
Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
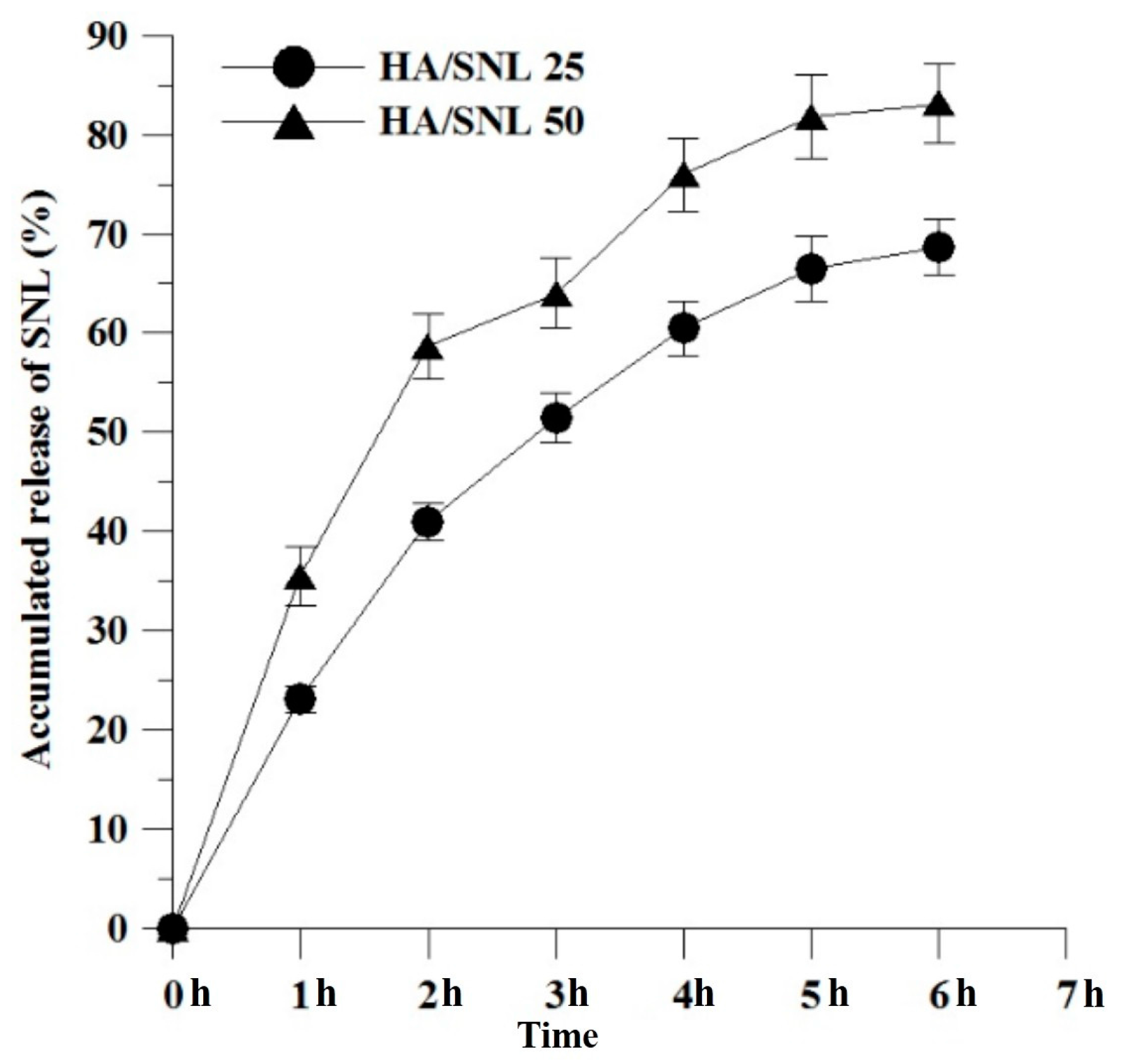
2.1. Characteristics of HA/SNL Aggregates
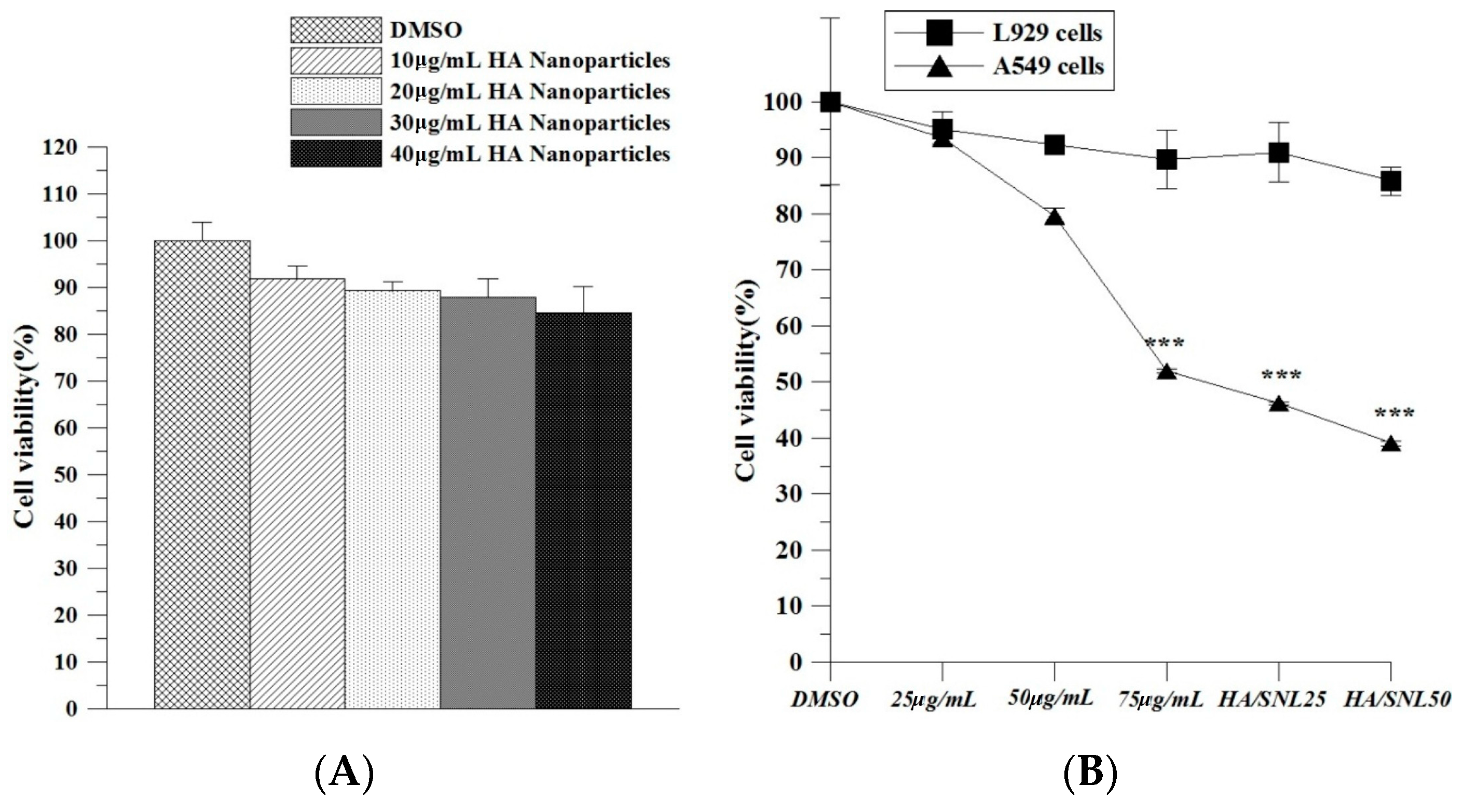
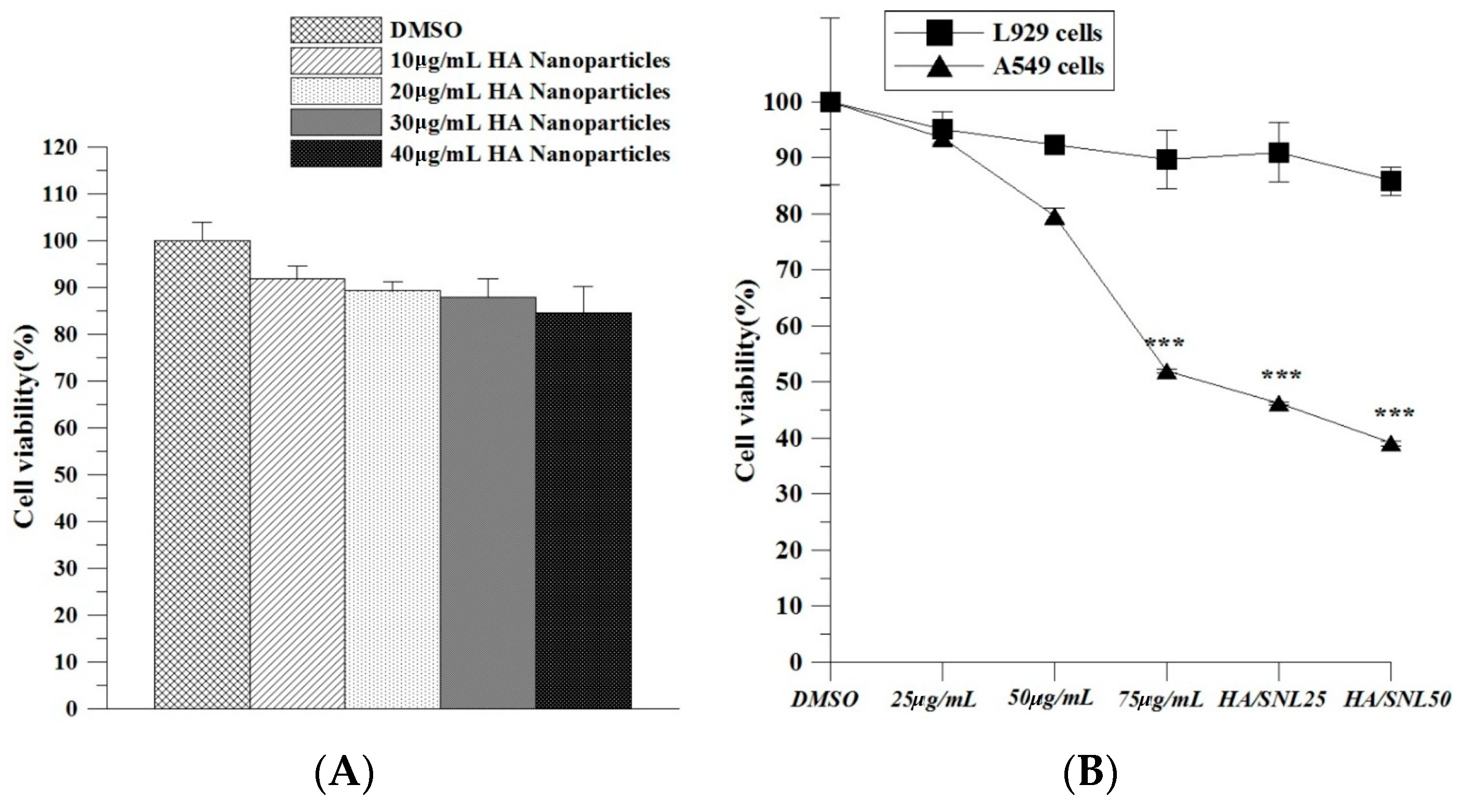
2.2. Hyaluronan Nanoparticle Cytotoxicity Assay
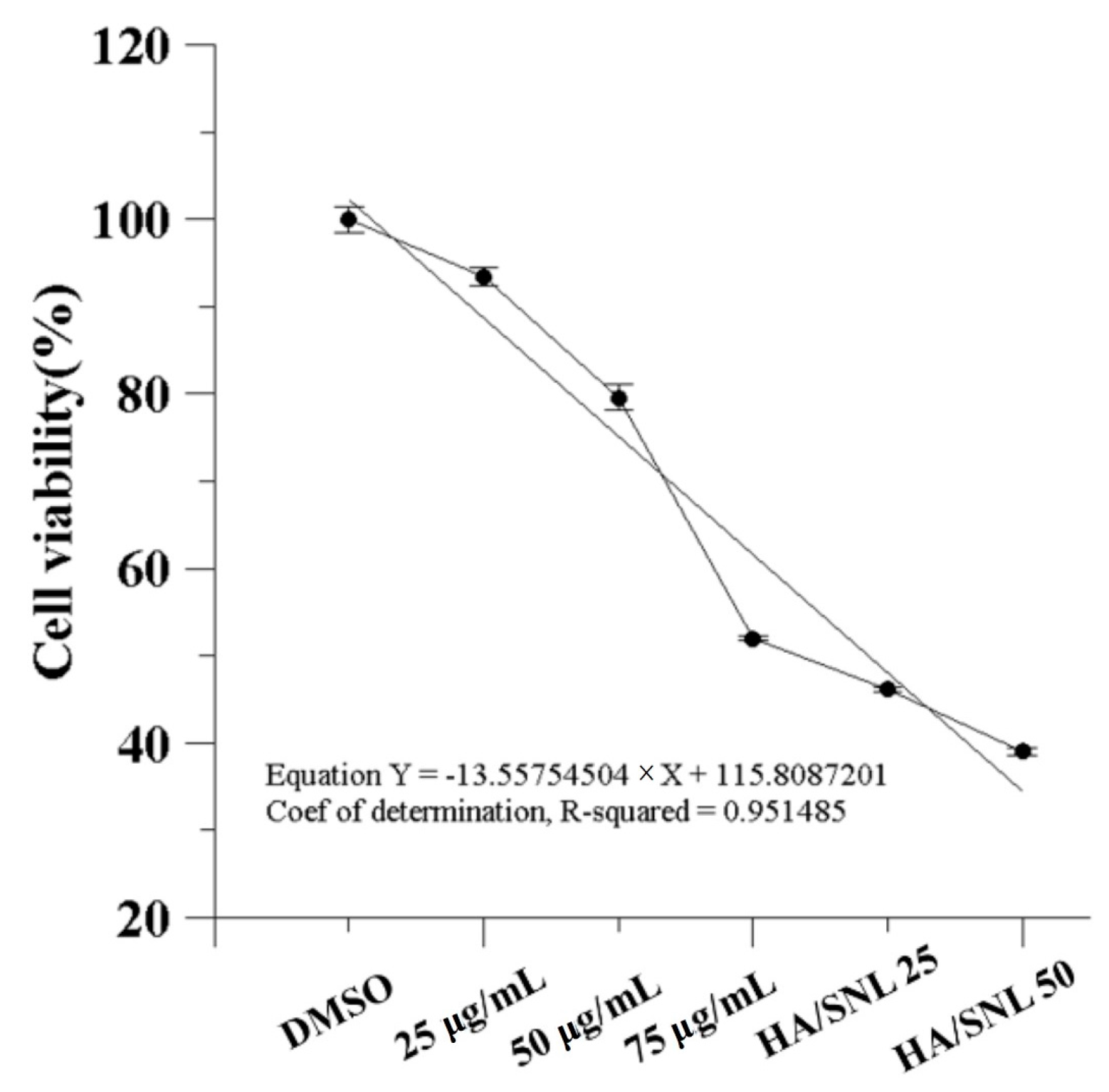
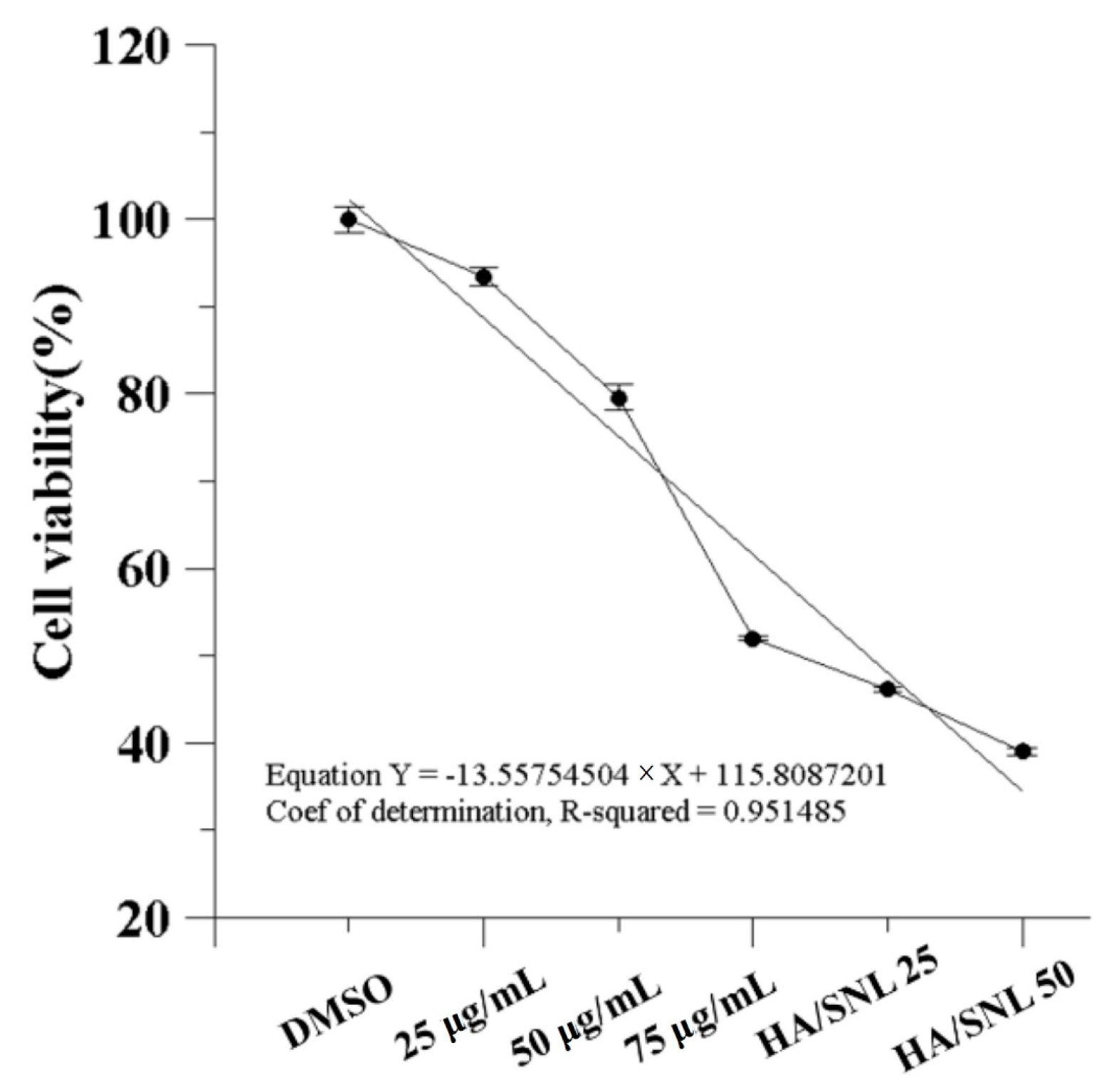
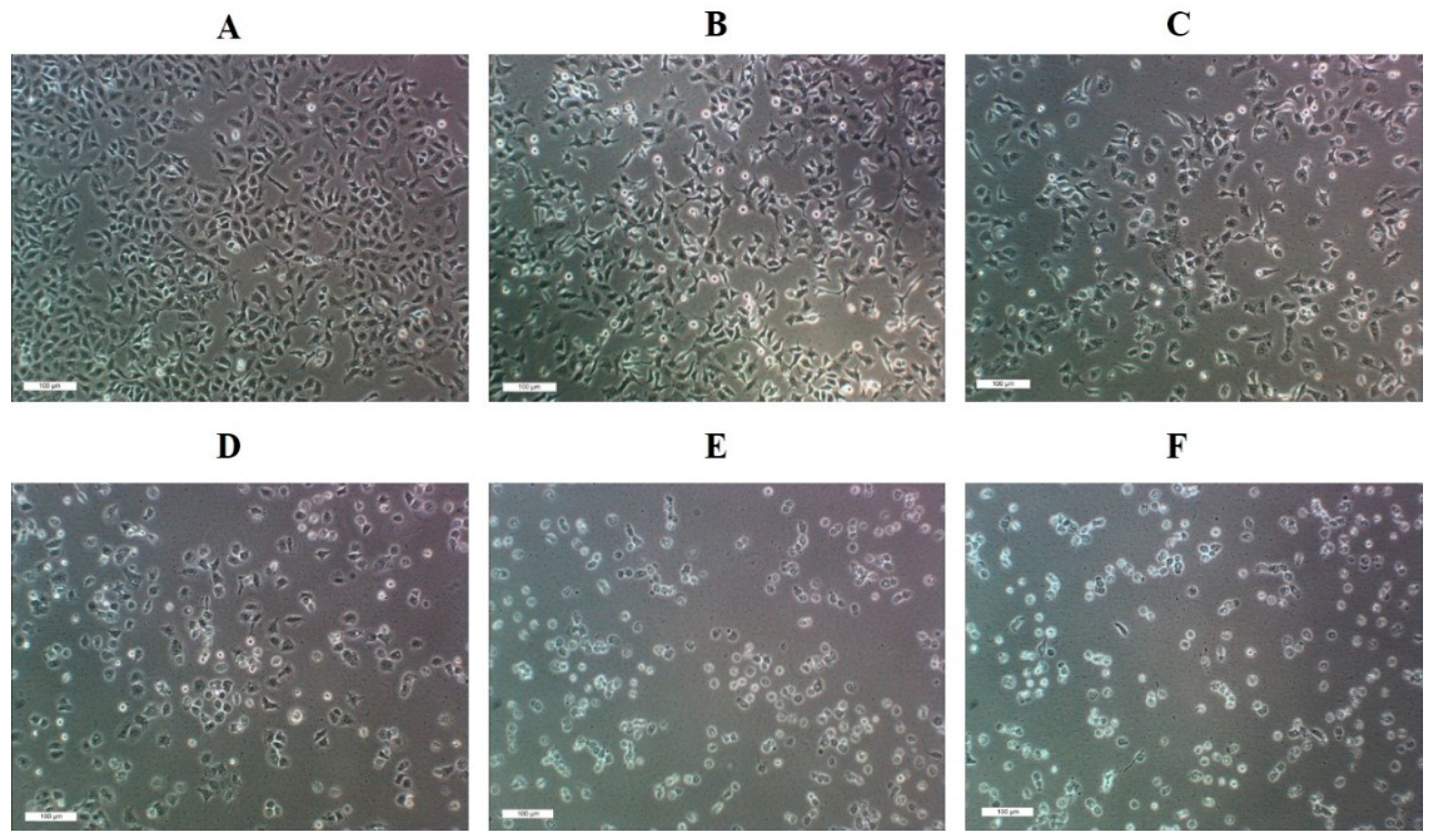
2.3. IC50 of SNL and in Vitro Cell Viability
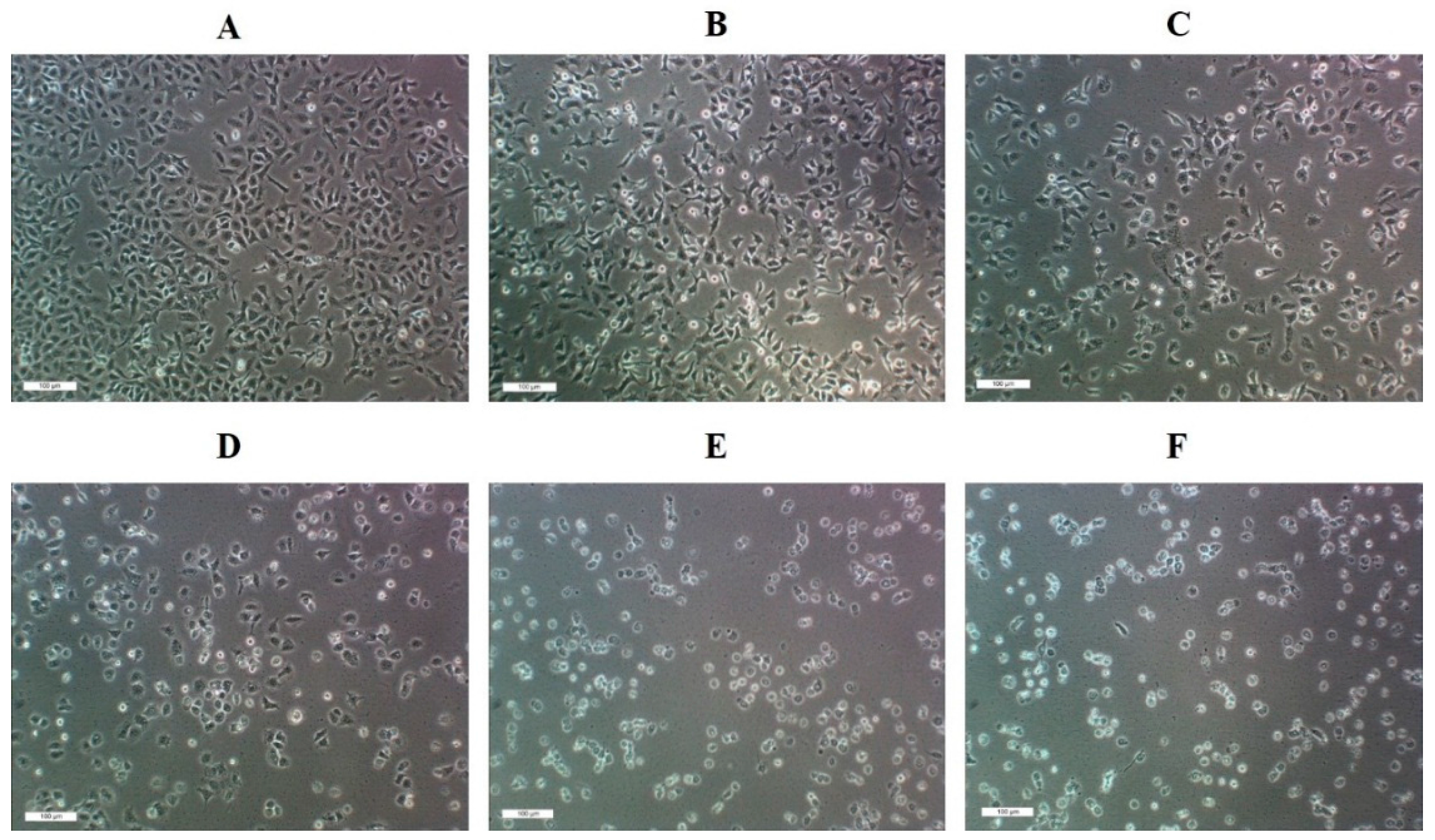
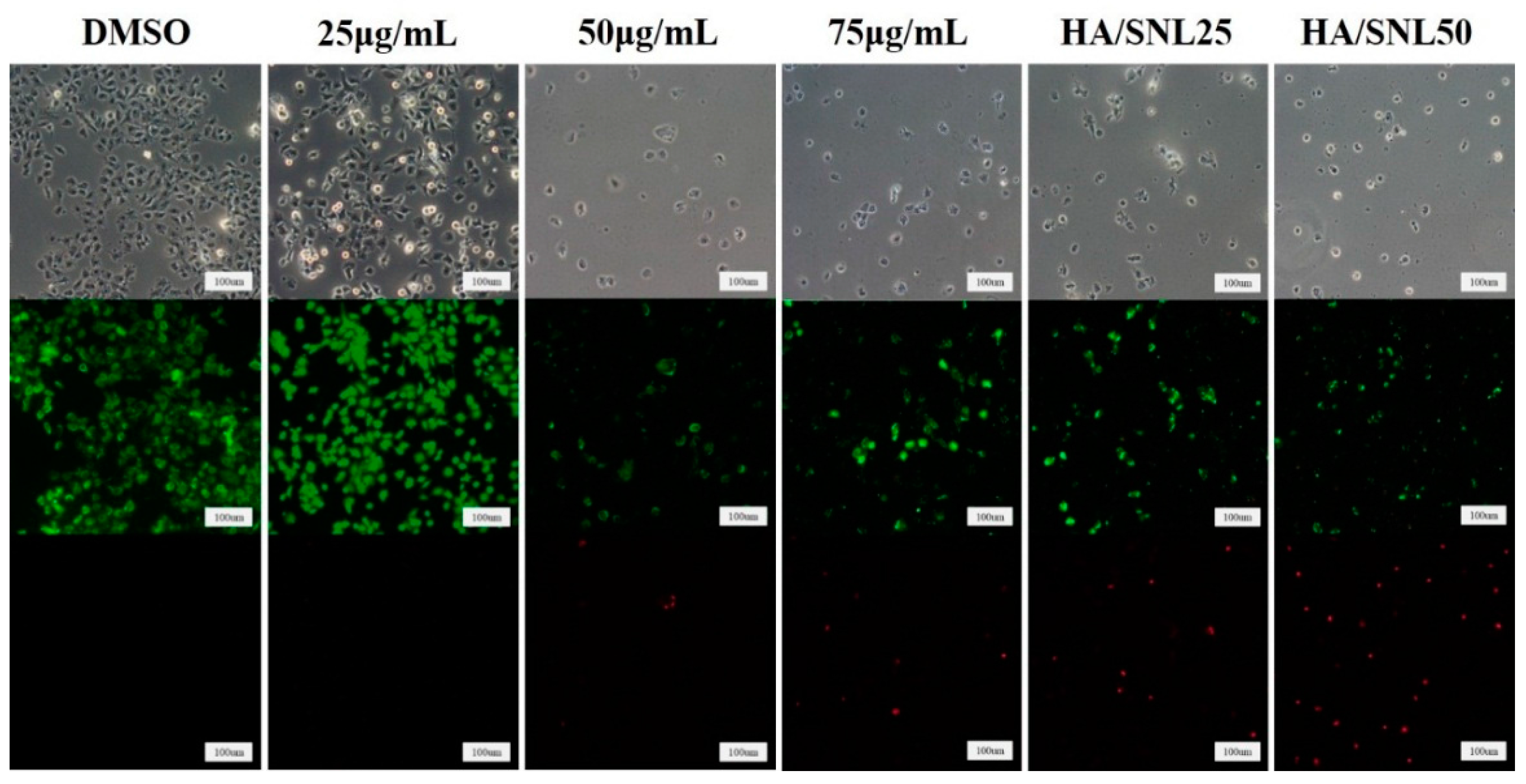
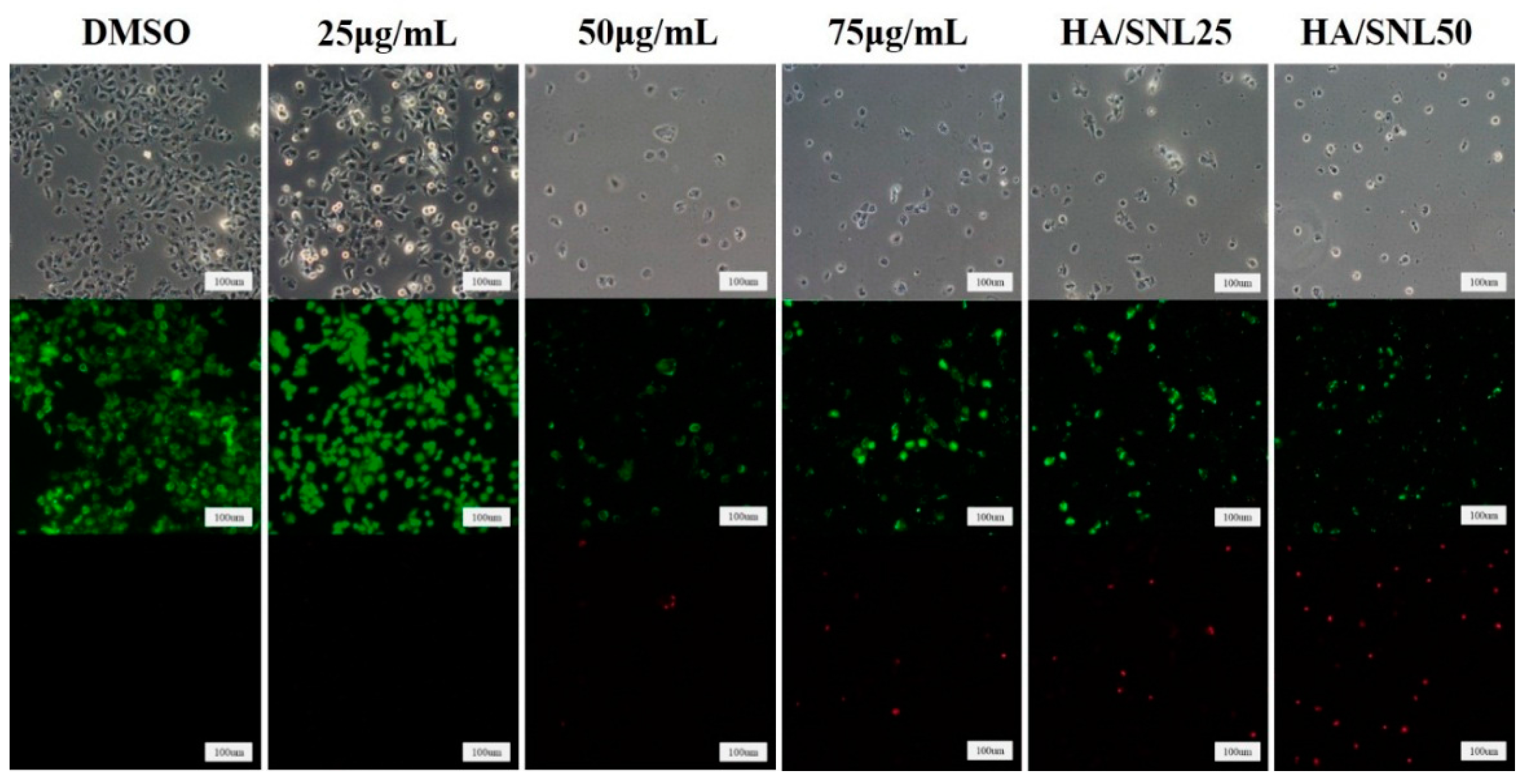
2.4. Live/dead Assay of A549 Cells
2.5. Antimigratory Effect of SNL and HA/SNL Aggregates on A549 Cells
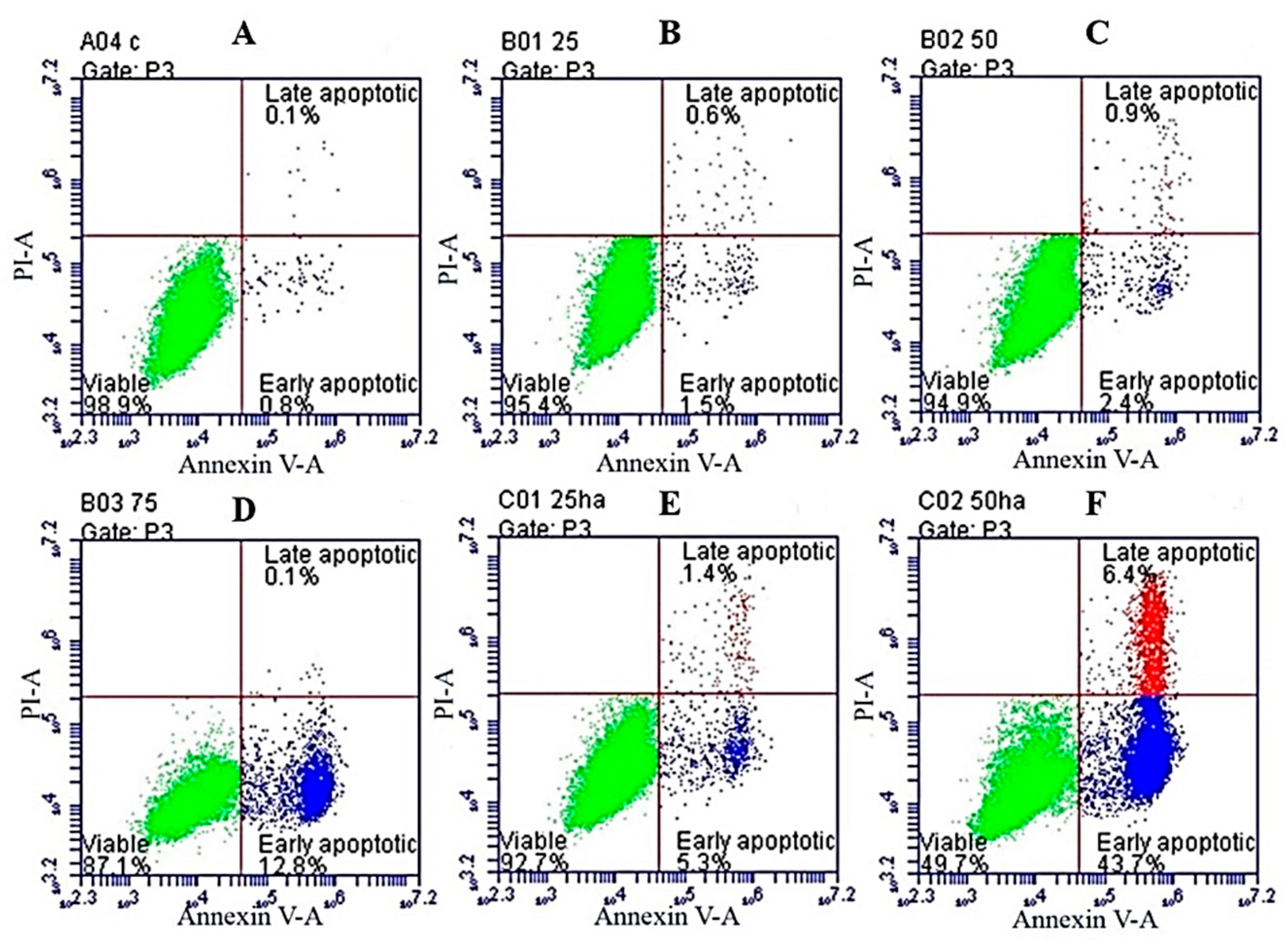
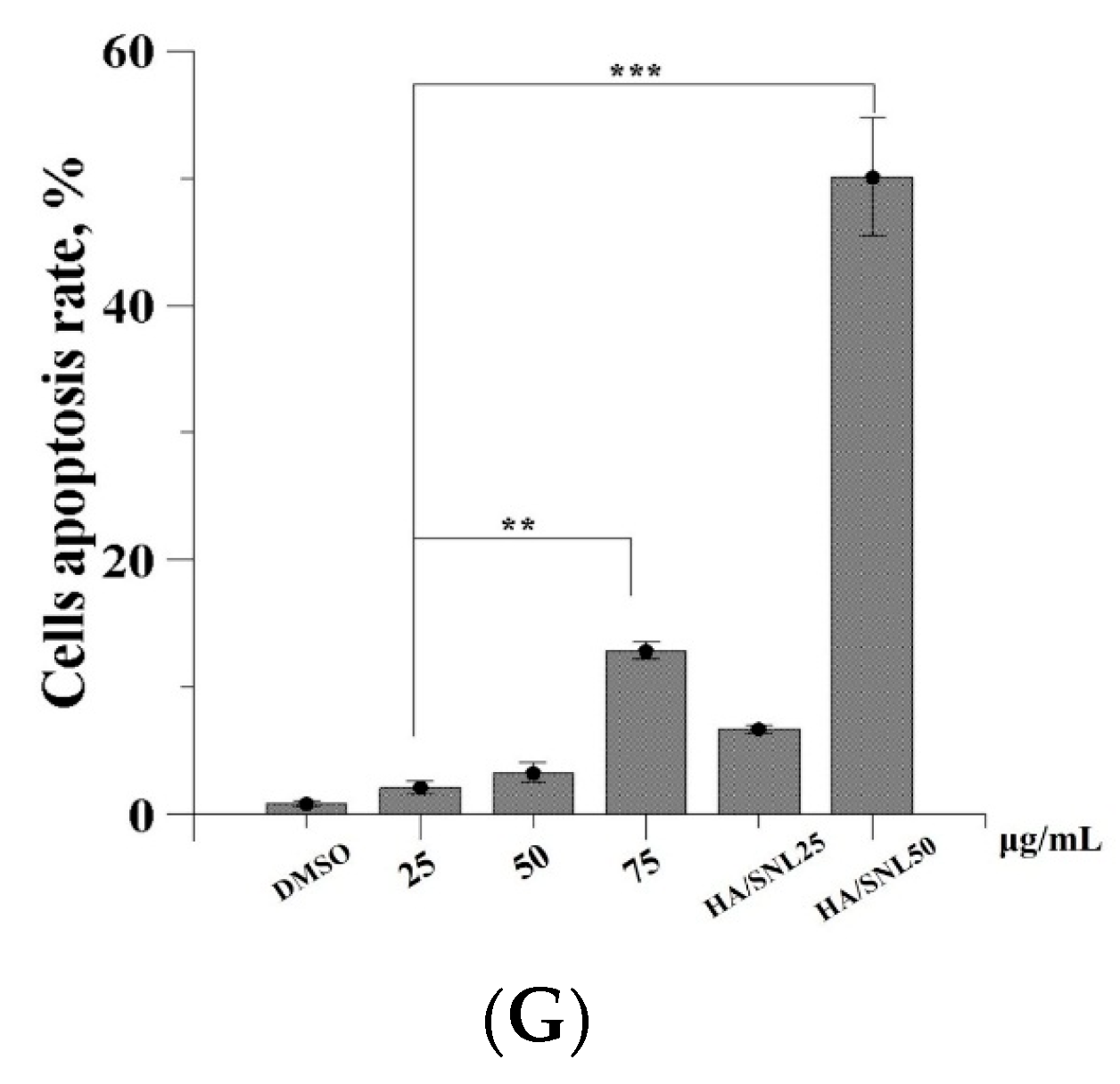
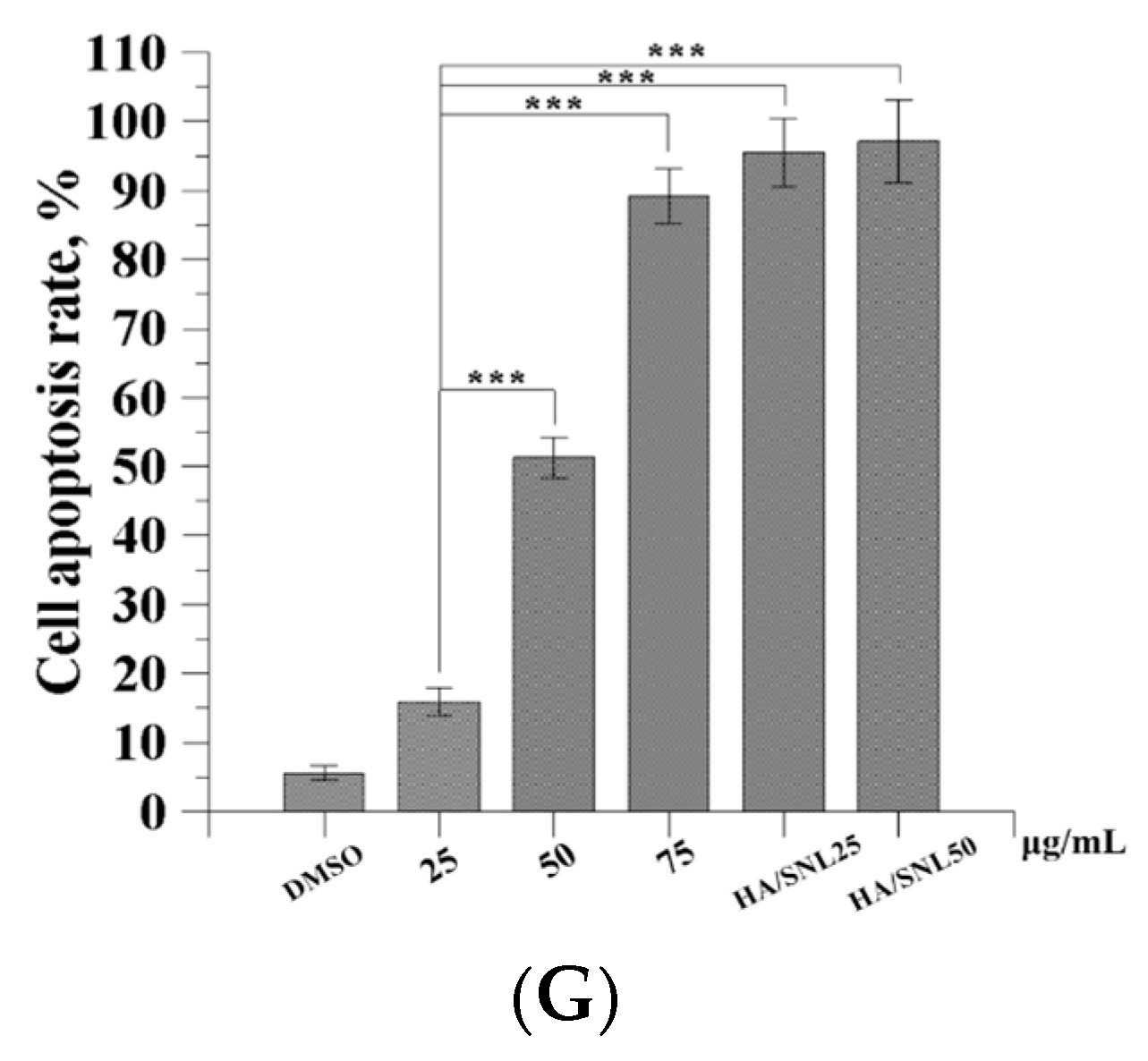
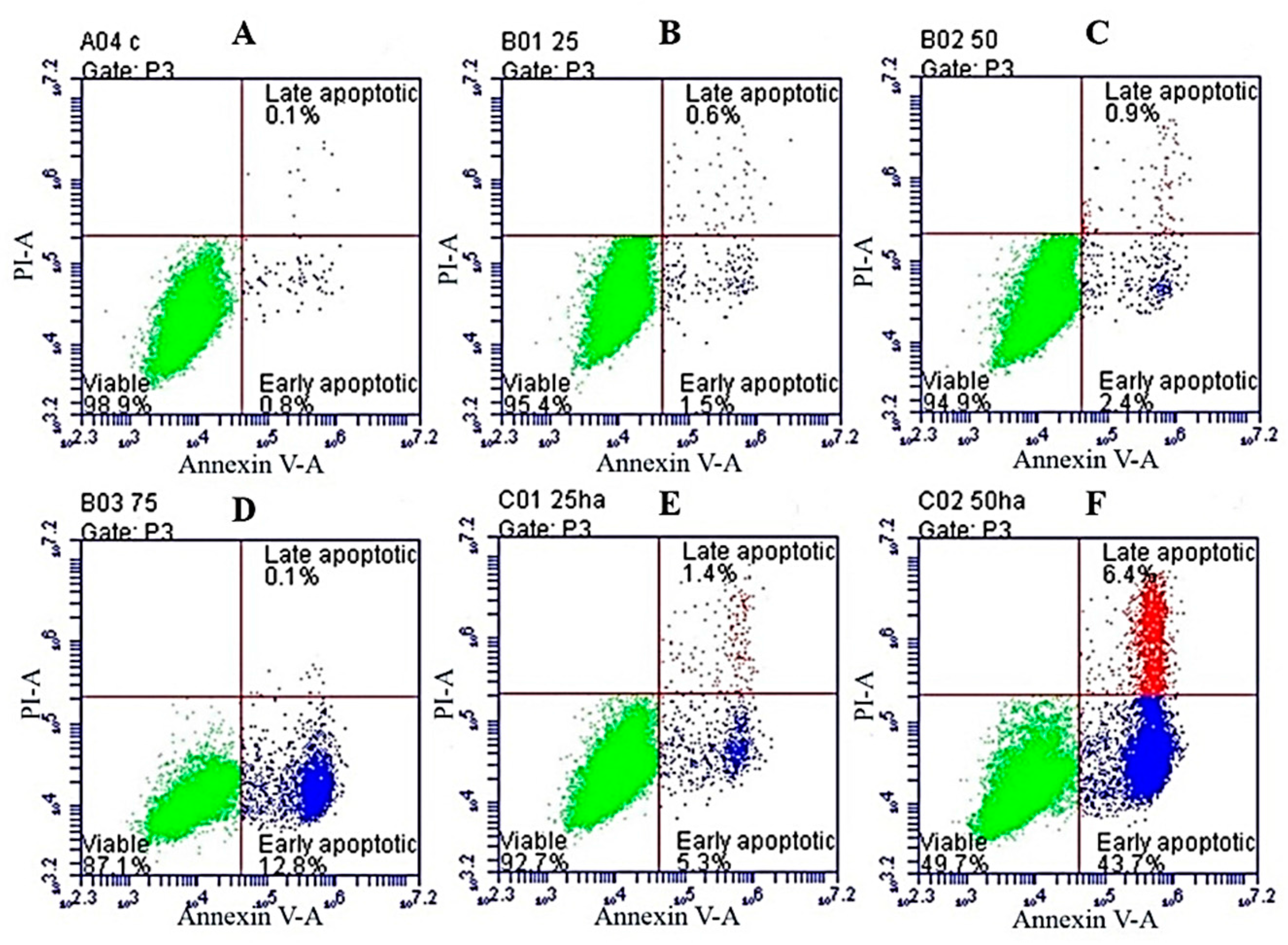
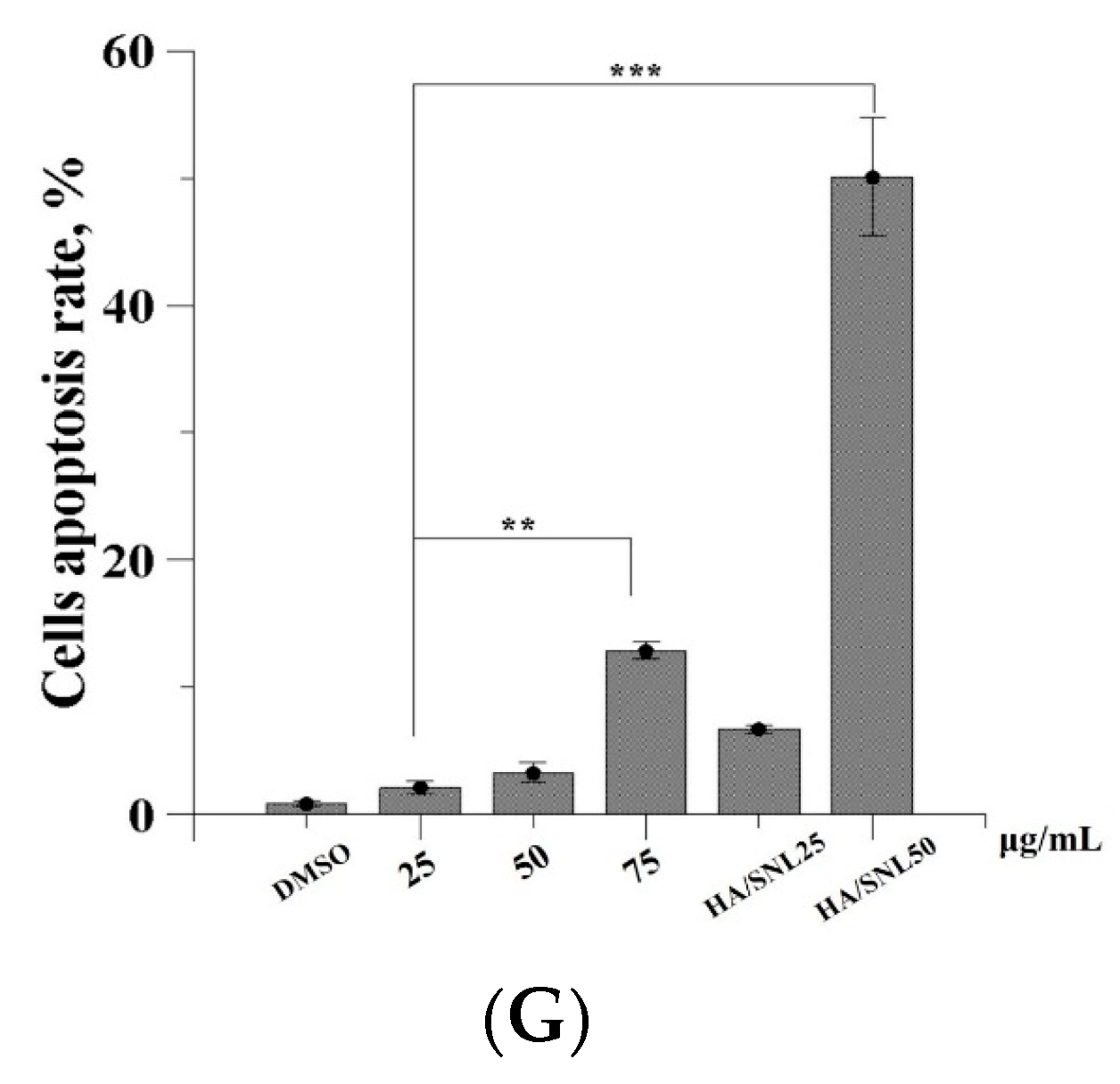
2.6. Apoptosis Assay through Flow Cytometry
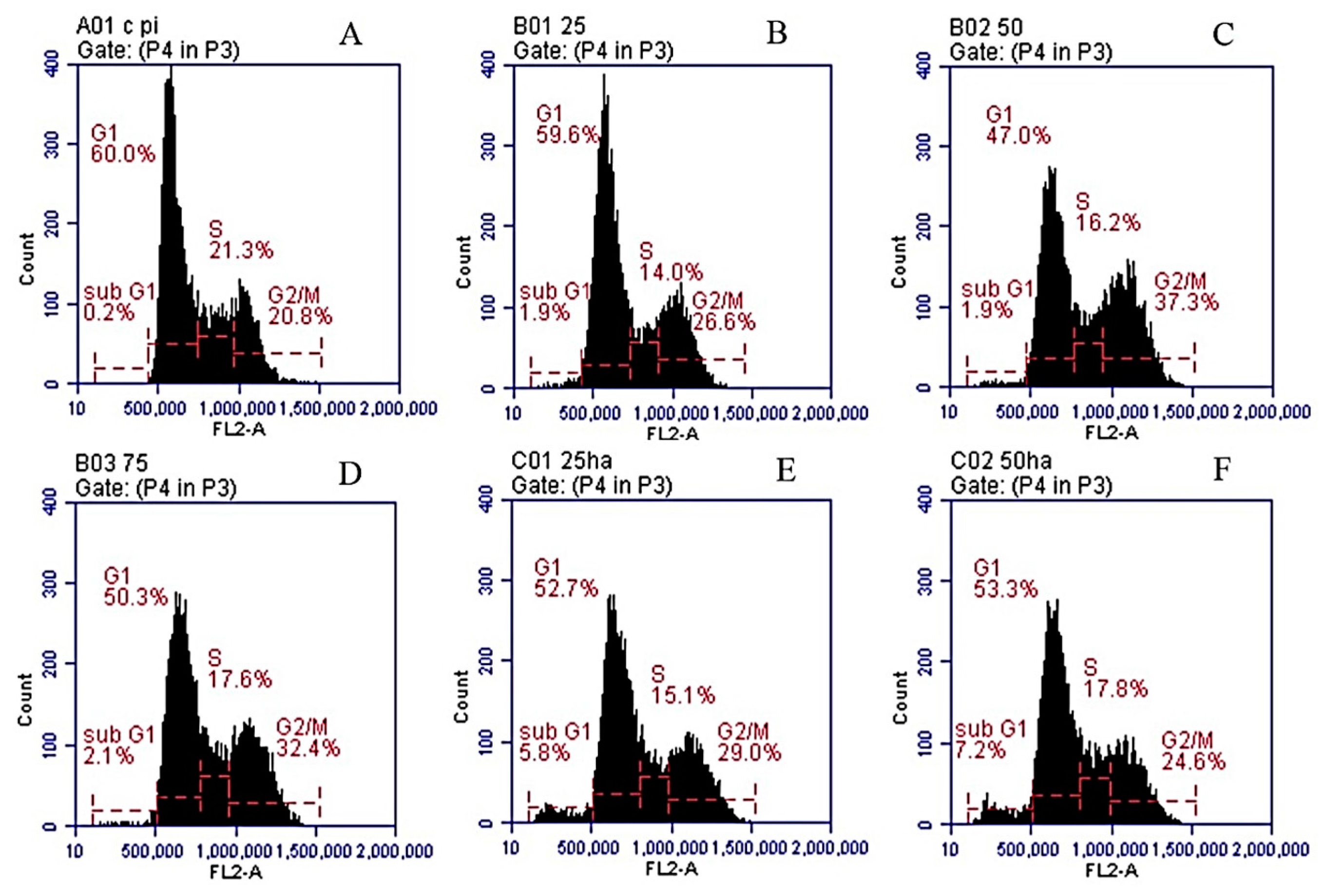
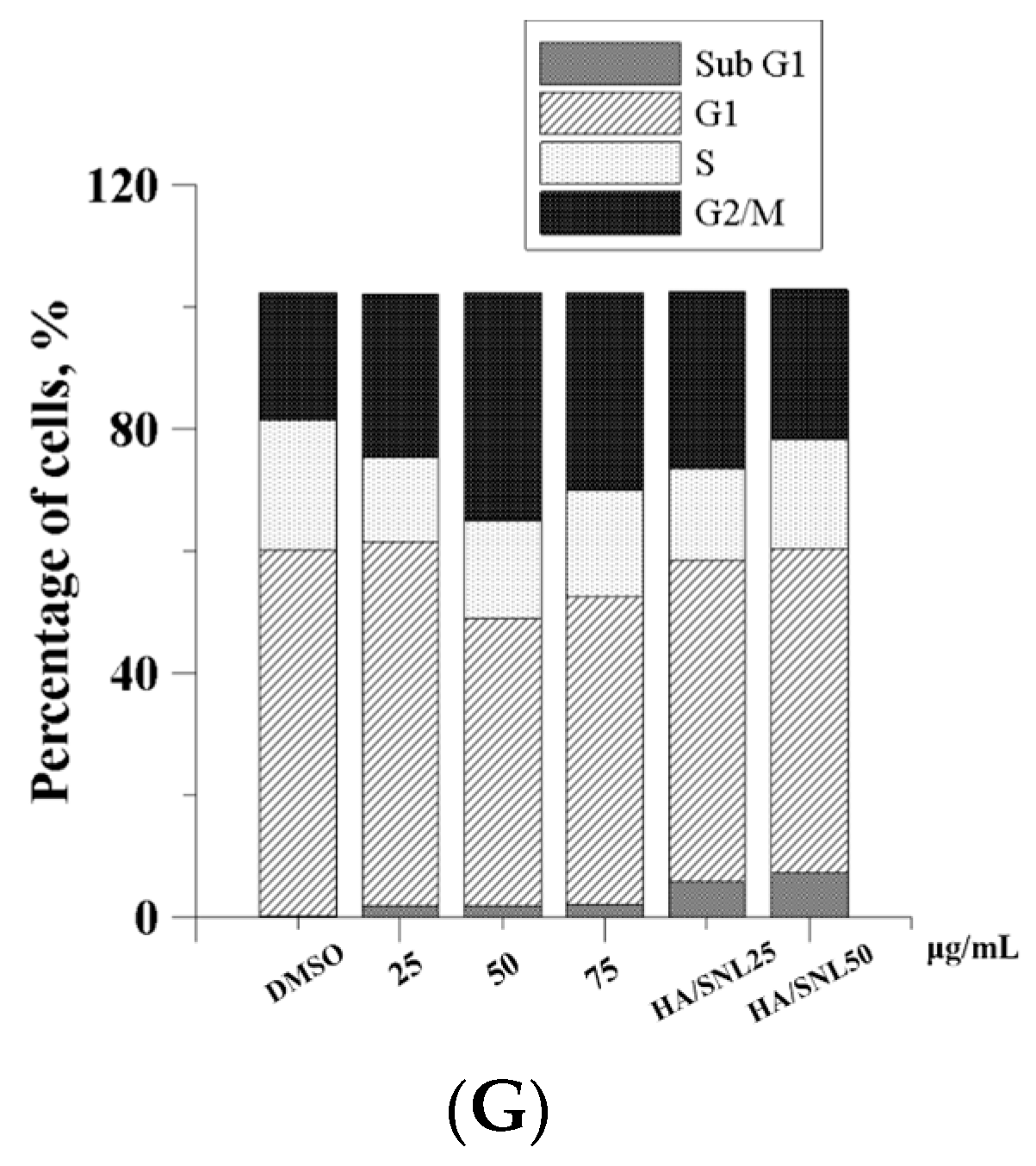
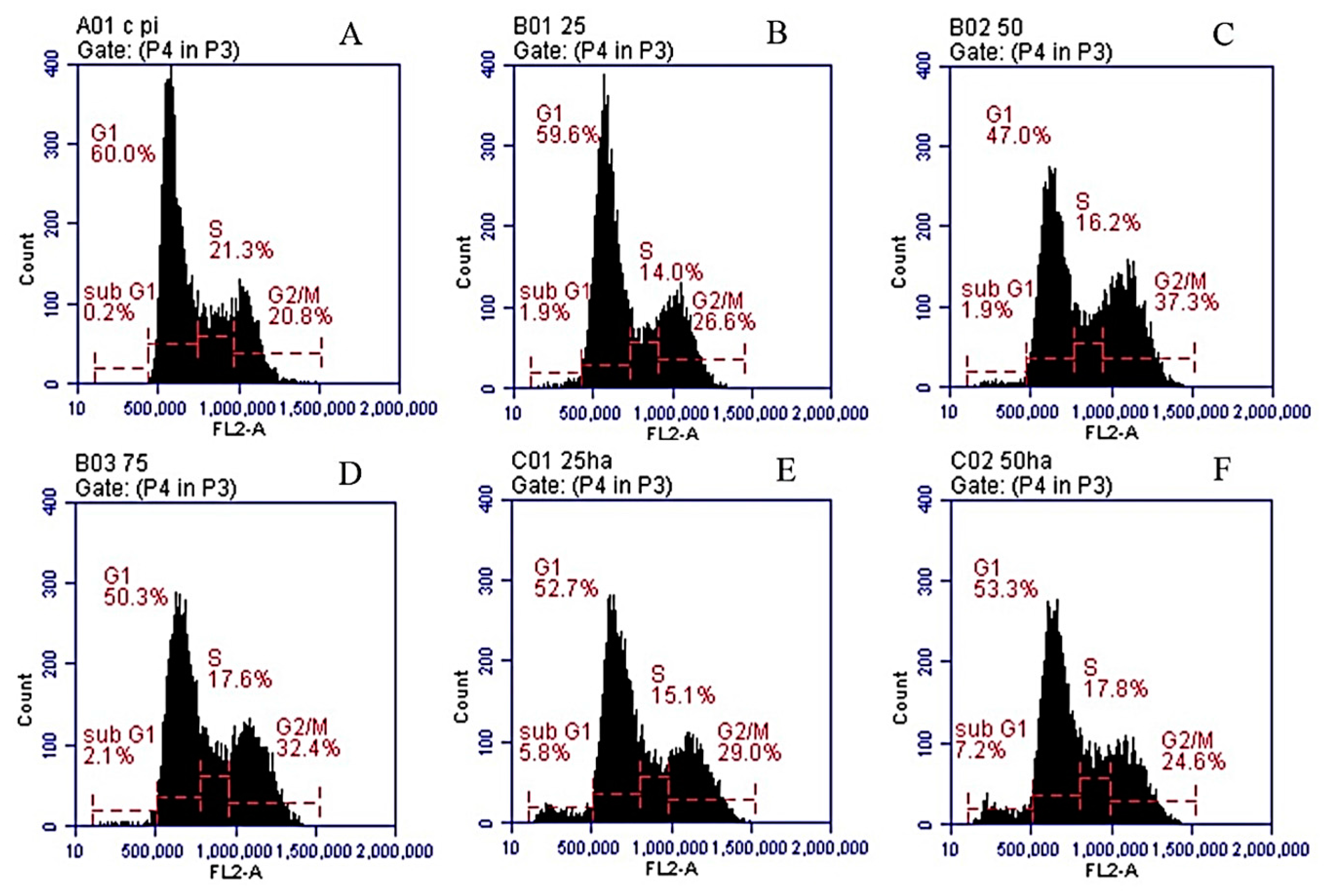
2.7. Cell Cycle Analysis
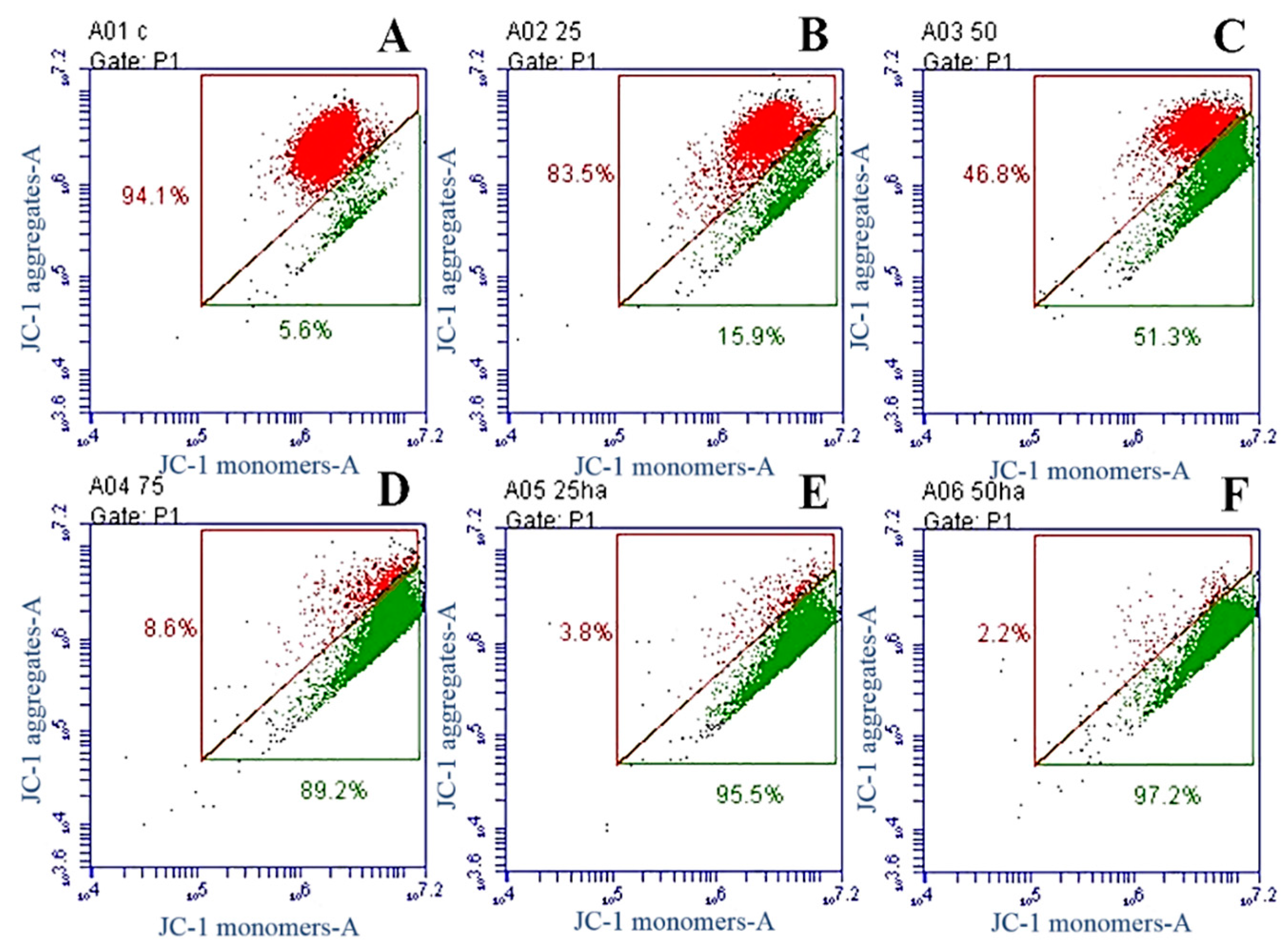
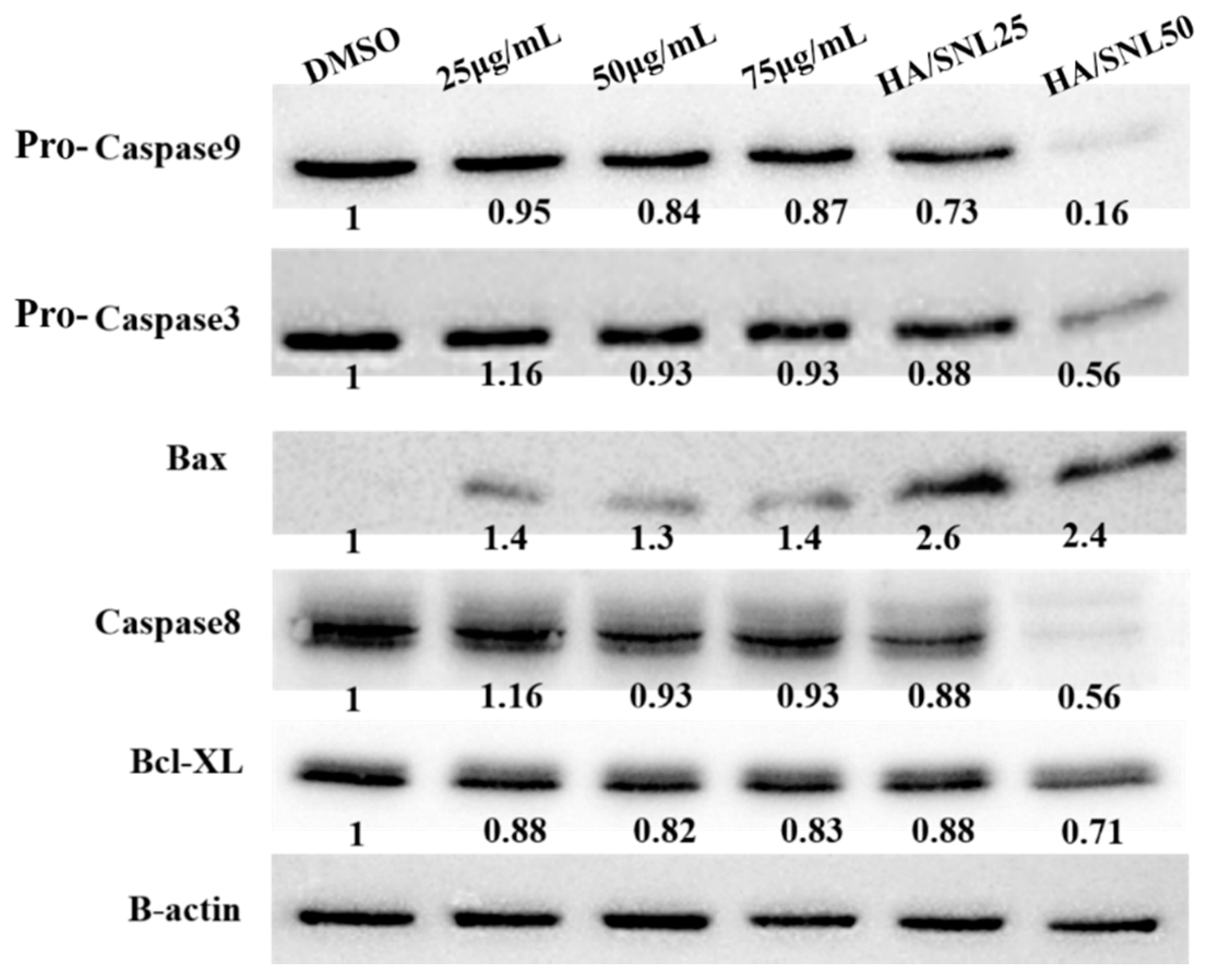
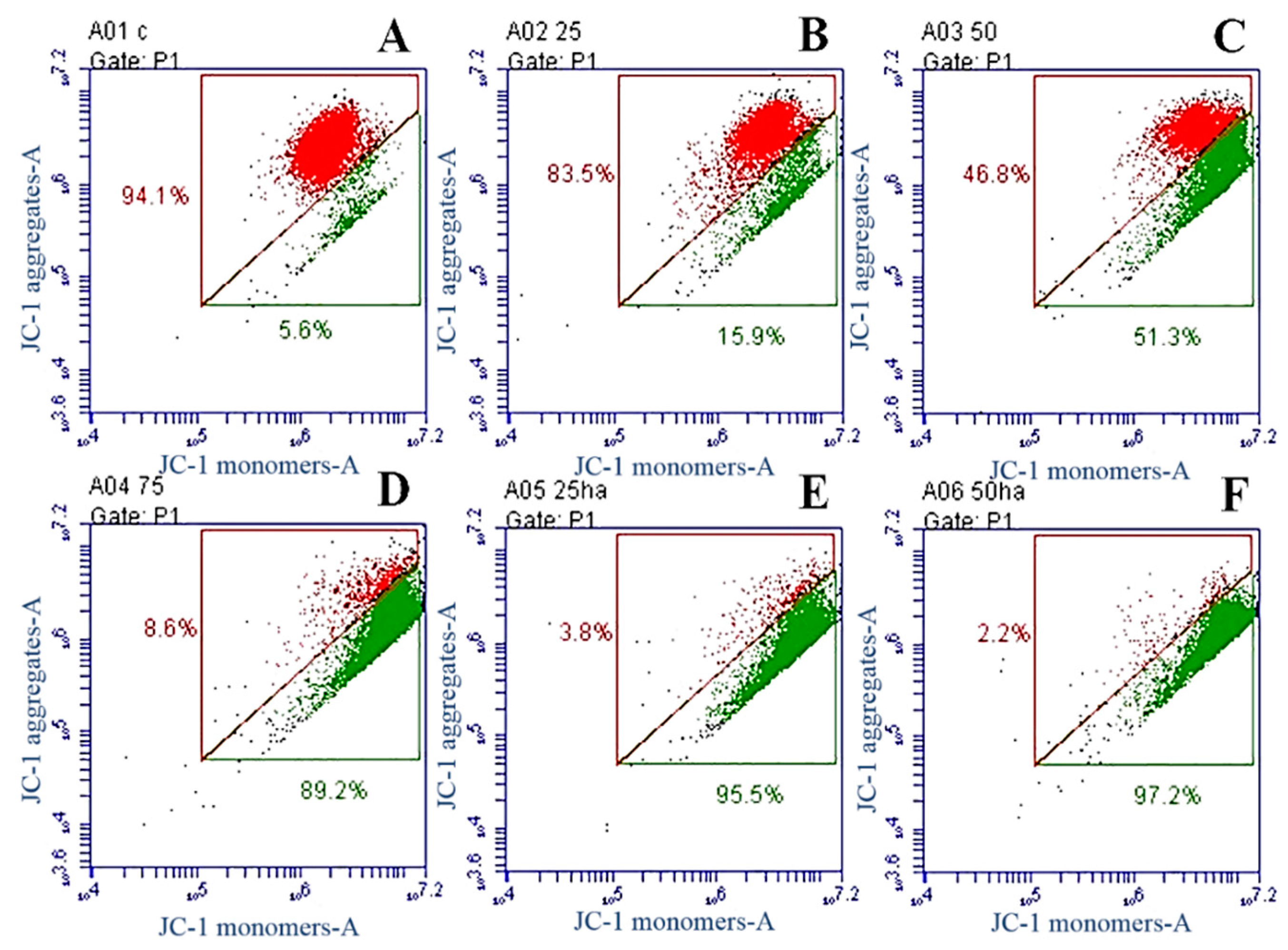
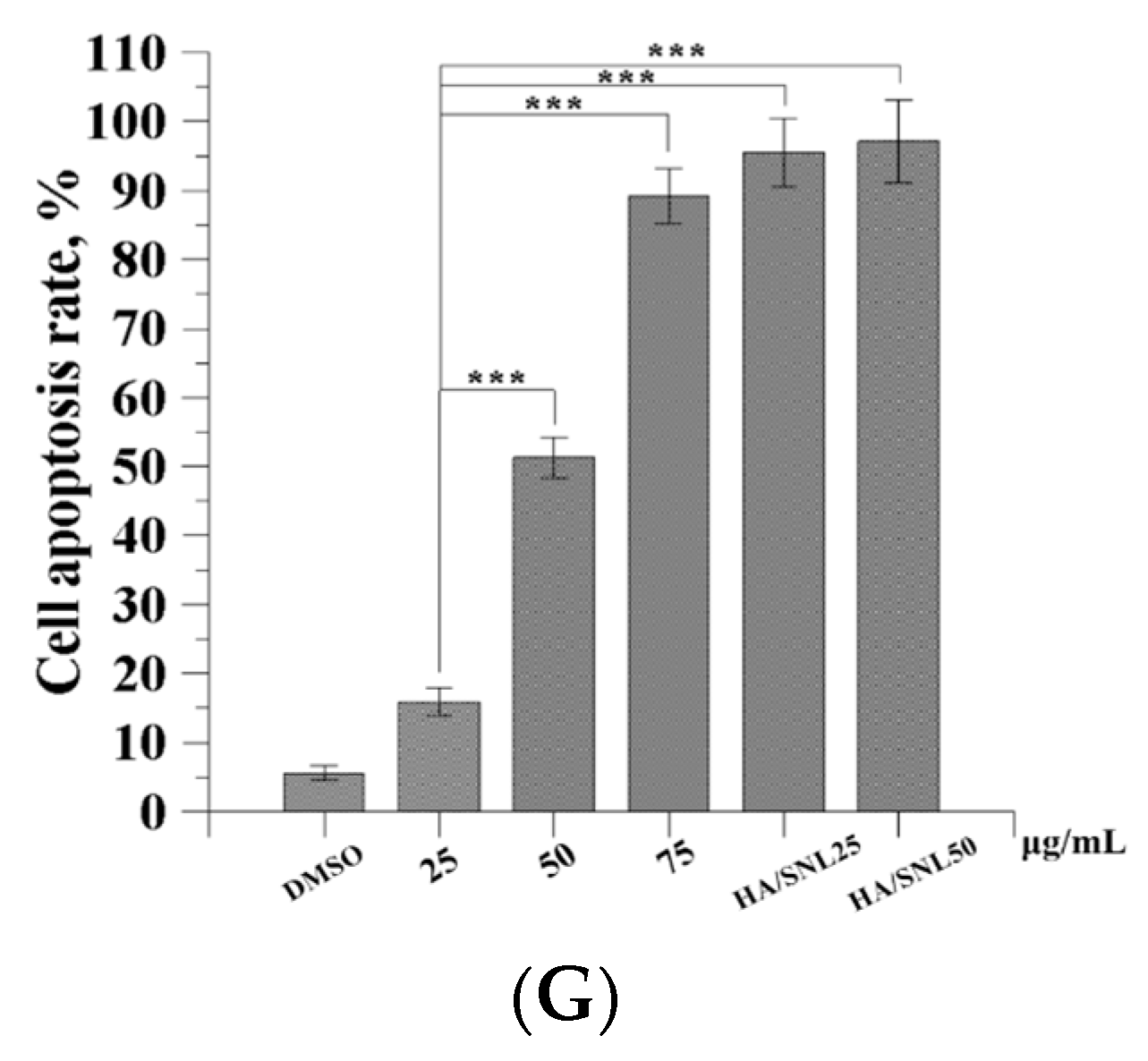
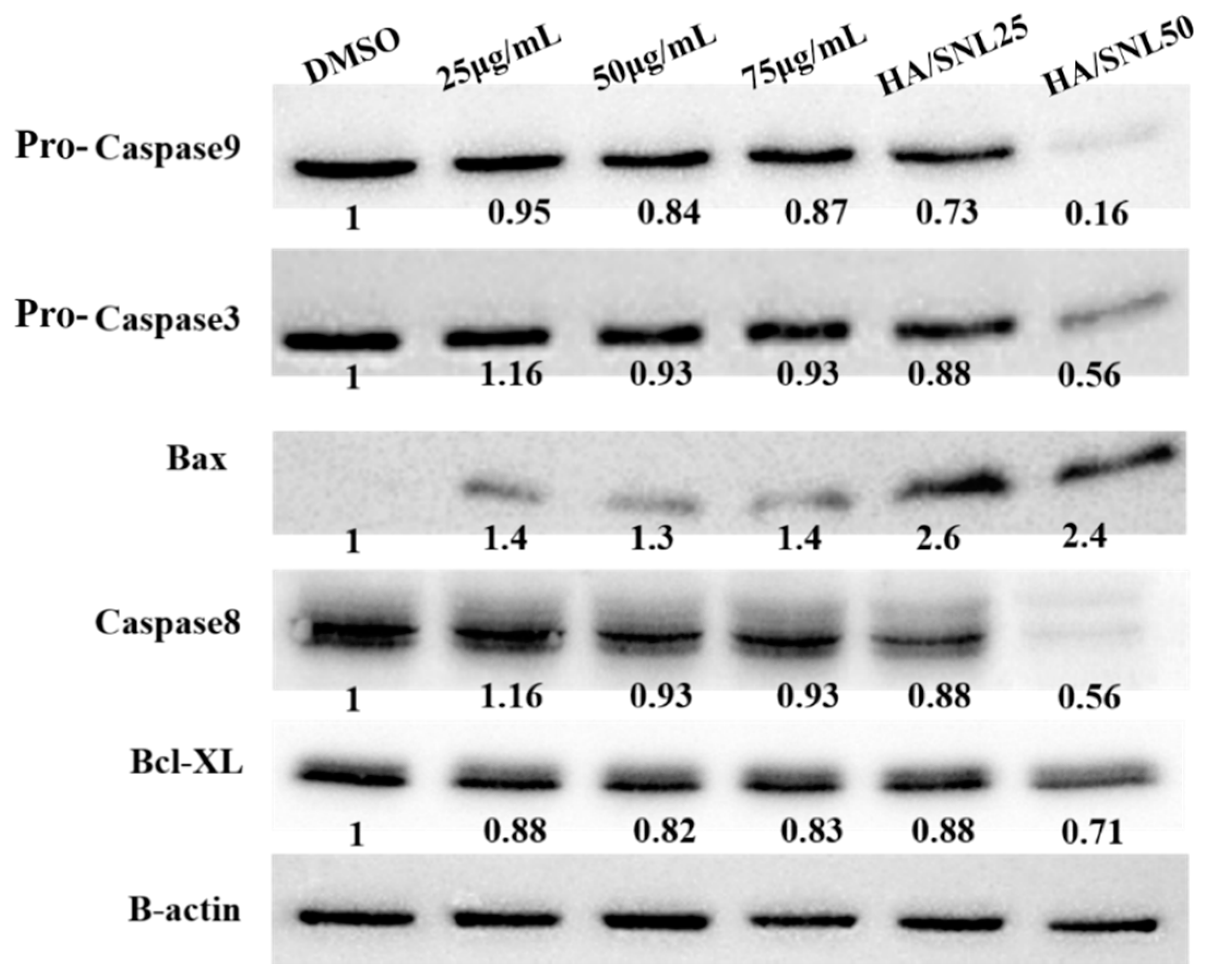
2.8. Western Blot Analysis
3. Experimental Section
3.1. Production of Hyaluronan/Sinulariolide Aggregates
3.2. SNL Incorporation Efficiency and in Vitro Release Study
3.3. Cell Culture and HA/SNL Aggregates Treatment
3.4. Flow Cytometric Analysis of Apoptosis
3.5. Cell Cycle Analysis
3.6. Western Blotting Analysis
3.7. Wound Healing Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. GLOBOCAN. Available online: http://globocan.iarc.fr/Pages/fact_sheets_cancer.aspx (accessed on 24 December 2014).
- Tanoue, L.T.; Detterbeck, F.C. New TNM classification for non-small-cell lung cancer. Expert Rev. Anticancer Ther. 2009, 9, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Sandler, A.; Gray, R.; Perry, M.C.; Brahmer, J.; Schiller, J.H.; Dowlati, A.; Lilenbaum, R.; Johnson, D.H. Paclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancer. N Engl. J. Med. 2006, 355, 2542–2550. [Google Scholar] [CrossRef] [PubMed]
- Hanna, N.; Shepherd, F.A.; Fossella, F.V.; Pereira, J.R.; de Marinis, F.; von Pawel, J.; Gatzemeier, U.; Tsao, T.C.; Pless, M.; Muller, T.; et al. Randomized phase III trial of pemetrexed vs. docetaxel in patients with non-small-cell lung cancer previously treated with chemotherapy. J. Clin. Oncol. 2004, 22, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Su, J.H.; Tsao, C.Y.; Hung, C.T.; Chao, H.H.; Lin, J.J.; Liao, M.H.; Yang, Z.Y.; Huang, H.H.; Tsai, F.J.; et al. Sinulariolide induced hepatocellular carcinoma apoptosis through activation of mitochondrial-related apoptotic and PERK/eLF2á/ATF4/CHOP pathway. Molecules 2013, 18, 10146–10161. [Google Scholar] [CrossRef] [PubMed]
- Li, H.H.; Su, J.H.; Chiu, C.C.; Lin, J.J.; Yang, Z.Y.; Hwang, W.I.; Chen, Y.K.; Lo, Y.H.; Wu, Y.J. Proteomic investigation of the sinulariolide-treated melanoma cells A375: Effects on the cell apoptosis through mitochondrial-related pathway and activation of caspase cascade. Mar. Drugs 2013, 11, 2625–2642. [Google Scholar] [CrossRef] [PubMed]
- Neoh, C.A.; Wang, R.Y.; Din, Z.H.; Su, J.H.; Chen, Y.K.; Tsai, F.J.; Weng, S.H.; Wu, Y.J. Induction of apoptosis by sinulariolide from soft coral through mitochondrial-related and p38MAPK pathways on human bladder carcinoma cells. Mar. Drugs 2012, 10, 2893–2911. [Google Scholar] [CrossRef] [PubMed]
- Gelperina, S.; Kisich, K.; Iseman, M.D.; Heifets, L. The potential advantages of nanoparticle drug delivery systems in chemotherapy of tuberculosis. Am. J. Respir. Crit. Care Med. 2005, 172, 1487–1490. [Google Scholar] [CrossRef] [PubMed]
- Teong, B.; Lin, C.Y.; Chang, S.J.; Niu, G.C.; Yao, C.H.; Chen, I.F.; Kuo, S.M. Enhanced anti-cancer activity by curcumin-loaded hydrogel nanoparticle derived aggregates on A549 lung adenocarcinoma cells. J. Mater. Sci. Mater. Med. 2015, 26, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Toole, B.P.; Kinney, S.D.; Kuo, J.W.; Stamenkovic, I. Inhibition of tumor growth in vivo by hyaluronan oligomers. Int. J. Cancer 1998, 77, 396–401. [Google Scholar] [CrossRef]
- Arpicco, S.; Milla, P.; Stella, B.; Dosio, F. Hyaluronic acid conjugates as vectors for the active targeting of drugs, genes and nanocomposites in cancer treatment. Molecules 2014, 19, 3193–3230. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.H.; Chang, S.J.; Chuang, C.W.; Kuo, S.M.; Chen, S.Y.; Lu, R.T. Preparation of bio-polymeric nanoparticles by electrostatic field system. Micro. Nano Lett. 2012, 7, 997–1000. [Google Scholar] [CrossRef]
- Mitra, S.; Gaur, U.; Ghosh, P.C.; Maitra, A.N. Tumour targeted delivery of encapsulated dextran–doxorubicin conjugate using chitosan nanoparticles as carrier. J. Control. Release 2001, 74, 317–323. [Google Scholar] [CrossRef]
- Huang, K.J.; Chen, Y.C.; El-Shazly, M.; Du, Y.C.; Su, J.H.; Tsao, C.W.; Yen, W.H.; Chang, W.B.; Su, Y.D.; Yeh, Y.T.; Lu, M.C. 5-Episinuleptolide acetate, a norcembranoidal diterpene from the formosan soft coral Sinularia sp., induces leukemia cell apoptosis through Hsp90 inhibition. Molecules 2013, 18, 2924–2933. [Google Scholar] [CrossRef] [PubMed]
- Su, C.C.; Su, J.H.; Lin, J.J.; Chen, C.C.; Hwang, W.I.; Huang, H.H.; Wu, Y.J. An investigation into the cytotoxic effects of 13-acetoxysarcocrassolide from the soft coral Sarcophyton crassocaule on bladder cancer cells. Mar. Drugs 2011, 9, 2622–2642. [Google Scholar] [CrossRef] [PubMed]
- Su, T.R.; Lin, J.J.; Chiu, C.C.; Chen, J.Y.; Su, J.H.; Cheng, Z.J.; Hwang, W.I.; Huang, H.H.; Wu, Y.J. Proteomic investigation of anti-tumor activities exerted by sinularin against A2058 melanoma cells. Electrophoresis 2012, 33, 1139–1152. [Google Scholar] [CrossRef] [PubMed]
- Bombelli, F.B.; Wedster, C.A.; Moncrieff, M.; Sherwood, V. The scope of nanoparticle therapies for future metastatic melanoma treatment. Lancet Oncol. 2014, 15, e22–e32. [Google Scholar] [CrossRef]
- Choi, K.Y.; Saravanakumar, G.; Park, J.H.; Park, K. Hyaluronic acid-based nanocarriers for intracellular targeting: Interfacial interactions with proteins in cancer. Colloids Surf. 2012, 99, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Bilensoy, E. Cationic nanoparticles for cancer therapy. Expert Opin. Drug Deliv. 2010, 7, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Rivkin, I.; Cohen, K.; Koffler, J.; Melikhov, D.; Peer, D.; Margalit, R. Paclitaxel-clusters coated with hyaluronan as selective tumor-targeted nanovectors. Biomaterials 2010, 31, 7106–7114. [Google Scholar] [CrossRef] [PubMed]
- Iversen, T.G.; Skotland, T.; Sandvig, K. Endocytosis and intracellular transport of nanoparticles: Present knowledge and need for future studies. Nano Today 2011, 6, 176–185. [Google Scholar] [CrossRef]
- Arora, H.C.; Jensen, M.P.; Yuan, Y. Nanocarriers enhance doxorubicin uptake in drug-resistant ovarian cancer cells. Cancer Res. 2012, 72, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Teong, B.; Chen, I.F.; Chang, S.J.; Gao, J.; Kuo, S.M. Enhanced apoptotic effects of dihydroartemisinin-aggregated gelatin and hyaluronan nanoparticles on human lung cancer cells. J. Biomed. Mater. Res. B Appl. Biomater. 2014, 102, 455–462. [Google Scholar] [CrossRef] [PubMed]
- El-Dakdouki, M.H.; Xia, J.; Zhu, D.C.; Kavunja, H.; Grieshaber, J.; O’Reilly, S.; McCormick, J.J.; Huang, X. Assessing the in vivo efficacy of doxorubicin loaded hyaluronan nanoparticles. ACS Appl. Mater. Interfaces 2014, 6, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Prestwich, G.D.; Marecak, D.M.; Marecek, J.F.; Vercruysse, K.P.; Ziebell, M.R. Controlled chemical modification of hyaluronic acid: Synthesis, applications, and biodegradation of hydrazide derivatives. J. Control. Release 1998, 531, 93–103. [Google Scholar] [CrossRef]
- Jaracz, S.; Chen, J.; Kuznetsova, L.V.; Ojima, I. Recent advances in tumor-targeting anticancer drug conjugates. Bioorg. Med. Chem. 2005, 13, 5043–5054. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Azpurua, J.; Hine, C.; Vaidya, A.; Myakishev-Rempel, M.; Ablaeva, J.; Mao, Z.; Nevo, E.; Gorbunova, V.; Seluanov, A. High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature 2013, 499, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Chan, T.M.; Wu, Y.J.; Chen, J.J. Review: Application of Nanoparticles in Urothelial Cancer of the Urinary Bladder. J. Med. Biol. Eng. 2015, 35, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Rosse, T.; Olivier, R.; Monney, L.; Rager, M.; Conus, S.; Fellay, I.; Jansen, B.; Borner, C. Bcl-2 prolongs cell survival after Bax-induced release of cytochrome c. Nature 1998, 391, 496–499. [Google Scholar] [PubMed]
- Pastorino, J.G.; Chen, S.T.; Tafani, M.; Snyder, J.W.; Farber, J.L. The overexpression of Bax produces cell death upon induction of the mitochondrial permeability transition. J. Biol. Chem. 1998, 273, 7770–7775. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Sinulariolide was purchased from National Museum of Marine Biology and Aquarium, Pingtung, Taiwan.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Early Apoptosis | Late Apoptosis |
|---|---|---|
| DMSO | 0.8% | 0.1% |
| SNL 25 μg/mL | 1.5% | 0.6% |
| SNL 50 μg/mL | 2.4% | 0.9% |
| SNL 75 μg/mL | 12.8% | 0.1% |
| HA/SNL 25 | 5.3% | 1.4% |
| HA/SNL 50 | 43.7% | 6.4% |
| Conditions | Sub G1 | G1 | S | G2/M |
|---|---|---|---|---|
| DMSO | 0.1% | 58.7% | 22.4% | 20.8% |
| SNL 25 μg/mL | 1.9% | 59.6% | 19.6% | 21.3% |
| SNL 50 μg/mL | 3.4% | 47.2% | 16.4% | 34.8% |
| SNL 75 μg/mL | 3.5% | 55.5% | 13.4% | 30.6% |
| HA/SNL 25 | 7.4% | 55.0% | 11.4% | 28.8% |
| HA/SNL 50 | 9.2% | 55.2% | 13.1% | 25.6% |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, K.Y.; Wu, Y.-J.; Liu, Z.N.; Chuang, C.W.; Huang, H.H.; Kuo, S.M. Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells. Molecules 2016, 21, 297. https://doi.org/10.3390/molecules21030297
Hsiao KY, Wu Y-J, Liu ZN, Chuang CW, Huang HH, Kuo SM. Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells. Molecules. 2016; 21(3):297. https://doi.org/10.3390/molecules21030297
Chicago/Turabian StyleHsiao, Kuan Yin, Yi-Jhen Wu, Zi Nong Liu, Chin Wen Chuang, Han Hsiang Huang, and Shyh Ming Kuo. 2016. "Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells" Molecules 21, no. 3: 297. https://doi.org/10.3390/molecules21030297
APA StyleHsiao, K. Y., Wu, Y.-J., Liu, Z. N., Chuang, C. W., Huang, H. H., & Kuo, S. M. (2016). Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells. Molecules, 21(3), 297. https://doi.org/10.3390/molecules21030297