
Biochemical Constituents and in Vitro Antioxidant and Anticholinesterase Potential of Seeds from Native Korean Persimmon Genotypes


Abstract
:1. Introduction
2. Results and Discussion
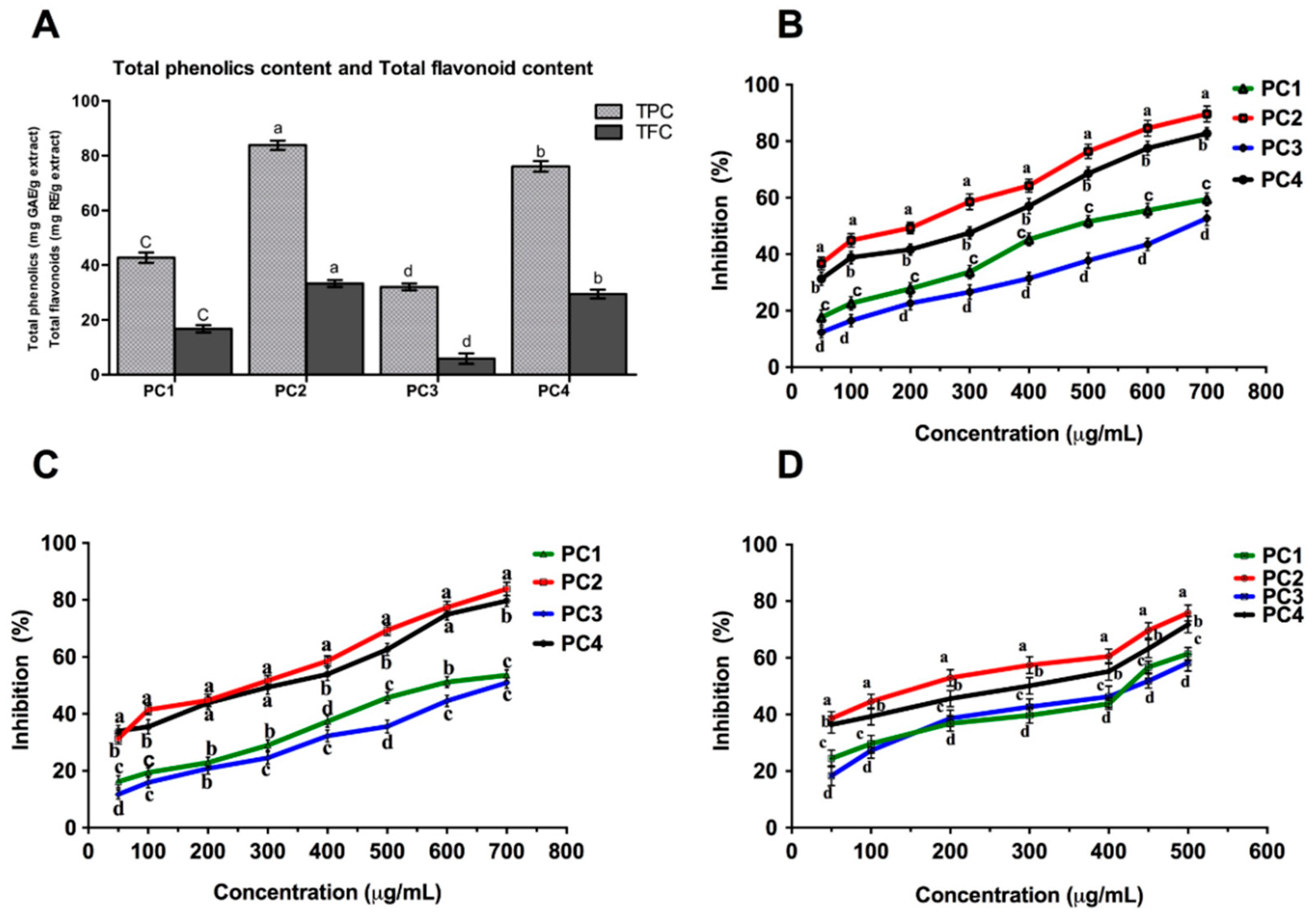
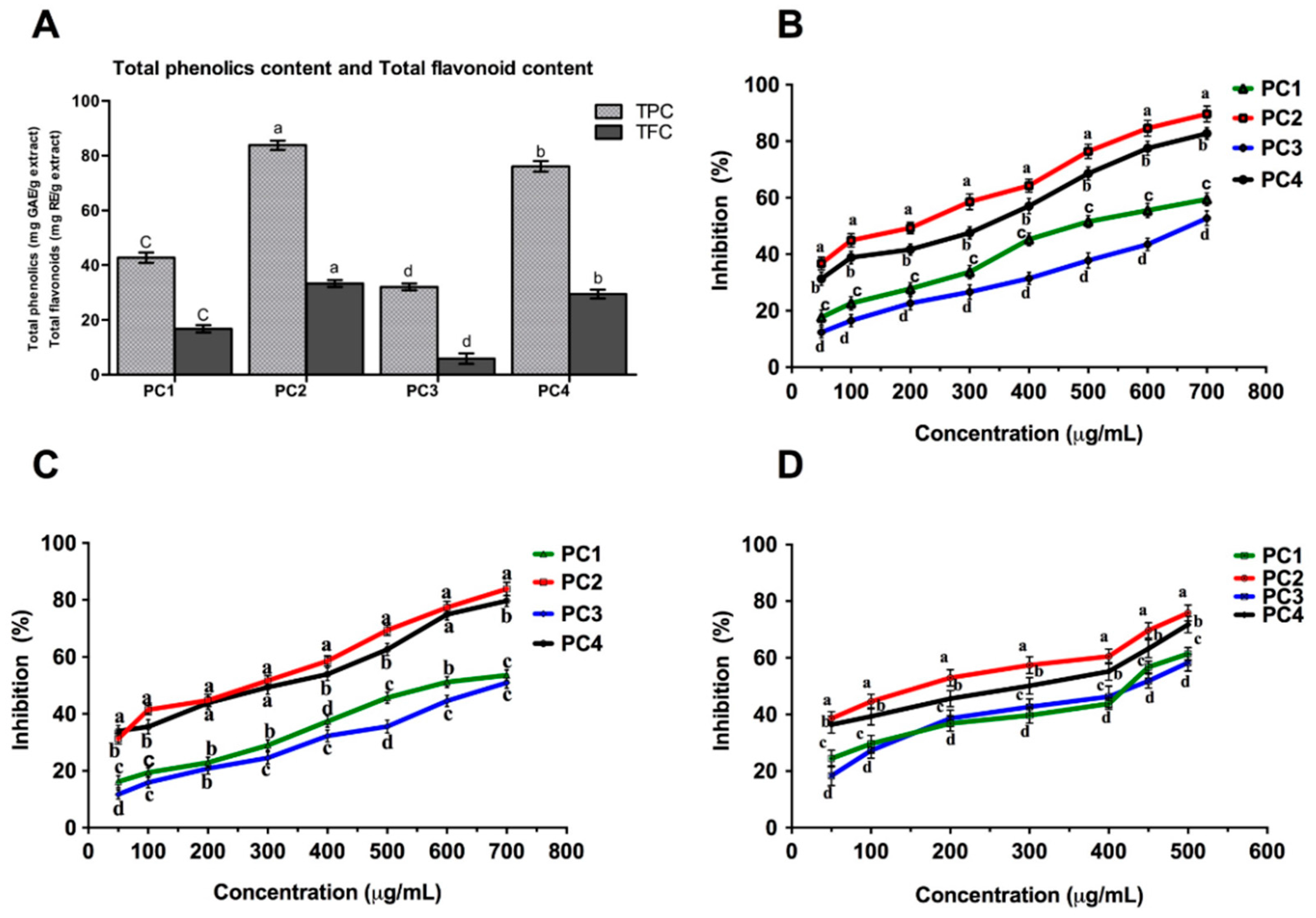
2.1. Total Phenolic Content (TPC)
2.2. Total Flavonoid Content (TFC)
2.3. Oxidant Scavenging Potentials of Persimmon Seeds
Radical Scavenging Activity (DPPH, ABTS and SO Assays)
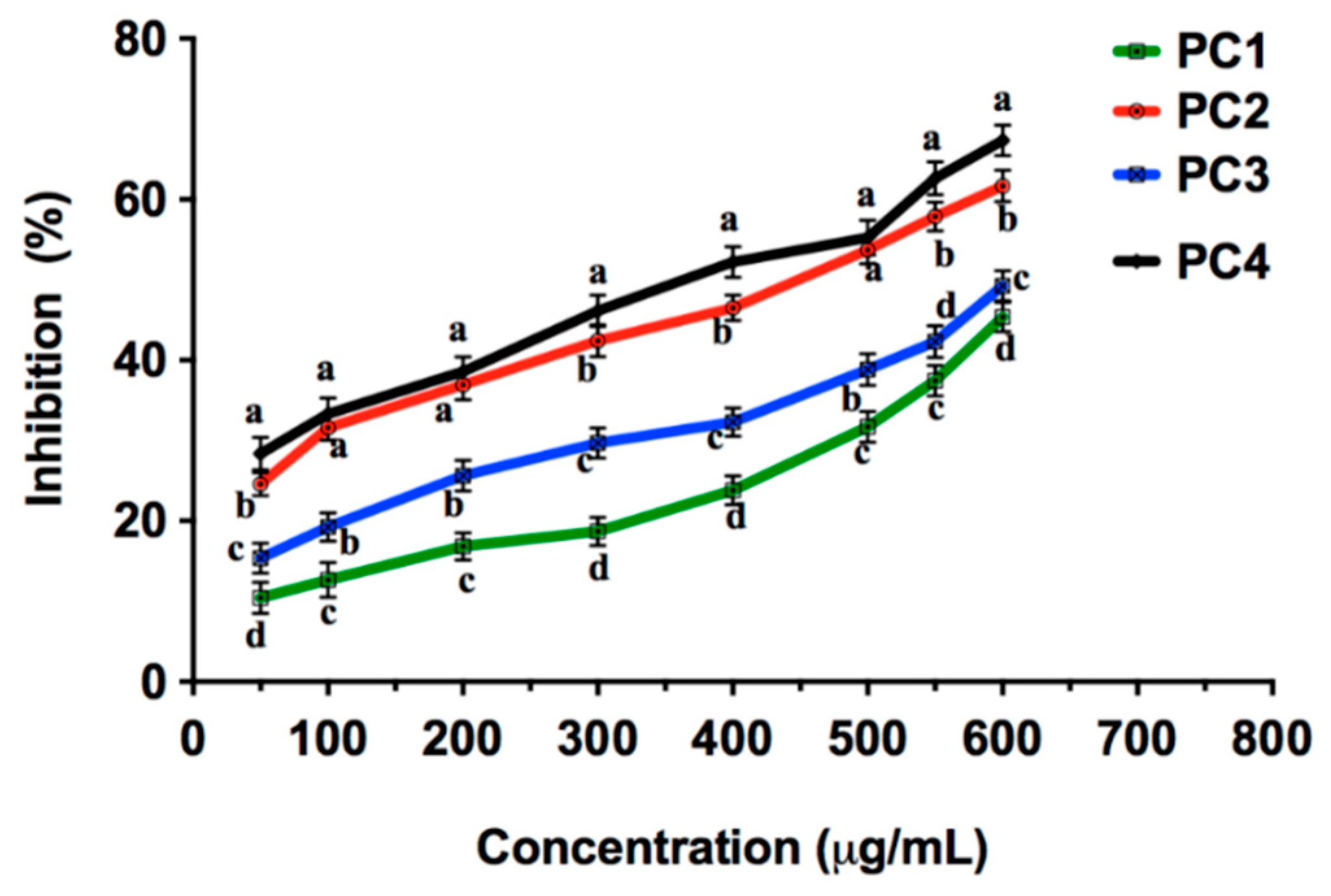
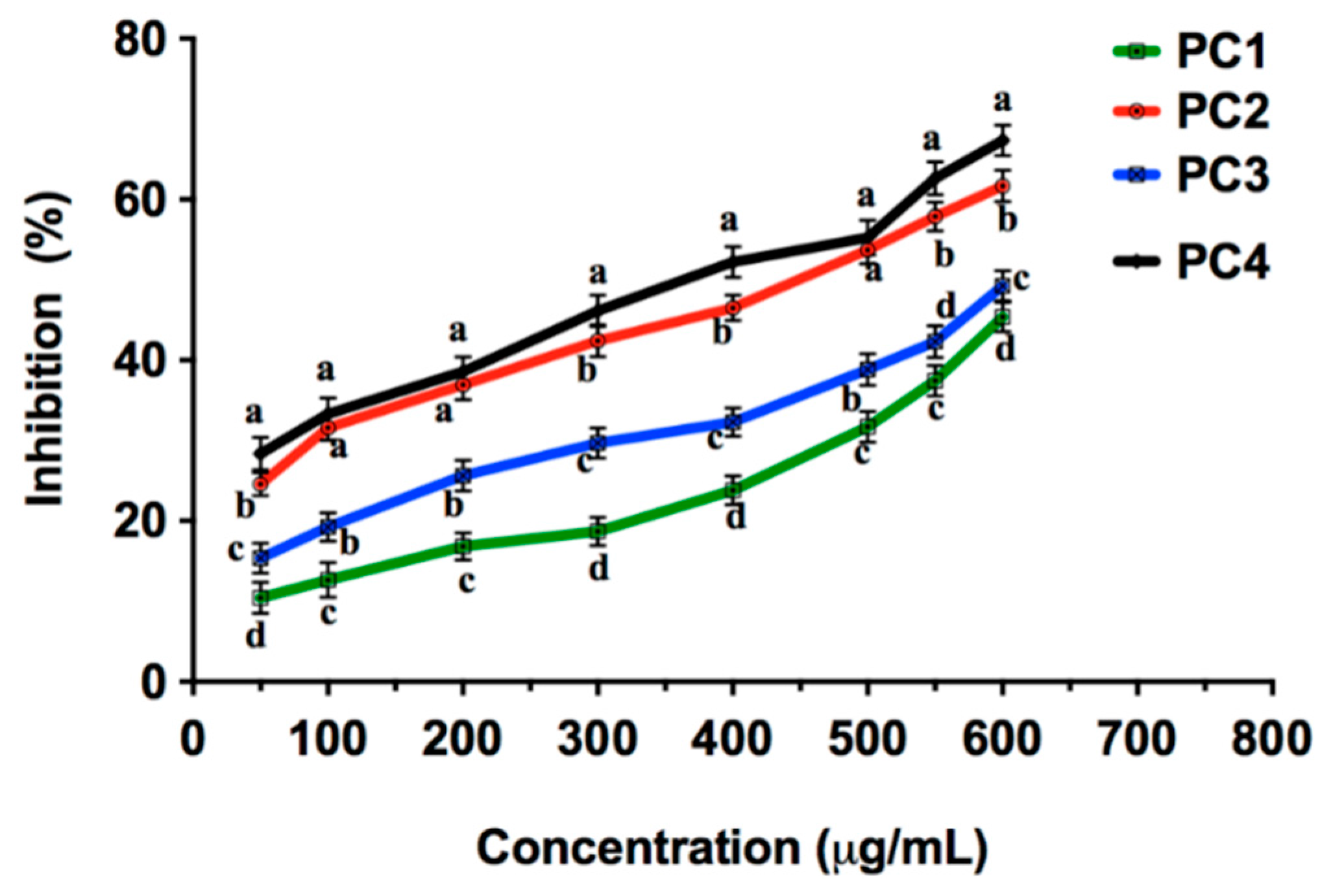
2.4. Acetylcholinesterase (AChE) Inhibition by Persimmon Seeds
2.5. Free Amino Acid in Persimmon Seeds
2.6. Fatty Acids in the Seeds of Persimmon
2.7. Organic Acids Content in Different Persimmon Cultivar Seeds
2.8. Relationship between Total Phenolic/Flavonoid Content, Antioxidant Activities, and Acetyl-cholinesterase Inhibition
3. Materials and Methods
3.1. Chemicals
3.2. Plant Materials
3.3. Preparation of Extracts of Persimmon Seeds
3.4. Determination of Total Phenolic Content
3.5. Determination of Total Flavonoid Content
3.6. DPPH Radical-Scavenging Activity
3.7. ABTS Radical-Scavenging Activity (RSA)
3.8. Superoxide Radical-Scavenging Activity
3.9. Micro-Plate Assay for Inhibition of Acetylcholinesterase
3.10. Free Amino Acid Composition
3.11. Fatty Acids Contents Analysis
3.12. Organic Acid Composition
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Datta, K.; Sinha, S.; Chattopadhyay, P. Reactive oxygen species in health and disease. Nat. Med. J. India 2000, 13, 304–310. [Google Scholar]
- Gülçin, İ.; Oktay, M.; Küfrevioğlu, Ö.İ.; Aslan, A. Determination of antioxidant activity of lichen Cetraria islandica (L) Ach. Ethnopharmacology 2002, 79, 325–329. [Google Scholar] [PubMed]
- Valko, M.; Rhodes, C.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [PubMed]
- Inoue, M.; Sato, E.F.; Nishikawa, M.; Park, A.M.; Kira, Y.; Imada, I.; Utsumi, K. Mitochondrial generation of reactive oxygen species and its role in aerobic life. Curr. Med. Chem. 2003, 10, 2495–2505. [Google Scholar] [PubMed]
- Bagchi, D.; Bagchi, M.; Stohs, S.J.; Das, D.K.; Ray, S.D.; Kuszynski, C.A.; Joshi, S.S.; Pruess, H.G. Free radicals and grape seed proanthocyanidin extract: Importance in human health and disease prevention. Toxicology 2000, 148, 187–197. [Google Scholar] [CrossRef]
- Klimczak, I.; Małecka, M.; Szlachta, M.; Gliszczyńska-Świgło, A. Effect of storage on the content of polyphenols, vitamin C and the antioxidant activity of orange juices. Food Compos. Anal. 2007, 20, 313–322. [Google Scholar]
- Jang, I.C.; Jo, E.K.; Bae, S.M.; Bae, M.S.; Lee, H.J.; Park, E.; Yuk, H.G.; Ahn, G.H.; Lee, S.C. Antioxidant activity and fatty acid composition of four different persimmon seeds. Food Sci. Technol. Res. 2010, 16, 577–584. [Google Scholar] [CrossRef]
- Moure, A.; Cruz, J.M.; Franco, D.; Domı́nguez, J.M.; Sineiro, J.; Domı́nguez, H.; Núñez, M.J.; Parajó, J.C. Natural antioxidants from residual sources. Food Chem. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Scherer, R.; Godoy, H.T. Antioxidant activity index (AAI) by the 2,2-diphenyl-1-picrylhydrazyl method. Food Chem. 2009, 112, 654–658. [Google Scholar]
- Luo, D.; Fang, B. Structural identification of ginseng polysaccharides and testing of their antioxidant activities. Carbohydr. Polym. 2008, 72, 376–381. [Google Scholar]
- Ajila, C.; Naidu, K.; Bhat, S.; Rao, U.P. Bioactive compounds and antioxidant potential of mango peel extract. Food Chem. 2007, 105, 982–988. [Google Scholar]
- Li, J.W.; Ding, S.D.; Ding, X.L. Comparison of antioxidant capacities of extracts from five cultivars of Chinese jujube. Process Biochem. 2005, 40, 3607–3613. [Google Scholar]
- Sochor, J.; Skutkova, H.; Babula, P.; Zitka, O.; Cernei, N.; Rop, O.; Krska, B.; Adam, V.; Provazník, I.; Kizek, R. Mathematical evaluation of the amino acid and polyphenol content and antioxidant activities of fruits from different apricot cultivars. Molecules 2011, 16, 7428–7457. [Google Scholar] [PubMed]
- Calder, P.C. Dietary fatty acids and the immune system. Lipids 1999, 34, S137–S140. [Google Scholar] [PubMed]
- Alana de Vasconcelos, A. D.; Oliveira, P.S.; Ritter, M.; Freitag, R.A.; Romano, R.L.; Quina, F.H.; Pizzuti, L.; Pereira, C.M.P.; Stefanello, F.M.; Barscha, A.G. Antioxidant capacity and environmentally friendly synthesis of dihydropyrimidin-(2H)-ones promoted by naturally occurring organic acids. Biochem. Mol. Toxicol. 2012, 26, 155–161. [Google Scholar]
- Yahia, E.M.; Gutierrez-Orozco, F.; Leon, C.A.D. Phytochemical and antioxidant characterization of the fruit of black sapote (Diospyros digyna Jacq.). Food Res. Int. 2011, 44, 2210–2216. [Google Scholar]
- Suzuki, T.; Someya, S.; Hu, F.; Tanokura, M. Comparative study of catechin compositions in five Japanese persimmons (Diospyros kaki). Food Chem. 2005, 93, 149–152. [Google Scholar]
- George, A.P.; Redpath, S. Health and medicinal benefits of persimmon fruit: A review. Adv. Hortic. Sci. 2008, 22, 244–249. [Google Scholar]
- Bei, W.; Zang, L.; Guo, J.; Peng, W.; Xu, A.; Good, D.A.; Hu, Y.; Wu, W.; Hu, D.; Zhu, X.; Wei, M.; Li, C. Neuroprotective effects of a standardized flavonoid extract from Diospyros kaki leaves. Ethnopharmacology 2009, 126, 134–142. [Google Scholar] [PubMed]
- Akter, M.S.; Ahmed, M.; Eun, J.B. Solvent effects on antioxidant properties of persimmon (Diospyros kaki L. cv. Daebong) seeds. Int. J. Food Sci. Technol. 2010, 45, 2258–2264. [Google Scholar]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar]
- Celep, E.; Aydın, A.; Yesilada, E. A comparative study on the in vitro antioxidant potentials of three edible fruits: Cornelian cherry, Japanese persimmon and cherry laurel. Food Chem. Toxicol. 2012, 50, 3329–3335. [Google Scholar] [PubMed]
- Ercisli, S.; Akbulut, M.; Ozdemir, O.; Sengul, M.; Orhan, E. Phenolic and antioxidant diversity among persimmon (Diospyrus kaki L.) genotypes in Turkey. Int. J. Food Sci. Nutr. 2008, 59, 477–482. [Google Scholar] [PubMed]
- Pu, F.; Ren, X.L.; Zhang, X.P. Phenolic compounds and antioxidant activity in fruits of six Diospyros kaki genotypes. Eur. Food Res. Technol. 2013, 237, 923–932. [Google Scholar]
- Li, P.M.; Du, G.R.; Ma, F.W. Phenolics concentration and antioxidant capacity of different fruit tissues of astringent versus non-astringent persimmons. Sci. Hortic. 2011, 129, 710–714. [Google Scholar]
- Simó, C.; Ibáez, C.; Valdés, A.; Cifuentes, A.; García-Cañas, V. Metabolomics of genetically modified crops. Int.J. Mol. Sci. 2014, 15, 18941–18966. [Google Scholar] [PubMed]
- Shon, M.Y.; Kim, T.H.; Sung, N.J. Antioxidants and free radical scavenging activity of Phellinus baumii (Phellinus of Hymenochaetaceae) extracts. Food Chem. 2003, 82, 593–597. [Google Scholar]
- Ahn, H.S.; Jeon, T.I.; Lee, J.Y.; Hwang, S.G.; Lim, Y.; Park, D.K. Antioxidative activity of persimmon and grape seed extract: In vitro and in vivo. Nutr. Res. 2002, 22, 1265–1273. [Google Scholar]
- Almeida, M.M.B.; de Sousa, P.H.M.; Arriaga, Â.M.C.; do Prado, G.M.; de Carvalho Magalhães, C.E.; Maia, G.A.; de Lemos, T.L.G. Bioactive compounds and antioxidant activity of fresh exotic fruits from northeastern Brazil. Food Res. Int. 2011, 44, 2155–2159. [Google Scholar]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, J.; Lu, X.; Zhang, L.; Zhang, Y. Evaluation to the antioxidant activity of total flavonoids extract from persimmon (Diospyros kaki L.) leaves and Food. Chem. Toxicol. 2012, 50, 3329–3335. [Google Scholar]
- Rufino, M.D.S.M.; Alves, R.E.; de Brito, E.S.; Pérez-Jiménez, J.; Saura-Calixto, F.; Mancini-Filho, J. Bioactive compounds and antioxidant capacities of 18 non-traditional tropical fruits from Brazil. Food Chem. 2010, 121, 996–1002. [Google Scholar]
- Moo-Huchin, V.M.; Estrada-Mota, I.; Estrada-León, R.; Cuevas-Glory, L.; Ortiz-Vázquez, E.; Vargas, M.D.L.V.Y.; Betancur-Ancona, D.; Sauri-Duch, E. Determination of some physicochemical characteristics, bioactive compounds and antioxidant activity of tropical fruits from Yucatan, Mexico. Food Chem. 2014, 152, 508–515. [Google Scholar] [PubMed]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [PubMed]
- Mahattanatawee, K.; Manthey, J.A.; Luzio, G.; Talcott, S.T.; Goonder, K.; Baldwin, E.A. Total antioxidant activity and fiber content of select florida-grown tropical fruits. J. Agric. Food Chem. 2006, 54, 7355–7363. [Google Scholar] [PubMed]
- Scalzo, J.; Politi, A.; Pellegrini, N.; Mezzetti, B.; Battino, M. Plant genotype affects total antioxidant capacity and phenolic contents in fruit. Nutrition 2005, 21, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Pervin, M.; Hasnat, M.A.; Lee, Y.M.; Kim, H.D.; Jo, J.E.; Lim, B.O. Antioxidant activity and acetylcholinesterase inhibition of grape skin anthocyanin (GSA). Molecules 2014, 19, 9403–9418. [Google Scholar] [CrossRef] [PubMed]
- Tiana, Y.; Zoua, B.; Yanga, L.; Xua, S.; Yanga, J.; Yaob, P.; Lia, C. High molecular weight persimmon tannin ameliorates cognition deficits and attenuates oxidative damage in senescent mice induced by d-galactose. Food Chem. Toxicol. 2011, 49, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.F.; Falé, P.L.V.; Araújo, M.E.M.; Serralheiro, M.L.M. Acetylcholinesterase inhibition and antioxidant activity of the water extracts of several Hypericum species. Food Chem. 2010, 120, 1076–1082. [Google Scholar] [CrossRef]
- Williams, P.; Sorribas, A.; Howes, M.J.R. Natural products as a source of Alzheimer’s drug leads. Nat. Prod. Rep. 2011, 28, 48–77. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, R.; Falleh, H.; Megdiche, W.; Trabelsi, N.; Mhamdi, B.; Chaieb, K.; Bakrouf, A.; Magné, C.; Abdelly, C. Antioxidant and antimicrobial activities of the edible medicinal halophyte Tamarix gallica L. and related polyphenolic constituents. Food Chem. Toxicol. 2009, 47, 2083–2091. [Google Scholar] [CrossRef] [PubMed]
- Ramassamy, C. Emerging role of polyphenolic compounds in the treatment of neurodegenerative diseases: A review of their intracellular targets. Eur.J. Pharmacol. 2006, 545, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Lim, J.H.; Kim, Y.C.; Kim, M.Y.; Lee, B.W.; Chung, S.K. Chemical composition and quality of persimmon peels according to cultivars. Korean Soc. Appl. Biol. Chem. 2005, 48, 70–76. [Google Scholar]
- Zhong, S.; Ding, T.L.; Li, Y.G.; Wu, S.G.; Lv, Z.Q. Determination and analysis of main nutritional ingredients in mulberry fruit of Morus nigra L. Chin. J. Sci. Seric. 2012, 38, 1067–1072. [Google Scholar]
- Maimaiti, Y.M.; Xia, Q.Y.; Wu, L.L.; Yin, G.; Zeng, F.J.; Yan, H.L. Study on desert mulberry from Xinjiang. Chin. J. North Seric. 2007, 28, 1–4. [Google Scholar]
- Ruth, M.R.; Field, C.J. The immune modifying effects of amino acids on gut-associated lymphoid tissue. J. Anim. Sci. Biotechnol. 2013, 4, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Holt, A.G.; Newman, S.W. Distribution of methionine and leucine enkephalin neurons within the social behavior circuitry of the male Syrian hamster brain. Brain Res. 2004, 1030, 28–48. [Google Scholar] [CrossRef] [PubMed]
- Layman, D.K.; Walker, D.A. Potential importance of leucine in treatment of obesity and the metabolic syndrome. Nutrition 2006, 136, 319S–323S. [Google Scholar]
- Gerich, J.E.; Meyer, C.; Woerle, H.J.; Stumvoll, M. Renal Gluconeogenesis Its importance in human glucose homeostasis. Diabetes Care 2001, 24, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Kirma, M.; Araújo, W.L.; Fernie, A.R.; Galili, G. The multifaceted role of aspartate-family amino acids in plant metabolism. Exp. Bot. 2012, 63, 4995–5001. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.E.; Geballe, M.T.; Stansfeld, P.J.; Johnston, A.R.; Yuan, H.; Jacob, A.L.; Snyder, J.P.; Traynelis, S.F.; Wyllie, D.J. Structural features of the glutamate binding site in recombinant NR1/NR2A N-methyl-d-aspartate receptors determined by site-directed mutagenesis and molecular modeling. Mol. Pharmacol. 2005, 67, 1470–1484. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, J.; Butterworth, R.F. The brain glutamate system in liver failure. J. Neurochem. 2006, 98, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, H.A.; Ebrahimi, S. Evaluation of the effects of charged amino acids on uncontrolled seizures. Neurol. Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Paredes, R.G.; Ågmo, A. GABA and behavior: The role of receptor subtypes. Neurosci. Biobehav. Rev. 1992, 16, 145–170. [Google Scholar] [CrossRef]
- Egydio, A.P.M.; Santa Catarina, C.; Floh, E.I.S.; dos Santos, D.Y.A.C. Free amino acid composition of Annona (Annonaceae) fruit species of economic interest. Ind. Crop. Prod. 2013, 45, 373–376. [Google Scholar] [CrossRef]
- Aktumsek, A.; Zengin, G.; Guler, G.O.; Cakmak, Y.S.; Duran, A. Assessment of the antioxidant potential and fatty acid composition of four Centaurea L. taxa from Turkey. Food Chem. 2013, 141, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Glew, R.H.; Ayaz, F.A.; Millson, M.; Huang, H.; Chuang, L.; Sanz, C.; Golding, J.B. Changes in sugars, acids and fatty acids in naturally parthenocarpic date plum persimmon (Diospyros lotus L.) fruit during maturation and ripening. Eur. Food Res. Technol. 2005, 221, 113–118. [Google Scholar] [CrossRef]
- Carrillo Pérez, C.; Cavia Camarero, M.D.M.; Alonso de la Torre, S. Role of oleic acid in immune system; mechanism of action; a review. Nutr. Hosp. 2012, 27, 978–990. [Google Scholar]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.; Ross, R.; Stanton, C. Health implications of high dietary ω-6 polyunsaturated fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef] [PubMed]
- Veberic, R.; Jurhar, J.; Mikulic-Petkovsek, M.; Stampar, F.; Schmitzer, V. Comparative study of primary and secondary metabolites in 11 cultivars of persimmon fruit (Diospyros kaki L.). Food Chem. 2010, 119, 477–483. [Google Scholar] [CrossRef]
- Pande, G.; Akoh, C.C. Organic acids, antioxidant capacity, phenolic content and lipid characterisation of Georgia-grown underutilized fruit crops. Food Chem. 2010, 120, 1067–1075. [Google Scholar] [CrossRef]
- Hernández, Y.; Lobo, M.G.; González, M. Determination of vitamin C in tropical fruits: A comparative evaluation of methods. Food Chem. 2006, 96, 654–664. [Google Scholar] [CrossRef]
- Igual, M.; García-Martínez, E.; Martín-Esparza, M.; Martínez-Navarrete, N. Effect of processing on the drying kinetics and functional value of dried apricot. Food Res. Int. 2012, 47, 284–290. [Google Scholar] [CrossRef]
- Abdel-Hameed, E.-S.S. Total phenolic contents and free radical scavenging activity of certain Egyptian Ficus species leaf samples. Food Chem. 2009, 114, 1271–1277. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. LWT-Food Sci. Technol. 2007, 40, 344–352. [Google Scholar] [CrossRef]
- Hasnat, M.A.; Pervin, M.; Lim, B.O. Acetylcholinesterase inhibition and in vitro and in vivo antioxidant activities of Ganoderma lucidum grown on germinated brown rice. Molecules 2013, 18, 6663–6678. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Naczk, M.; Shahidi, F. Phenolics in cereals, fruits and vegetables: Occurrence, extraction and analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Aydin, A.; Orhan, H.; Sayal, A.; Özata, M.; Sahin, G.; Isimer, A. Oxidative stress and nitric oxide related parameters in type II diabetes mellitus: Effects of glycemic control. Clin. Biochem. 2001, 34, 65–70. [Google Scholar] [CrossRef]
- Moyo, M.; Ndhlala, A.R.; Finnie, J.F.; Staden, J.V. Phenolic composition, antioxidant and acetylcholinesterase inhibitory activities of Sclerocarya birrea and Harpephyllum caffrum (Anacardiaceae) extracts. Food Chem. 2010, 123, 69–76. [Google Scholar] [CrossRef]
- Waqas, M.; Shahzad, R.; Khan, A.L.; Asaf, S.; Kim, Y.H.; Kang, S.M.; Bilal, S.; Hamayun, M.; Lee, I.J. Salvaging effect of triacontanol on plant growth, thermotolerance, macro-nutrient content, amino acid concentration and modulation of defense hormonal levels under heat stress. Plant Physiol. Biochem. 2016, 99, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Khan, A.L.; Kim, D.H.; Lee, S.Y.; Kim, K.M.; Waqas, M.; Jung, H.Y.; Shin, J.H.; Kim, J.G.; Lee, I.J. Silicon mitigates heavy metal stress by regulating p-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Khan, A.L.; Lee, I.J. Bioactive chemical constituents produced by endophytes and effects on rice plant growth. J. Plant Interact. 2014, 9, 478–487. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.

{kind=link}
{kind=link}
| Samples | DPPH Radical Scavenging | ABTS Radical Scavenging | AChE Inhibition | SO Radical Scavenging |
|---|---|---|---|---|
| PC1 | 609.03 ± 4.11 | 517.74 ± 3.37 | 779.16 ± 4.19 | 396.34 ± 3.97 |
| PC2 | 262.65 ± 2.04 | 195.83 ± 1.95 | 427.68 ± 3.78 | 190.27 ± 2.96 |
| PC3 | 706.37 ± 4.06 | 692.95 ± 4.17 | 659.96 ± 4.06 | 408.75 ± 3.72 |
| PC4 | 297.55 ± 2.42 | 285.66 ± 2.73 | 368.29 ± 2.68 | 260.51 ± 4.1 |
| Ascorbic acid | 112.23 ± 2.51 | 101.41 ± 1.96 | ||
| Galantamine | 0.78 ± 0.2 | |||
| Rutin | 185.63 ± 3.62 |
| Assays | Concentrations of Seed Extracts | Types of Persimmon Seeds | Interaction of Concentration x Types | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MS | P | % of Total Variance | MS | P | % of Total Variance | MS | P | % of Total Variance | |
| DPPH inhibition | 3040 | 0.0001 | 59.16% | 4657 | 0.0001 | 38.84% | 21.76 | 0.0001 | 1.27% |
| ABTS inhibition | 3419 | 0.0001 | 58.42% | 5346 | 0.0001 | 39.15% | 30.35 | 0.0001 | 1.56% |
| SO inhibition | 2095 | 0.0001 | 73.00% | 1335 | 0.0001 | 23.26% | 10.81 | 0.1971 | 1.13% |
| AChE inhibition | 1919 | 0.0001 | 59.74% | 2865 | 0.0001 | 38.22% | 11.15 | 0.0002 | 1.04% |
| Amino Acids | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Threonine | 129.97 ± 2.6 d | 141.18 ± 1.7 c | 167.03 ± 3.6 b | 289.078 ± 2.64 a |
| Valine | 140.51 ± 3.4 d | 186.62 ± 3 c | 228.35 ± 3.21 b | 269.18 ± 3.3 a |
| Methionine | 36.32 ± 4.3 d | 44.6 ± 2.6 c | 93.45 ± 2 b | 127.63 ± 1.2 a |
| Isoleucine | 98.85 ± 1.1 d | 134.41 ± 2.7 c | 142.04 ± 2.5 b | 251.11 ± 2 a |
| Leucine | 94.48 ± 2.16 d | 123.59 ± 2 c | 218.31 ± 3.5 b | 293.87 ± 2 d |
| Phenylalanine | 66.04 ± 4.5 d | 74.24 ± 1.1 c | 119.13 ± 0.9 b | 168.9 ± 3 a |
| Lysine | 52.28 ± 1.2 d | 60.67 ± 4.6 c | 102.46 ± 2.2 b | 157.9 ± 4 a |
| Histidine | 18.49 ± 1.3 a | ND c | 14.59 ± 3.8 b | ND c |
| Aspartic acid | 292.98 ± 3.6 b | 275.4 ± 3.01 c | 253.26 ± 1.7 d | 662.81 ± 1.1 a |
| Alanine | 458.46 ± 1.3 b | 522.44 ± 5.1 a | 436.25 ± 1.6 c | 304.55 ± 2.2 d |
| Cystine | 24.14 ± 1.09 d | 64.53 ± 3.1 b | 73.93 ± 4.3 a | 42.17 ± 2.01 c |
| Glutamc acid | 932.74 ± 1.2 a | 444.02 ± 3 b | 264.24 ± 4.4 d | 369.56 ± 2.0 c |
| Glycine | 51.31 ± 3.2 c | 139.61 ± 1.7 a | 126.54 ± 2.2 b | 140.77 ± 4.03 a |
| Sarcosine | 101.44 ± 4.2 a | 55.96 ± 3.0 b | 101.17 ± 2.1 a | 84.94 ± 2.1 c |
| Tyrosine | 94.39 ± 2.9 d | 118.34 ± 1.05 c | 148.29 ± 2.01 b | 194.21 ± 1.6 a |
| Serine | 205.95 ± 3.01 c | 231.66 ± 2.03 b | 207.79 ± 2.2 c | 416.29 ± 3.1 a |
| Arginine | ND a | ND a | ND a | ND a |
| Citrulline | 1777.15 ± 19.1 d | 2205.1 ± 15.6 c | 2914.29 ± 14.7 a | 2454.09 ± 12.4 b |
| β-Alanine | 42.89 ± 2.05 c | 55.34 ± 2.2 b | 76.81 ± 3.4 a | 53.47 ± 3.1 b |
| γ-Amino-n-butyric acid | 174.81 ± 7.77 c | 620.44 ± 11.1 a | 599.41 ± 12.87 ab | 577.91 ± 18.01 b |
| Ethanolamine | 44.65 ± 3.3 b | 65.05 ± 4.8 a | 63.81 ± 2.0 a | 45.4 ± 4.1 b |
| Ammonia | 26.64 ± 1.2 bc | 28.7 ± 2.2 b | 24.52 ± 1.04 c | 33.01 ± 2.9 a |
| 1-Methylhistidine | 174.88 ± 3.1 d | 254.58 ± 1.4 c | 322.48 ± 3.2 b | 399.01 ± 3.02 a |
| Total A. A | 2798.35 | 2617.31 | 2696.91 | 3773.05 |
| Total essential A. A | 636.98 | 765.34 | 1085.4 | 1557.69 |
| Total Metabolites | 2241.0005 | 3229.21 | 4001.33 | 3562.89 |
| Cultivars | Palmitic | Stearic | Oleic | Linoleic | Linolenic | Arachidic | Total µg/g |
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | C18:1 | C18:2 | C18:3 | C20:0 | ||
| PC1 | 17.67 ± 1.17 b | 3.26 ± 0.34 a | 37.51 ± 0.91 b | 38.05 ± 1.02 b | 3.49 ± 1.01 b | ND | 1180.5 d |
| PC2 | 21.53 ± 1.08 b | 3.46 ± 0.81 a | 38.82 ± 1.02 b | 33.21 ± 0.72 c | 2.97 ± 0.69 b | ND | 2503.5 a |
| PC3 | 17.54 ± 2.1 b | 3.57 ± 0.78 a | 36.27 ± 1.1 bc | 39.63 ± 0.4 a | 2.97 ± 0.98 b | ND | 1701.25 c |
| PC4 | 19.95 ± 1.9 ab | 3.46 ± 1.2 a | 43.22 ± 1.2 a | 30.60 ± 0.88 d | 2.76 ± 1.1 b | ND | 1851.1 b |
| Cultivars | Oxalic Acid | Citric Acid | Malic Acid | Succinic Acid | Lactic Acid | Acetic Acid | Total |
|---|---|---|---|---|---|---|---|
| PC1 | 111 ± 1.0 d | 248 ± 1.9 d | 3510 ± 15.1 b | 46 ± 1.7 a | 23 ± 0.7 a | ND c | 3938 c |
| PC2 | 144 ± 2.0 c | 307 ± 4.2 b | 1985 ± 4.3 d | 21 ± 0.8 c | 18 ± 1.0 b | 23 ± 1.8 a | 2498 d |
| PC3 | 178 ± 2.4 b | 351 ± 4.3 a | 3263 ± 8.9 c | 21 ± 1.2 c | 11 ± 0.8 c | 15 ± 1.0 b | 3839 b |
| PC4 | 193 ± 3.1 a | 270 ± 3.4 c | 3708 ± 6.1 a | 35 ± 0.9 b | 6 ± 0.5 d | ND c | 4212 a |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilal, S.; Khan, A.L.; Waqas, M.; Shahzad, R.; Kim, I.-D.; Lee, I.-J.; Shin, D.-H. Biochemical Constituents and in Vitro Antioxidant and Anticholinesterase Potential of Seeds from Native Korean Persimmon Genotypes. Molecules 2016, 21, 893. https://doi.org/10.3390/molecules21070893
Bilal S, Khan AL, Waqas M, Shahzad R, Kim I-D, Lee I-J, Shin D-H. Biochemical Constituents and in Vitro Antioxidant and Anticholinesterase Potential of Seeds from Native Korean Persimmon Genotypes. Molecules. 2016; 21(7):893. https://doi.org/10.3390/molecules21070893
Chicago/Turabian StyleBilal, Saqib, Abdul Latif Khan, Muhammad Waqas, Raheem Shahzad, Il-Doo Kim, In-Jung Lee, and Dong-Hyun Shin. 2016. "Biochemical Constituents and in Vitro Antioxidant and Anticholinesterase Potential of Seeds from Native Korean Persimmon Genotypes" Molecules 21, no. 7: 893. https://doi.org/10.3390/molecules21070893
APA StyleBilal, S., Khan, A. L., Waqas, M., Shahzad, R., Kim, I.-D., Lee, I.-J., & Shin, D.-H. (2016). Biochemical Constituents and in Vitro Antioxidant and Anticholinesterase Potential of Seeds from Native Korean Persimmon Genotypes. Molecules, 21(7), 893. https://doi.org/10.3390/molecules21070893