, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, G.A.; Soundarapandian, M.M.; Tyrberg, B. Sweet taste receptor signaling in beta cells mediates fructose-induced potentiation of glucose-stimulated insulin secretion. Proc. Natl. Acad. Sci. USA 2012, 109, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Simon, B.R.; Learman, B.S.; Parlee, S.D.; Scheller, E.L.; Mori, H.; Cawthorn, W.P.; Ning, X.; Krishnan, V.; Ma, Y.L.; Tyrberg, B.; et al. Sweet taste receptor deficient mice have decreased adiposity and increased bone mass. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Young, R.L. Sensing via intestinal sweet taste pathways. Front. Neurosci. 2011, 5. [Google Scholar] [CrossRef] [PubMed]
- Kokrashvili, Z.; Mosinger, B.; Margolskee, R.F. Taste signaling elements expressed in gut enteroendocrine cells regulate nutrient-responsive secretion of gut hormones. Am. J. Clin. Nutr. 2009, 90, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Howitt, M.R.; Lavoie, S.; Michaud, M.; Blum, A.M.; Tran, S.V.; Weinstock, J.V.; Gallini, C.A.; Redding, K.; Margolskee, R.F.; Osborne, L.C.; et al. Tuft cells, taste-chemosensory cells, orchestrate parasite type 2 immunity in the gut. Science 2016, 351, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Baquero, A.F.; Gilbertson, T.A. Insulin activates epithelial sodium channel (enac) via phosphoinositide 3-kinase in mammalian taste receptor cells. Am. J. Physiol. Cell Physiol. 2011, 300, 860–871. [Google Scholar] [CrossRef] [PubMed]
- Renwick, A.G.; Molinary, S.V. Sweet-taste receptors, low-energy sweeteners, glucose absorption and insulin release. Br. J. Nutr. 2010, 104, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, N.; Roper, S.D. The cell biology of taste. J. Cell Biol. 2010, 190, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Egan, J.M.; Margolskee, R.F. Taste cells of the gut and gastrointestinal chemosensation. Mol. Interv. 2008, 8, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Glendinning, J.I.; Stano, S.; Holter, M.; Azenkot, T.; Goldman, O.; Margolskee, R.F.; Vasselli, J.R.; Sclafani, A. Sugar-induced cephalic-phase insulin release is mediated by a T1R2 + T1R3-independent taste transduction pathway in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R552–R560. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, G.A.; Smith, K.R.; Tyrberg, B.; Hussain, T.; Pratley, R.E. Sweet taste receptors regulate basal insulin secretion and contribute to compensatory insulin hypersecretion during the development of diabetes in male mice. Endocrinology 2014, 155, 2112–2121. [Google Scholar] [CrossRef] [PubMed]
- Oya, M.; Suzuki, H.; Watanabe, Y.; Sato, M.; Tsuboi, T. Amino acid taste receptor regulates insulin secretion in pancreatic beta-cell line min6 cells. Genes Cells 2011, 16, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, K.J.; Gillespie, P.G. Monitoring intracellular calcium ion dynamics in hair cell populations with fluo-4 am. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Gerspach, A.C.; Gutmann, H.; Asarian, L.; Drewe, J.; Beglinger, C. The functional involvement of gut-expressed sweet taste receptors in glucose-stimulated secretion of glucagon-like peptide-1 (glp-1) and peptide yy (pyy). Clin. Nutr. 2011, 30, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Maruoka, D.; Arai, M.; Tanaka, T.; Okimoto, K.; Oyamada, A.; Minemura, S.; Tsuboi, M.; Matsumura, T.; Nakagawa, T.; Kanda, T.; et al. Mosapride citrate increases postprandial glucagon-like peptide-1, insulin, and gene expression of sweet taste receptors. Dig. Dis. Sci. 2015, 60, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, R.A.; Nathan, M.S.; Beattie, P. Oral agents for the treatment of type 2 diabetes mellitus: Pharmacology, toxicity, and treatment. Ann. Emerg. Med. 2001, 38, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, J.R.; Chisholm, D.J. How sweet it is: Intestinal sweet taste receptors in type 2 diabetes. Diabetes 2013, 62, 3336–3337. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Egan, J.M.; Jang, H.J. Denatonium induces secretion of glucagon-like peptide-1 through activation of bitter taste receptor pathways. Diabetologia 2014, 57, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Avau, B.; Rotondo, A.; Thijs, T.; Andrews, C.N.; Janssen, P.; Tack, J.; Depoortere, I. Targeting extra-oral bitter taste receptors modulates gastrointestinal motility with effects on satiation. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Duskova, M.; Macourek, M.; Sramkova, M.; Hill, M.; Starka, L. The role of taste in cephalic phase of insulin secretion. Prague Med. Rep. 2013, 114, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, S.J.; Naim, M.; Zehavi, U.; Lindemann, B. Changes in IP3 and cytosolic Ca2+ in response to sugars and non-sugar sweeteners in transduction of sweet taste in the rat. J. Physiol. Lond. 1996, 490, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Ogura, T.; Margolskee, R.F.; Kinnamon, S.C. Taste receptor cell responses to the bitter stimulus denatonium involve Ca2+ influx via store-operated channels. J. Neurophysiol. 2002, 87, 3152–3155. [Google Scholar] [PubMed]
- Yamamoto, K.; Ishimaru, Y. Oral and extra-oral taste perception. Semin. Cell Dev. Biol. 2013, 24, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, Y.; Nakagawa, Y.; Nagasawa, M.; Takeda, S.; Arakawa, H.; Kojima, I. Diverse signaling systems activated by the sweet taste receptor in human Glp-1-secreting cells. Mol. Cell Endocrinol. 2014, 394, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Avau, B.; Depoortere, I. The bitter truth about bitter taste receptors: Beyond sensing bitter in the oral cavity. Acta Physiol. 2016, 216, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Faley, S.; Seale, K.; Hughey, J.; Schaffer, D.K.; VanCompernolle, S.; McKinney, B.; Baudenbacher, F.; Unutmaz, D.; Wikswo, J.P. Microfluidic platform for real-time signaling analysis of multiple single t cells in parallel. Lab Chip 2008, 8, 1700–1712. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, P.C. Microfluidic selection and retention of a single cardiac myocyte, on-chip dye loading, cell contraction by chemical stimulation, and quantitative fluorescent analysis of intracellular calcium. Anal. Chem. 2005, 77, 4315–4322. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, J.; Tibbits, G.F.; Li, P.C. Real-time monitoring of intracellular calcium dynamic mobilization of a single cardiomyocyte in a microfluidic chip pertaining to drug discovery. Electrophoresis 2007, 28, 4723–4733. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, T.V.; Bazopoulou, D.; Chronis, N. An automated microfluidic platform for calcium imaging of chemosensory neurons in caenorhabditis elegans. Lab Chip 2010, 10, 2758–2763. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.; Bushell, T.J.; Zagnoni, M. Chemically induced synaptic activity between mixed primary hippocampal co-cultures in a microfluidic system. Integr. Biol. 2014, 6, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Renault, R.; Sukenik, N.; Descroix, S.; Malaquin, L.; Viovy, J.L.; Peyrin, J.M.; Bottani, S.; Monceau, P.; Moses, E.; Vignes, M. Combining microfluidics, optogenetics and calcium imaging to study neuronal communication in vitro. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Weir, K.; Easton, C.; Huynh, W.; Moody, W.J.; Folch, A. A microfluidic microelectrode array for simultaneous electrophysiology, chemical stimulation, and imaging of brain slices. Lab Chip 2013, 13, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, X.A.; Cooksey, G.A.; Votaw, S.V.; Horowitz, L.F.; Folch, A. Large-scale investigation of the olfactory receptor space using a microfluidic microwell array. Lab Chip 2010, 10, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Yang, H.J.; Lee, I.S.; Kim, K.H.; Park, J.; Jeong, H.S.; Kim, Y.; Ahn, K.S.; Na, Y.C.; Jang, H.J. The aglycone of ginsenoside Rg3 enables glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in type 2 diabetic mice. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Nagasawa, M.; Medina, J.; Kojima, I. Glucose evokes rapid Ca2+ and cyclic amp signals by activating the cell-surface glucose-sensing receptor in pancreatic beta-cells. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Tomonari, H.; Miura, H.; Nakayama, A.; Matsumura, E.; Ooki, M.; Ninomiya, Y.; Harada, S. Galpha-gustducin is extensively coexpressed with sweet and bitter taste receptors in both the soft palate and fungiform papillae but has a different functional significance. Chem. Senses 2012, 37, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Grapengiesser, E.; Gylfe, E.; Hellman, B. Glucose-induced oscillations of cytoplasmic Ca2+ in the pancreatic beta-cell. Biochem. Biophys. Res. Commun. 1988, 151, 1299–1304. [Google Scholar] [CrossRef]
- Shiyovich, A.; Sztarkier, I.; Nesher, L. Toxic hepatitis induced by gymnema sylvestre, a natural remedy for type 2 diabetes mellitus. Am. J. Med. Sci. 2010, 340, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, A.; Driscoll, M.; Brent, R. Using measures of single-cell physiology and physiological state to understand organismic aging. Aging Cell 2016, 15, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Taheri-Araghi, S.; Brown, S.D.; Sauls, J.T.; McIntosh, D.B.; Jun, S. Single-cell physiology. Annu. Rev. Biophys. 2015, 44, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Eiriz, M.F.; Grade, S.; Rosa, A.; Xapelli, S.; Bernardino, L.; Agasse, F.; Malva, J.O. Functional evaluation of neural stem cell differentiation by single cell calcium imaging. Curr. Stem Cell Res. Ther. 2011, 6, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Badea, T.; Goldberg, J.; Mao, B.; Yuste, R. Calcium imaging of epileptiform events with single-cell resolution. J. Neurobiol. 2001, 48, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Marsh, K.A. Single-cell calcium imaging. Methods Mol. Biol. 1995, 41, 229–238. [Google Scholar] [PubMed]
- Patel, T.P.; Man, K.; Firestein, B.L.; Meaney, D.F. Automated quantification of neuronal networks and single-cell calcium dynamics using calcium imaging. J. Neurosci. Methods 2015, 243, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Hamano, K.; Nakagawa, Y.; Ohtsu, Y.; Li, L.; Medina, J.; Tanaka, Y.; Masuda, K.; Komatsu, M.; Kojima, I. Lactisole inhibits the glucose-sensing receptor T1R3 expressed in mouse pancreatic beta-cells. J. Endocrinol. 2015, 226, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.X.; Wu, W.; Zhang, W.B.; Deng, S.P. Thermodynamics of the interaction of sweeteners and lactisole with fullerenols as an artificial sweet taste receptor model. Food Chem. 2011, 128, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Theodorakis, M.J. Comment on faerch et al. Glp-1 response to oral glucose is reduced in prediabetes, screen-detected type 2 diabetes, and obesity and influenced by sex: The addition-pro study. Diabetes 2015, 64, 2513–2525. [Google Scholar] [CrossRef] [PubMed]
- Temizkan, S.; Deyneli, O.; Yasar, M.; Arpa, M.; Gunes, M.; Yazici, D.; Sirikci, O.; Haklar, G.; Imeryuz, N.; Yavuz, D.G. Sucralose enhances Glp-1 release and lowers blood glucose in the presence of carbohydrate in healthy subjects but not in patients with type 2 diabetes. Eur. J. Clin. Nutr. 2015, 69, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Casamitjana, R.; Viaplana-Masclans, J.; Lacy, A.; Vidal, J. Glp-1 action and glucose tolerance in subjects with remission of type 2 diabetes after gastric bypass surgery. Diabetes Care 2013, 36, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Pirhonen, E.; Moimas, L.; Brink, M. Mechanical properties of bioactive glass 9–93 fibres. Acta Biomater. 2006, 2, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.V.; Beierlein, M.; Morgan, J.L.; Rothe, A.; Gee, K.R. Novel fluo-4 analogs for fluorescent calcium measurements. Cell Calcium 2004, 36, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Statistical power analysis. Curr. Dir. Psychol. Sci. 1992, 1, 98–101. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calcium Imaging Responses | Peak Fold Change | p-Value 1 | Cohen’s Effect Size (d) | Range | Quartile Coefficient of Dispersion |
|---|---|---|---|---|---|
| Automatic chip (n = 1382) | 1.39 ± 0.27 | <0.001 | 0.65 | 4.32 | 7.7% |
| Bath perfusion (n = 64) | 1.21 ± 0.20 | 0.81 | 10.7% |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, Y.-H.; Hsu, C.-H.; Chen, C. A High-Throughput Automated Microfluidic Platform for Calcium Imaging of Taste Sensing. Molecules 2016, 21, 896. https://doi.org/10.3390/molecules21070896
Hsiao Y-H, Hsu C-H, Chen C. A High-Throughput Automated Microfluidic Platform for Calcium Imaging of Taste Sensing. Molecules. 2016; 21(7):896. https://doi.org/10.3390/molecules21070896
Chicago/Turabian StyleHsiao, Yi-Hsing, Chia-Hsien Hsu, and Chihchen Chen. 2016. "A High-Throughput Automated Microfluidic Platform for Calcium Imaging of Taste Sensing" Molecules 21, no. 7: 896. https://doi.org/10.3390/molecules21070896
APA StyleHsiao, Y.-H., Hsu, C.-H., & Chen, C. (2016). A High-Throughput Automated Microfluidic Platform for Calcium Imaging of Taste Sensing. Molecules, 21(7), 896. https://doi.org/10.3390/molecules21070896