
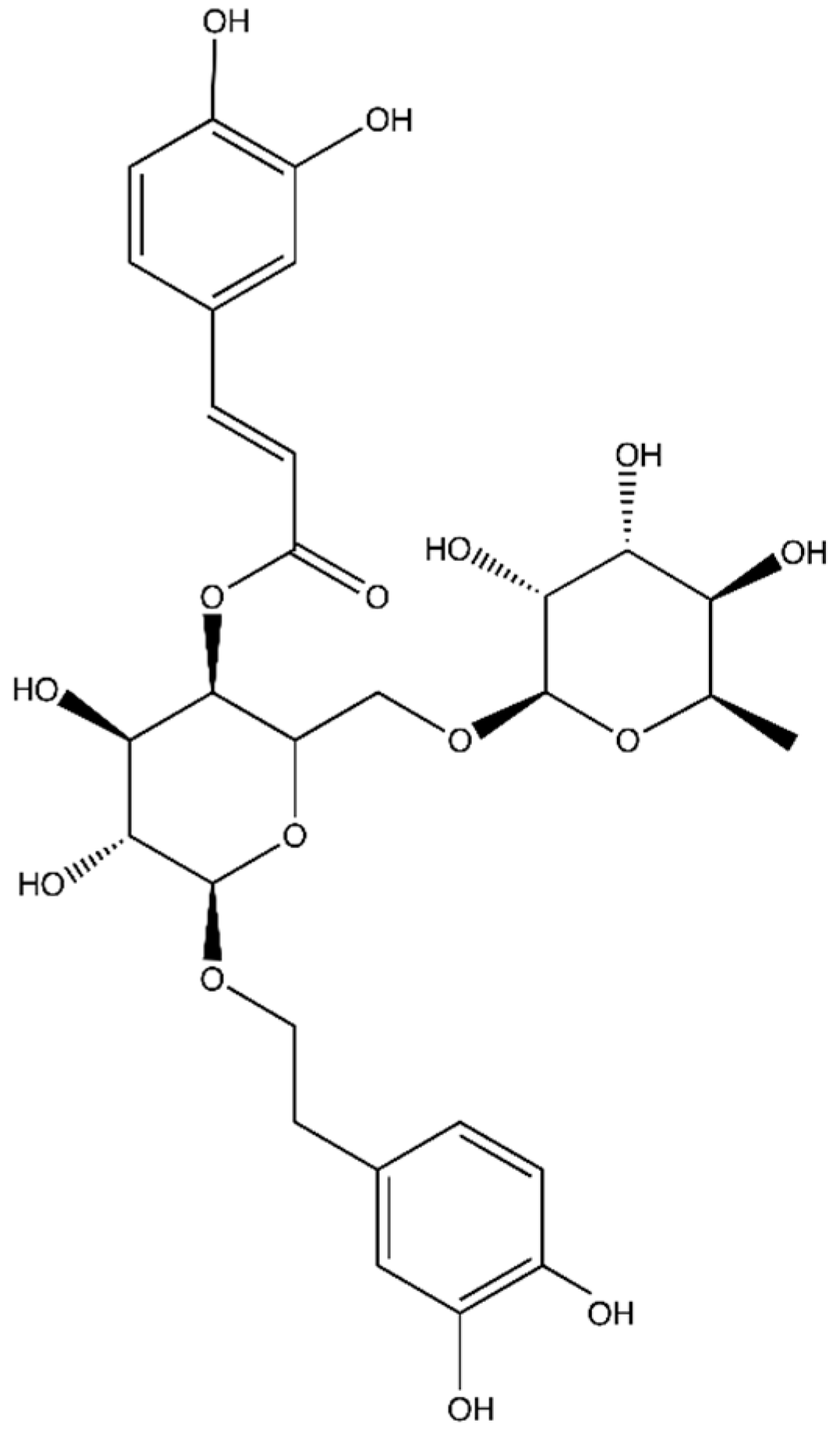
Forsythoside A Controls Influenza A Virus Infection and Improves the Prognosis by Inhibiting Virus Replication in Mice

Abstract
:1. Introduction
2. Results
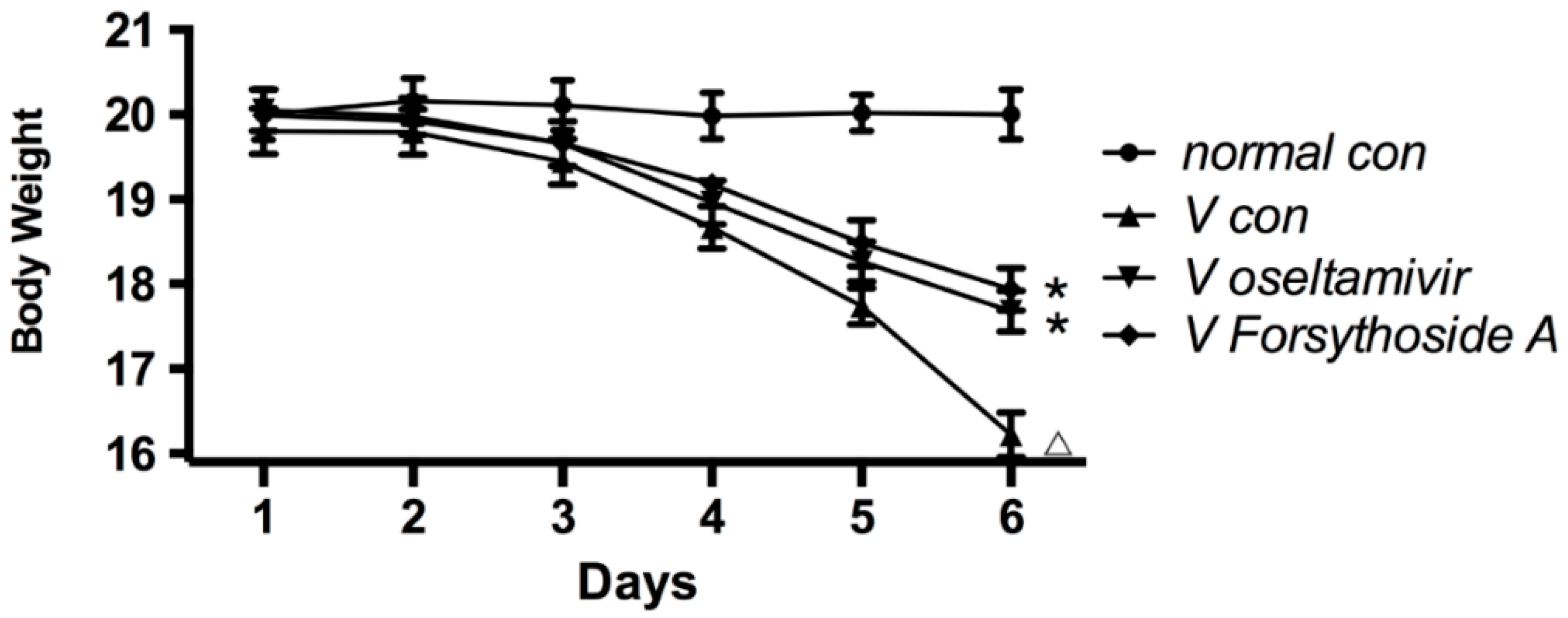
2.1. Changes in Body Weight
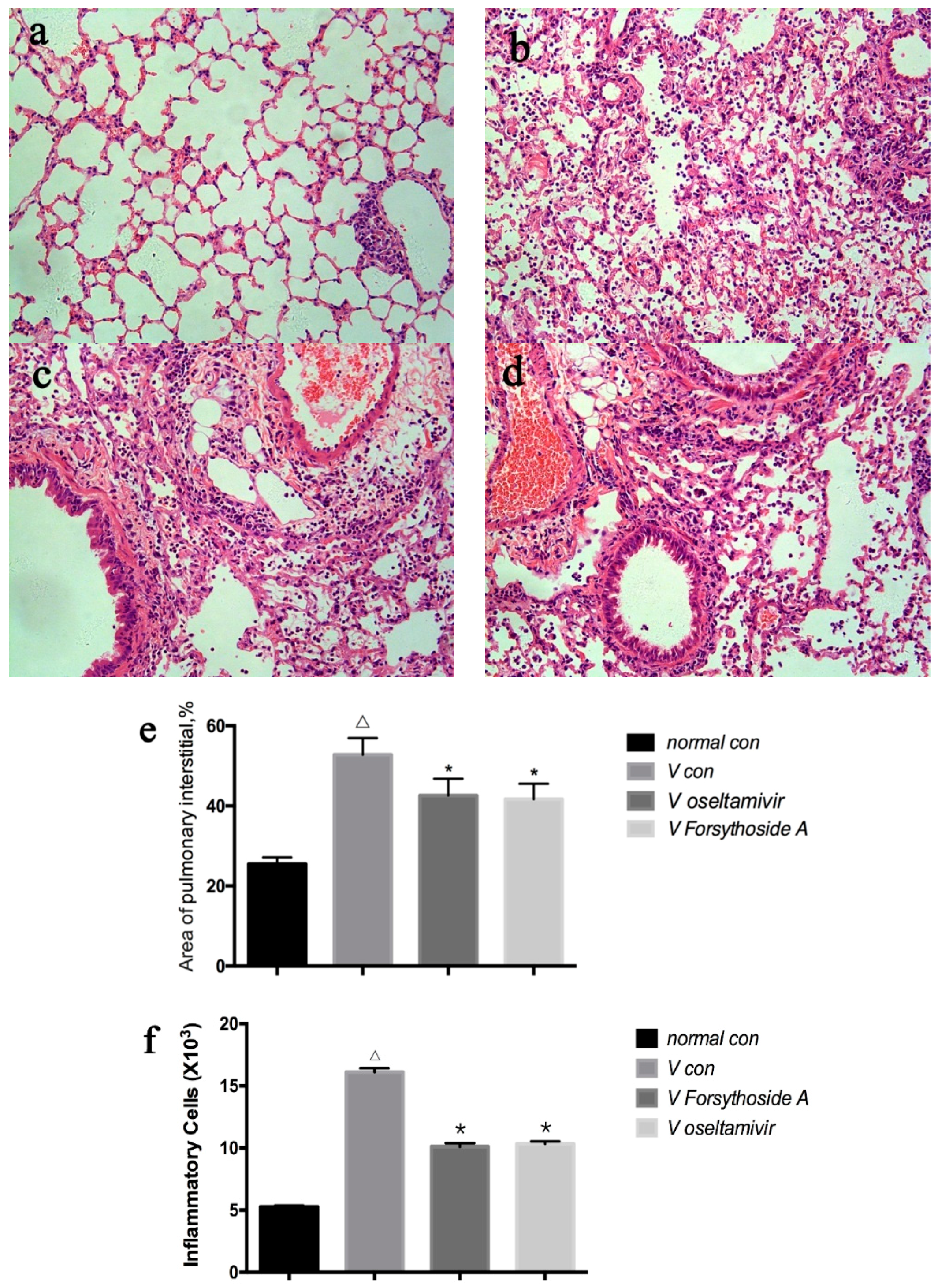
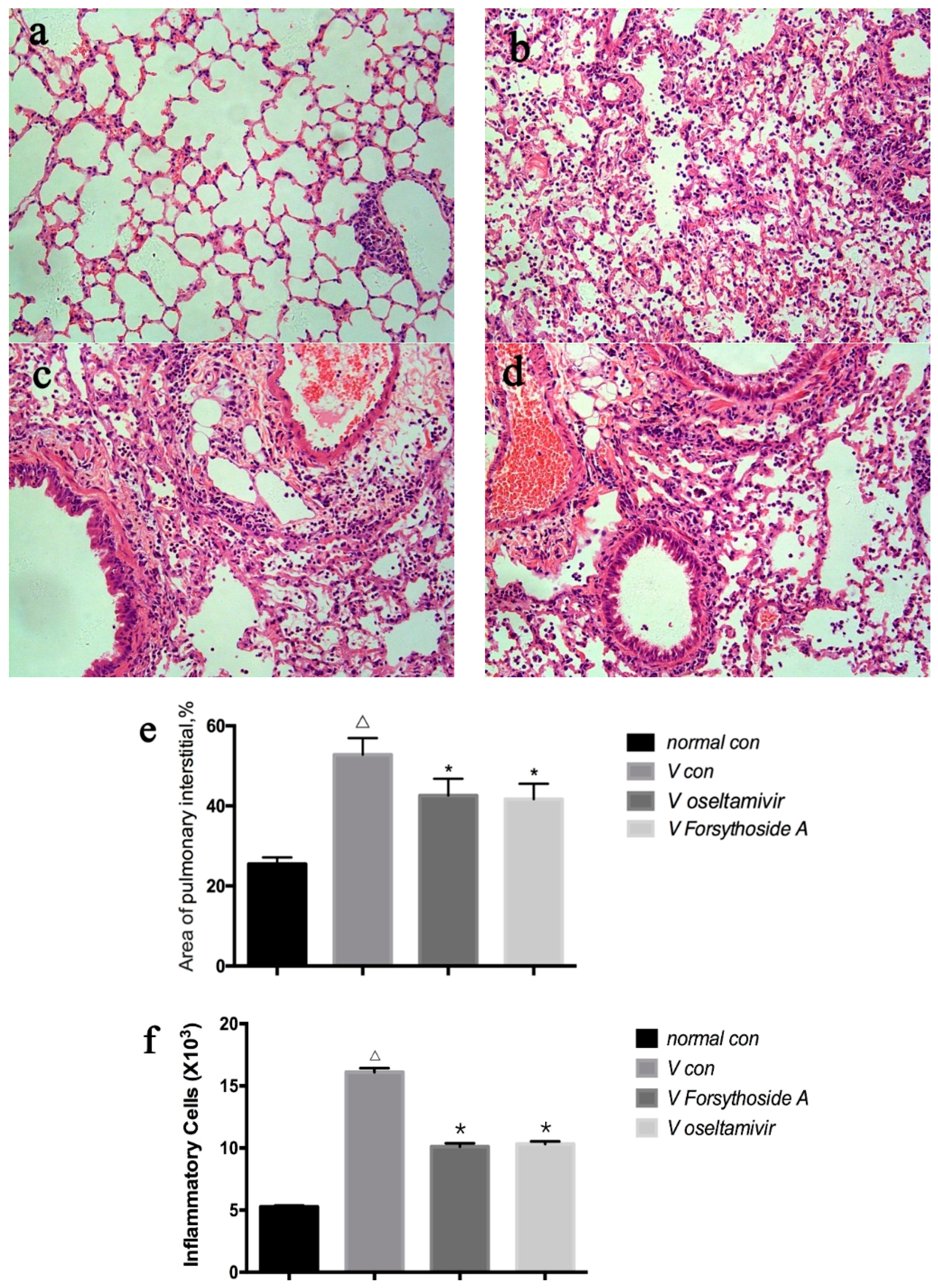
2.2. Changes in Lung Tissue
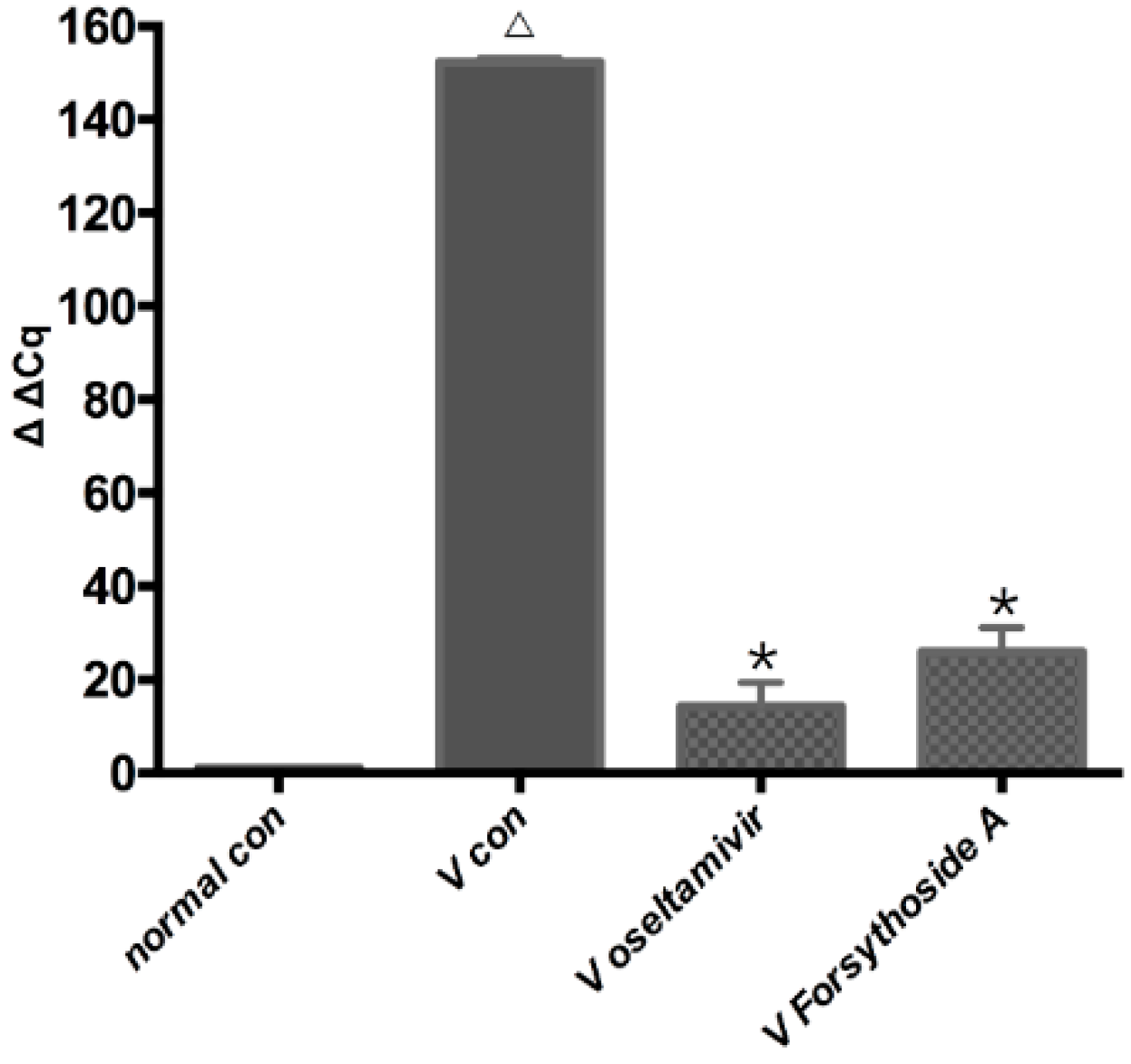
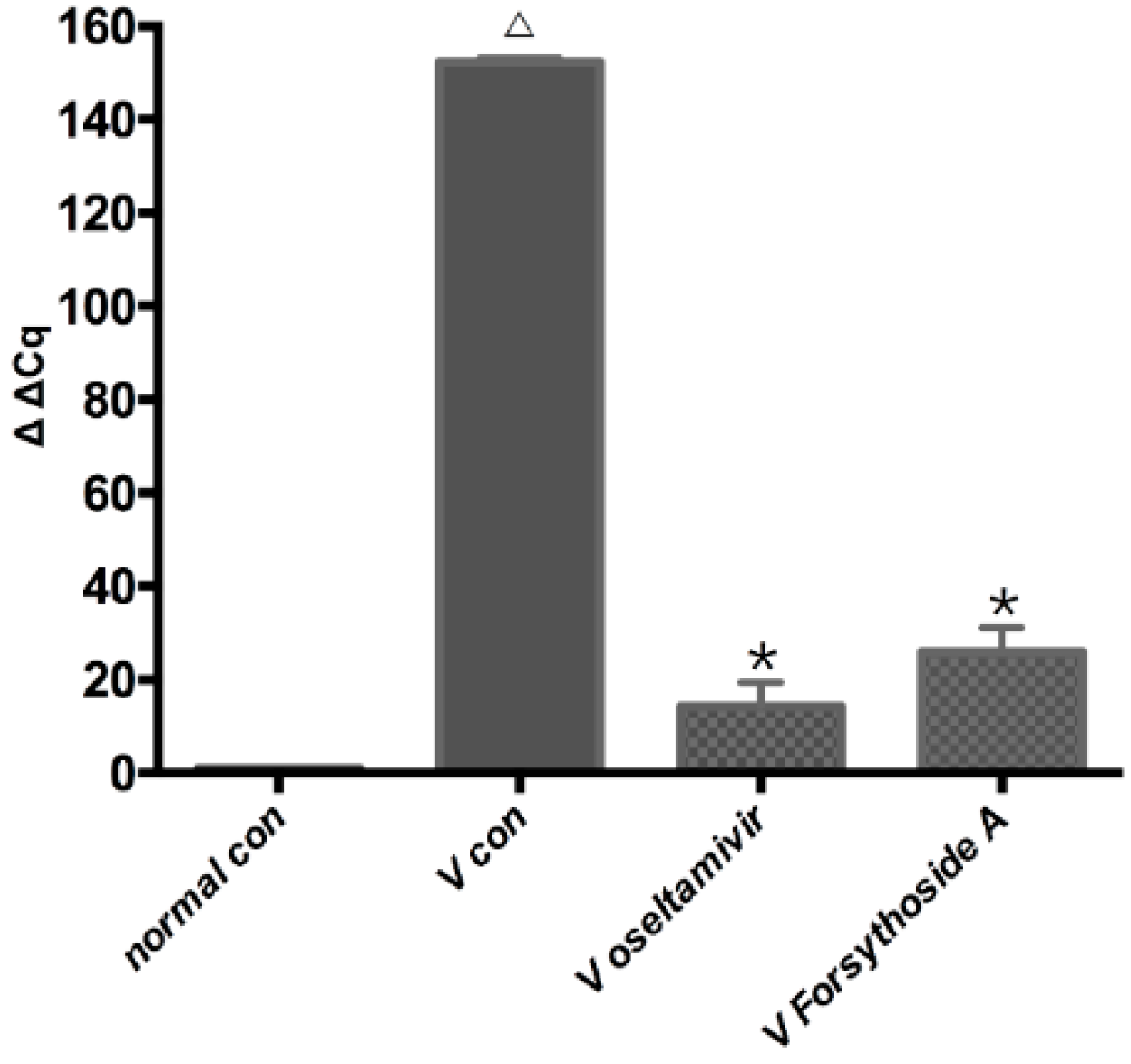
2.3. Changes in Relative Expression of the Influenza A Virus Replication in Lung
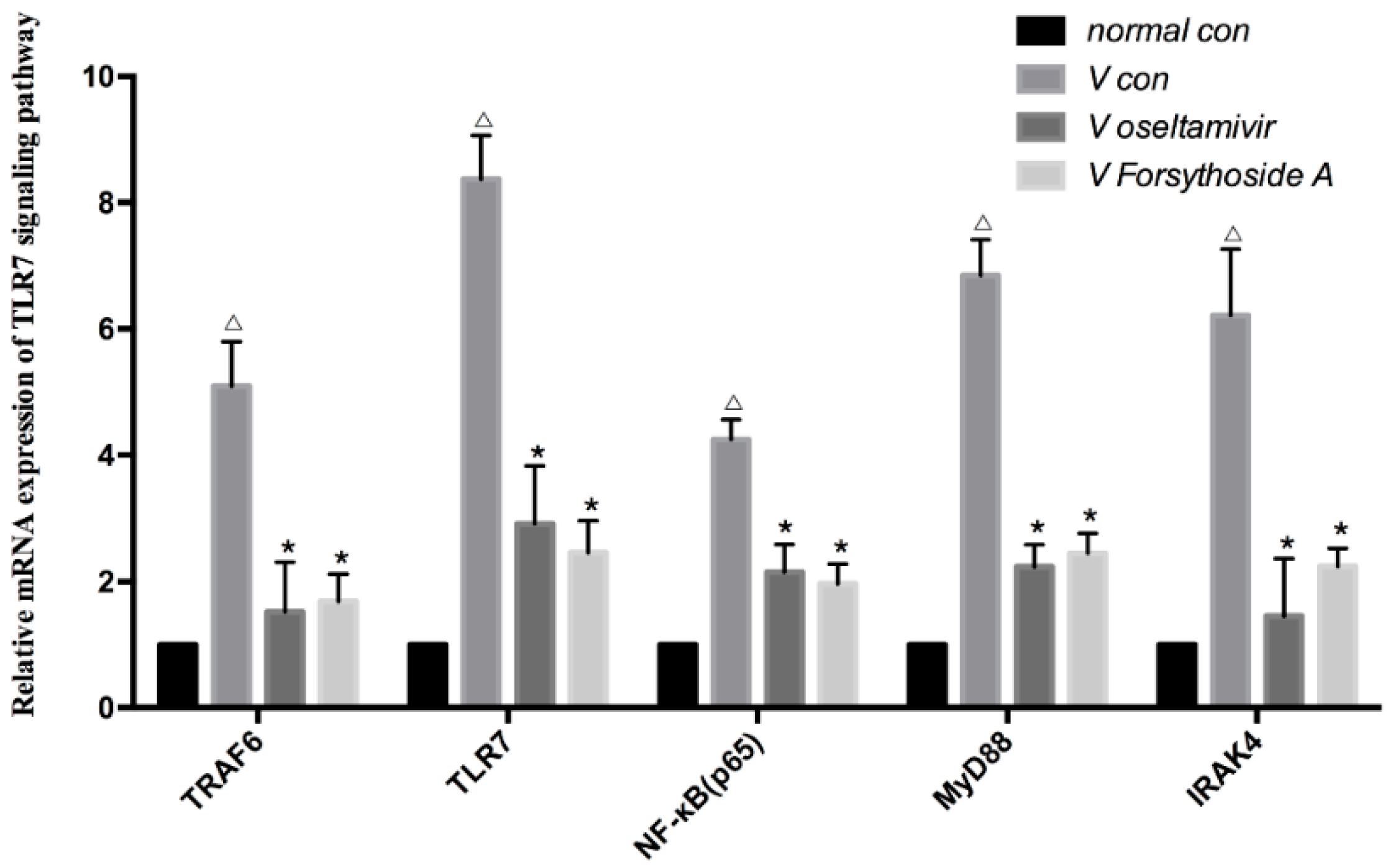
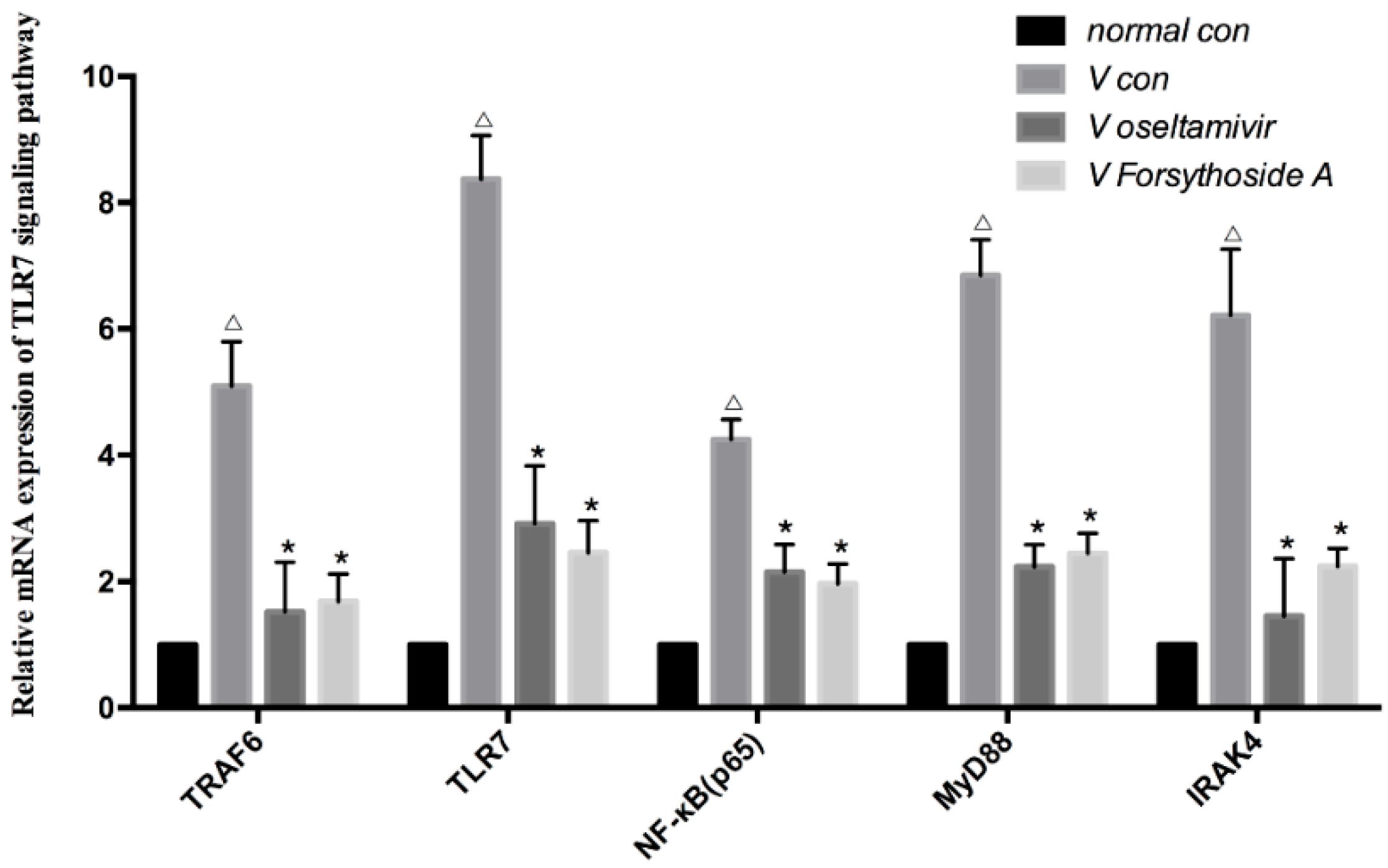
2.4. Relative mRNA Expression of TLR7, MyD88, TRAF6, IRAK4 and NF-κB
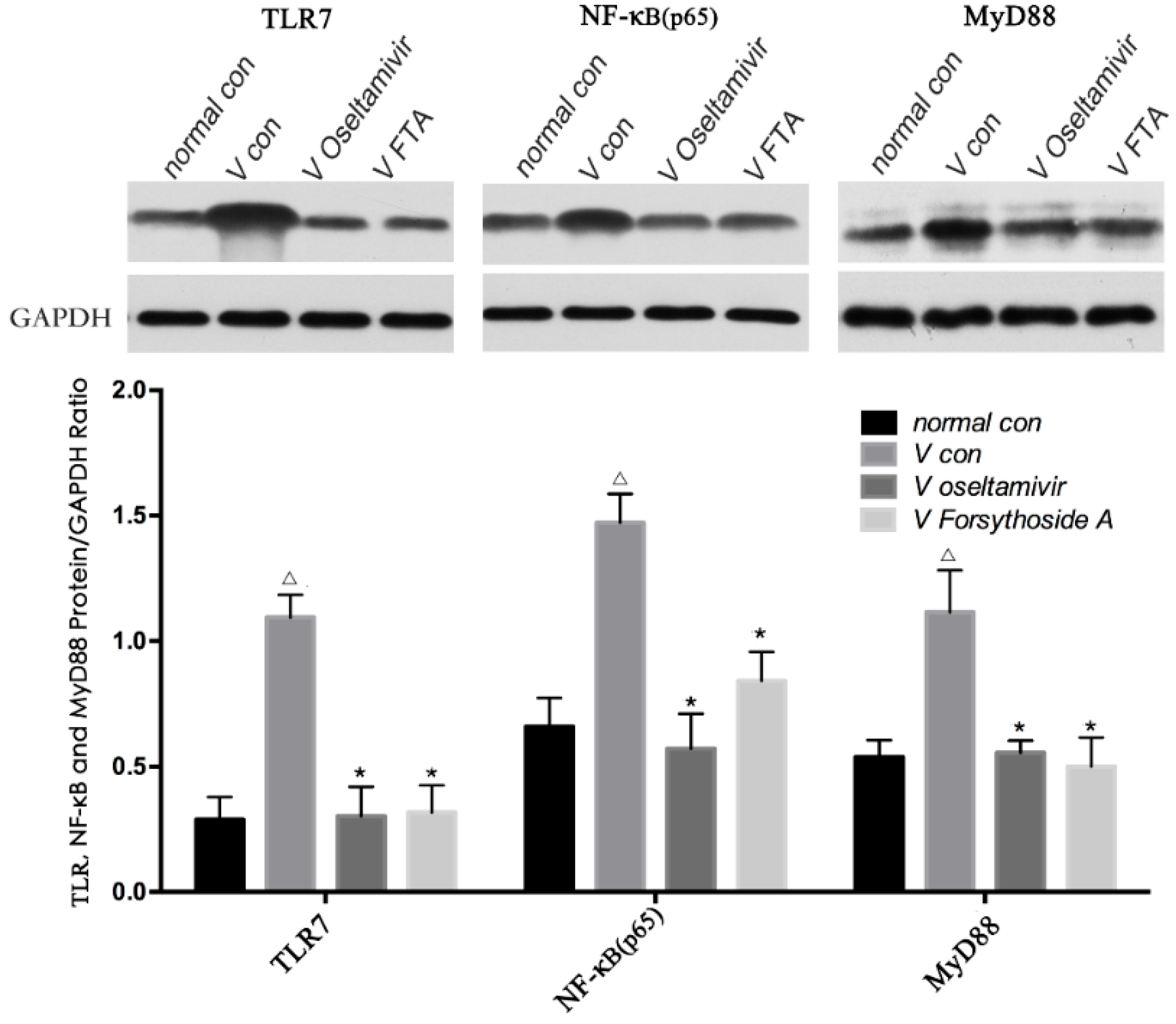
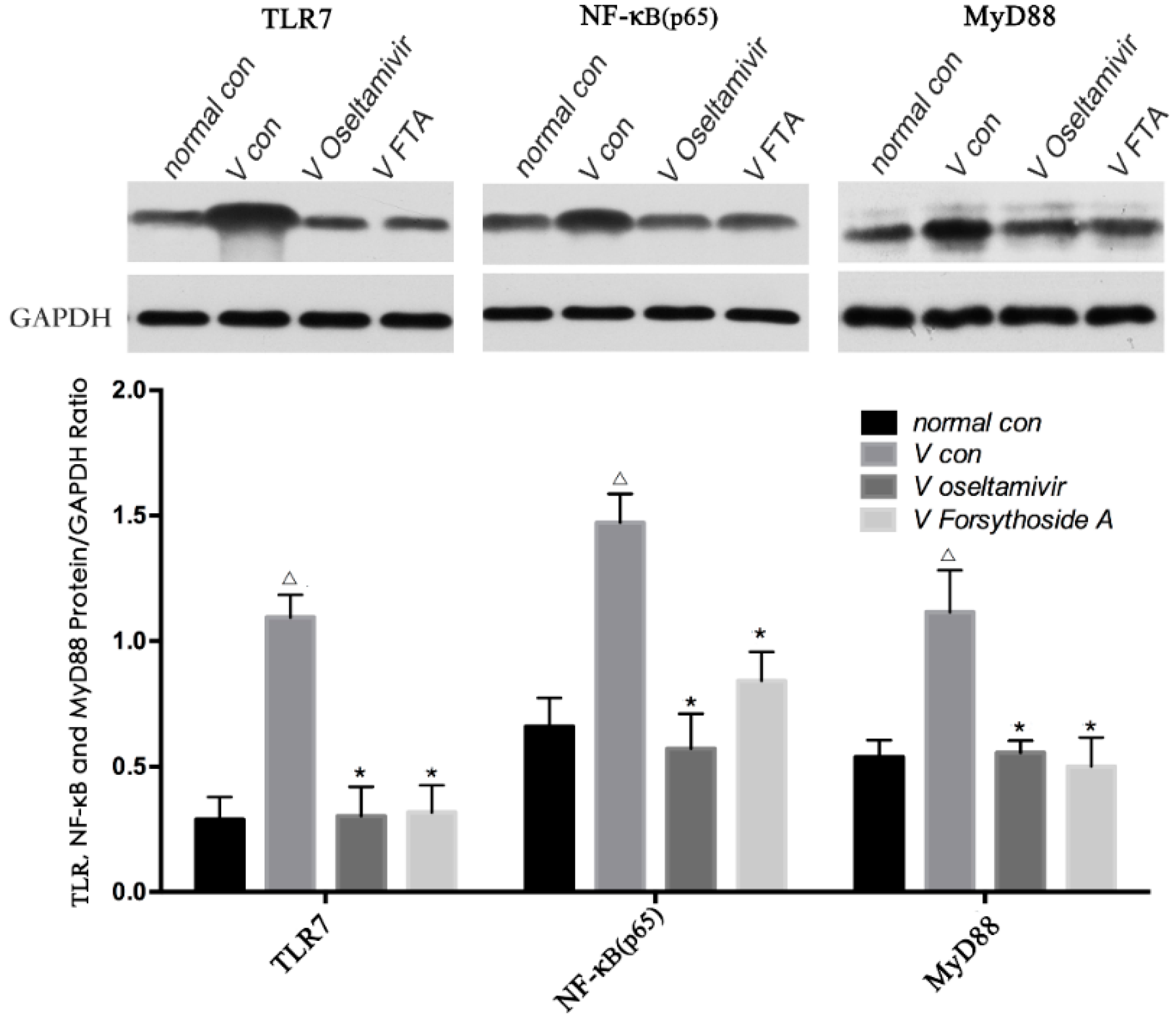
2.5. Relative Protein Expression of TLR7, MyD88 and NF-κB p65
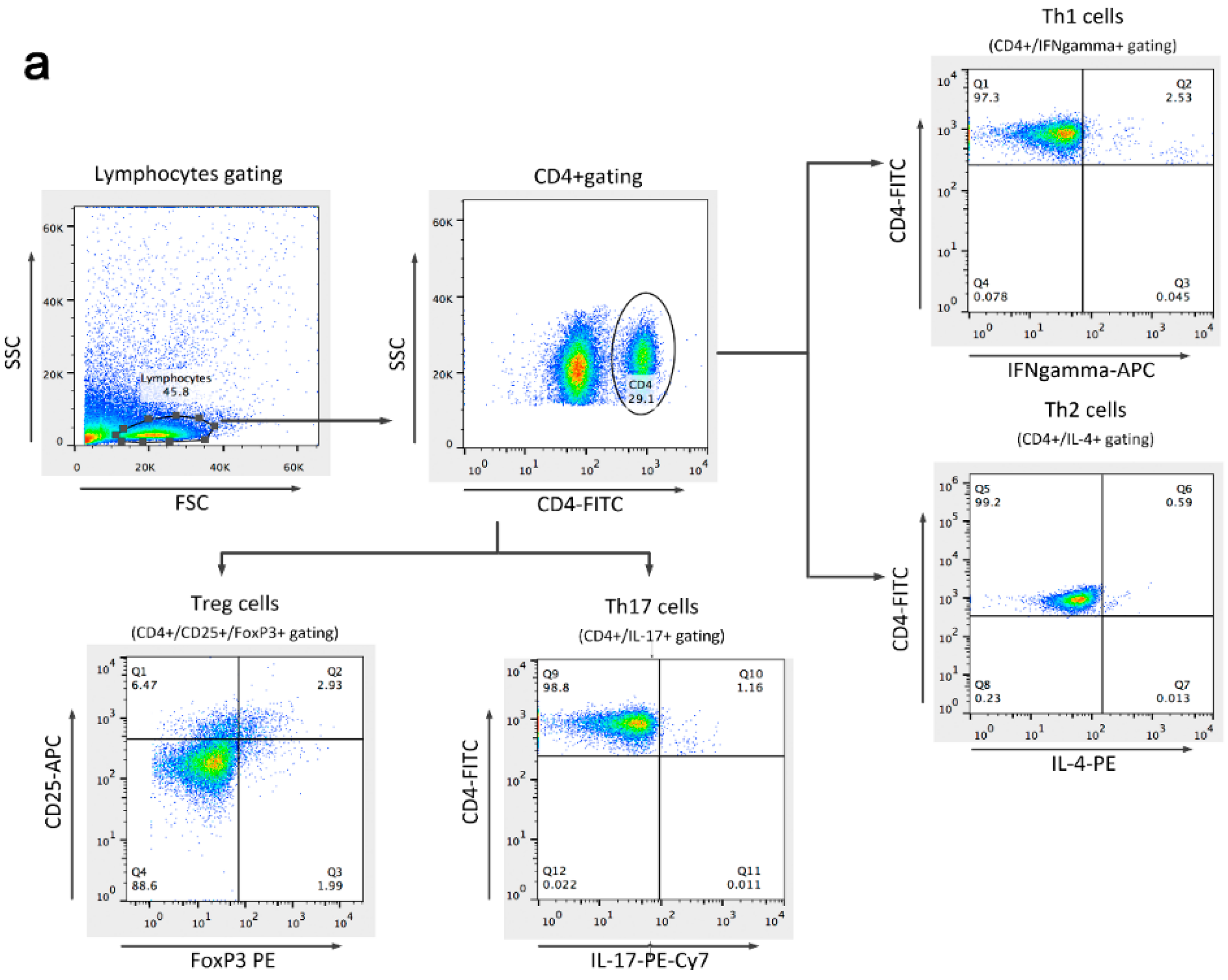
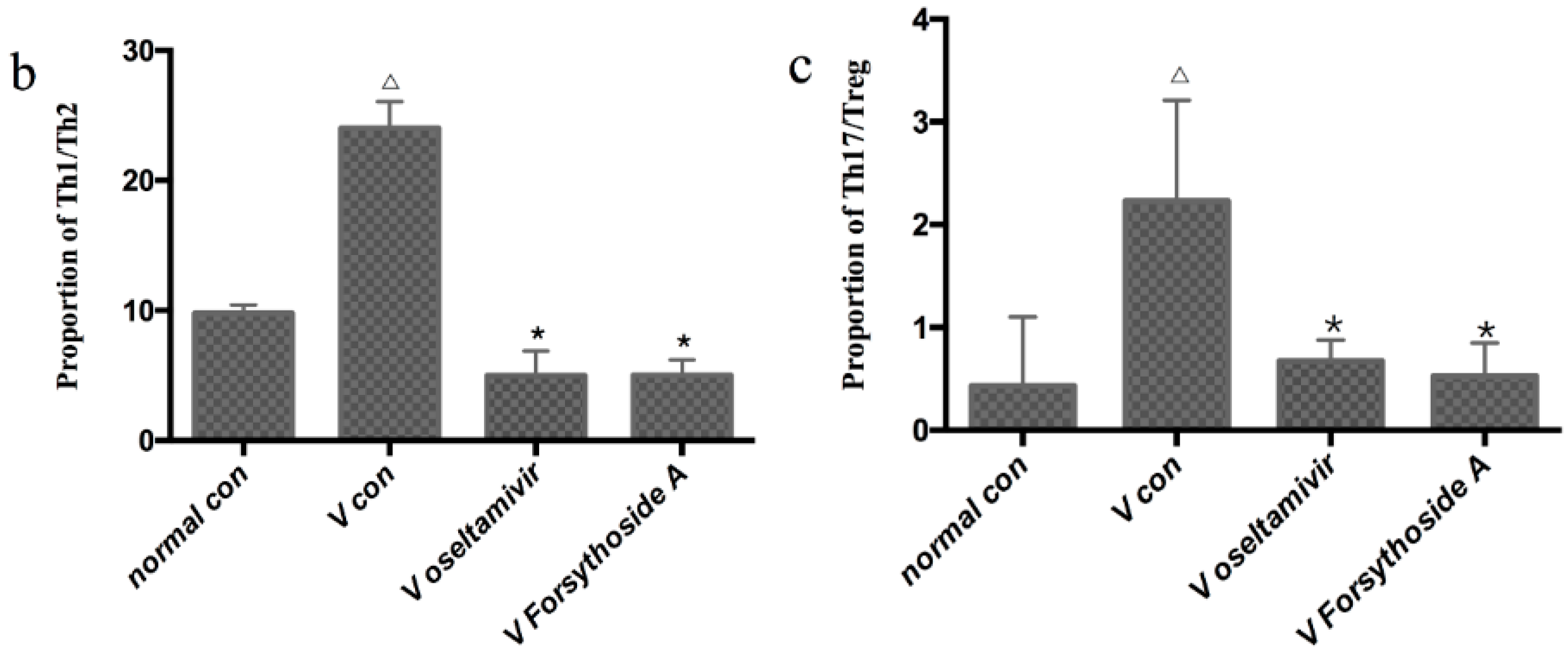
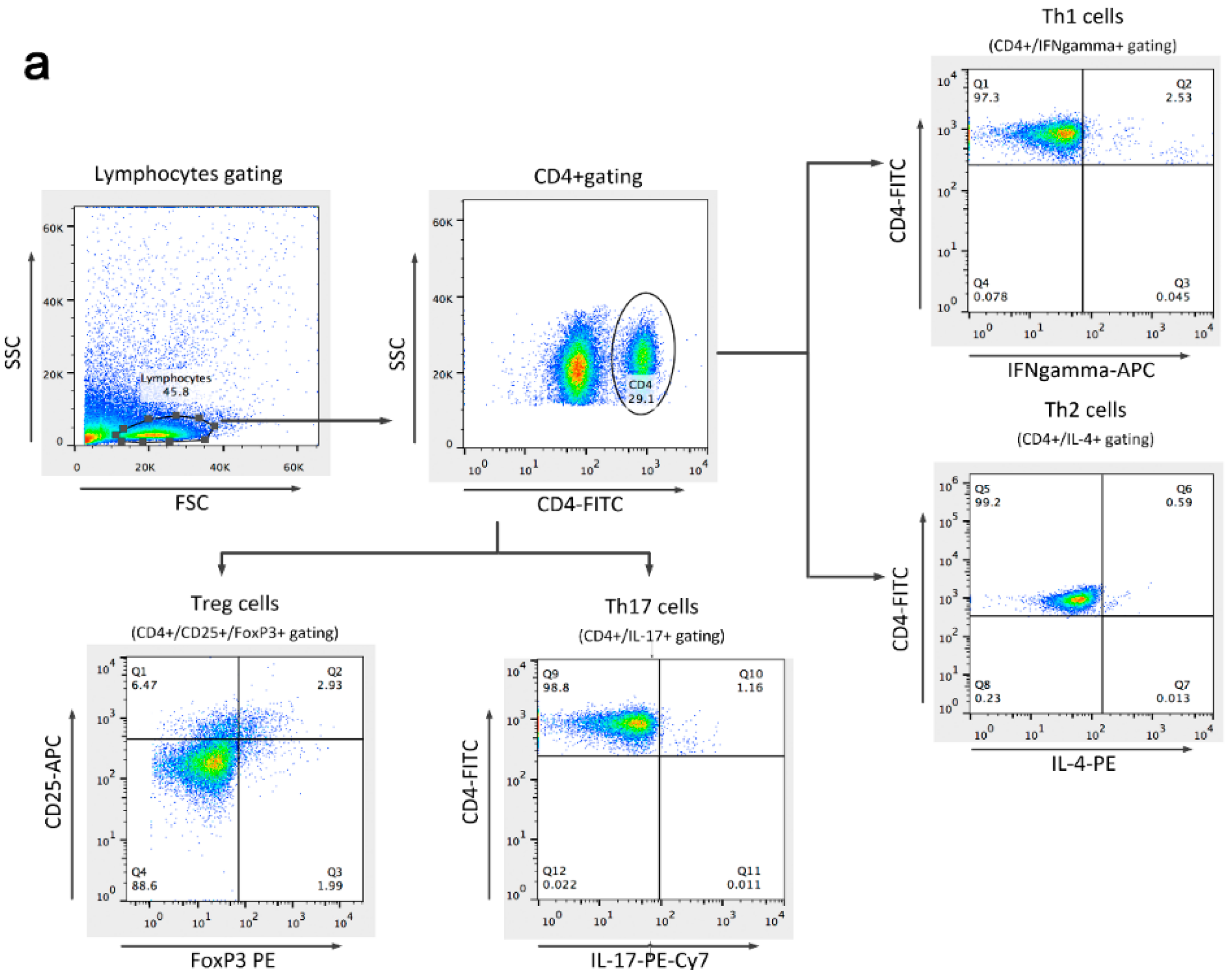
2.6. Detection of Th1, Th2, Th17, and Treg Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Grouping, Virus and Treatment
4.3. Histopathological Examination
4.4. RT-qPCR of TLR7, MyD88, TRAF6, IRAK4 and NF-κB p65 mRNA and Relative Expression of the Influenza A Virus in Lung
4.5. Western Blot for TLR7, MyD88 and NF-κB p65
4.6. Immunofluorescence Labeling and Flow Cytometry
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oxford, J.S. Influenza A pandemics of the 20th century with special reference to 1918: Virology, pathology and epidemiology. Rev. Med. Virol. 2000, 10, 119–133. [Google Scholar] [CrossRef]
- Potter, C.W. A history of influenza. J. Appl. Microbiol. 2001, 91, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Yuen, K.Y.; Chan, P.K.; Peiris, M.; Tsang, D.N.; Que, T.L.; Shortridge, K.F.; Cheung, P.T.; To, W.K.; Ho, E.T.; Sung, R.; et al. Clinical features and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. Lancet 1998, 351, 467–471. [Google Scholar] [CrossRef]
- Wu, S.; Jiang, Z.-Y.; Sun, Y.-F.; Yu, B.; Chen, J.; Dai, C.-Q.; Wu, X.-L.; Tang, X.-L.; Chen, X.-Y. Microbiota regulates the TLR7 signaling pathway against respiratory tract influenza A virus infection. Curr. Microbiol. 2013, 67, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.-J.; Hung, T.M.; Min, B.-S.; Kim, J.-C.; Woo, M.H.; Choi, J.S.; Lee, H.K.; Bae, K. Lignans from the Fruits of Forsythia suspensa (Thunb.) Vahl Protect High-Density Lipoprotein during Oxidative Stress. Biosci. Biotechnol. Biochem. 2008, 72, 2750–2755. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Fu, Y.-J.; Zu, Y.-G.; Luo, M.; Wang, W.; Zhang, L.; Li, J. Enzyme-assisted microwave hydro-distillation essential oil from Fructus forsythia, chemical constituents, and its antimicrobial and antioxidant activities. Food Chem. 2012, 134, 235–243. [Google Scholar] [CrossRef]
- Xing, S.; Peng, Y.; Wang, M.; Chen, D.; Li, X. In vitro human fecal microbial metabolism of Forsythoside A and biological activities of its metabolites. Fitoterapia 2014, 99, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Zhang, Z.W.; Li, H.W.; Sun, J.H.; Xu, C.Y.; Wu, G.J. Effects of forsythoside A on the Expression of IFN-α and Mx1. Sci. Agric. Sin. 2010. [Google Scholar] [CrossRef]
- Saenz, R.A.; Quinlivan, M.; Elton, D.; MacRae, S.; Blunden, A.S.; Mumford, J.A.; Daly, J.M.; Digard, P.; Cullinane, A.; Grenfell, B.T.; et al. Dynamics of Influenza Virus Infection and Pathology. J. Virol. 2010, 84, 3974–3983. [Google Scholar] [CrossRef] [PubMed]
- Myers, K.P.; Olsen, C.W.; Gray, G.C. Cases of swine influenza in humans: A review of the literature. Clin. Infect. Dis. 2007, 44, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Dotis, J.; Roilides, E. H1N1 Influenza A infection. Hippokratia 2009, 13, 135–138. [Google Scholar] [PubMed]
- McClellan, K.; Perry, C.M. Oseltamivir—A review of its use in influenza. Drugs 2001, 61, 263–283. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, T.; Jones, M.; Doshi, P.; Spencer, E.A.; Onakpoya, I.; Heneghan, C.J. Oseltamivir for influenza in adults and children: Systematic review of clinical study reports and summary of regulatory comments. Br. Med. J. 2014, 348. [Google Scholar] [CrossRef] [PubMed]
- Wallensten, A.; Oliver, I.; Lewis, D.; Harrison, S. Compliance and Side Effects of Prophylactic Oseltamivir Treatment in a School in South West England. Eurosurveillance 2009, 14, 6–9. [Google Scholar]
- Matsuzaki, Y.; Mizuta, K.; Aoki, Y.; Suto, A.; Abiko, C.; Sanjoh, K.; Sugawara, K.; Takashita, E.; Itagaki, T.; Katsushima, Y.; et al. A two-year survey of the oseltamivir-resistant influenza A(H1N1) virus in Yamagata, Japan and the clinical effectiveness of oseltamivir and zanamivir. Virol. J. 2010, 7. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Huifu, X.; Hao, H. Effects of the extract of Forsythia suspensa on influenza A H1N1 infection in vitro. J. Med. Plants Res. 2010, 4, 1468–1471. [Google Scholar]
- Baharom, F.; Thomas, S.; Bieder, A.; Hellmer, M.; Volz, J.; Sandgren, K.J.; McInerney, G.M.; Karlsson Hedestam, G.B.; Mellman, I.; Smed-Sorensen, A. Protection of human myeloid dendritic cell subsets against influenza A virus infection is differentially regulated upon TLR stimulation. J. Immunol. 2015, 194, 4422–4430. [Google Scholar] [CrossRef] [PubMed]
- Koyama, S.; Ishii, K.J.; Kumar, H.; Tanimoto, T.; Coban, C.; Uematsu, S.; Kawai, T.; Akira, S. Differential role of TLR- and RLR-signaling in the immune responses to influenza A virus infection and vaccination. J. Immunol. 2007, 179, 4711–4720. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Wang, H.; Han, X.; Lin, Y.; Yang, Y.; Gu, L.; Zhao, J.; Wang, L.; Huang, L.; Li, Y.; et al. Baicalin inhibits TLR7/MYD88 signaling pathway activation to suppress lung inflammation in mice infected with influenza A virus. Biomed. Rep. 2014, 2, 437–441. [Google Scholar] [PubMed]
- Brightbill, H.D.; Modlin, R.L. Toll‐like receptors: Molecular mechanisms of the mammalian immune response. Immunology 2000, 101. [Google Scholar] [CrossRef]
- Zhang, A.J.X.; Li, C.; To, K.K.W.; Zhu, H.-S.; Lee, A.C.Y.; Li, C.-G.; Chan, J.F.W.; Hung, I.F.N.; Yuen, K.-Y. Toll-like receptor 7 agonist imiquimod in combination with influenza vaccine expedites and augments humoral immune responses against influenza A(H1N1)pdm09 virus infection in BALB/c mice. Clin. Vaccine Immunol. 2014, 21, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Galgani, M.; de Rosa, V.; Matarese, G. T cell metabolism and susceptibility to autoimmune diseases. Mol. Immunol. 2015, 68, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.E.; Slaughter, N. T-cell activation through the antigen receptor. Part 2: Role of signaling cascades in T-cell differentiation, anergy, immune senescence, and development of immunotherapy. J. Allergy Clin. Immunol. 2002, 109, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Yamamoto, M.; Ruan, J.; Choi, J.Y.; Gauvreau, G.M.; Olek, S.; Hoffmueller, U.; Carlsten, C.; FitzGerald, J.M.; Boulet, L.-P.; et al. Th17/Treg ratio derived using DNA methylation analysis is associated with the late phase asthmatic response. Allergy Asthma Clin. Immunol. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.H.; Fang, Y.Q.; Ryan, U.; Guo, Y.X.; Wu, F.; Du, S.Z.; Chen, D.K.; Lin, Q. Dynamics of Th17 associating cytokines in Cryptosporidium parvum-infected mice. Parasitol. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, G.; Kumberova, A.; Croft, N.M.; McKenzie, C.; Steer, H.W.; MacDonald, T.T. Blocking Smad7 restores TGF-β1 signaling in chronic inflammatory bowel disease. J. Clin. Investig. 2001, 108, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Bierie, B.; Moses, H.L. Transforming growth factor beta (TGF-β) and inflammation in cancer. Cytokine Growth Factor Rev. 2010, 21, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, J.; Zhang, Z.; Ma, Y.; Liao, F.; Zhang, Y.; Wu, G. Forsythoside A Inhibits the Avian Infectious Bronchitis Virus in Cell Culture. Phytother. Res. 2011, 25, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Kernbauer, E.; Ding, Y.; Cadwell, K. An enteric virus can replace the beneficial function of commensal bacteria. Nature 2014, 516, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.S.; Bauer, C.M.T.; Vujicic, N.; Gaschler, G.J.; Lichty, B.D.; Brown, E.G.; Stämpfli, M.R. Cigarette Smoke Impacts Immune Inflammatory Responses to Influenza in Mice. Am. J. Respir. Crit. Care Med. 2012, 174, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| GAPDH | Forward primer | 5′-TGATGACATCAAGAAGGTGGTGAAG-3′ |
| Reverse primer | 5′-TCCTTGGAGGCCATGTAGGCCAT-3′ | |
| TLR7 | Forward primer | 5′-GGGTCCAAAGCCAATGTG-3′ |
| Reverse primer | 5′-TGTTAGATTCTCCTTCGTGATG-3′ | |
| MyD88 | Forward primer | 5′-CGATTATCTACAGAGCAAGGAATG-3′ |
| Reverse primer | 5′-ATAGTGATGAACCGCAGGATAC-3′ | |
| TRAF6 | Forward primer | 5′-TTGGAGAGTCGCCTAGTAAG-3′ |
| Reverse primer | 5′-GTTACACTGCTGTGCTTCC-3′ | |
| IRAK4 | Forward primer | 5′-CATCGTGGCGGTGAAGAAG-3′ |
| Reverse primer | 5′-AGCATACACTAAGCACAGGTTG-3′ | |
| NF-κB p65 | Forward primer | 5′-ATTCTGACCTTGCCTATCTAC-3′ |
| Reverse primer | 5′-TCCAGTCTCCGAGTGAAG-3′ | |
| Virus Replication | Forward primer | 5′-GACCAATCCTGTCACCTCTGAC-3′ |
| Reverse primer | 5′-GGGCATTTGGACAAACGTCTACG-3′ |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, L.; Pang, P.; Zheng, K.; Nie, J.; Xu, H.; Wu, S.; Chen, J.; Chen, X. Forsythoside A Controls Influenza A Virus Infection and Improves the Prognosis by Inhibiting Virus Replication in Mice. Molecules 2016, 21, 524. https://doi.org/10.3390/molecules21050524
Deng L, Pang P, Zheng K, Nie J, Xu H, Wu S, Chen J, Chen X. Forsythoside A Controls Influenza A Virus Infection and Improves the Prognosis by Inhibiting Virus Replication in Mice. Molecules. 2016; 21(5):524. https://doi.org/10.3390/molecules21050524
Chicago/Turabian StyleDeng, Li, Peng Pang, Ke Zheng, Jiao Nie, Huachong Xu, Sizhi Wu, Jia Chen, and Xiaoyin Chen. 2016. "Forsythoside A Controls Influenza A Virus Infection and Improves the Prognosis by Inhibiting Virus Replication in Mice" Molecules 21, no. 5: 524. https://doi.org/10.3390/molecules21050524
APA StyleDeng, L., Pang, P., Zheng, K., Nie, J., Xu, H., Wu, S., Chen, J., & Chen, X. (2016). Forsythoside A Controls Influenza A Virus Infection and Improves the Prognosis by Inhibiting Virus Replication in Mice. Molecules, 21(5), 524. https://doi.org/10.3390/molecules21050524