
The Effects of Fungicide, Soil Fumigant, Bio-Organic Fertilizer and Their Combined Application on Chrysanthemum Fusarium Wilt Controlling, Soil Enzyme Activities and Microbial Properties




Abstract
:1. Introduction
2. Results
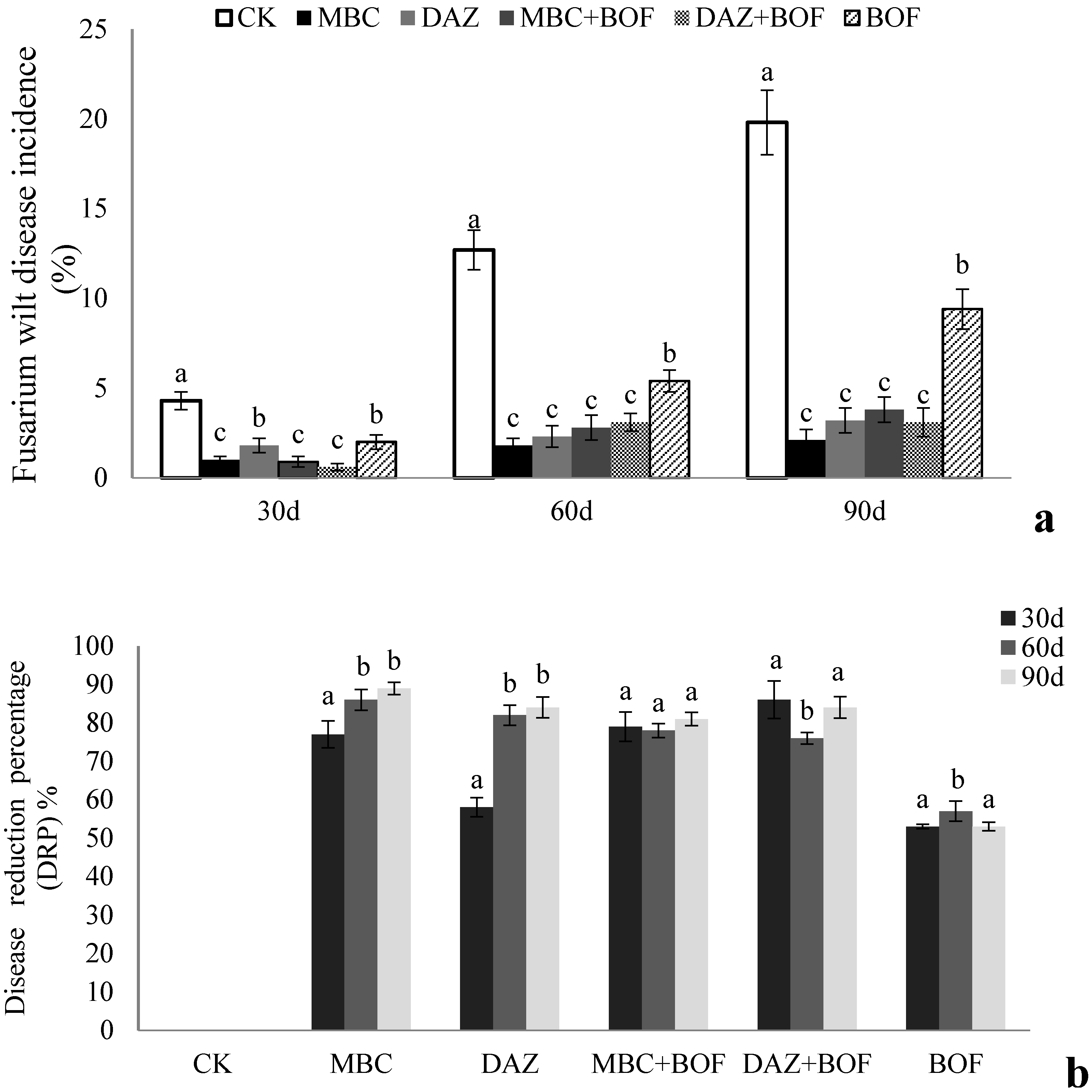
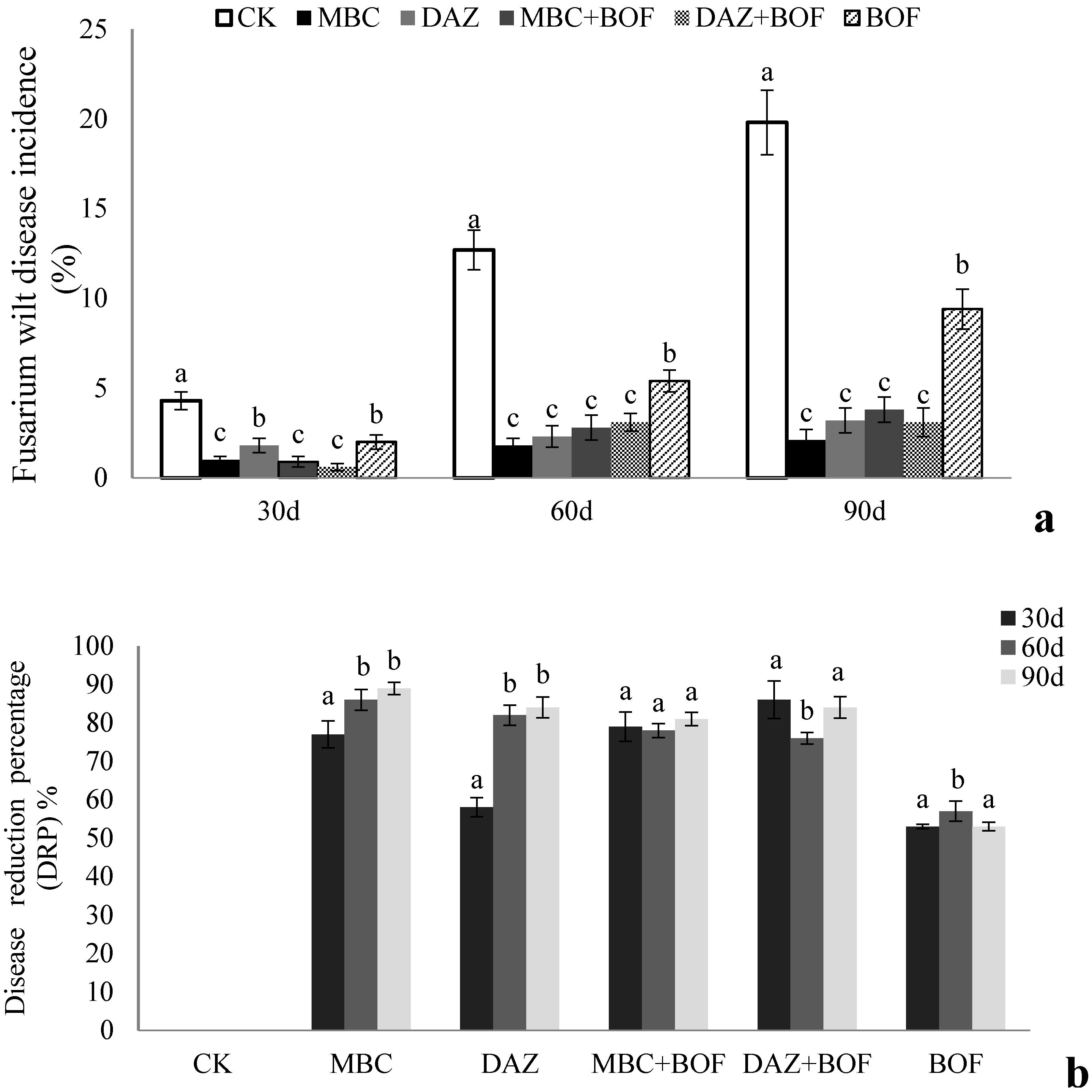
2.1. The Impact of Fusarium Wilt on Chrysanthemum Growth
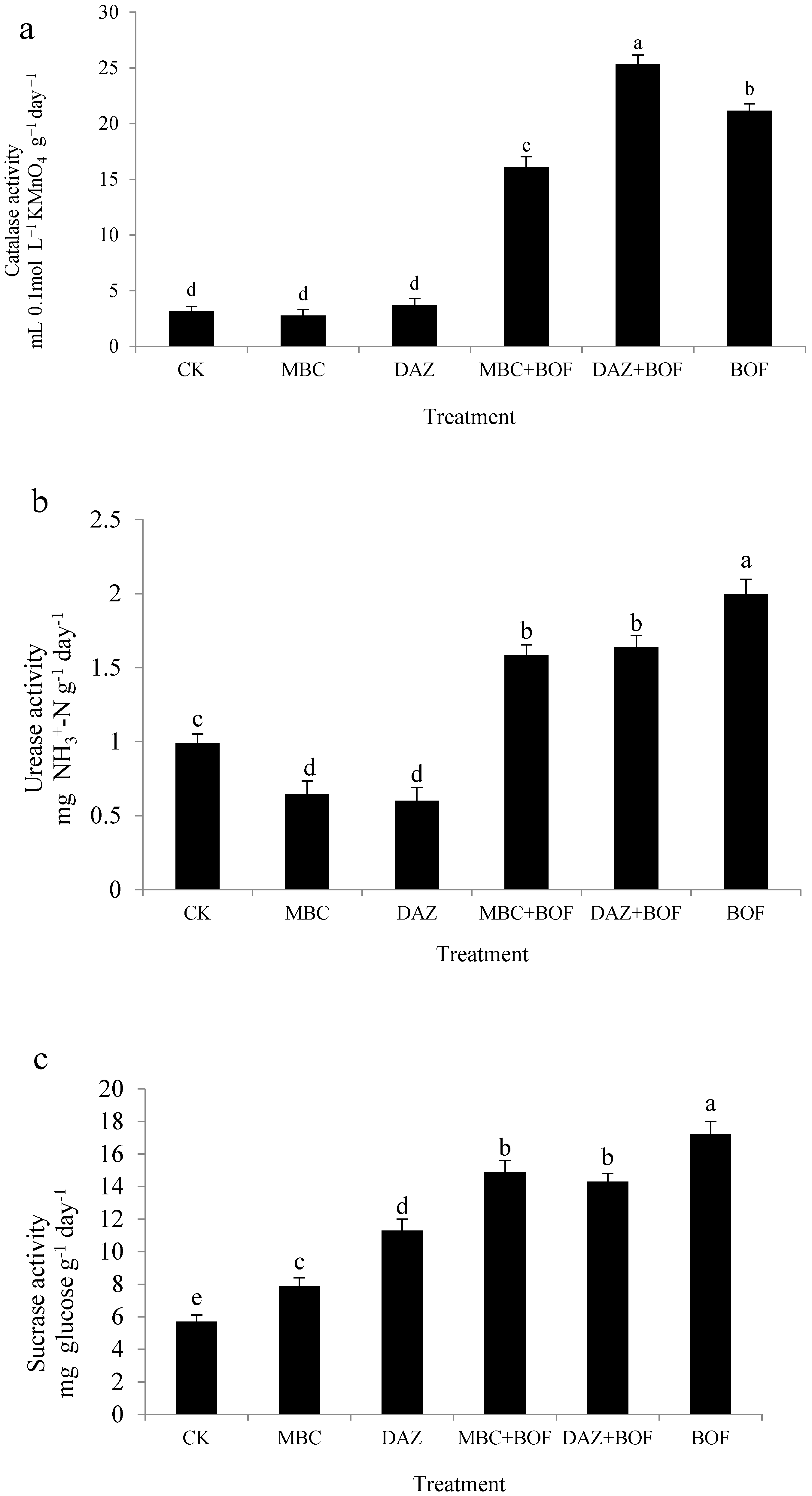
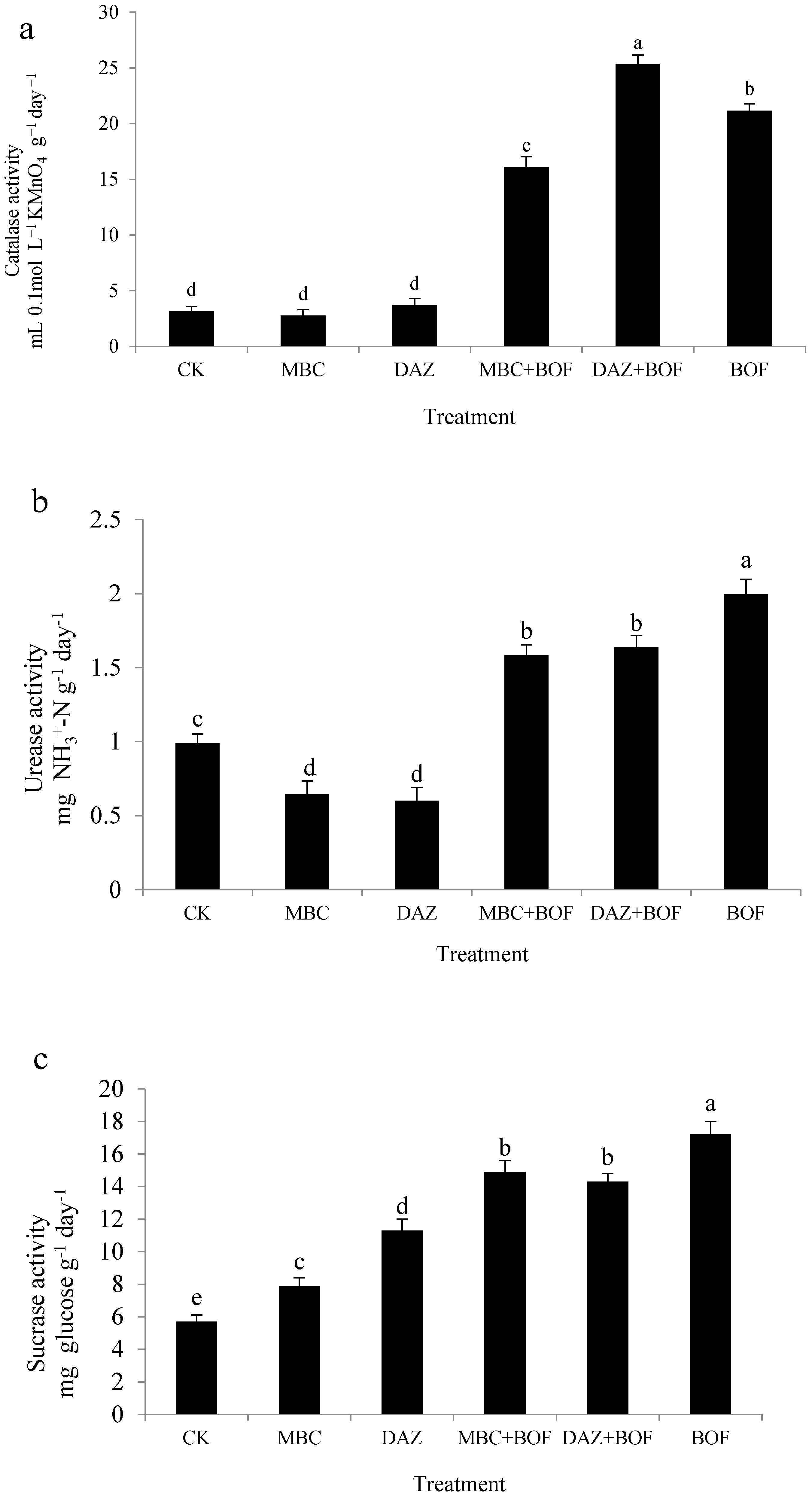
2.2 Enzymatic Activity
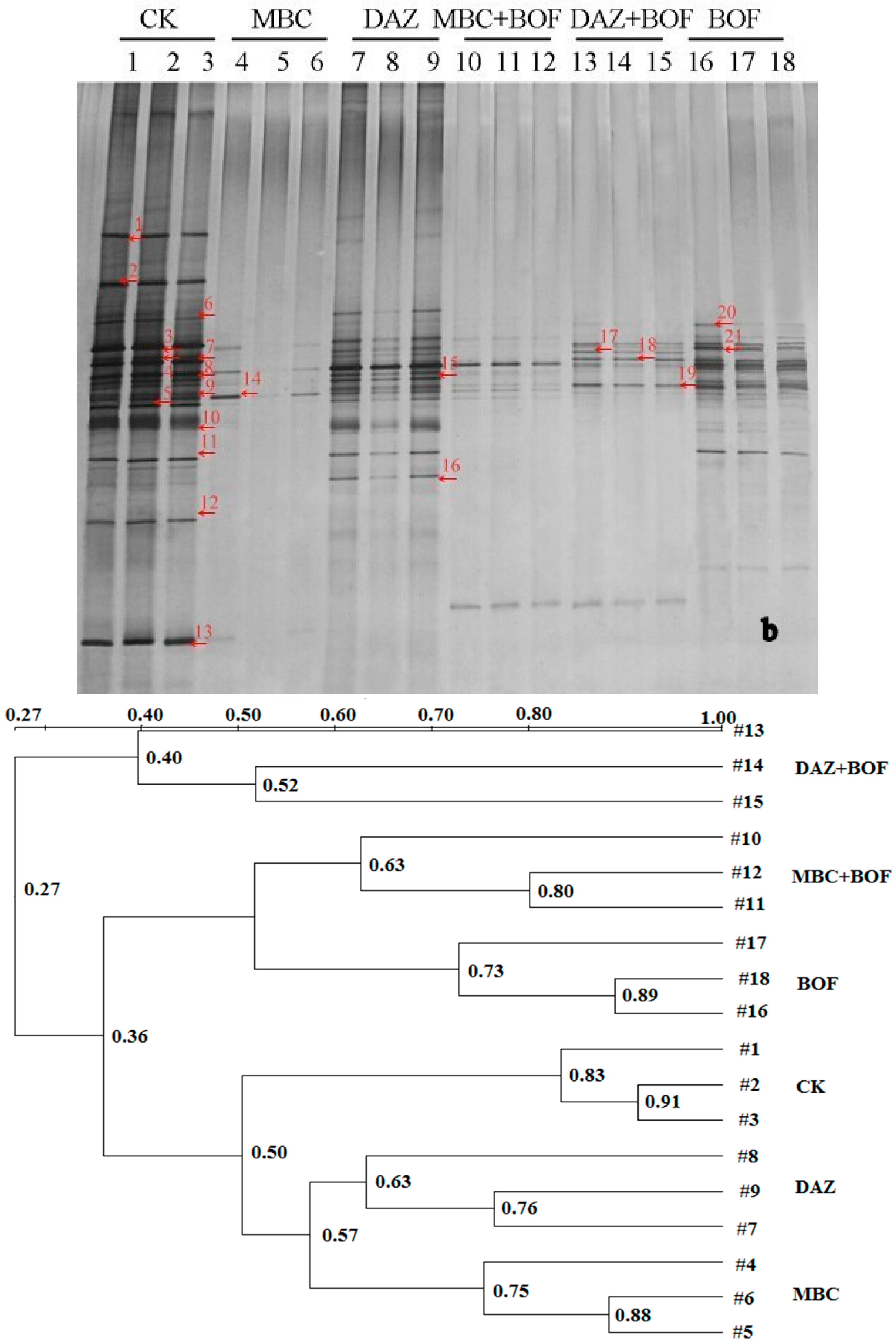
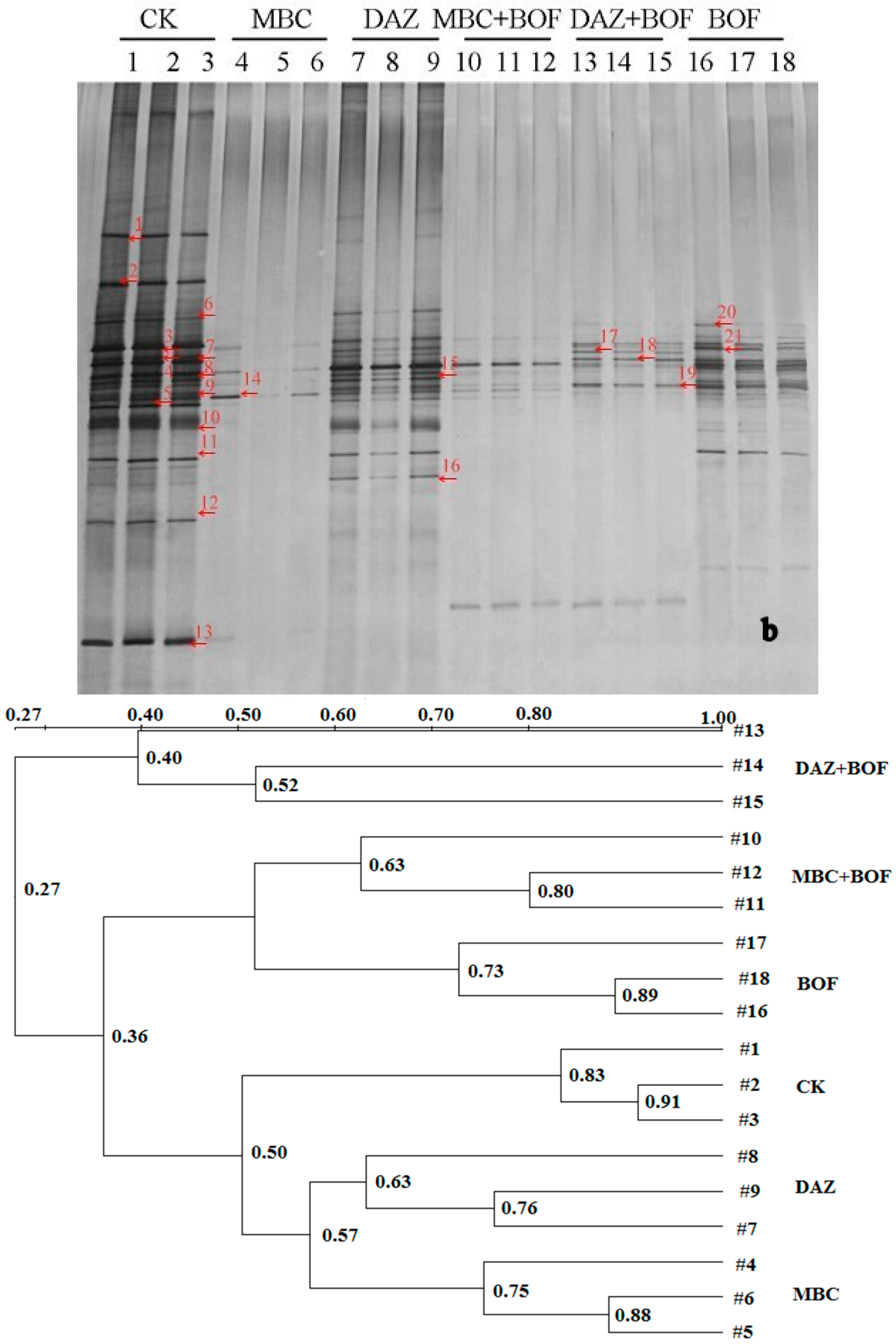
2.3. Diversity of the Soil Microbiota
3. Discussion
3.1. The Effects of the Soil Treatments on Fusarium Wilt Incidence
3.2. The Effect of the Various Soil Treatments on Soil Microbiota Composition
3.3. The Effect of the Various Treatments on Chrysanthemum Productivity
4. Materials and Methods
4.1. Site Description and Plant Material
4.2. BOF and Fungicide and Soil Fumigant
4.3. Soil Treatment and Sampling
4.3.1. The Measurement of Growth and Disease Incidence
4.3.2. Soil DNA Extraction and PCR Amplification
4.3.3. Denaturing Gradient Gel Electrophoresis (DGGE) Profiling
4.3.4. Real-Time PCR
4.3.5. Enzymatic Activity
4.4. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duineveld, B.M.; Rosado, A.S.; van Elsas, J.D.; van Veen, J.A. Analysis of the dynamics of bacterial communitiesin the rhizosphere of chrysanthemum via denaturing gradient gelelectrophoresis and substrate utilization patterns. Appl. Environ. Microbiol. 1998, 64, 4950–4957. [Google Scholar] [PubMed]
- Duineveld, B.M.; Kowalchuk, G.A.; Keijzer, A.; van Elsas, J.D.; van Veen, J.A. Analysis of bacterial communities in the rhizosphere of chrysanthemum via denaturing gradient gel electrophoresis of PCR-Amplified 16S rRNA as well as DNA fragments coding for 16S rRNA. Appl. Environ. Microbiol. 2001, 67, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Farias, M.F.; Saad, J.C.C. Growth and quality of chrysanthemum produced in greenhouse. Hortic. Bras. 2005, 23, 740–742. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Sawicka, A. Effect of carbendazim, imazetapir and thiram on nitrogenase activity, number of microorganisms in soil and yield of hybrid lucerne (Medicago media). Pol. J. Environ. Stud. 2002, 11, 737–744. [Google Scholar]
- Nannipieri, P.; Grego, S.; Ceccanti, B. Ecological significance of the biological activityin soil. In Soil Biochemistry; Bollag, J.M., Stotzky, G., Eds.; Marcel Dekker: New York, NY, USA, 1990; Volume 6, pp. 293–355. [Google Scholar]
- Chen, S.K.; Edwards, C.A.; Subler, S. Effect of fungicides benomyl, captan and chlorothalonil on soil microbial activity and nitrogen dynamics in laboratory incubations. Soil Biol. Biochem. 2001, 33, 1971–1980. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Lagomarsino, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasierm, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Ai, Liang; Sun, C.G.J.; Wang, X.; Zhou, W. Responses of extracellular enzyme activities and microbial community in both the rhizosphere and bulk soil to long-term fertilization practices in a fluvo-aquic soil. Geoderma 2012, 330–338, 173–174. [Google Scholar]
- Speir, T.W.; Lee, R.; Pansier, E.A.; Cairns, A. A comparison of sulphatase, urease and protease activities in planted and in fallow soils. Soil Biol. Biochem. 1980, 12, 281–291. [Google Scholar] [CrossRef]
- Frankenberger, W.T.; Dick, W.A. Relationships between enzyme ctivities and microbial growth and activity in soil. Soil Sci. Soc. Am. J. 1983, 47, 945–951. [Google Scholar] [CrossRef]
- Zhang, S.; Raza, W.; Yang, X.M.; Hu, J.; Huang, Q.W.; Xu, Y.C.; Liu, X.; Ran, W.; Shen, Q.R. Control of Fusarium wilt disease of cucumber plants with the application of a bioorganic fertilizer. Biol. Fertil. Soils 2008, 4, 1073–1080. [Google Scholar] [CrossRef]
- Zhang, N.; Wu, K.; He, X.; Li, S.Q.; Zhang, Z.H.; Shen, B.; Yang, X.M.; Zhang, R.F.; Huang, Q.W.; Shen, Q.R. A new bioorganic fertilizer can effectively control banana wilt by strong colonization of Bacillus subtilis N11. Plant Soil 2011, 344, 87–97. [Google Scholar] [CrossRef]
- Luo, J.; Ran, W.; Hu, J.; Yang, X.M.; Xu, Y.C.; Shen, Q.R. Application of bio-organic fertilizer significantly affected fungal diversity of soils. Soil Sci. Soc. Am. J. 2010, 74, 2039–2048. [Google Scholar] [CrossRef]
- Shen, Z.Z.; Zhong, S.T.; Wang, Y.G.; Mei, X.L.; Wang, B.B.; Li, R.; Ruan, Y.Z.; Shen, Q.R. Induced soil microbial suppression of banana Fusarium wilt disease using compost and bio fertilizers to improve yield and quality. Eur. J. Soil Biol. 2013, l57, 1–8. [Google Scholar] [CrossRef]
- Minuto, A.; Davide, S.; Garibaldi, A.; Gullino, M.L. Control of soil-borne pathogens of tomato using a commercial formulation of Streptomyces griseoviridis and solarization. Crop Prot. 2006, 25, 468–475. [Google Scholar] [CrossRef]
- Ling, N.; Zhang, W.W.; Tan, S.Y.; Huang, Q.W.; Shen, Q.R. Effect of the nursery application of bioorganic fertilizer on spatial distribution of Fusarium oxysporum f. sp. niveum and its antagonistic bacterium in the rhizosphere of watermelon. Appl. Soil Ecol. 2012, 59, 13–19. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, D.Y.; Ling, N.; Chen, F.; Fan, W.; Shen, Q.R. Bio-organic fertilizer application significantly reduces the Fusarium oxysporum population and alters the composition of fungi communities of watermelon Fusarium wilt rhizosphere soil. Biol. Fertil. Soils 2014, 50, 76. [Google Scholar] [CrossRef]
- Dick, R.P. Soil enzyme activities as indicators of soil quality. In Defining Soil Quality for A Sustainable Environment; No. 35.; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; SSSA Special Publication: ASA, CSSA, SSA, Madison, WI, USA, 1994; pp. 107–124. [Google Scholar]
- Bending, G.D.; Turner, M.K.; Rayns, F.; Marx, M.C.; Wood, M. Microbial and biochemical soil quality indicators and their potential for differentiating areas under contrasting agricultural management regimes. Soil Biol. Biochem. 2004, 36, 1785–1792. [Google Scholar] [CrossRef]
- Ingram, C.W.; Coyne, M.S.; Williams, D.W. Effects of commercial diazinon and imidacloprid on microbial urease activity in soil. J. Environ. Qual. 2005, 34, 1573–1580. [Google Scholar] [CrossRef] [PubMed]
- Ling, N.; Wang, D.S.; Zhu, C.; Song, Y.; Yu, G.H.; Ran, W.; Huang, Q.W.; Guo, S.W.; Shen, Q.R. Response of the population size and community structure of Paenibacillus spp. to different fertilization regimes in a long-term experiment of red soil. Plant Soil 2014, 383, 87–98. [Google Scholar] [CrossRef]
- Ling, N.; Sun, Y.M.; Ma, J.H.; Guo, J.J.; Zhu, P.; Peng, C.; Yu, G.H.; Ran, W.; Guo, S.W.; Shen, Q.R. Response of the bacterial diversity and soil enzyme activity in particle-size fractions of Mollisol after different fertilization in along-term experiment. Biol. Fertil. Soils. 2013, 50, 901–911. [Google Scholar] [CrossRef]
- Boldrin, B.; Tiehm, A.; Fritzsche, C. Degradation of phenanthrene, fluorene, fluoranthene, and pyrene by a Mycobacterium sp. Appl. Environ. Microbiol. 1993, 59, 1927–1930. [Google Scholar] [PubMed]
- Gai, Z.H.; Gai, Z.; Yu, B.; Li, L.; Wang, Y.; Ma, C.; Feng, J.; Deng, Z.; Xu, P. Cometabolic degradation of dibenzofuran and dibenzothiophene by a newly isolated carbazole-degrading Sphingomonas sp. strain. Appl. Environ. Microbiol. 2007, 73, 2832–2838. [Google Scholar] [CrossRef] [PubMed]
- Dussault, M.; Becaert, V.; Francois, M.; Sauve, S.; Deschenes, L. Effect of copper on soil functional stability measured by relative soil stability index (RSSI) based on two enzyme activities. Chemosphere 2008, 72, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Cunliffe, M.; Kertesz, M.A. Effect of Sphingobium yanoikuyae B1 inoculation on bacterial community dynamics and polycyclic aromatic hydrocarbon degradation in aged and freshly PAH-contaminated soils. Environ. Pollut. 2006, 144, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, S.R.; Albers, C.N.; Aamand, J. Rapid mineralization of the phenylurea herbicide diuron by Variovorax sp. strain SRS16 in pure culture and within a two-member consortium. Appl. Environ. Microbiol. 2008, 74, 2332–2340. [Google Scholar] [CrossRef] [PubMed]
- Dijksterhuis, J.; Sanders, M.; Gorris, L.G.M.; Smid, E.J. Antibiosis plays a role in the context of direct interaction during antagonism of Paenibacillus polymyxa towards Fusarium oxysporum. J. Appl. Microbiol. 1999, 86, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Wang, Y.; Shen, Q. R. Paenibacillus polymyxa: antibiotics, hydrolytic enzymes and hazard assessment. J. Plant Pathol. 2008, 90, 419–430. [Google Scholar]
- Lang, J.J.; Hu, J.; Ran, W.; Xu, Y.C.; Shen, Q. Control of cotton Verticillium wilt and fungal diversity of rhizosphere soils by bio-organic fertilizer. Biol. Fertil. Soils 2012, 48, 191–203. [Google Scholar] [CrossRef]
- Chen, L.H.; Huang, X.Q.; Zhang, F.G.; Zhao, D.K.; Yang, X.M.; Shen, Q.R. Applications of Trichoderma harzianum SQR-T037 bioorganic fertilizer significantly affect the microbial communities of continuously cropped soil of cucumber. J. Sci. Food Agric. 2012, 92, 2465–2470. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.H.; Zhang, R.F.; Xue, C.; Zhang, S.S.; Li, S.Q.; Zhang, N.; Shen, Q.R. Application of a novel bioorganic fertilizer can control Fusarium wilt by regulating microbial community of cucumber rhizosphere soils. Biol. Fertil. Soils 2012, 48, 807–816. [Google Scholar] [CrossRef]
- Wang, B.B.; Yuan, J.; Zhang, J.; Shen, Z.Z.; Zhang, M.X.; Li, R.; Ruan, Y.Z.; Shen, Q.R. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Burrows, L.A.; Edwards, C.A. The use of integrated soil microcosms to assess the impact of carbendazim on soil ecosystems. Ecotoxicology 2004, 13, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes, application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.Y.; Zhang, D.S.; Zhang, Z.M. Soil Enzymes and Its Methodology; Agricultural Press: Beijing, China, 1986; pp. 274–338. [Google Scholar]
- Liu, D.; Fang, S.; Tian, Y.; Dun, X. Variation in rhizosphere soil microbial index of tree species on seasonal flooding land: An in situ rhizobox approach. Appl. Soil Ecol. 2012, 59, 1–11. [Google Scholar] [CrossRef]
- Fang, S.Z.; Liu, D.; Tian, Y.; Deng, S.P.; Shang, X.L. Tree species composition influences enzyme activities and microbial biomass in the rhizosphere: A rhizobox approach. PLoS ONE 2013, 8, e61461. [Google Scholar] [CrossRef] [PubMed]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soil. Appl. Environ. Microbiol. 2003, 69, 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds MBC, DAZ and BOF, are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot | Leaf | Flower | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Height (cm) | Diameter (cm) | Fresh wt (g/plant) | Dry wt (g/plant) | Width (cm) | Length (cm) | SPAD Value (%) | Diameter (cm) | Ray Floret Number (No.) | |
| CK | 56.45 ± 4.01 d | 3.35 ± 0.32 d | 49.31 ± 4.75 d | 8.95 ± 0.89 d | 3.24 ± 0.31 d | 6.48 ± 0.55 d | 13.67 ± 1.73 d | 8.25 ± 0.46 d | 231.83 ± 17.34 f |
| MBC | 62.63 ± 3.32 c | 4.39 ± 0.41 c | 56.61 ± 4.57 c | 9.97 ± 0.73 c | 4.01 ± 0.24 c | 7.46 ± 0.46 c | 14.92 ± 1.66 c | 9.32 ± 0.52 c | 249.20 ± 18.56 e |
| DAZ | 63.01 ± 3.49 c | 4.91 ± 0.34 c | 57.05 ± 5.52 c | 10.16 ± 0.87 c | 4.28 ± 0.27 c | 7.51 ± 0.38 c | 14.53 ± 1.62 c | 9.39 ± 0.43 c | 259.55 ± 14.39 d |
| BOF | 69.83 ± 4.18 b | 5.92 ± 0.37 b | 68.79 ± 4.28 b | 14.23 ± 0.83 b | 5.43 ± 0.29 b | 8.32 ± 0.42 b | 17.35 ± 1.71 b | 10.50 ± 0.51 b | 261.89 ± 17.77 c |
| MBC + BOF | 69.98 ± 4.41 b | 5.99 ± 0.35 b | 69.08 ± 4.98 b | 14.76 ± 0.64 b | 5.27 ± 0.31 b | 8.45 ± 0.34 b | 17.07 ± 1.63 b | 10.43 ± 0.49 b | 269.34 ± 21.97 b |
| DAZ + BOF | 72.01 ± 3.92 a | 6.96 ± 0.33 a | 72.92 ± 4.82 a | 16.17 ± 0.91 a | 6.34 ± 0.25 a | 9.46 ± 0.36 a | 18.39 ± 1.92 a | 11.49 ± 0.40 a | 271.49 ± 19.72 a |
| Treatment | Bacteria (104 cfu·g−1 soil) | Fungi (104 cfu·g−1 soil) | Bacteria/Fungi Ratio |
|---|---|---|---|
| CK | 89.6 ± 0.59 c | 8.66 ± 0.39 a | 10.3 ± 0.9 d |
| MBC | 92.8 ± 0.48 c | 7.34 ± 0.37 bc | 13.7 ± 1.0 c |
| DAZ | 91.7 ± 0.68 c | 4.94 ± 0.22 d | 18.6 ± 1.5 b |
| MBC + BOF | 145.0 ± 0.72 b | 6.78 ± 0.39 b | 19.8 ± 1.2 b |
| DAZ + BOF | 179.0 ± 0.73 a | 5.86 ± 0.38 c | 30.5 ± 1.1 a |
| BOF | 140.0 ± 0.82 b | 6.47 ± 0.42 bc | 21.6 ± 1.2 b |
| Treatments | Bacteria | Fungi | ||||
|---|---|---|---|---|---|---|
| n | H′ | J | n | H′ | J | |
| CK | 25 | 3.10 ± 0.30 a | 0.96 ± 0.02 a | 24 | 3.00 ± 0.01 a | 0.66 ± 0.07 d |
| MBC | 32 | 3.15 ± 0.33 a | 0.98 ± 0.09 a | 5 | 1.24 ± 0.10 f | 0.77 ± 0.06 c |
| DAZ | 24 | 3.12 ± 0.23 a | 0.98 ± 0.04 a | 16 | 1.65 ± 0.12 b | 0.81 ± 0.02 b |
| MBC+BOF | 25 | 3.14 ± 0.37 a | 0.97 ± 0.03 a | 7 | 1.81 ± 0.18 e | 0.89 ± 0.00 b |
| DAZ+BOF | 26 | 3.19 ± 0.11 a | 0.97 ± 0.02 a | 8 | 1.99 ± 0.20 d | 0.96 ± 0.01 a |
| BOF | 35 | 3.23 ± 0.22 a | 0.96 ± 0.11 a | 14 | 2.57 ± 0.11 c | 0.98 ± 0.04 a |
| DGGE Band | Closest Relatives Microorganisms (Phylogenic Affiliations) | Similarity (%) | Genebank Accession No. |
|---|---|---|---|
| 1 | Uncultured Xanthomonadaceae bacterium | 96 | FJ53688611 |
| 2 | Uncultured bacterium | 97 | EU362858.1 |
| 3 | Uncultured Ohtaekwangia sp. | 97 | JX493344.1 |
| 4 | Uncultured Bacillus sp. | 99 | HQ1791481 |
| 5 | Alpha proteobacterium | 96 | JQ608334.2 |
| 6 | Uncultured Bradyrhizobium sp. | 95 | HE654679.1 |
| 7 | Uncultured Xanthomonadaceae bacterium | 97 | FJ88933811 |
| 8 | Agricultural soil bacterium | 98 | HQ132702.1 |
| 9 | Uncultured beta proteobacterium | 98 | AJ318162.1 |
| 10 | Uncultured methylovirgula sp. | 99 | KC297188.1 |
| 11 | Alpha proteobacterium | 97 | AB470422.1 |
| 12 | Rhizobium sp. | 95 | HG423545.1 |
| 13 | Variovorax sp. | 97 | HM484318.1 |
| 14 | Microbacterium sp. | 98 | AJ318162.1 |
| 15 | Sphingomonas sp. | 98 | JQ608334.2 |
| 16 | Uncultured bacterium | 97 | FJ796671.1 |
| 17 | Anoxybacillus flavithermus | 98 | KF279366.1 |
| 18 | Sphingobium yanoikuyae | 98 | GQ214010.1 |
| 19 | Uncultured Chloroflexi bacterium | 94 | HM164420.1 |
| 20 | Flavobacterium sp. | 100 | KF891387.1 |
| 21 | Uncultured bacterium | 96 | JQ890611.1 |
| 22 | Dechlorospirillum | 98 | GU167977.1 |
| 23 | Uncultured Flexibacter sp. | 96 | GU201555.1 |
| 24 | Ochrobactrum sp. | 97 | HQ659714.1 |
| 25 | Agricultural soil bacterium | 95 | KC193578.1 |
| 26 | Uncultured bacterium | 99 | EU419388.1 |
| 27 | Mycobacterium sp. | 97 | DQ658940.1 |
| 28 | Uncultured Methylocystis sp. | 97 | GU227561.1 |
| 29 | Mycobacterium gilvum | 97 | AJ699170.3 |
| 30 | Clostridium neonatale | 96 | GU227558.1 |
| DGGE Band | Closest Relatives Microorganisms (Phylogenic Affiliations) | Similarity (%) | Genebank Accession No. |
|---|---|---|---|
| 1 | Aspergillus terreus | 98 | JN639854.1 |
| 2 | Leptosphaeria maculans | 98 | NW003533867.1 |
| 3 | Fusarium sp. | 97 | EU381149.1 |
| 4 | Ophiobolus herpotrichus | 100 | U43453.1 |
| 5 | Psilocybe silvatica | 97 | DQ851583.1 |
| 6 | Alnicola sp. | 98 | JN939094.1 |
| 7 | Uncultured Clitopilus | 96 | GQ995701.1 |
| 8 | Mythicomyces corneipes | 98 | DQ092917.1 |
| 9 | Arthrobotrys oligospora | 98 | JQ809337.1 |
| 10 | Aspergillus ochraceus | 97 | AB002068.1 |
| 11 | Cucurbitaria sp. | 99 | FJ215704.1 |
| 12 | Phoma macrostoma | 97 | AB454217.1 |
| 13 | Phoma sp. | 97 | AY293772.1 |
| 14 | Leptosphaeria microscopica | 99 | U04235.1 |
| 15 | Gondwanamyces proteae | 97 | AY271804.1 |
| 16 | Penicillium sp. | 97 | JF950269.1 |
| 17 | Monodictys arctica | 97 | EU686519.1 |
| 18 | Graphium putredinis | 99 | AB007683.1 |
| 19 | Saccobolus dilutellus | 99 | FJ176814.1 |
| 20 | Orbilia auricolor | 99 | DQ471001.1 |
| 21 | Uncultured soil fungi | 97 | AJ877196 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Chen, X.; Deng, S.; Dong, X.; Song, A.; Yao, J.; Fang, W.; Chen, F. The Effects of Fungicide, Soil Fumigant, Bio-Organic Fertilizer and Their Combined Application on Chrysanthemum Fusarium Wilt Controlling, Soil Enzyme Activities and Microbial Properties. Molecules 2016, 21, 526. https://doi.org/10.3390/molecules21040526
Zhao S, Chen X, Deng S, Dong X, Song A, Yao J, Fang W, Chen F. The Effects of Fungicide, Soil Fumigant, Bio-Organic Fertilizer and Their Combined Application on Chrysanthemum Fusarium Wilt Controlling, Soil Enzyme Activities and Microbial Properties. Molecules. 2016; 21(4):526. https://doi.org/10.3390/molecules21040526
Chicago/Turabian StyleZhao, Shuang, Xi Chen, Shiping Deng, Xuena Dong, Aiping Song, Jianjun Yao, Weimin Fang, and Fadi Chen. 2016. "The Effects of Fungicide, Soil Fumigant, Bio-Organic Fertilizer and Their Combined Application on Chrysanthemum Fusarium Wilt Controlling, Soil Enzyme Activities and Microbial Properties" Molecules 21, no. 4: 526. https://doi.org/10.3390/molecules21040526
APA StyleZhao, S., Chen, X., Deng, S., Dong, X., Song, A., Yao, J., Fang, W., & Chen, F. (2016). The Effects of Fungicide, Soil Fumigant, Bio-Organic Fertilizer and Their Combined Application on Chrysanthemum Fusarium Wilt Controlling, Soil Enzyme Activities and Microbial Properties. Molecules, 21(4), 526. https://doi.org/10.3390/molecules21040526