Abstract
Most red wines commercialized in the market use the malolactic fermentation process in order to ensure stability from a microbiological point of view. In this second fermentation, malic acid is converted into l-lactic acid under controlled setups. However this process is not free from possible collateral effects that on some occasions produce off-flavors, wine quality loss and human health problems. In warm viticulture regions such as the south of Spain, the risk of suffering a deviation during the malolactic fermentation process increases due to the high must pH. This contributes to produce wines with high volatile acidity and biogenic amine values. This manuscript develops a new red winemaking methodology that consists of combining the use of two non-Saccharomyces yeast strains as an alternative to the traditional malolactic fermentation. In this method, malic acid is totally consumed by Schizosaccharomyces pombe, thus achieving the microbiological stabilization objective, while Lachancea thermotolerans produces lactic acid in order not to reduce and even increase the acidity of wines produced from low acidity musts. This technique reduces the risks inherent to the malolactic fermentation process when performed in warm regions. The result is more fruity wines that contain less acetic acid and biogenic amines than the traditional controls that have undergone the classical malolactic fermentation.
1. Introduction
Pasteur, at the beginning of his oenological studies, considered malolactic fermentation as something unwanted, as he viewed lactic bacteria to be wine spoilage microorganisms. Later on, it has been assumed that to perform malolactic fermentations under controlled conditions is the best and almost unique way to stabilize a red wine from a microbiological point of view. Nevertheless in the last few years it has been proved that there are other different yeast species able to consume malic acid [1,2,3,4] and also others able to produce lactic acid [4,5,6,7,8].
The presence of non-Saccharomyces wild yeasts in fermentations was traditionally associated with high levels of acetic acid and other off-flavours. Nevertheless, nowadays researchers and winemakers are aware of the positive influence of non-Saccharomyces in wine quality complexity [8]. When the main objective is to produce dry wine, the difficulty with which non-Saccharomyces wine yeast finishes the alcoholic fermentation requires the development of multi-starter fermentations with Saccharomyces cerevisiae or another high fermentative yeast species as a binding partner. Nevertheless, some of these non-Saccharomyces could be used alone in the production of sweet wines. Some enzymatic properties (glycosidases, β-lyase, etc.), ethanol reduction and the release of some interesting metabolites such as glycerol, pyruvic acid, and mannoproteins among others, are the main highlights that justify the interest in these mixed fermentations [1,9,10].
Some studies have analyzed the use and influence of different non-Saccharomyces species in wine quality. Some of these yeast species are Kloeckera apiculata [11], Hanseniaspora uvarum [12], Hanseniaspora viane [13], Torulospora delbrueckii [14,15,16], Candida pulcherrima [16,17,18], Candida zemplinina [19], Zygosaccharomyces bailii [20,21], Schizosaccharomyces pombe [22], Lachancea thermotolerans [7] and Hansenula anomala [23,24]. Most of these studies report sequential inoculation of a non-Saccharomyces and a Saccharomyces cerevisiae as the best option.
Among non-Saccharomyces yeast species, Schizosaccharomyces pombe has been used for deacidification purposes, due to its ability to convert l-malic acid into ethanol [25]. On the other hand, during the last years new uses of this genus have been developed [1]. One of these new uses is its application in ageing over lees, due to their polysaccharide release superiority [26]. The literature also describes the use of certain Schizosaccharomyces mutants to reduce the initial content of gluconic acid in spoiled grape musts [27,28,29,30,31]. S. pombe fermentation also provides a way of increasing the overall pyranoanthocyanin content in red wines [32,33]. Nevertheless, due to the great variability in the genetical composition of S. pombe [34], further selection processes must be performed [35,36] in order to obtain proper strains for winemaking. Lachancea thermotolerans has been recently described for acidification of low acidic musts [5,6,7].
This study demonstrates that it is possible to produce a quality wine without using the genera Saccharomyces and to avoid any possible collateral effects produced by lactic bacteria in wines with high pH and high alcohol content. In these cases it is very difficult to develop a proper malolactic fermentation process without any deviation. For these reasons, the combined use of Lachancea thermotolerans and Schizosaccharomyces pombe is proposed as an alternative to the classical malolactic fermentation in red wine.
2. Results and Discussion
2.1. Fermentation Kinetics
2.1.1. Yeast Population Kinetic
Figure 1 shows the different yeast strain population development during the fermentation processes. In sequential fermentations, when Saccharomyces cerevisiae 87 or Schizosaccharomyces pombe V2 were inoculated, Kluyveromyces thermotolerans CONCERTO™ started to decline fast by day 4, although it was faster in the case involving Saccharomyces.
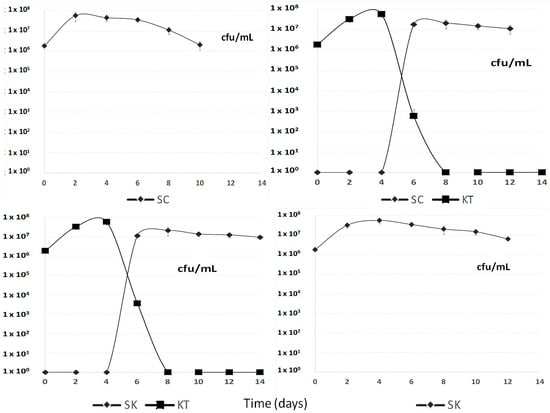
Figure 1.
Population development of Saccharomyces cerevisiae 87 (SC), Kluyveromyces thermotolerans CONCERTO™ (KT) and Schizosaccharomyces pombe V2 (SK) during the different sequential fermentation processes.
Figure 1.
Population development of Saccharomyces cerevisiae 87 (SC), Kluyveromyces thermotolerans CONCERTO™ (KT) and Schizosaccharomyces pombe V2 (SK) during the different sequential fermentation processes.
The early disappearance of Kluyveromyces thermotolerans CONCERTO™ could be explained due to presence of an ethanol concentration higher than 6% v/v by day 4, although this species has been reported to tolerate up to 9% v/v ethanol when it ferments by itself [5]. This low alcohol tolerance by Lachancea thermotolerans, make it impossible to produce a dry red wine in warm regions without using another more fermentative yeast in a sequential fermentation.
2.1.2. Sugar Consumption Kinetics
The fermentations involving Saccharomyces cerevisiae 87 (SC) (Figure 2A) consumed the sugar the fastest. Slower glucose and fructose consumption kinetics have been described before for Schizosaccharomyces pombe in spite of the fact that this yeast is able to consume all sugar in a regular must [22,36]. All the studied fermentations were finished properly between days 10 and 14 reaching values lower than 3 g/L in the sum of glucose and fructose, although there were some differences between them (Figure 2A).
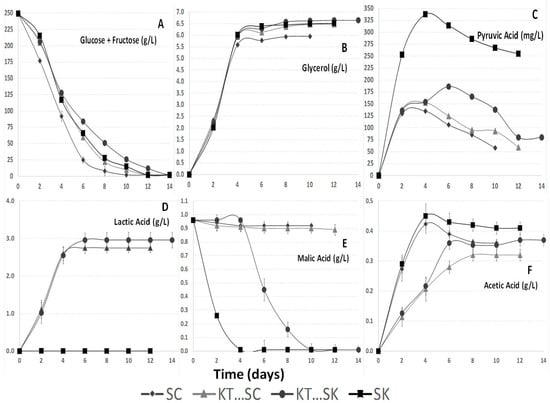
Figure 2.
(A) Glucose + fructose concentrations (g/L); (B) Glycerol concentrations (g/L); (C) Pyruvic acid (mg/L); (D) l-Lactic acid concentrations (g/L); (E) l-Malic acid concentrations (g/L); (F) Acetic acid concentrations (g/L). Parameters of the studied wines based on Tempranillo variety during fermentations performed by Saccharomyces cerevisiae 87 by itself (SC), sequential fermentation with Saccharomyces cerevisiae 87 and Kluyveromyces thermotolerans CONCERTO™ (KT···SC), sequential fermentation with Schizosaccharomyces pombe V2 and Kluyveromyces thermotolerans CONCERTO™ (KT…SK), and Schizosaccharomyces pombe V2 by itself (SK).
Figure 2.
(A) Glucose + fructose concentrations (g/L); (B) Glycerol concentrations (g/L); (C) Pyruvic acid (mg/L); (D) l-Lactic acid concentrations (g/L); (E) l-Malic acid concentrations (g/L); (F) Acetic acid concentrations (g/L). Parameters of the studied wines based on Tempranillo variety during fermentations performed by Saccharomyces cerevisiae 87 by itself (SC), sequential fermentation with Saccharomyces cerevisiae 87 and Kluyveromyces thermotolerans CONCERTO™ (KT···SC), sequential fermentation with Schizosaccharomyces pombe V2 and Kluyveromyces thermotolerans CONCERTO™ (KT…SK), and Schizosaccharomyces pombe V2 by itself (SK).
2.2. Chemical Parameter Monitoring
2.2.1. Glycerol
Most glycerol was produced during the first days of fermentation (Figure 2B). The SC fermentation reached the lowest level in glycerol and KT···SK fermentation showed the highest final content. Lachancea and Schizosaccharomyces genera have been reported before as higher glycerol producers than Saccharomyces [7,33,37]. Final levels of glycerol varied from 5.96 g/L to 6.65 g/L (Table 1). Increased glycerol content is described as one of the main contributions of non-Saccharomyces strains on wine quality [38].

Table 1.
Final analysis of Saccharomyces cerevisiae 87 by itself (SC), sequential fermentation with Saccharomyces cerevisiae 87 and Kluyveromyces thermotolerans CONCERTO™ (KT···SC), sequential fermentation with Schizosaccharomyces pombe V2 and Kluyveromyces thermotolerans CONCERTO™ (KT…SK), Schizosaccharomyces pombe V2 by itself (SK), and fermentations after malolactic fermentation with Oenococcus oeni 217 (+ MLF).
| Compounds | SC | SC + MLF | KT···SC | KT···SC + MLF | KT···SK | SK |
|---|---|---|---|---|---|---|
| l-Lactic Acid (g/L) | 0.01 ± 0.01a | 0.54 ± 0.08b | 2.75 ± 0.12c | 3.27 ± 0.19d | 2.96 ± 0.21c | 0.01 ± 0.01a |
| l-Malic Acid (g/L) | 0.92 ± 0.02b | 0.01 ± 0.01a | 0.89 ± 0.04b | 0.01 ± 0.01a | 0.01 ± 0.01a | 0.01 ± 0.01a |
| Acetic Acid (g/L) | 0.36 ± 0.01b | 0.44 ± 0.05c | 0.32 ± 0.02a | 0.39 ± 0.04bc | 0.37 ± 0.02b | 0.41 ± 0.02c |
| Residual Sugar (g/L) | 2.08 ± 0.30b | 0.12 ± 0.04a | 2.22 ± 052b | 0.16 ± 0.04a | 2.41 ± 0.58b | 2.13 ± 0.17b |
| Glycerol (g/L) | 5.96 ± 0.02a | 5.89 ± 0.05a | 6.48 ± 0.05b | 6.36 ± 0.06b | 6.65 ± 0.04c | 6.59 ± 0.03bc |
| Free SO2 (mg/L) | 26.12 ± 2.38a | 25.25 ± 3.43ab | 25.25 ± 3.28ab | 21.15 ± 1.28b | ||
| Total SO2 (mg/L) | 56.52 ± 2.43b | 44.13 ± 3.16a | 46.50 ± 3.21a | 58.58 ± 1.15b | ||
| Alcohol (% v/v) | 14.56 ± 0.01c | 14.54 ± 0.02c | 14.20 ± 0.04b | 14.18 ± 0.06b | 14.03 ± 0.05a | 14.23 ± 0.02b |
| pH | 3.94 ± 0.01c | 3.99 ± 0.02d | 3.74 ± 0.02a | 3.79 ± 0.02b | 3.83 ± 0.02b | 4.03 ± 0.02d |
| Urea | 1.43 ± 0.01b | 1.45 ± 0.02b | 0.12 ± 0.04a | 0.08 ± 0.01a | ||
| Color Intensity | 6.16 ± 0.03b | 5.38 ± 0.06a | 6.29 ± 0.06c | 5.51 ± 0.07a | 6.42 ± 0.08c | 6.88 ± 0.03d |
| Citric Acid (g/L) | 0.22 ± 0.01a | 0.03 ± 0.02b | 0.24 ± 0.03a | 0.04 ± 0.03b | 0.23 ± 0.03a | 0.22 ± 0.02a |
Results represent the mean ± SD for three replicates. Means in the same row with the same letter are not significantly different (p < 0.05).
2.2.2. Pyruvic Acid
The highest levels of pyruvic acid were formed during the first days of fermentation (Figure 2C), except for the KT···SK fermentation where another pyruvic formation peak appeared at day 6. The non-Saccharomyces yeast produce occasionally more pyruvic acid and more glycerol, both being derived from the glyceropyruvic pathway [38,39,40]. The maximum pyruvic acid concentrations reached were higher than those recorded in earlier works performed using Saccharomyces cerevisiae strains selected for their ability to produce pyruvic acid; these produced only between 60 and 130 mg/L of pyruvic acid [36] compared to the 186.38 mg/L reached in this study (Figure 2C) by KT···SK fermentation and the 337.67 mg/L produced by SK. In fermentations where Schizosaccharomyces was involved the pyruvic acid production was the highest. Similar results have been reported before [1]. Higher levels of pyruvic acid could be interesting for red wines because it contributes to the production of highly stable color compounds [32,36].
2.2.3. Alcohol
The alcohol levels varied from 14.03 to 14.56 (% vol/vol) (Table 1). The sugar consumption can also be used to produce higher amounts of compounds other than ethanol, such as glycerol or pyruvic acid, or to increase the yeast biomass [41,42]. The results obtained showed that fermentations involving non-Saccharomyces produced lower ethanol levels. These data agree with other authors who confirmed that some non-Saccharomyces types of yeast give lower ethanol yields than Saccharomyces [10,17,43,44]. Previous studies showed similar results for Lachancea thermotolerans [7] and Schizosaccharomyces pombe [22].
2.2.4. SO2
The final total SO2 levels varied from 44.13 to 58.58 mg/L (Table 1). Lachancea thermotolerans fermentations showed lower final concentrations of total SO2 than fermentations with SC and SK. The manufacturer (Chr. Hansen, Hørsholm, Denmark) offers it as a way of reducing the risk of presenting H2S faults. A possible reason for lower sulfur metabolism could be the lower SO2 tolerance reported for several strains of non-Saccharomyces.
2.2.5. l-Lactic Acid
Figure 2D shows that Kluyveromyces thermotolerans Concerto™ (KT) produced l-lactic acid (Table 1) during alcoholic fermentation. The final l-lactic acid produced by Lachancea thermotolerans in this study varied from 2.75 to 2.96 g/L, which clearly influenced the final pH (Table 1). Other authors [6] have also observed significant acidification using mixed cultures of Lachancea thermotolerans with the main objective of increasing must acidity. The production of l-lactic is also linked to the viable cell concentration [37]. In this study l-lactic production stopped when the Lachancea thermotolerans population started to decrease. The assays performed on malolactic fermentations showed an increase in l-lactic acid of about 0.54 g/L (Table 1). These levels were lower than the cases involving Lachancea thermotolerans, due to the low initial level of malic acid in the studied must.
2.2.6. l-Malic Acid
Figure 2E shows a progressive decrease to about 0 g/L in malic acid in all fermentations involving Schizosaccharomyces pombe. Schizosaccharomyces is the only yeast genus able to reduce efficiently malic acid concentration in must [1] during alcoholic fermentation.
2.2.7. Acetic Acid
Previous experiments with Lachancea thermotolerans reported significant reductions in acetic acid content [7,37]. On the other hand Schizosaccharomyces has been reported to produce acetic acid concentrations up to 1 g/L as main collateral effect [32]. Nevertheless, nowadays there are strains with reduced collateral effects [36]. The acetic acid levels obtained after alcoholic fermentation varied from of 0.32 to 0.41 g/L (Figure 2F). Those values were not excessive and they did not affect wine quality negatively. After malolactic fermentation took place in fermentations involving Saccharomyces cerevisiae, small statistical differences were reported (Table 1).
2.2.8. Biogenic Amines
The final levels of biogenic amines were lower than 2 mg/L (Table 2). This histamine value is considered the lowest level [45]. Fermentations involving Schizosaccharomyces pombe showed lower levels than those that where malolactic fermentation was performed (Table 2). The use of Schizosaccharomyces is of interest to reduce the possibility of lactic acid bacteria growing by removing malic acid (another nutrient source), thus reducing the risk of biogenic amine [1,46] or ethyl carbamate [47] formation. The urea content of the finished wines was less than 0.2 mg/L (Table 1) for fermentations involving Schizosaccharomyces pombe. The reported differences were attributed to the special ability of Schizosaccharomyces to metabolize urea [48]. This enzymatic activity also could reduce the initial level of ethyl carbamate precursors [1].
Table 2.
Biogenic amines analysis of Saccharomyces cerevisiae 87 by itself (SC), sequential fermentation with Saccharomyces cerevisiae 87 and Kluyveromyces thermotolerans CONCERTO™ (KT···SC), sequential fermentation with Schizosaccharomyces pombe V2 and Kluyveromyces thermotolerans CONCERTO™ (KT…SK), Schizosaccharomyces pombe V2 by itself (SK), and fermentations after malolactic fermentation with Oenococcus oeni 217 (+ MLF).
| Compounds | SC | SC + MLF | KT···SC | KT···SC + MLF | KT···SK | SK |
|---|---|---|---|---|---|---|
| Histamine (mg/L) | 0.43 ± 0.02a | 1.46 ± 0.06b | 0.42 ± 0.04a | 1.48 ± 0.15b | 0.44 ± 0.04a | 0.38 ± 0.02a |
| Tiramine (mg/L) | 0.25 ± 0.01a | 0.36 ± 0.04b | 0.26 ± 0.02a | 0.38 ± 0.06b | 0.22 ± 0.03a | 0.26 ± 0.03a |
| Phenylethylamine (g/L) | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Putrescine (g/L) | 1.78 ± 0.03a | 2.18 ± 0.18b | 1.82 ± 0.11a | 2.24 ± 0.21b | 1.71 ± 0.08a | 1.88 ± 0.07a |
| Cadaverine (g/L) | 0.51 ± 0.02a | 0.65 ± 0.04b | 0.49 ± 0.05a | 0.69 ± 0.07b | 0.52 ± 0.03ab | 0.55 ± 0.03a |
Results represent the mean ± SD for three replicates. Means in the same row with the same letter are not significantly different (p < 0.05).
2.3. Sensory Evaluation
During an informal tasting session differences in color, aroma, and taste were found between the wines. No apparent off-flavors were detectable. However, a full sensory analysis is needed to confirm and validate these findings.
3. Experimental Section
3.1. Microorganisms
The following yeasts were used for the experimental fermentations: Kluyveromyces thermotolerans Concerto™ (Hansen, Hørsholm, Denmark; www.chr-hansen.com) that belongs to the yeast species Lachancea thermotolerans, Saccharomyces cerevisiae 87 (Spanish Type Culture Collection, Valencia, Spain) and Schizosaccharomyces pombe V2 (Chemistry and Food Technology department, Polytechnic University of Madrid, Spain [36]). The strain of lactic acid bacteria used was Oenococcus oeni 217 (Spanish Type Culture Collection, Valencia, Spain).
3.2. Vinification
All fermentations were undertaken using the must of Vitis vinifera L. cultivar Tempranillo grapes grown in El Socorro Experimental vineyard (Madrid, Spain). The must was pasteurized at 105 °C for 5 min. A microvinification method similar to those described in scientific literature was used [15,22,32,33,49]. Pasteurized must (4 L) was placed in 5 L glass tanks. This allowed an adequate space for the release of carbon dioxide during the fermentation process. No sulphur dioxide was added to any vessel. Sugar concentration was 249.33 g/L, pH = 3.92, primary amino nitrogen (PAN) 167 g/L, malic acid 0.96 g/L, citric acid 0.24 g/L, lactic and acetic acid bellow 0.1 g/L. To provide nutrition 60 g/hL of Actimax NATURA (Agrovín S.A., Ciudad Real, Spain) were added. Four assays were performed (all in triplicate): (i) inoculation of the must with S. cerevisiae 87 (106 CFU/mL) alone (SC); (ii) inoculation of the must with K. thermotolerans Concerto™ (107 CFU/mL) followed by S. cerevisiae 87 (106 CFU/mL) 96 h later (KT···SC); (iii) inoculation of the must with K. thermotolerans Concerto™ (106 CFU/mL) followed by S. pombe V2 (106 CFU/mL) 96 h later (KT···SK); and (iv) inoculation of the must with S. pombe V2 alone (SK). Yeast inocula were performed using 100 mL of sterilized must with 1 mL of yeast extract dextrose peptone liquid medium [50] containing 108 CFU/mL (determined using a Thomas chamber). To reach this population, 100 μL of each yeast suspension were cultivated in 10 mL of YEPD at 25 °C for 24 h. This procedure was repeated three successive times before the final inoculation of 1 mL in the inocula. All inocula were performed in 250-mL flasks sealed with a Müller valve filled with 98% H2SO4 (Panreac, Barcelona, Spain), which allowed the release of CO2 while avoiding microbial contamination [51]. The temperature was maintained at 25 °C for 48 h. The progress of the inocula was developed under anaerobic conditions. All fermentations were performed in triplicate. All fermentation processes were carried out at 20 °C. When the sugar content was below 3 g/L, the wines were racked and stabilized during 7 days at 4 °C concluding with the final product being bottled. Then a concentration of 50 mg/L of sulphur dioxide in potassium metabisulfite form was added. Sealed bottles were placed horizontally in a climate chamber at 4 °C until the sensory evaluation took place. The wines fermented with Saccharomyces cerevisiae by itself (SC), were stabilized and racked following the same procedure, since they finished malolactic fermentation by Oenococcus oeni 217 (106 CFU/mL) in 2.8 L vessels at 18 °C. Then they remained under the same final storage conditions described above, for one month before tasting sessions took place.
3.3. Analytical Determinations of Non-Volatile Compounds
Glucose and fructose, l-lactic acid, acetic acid, glycerol, pyruvic acid, citric acid, l-malic acid, urea and primary amino nitrogen were all determined using a Y15 enzymatic autoanalyzer (Biosystems S.A, Barcelona, Spain) and its proper kits. Ethanol, pH, free SO2, total SO2 were determined following the methods in the Compendium of International Methods of Analysis of Musts and Wines [52].
3.4. Microvinifications Growth Kinetics
During fermentations, aliquots were taken periodically under aseptic conditions and further seria lten-fold dilutions were made. Yeast growth kinetics were monitored by plating 100 μL of the appropriate dilution on lysine media (non-Saccharomyces counts; [53]), YEPD media (total yeast counts; [50]) and YEPDActBzCl media (Schizosaccharomyces counts; [35,36]) based on actidione and benzoic acid as main inhibitor agents. In KT···SC fermentations the population of Lachancea thermotolerans was estimated by the difference between YEPD and Lysine media counts. In KT···SK fermentations the population of Lachancea thermotolerans was estimated by the difference between YEPD and YEPDActBzCl media counts. Colonies were counted after growth at 30 °C for 48–72 h. Lactic bacteria were monitored in MRS agar (Oxoid, Basingstoke, UK).
3.5. Analytical Determinations of Biogenic Amines
The aminoacids were analysed using a Jasco (Tokyo, Japan) UHPLC chromatograph series X-LCTM, equipped with a 3120-FP fluorescence detector. Gradients of solvent A (methanol/acetonitrile, 50:50, v/v) and B (sodium acetate /tetrahydrofuran, 99:1, v/v) were used in a C18 (HALO, city, state abbrev USA) column (100 mm × 2.1 mm; particle size 2.7 µm) as follows: 90% B (0.25 mL/min) from 0 to 6 min, 90%–78% B linear (0.2 mL/min) from 6 to 7.5 min, 78% B from 7.5 to 8 min, 78%–74% B linear (0.2 mL/min) from 8 to 8.5 min, 74% B (0.2 mL/min) from 8.5 to 11 min, 74%–50% B linear (0.2 mL/min) from 11 to 15 min, 50% B (0.2 mL/min) from 15 to 17 min, 50%–20% B linear (0.2 mL/min) from 17 to 21 min, 20%–90% B linear (0.2 mL/min) from 21 to 25 min and re-equilibration of the column from 25 to 26 min. Detection was performed by scanning in the 340–455 nm range. Quantification was performed by comparison against external standards of the studied amines. The different amines were identified by their retention times.
3.6. Sensory Evaluation
The experimental wines were evaluated by a team of 15 experienced wine tasters (five females and ten males), all employees of the Chemistry and Food Technology Department (Madrid, Spain) and the Estación Enológica de Haro (Haro, Spain). Two visual descriptors, four taste parameters and five aromas were used to evaluate the final fermentations. No specific training was carried out prior to tasting sessions. Twelve wines were evaluated in randomized order. The wines were presented in clear tasting glasses [54] identified by numbers from 1 to twelve and in an air-conditioned (20 °C) tasting room equipped with individual booths. Twenty five milliliters of each wine were served at 14 °C in randomized order. The panelists were asked to rate typicality regarding their personal Tempranillo wine concept after testing on an unstructured 10 cm scale, from 0 (no defect) to 10 (very strong defect perceptible), to rate the intensity of the 12 attributes. Additionally, the panelists were asked to name descriptors as free comments for each wine.
3.7. Statistical Analysis
All statistical analyses were performed using PC Statgraphics v.5 software (Graphics Software Systems, Rockville, MD, USA). The significance was set to p < 0.05 for the ANOVA matrix F value. The multiple range test was used to compare the means.
4. Conclusions
The comparison of the results from the fermentation trials showed differences in several analyzed parameters. The combination of the non-Saccharomyces Lachancea thermotolerans and Schizosaccharomyces pombe positively influenced wine quality in the studied case of a low acidic Tempranillo must. Fermentation kinetics showed a fast decline of Lachancea thermotolerans yeast immediately after a more fermentative yeast specie was inoculated. All non-Saccharomyces fermentations produced higher levels of glycerol and pyruvic acid without increasing acetic acidity. Lachancea thermotolerans sulphur dioxide production was significantly lower. All non-Saccharomyces produced reduced ethanol levels. The combination of Lachancea thermotolerans and a selected Schizosaccharomyces pombe strain produced wines stabilised from a malic acid point of view without any need of performing a malolactic fermentation. These wines also showed lower final levels of biogenic amines than the controls that underwent malolactic fermentation.
Acknowledgments
The authors are very grateful to Biosystems S.A., for the donation of the enzyme kits used in this work, and to the accredited Estación Enológica de Haro laboratory, where the biogenic amines analysis were performed, specially to Montserrat Iñiguez and Elena Melendez.
Author Contributions
All authors conceived and designed the experiments; A. Benito performed the vinificatons; S. Benito performed the Biogenic Amines analyses; A. Benito and F. Calderón performed the Microvinifications Growth Kinetics analyses; A. Benito, F. Palomero and S. Benito performed the Analytical Determinations of Non-Volatile Compounds; All authors analyzed and interpreted data, and drafted the paper.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Benito, S.; Palomero, P.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Schizosaccharomyces . In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 3, pp. 365–370. [Google Scholar]
- Rodriguez, S.B.; Roy, J. Factors influencing the utilisation of l-malate by yeasts. Thornton FEMS Microbiol. Lett. 1990, 60, 17–22. [Google Scholar]
- Thornton, R.J.; Rodriguez, S.B. Deacidification of red and white wines by a mutant of Schizosaccharomyces malidevorans under commercial winemaking conditions. Food Microbiol. 1996, 13, 475–482. [Google Scholar] [CrossRef]
- Su, J.; Wang, T.; Wang, Y.; Li, Y.Y.; Li, H. The use of lactic acid-producing, malic acid-producing, or malic acid-degrading yeast strains for acidity adjustment in the wine industry. Appl. Microbiol. Biotechnol. 2014, 98, 2395–2413. [Google Scholar] [CrossRef] [PubMed]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–333. [Google Scholar] [CrossRef] [PubMed]
- Contreras., A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin., C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction of Alcohol Content in Wine. Appl. Environ. Microb. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, T.; Reglero, G.; Herraiz, M.; Martin-Alvarez, P.J.; Cabezudo, M.D. The influence of the yeast and type of culture on the volatile composition of wines fermented without sulfur dioxide. Am. J. Enol. Vitic. 1990, 41, 313–318. [Google Scholar]
- Zironi, R.; Romano, P.; Suzzi, G.; Battistutta, F.; Comi, G. Volatile metabolites produced in wine by mixed and sequential cultures of Hanseniaspora guilliermondii or Kloeckera apiculata and Saccharomyces cerevisiae. Biotechnol. Lett. 1993, 15, 235–238. [Google Scholar] [CrossRef]
- Viana, F.; Belloch, C.; Valles, S.; Manzanares, P. Monitoring a mixed starter of Hanseniaspora vineae-Saccharomyces cerevisiae in natural must: Impact on 2-phenylethyl acetate production. Int. J. Food Microbiol. 2011, 151, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Azzolini, M.; Fedrizzi, B.; Tosi, E.; Finato, F.; Vagnoli, P.; Scrinzi, C.; Zapparoli, G. Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Eur. Food Res. Technol. 2012, 235, 303–313. [Google Scholar] [CrossRef]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderón, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.M.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Marechal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacon, J.J.; Ballester, J.; Vichi, S.; Guerin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, S.; Genna, G.; Gandolfo, V.; Amore, G.; Ciaccio, M.; Oliva, D. Presence of Candida zemplinina in Sicilian musts and selection of a strain for wine mixed fermentations. S. Afr. J. Enol. Vitic. 2012, 33, 80–87. [Google Scholar]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Romani, C.; Comitini, F.; Gobbi, M.; Lencioni, L.; Mannazzu, I.; Ciani, M. Potential spoilage non-Saccharomyces yeasts in mixed cultures with Saccharomyces cerevisiae. Ann. Microbiol. 2011, 61, 137–144. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderon, F.; Palmero, D.; Suarez-Lepe, J.A. Physiological features of Schizosaccharomyces pombe of interest in making of white wines. Eur. Food Res. Technol. 2013, 236, 29–36. [Google Scholar] [CrossRef]
- Izquierdo-Cañas, P.M.; Palacios-Garcia, A.T.; Garcia-Romero, E. Enhancement of flavour properties in wines using sequential inoculations of non-Saccharomyces (Hansenula and Torulaspora) and Saccharomyces yeast starter. Vitis 2011, 50, 177–182. [Google Scholar]
- Izquierdo-Cañas, P.M.; García-Romero, E.; Heras, J.M.; Fernández-González, M. Influence of sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the quality of red wines. Eur. Food Res. Technol. 2014, 239, 279–286. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Palomero, F.; Benito, S.; Calderón, F.; Morata, A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012, 235, 375–383. [Google Scholar] [CrossRef]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar]
- Peinado, R.A.; Moreno, J.J.; Maestre, O.; Ortega, J.M.; Medina, M.; Mauricio, J.C. Gluconic acid consumption in wines by Schizosaccharomyces pombe and its effect on the concentrations of major volatile compounds and polyols. J. Agric. Food. Chem. 2004, 52, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Mauricio, J.C.; Medina, M.; Moreno, J.J. Effect of Schizosaccharomyces pombe on aromatic compounds in dry sherry wines containing high levels of gluconic acid. J. Agric. Food. Chem. 2004, 52, 4529–4534. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Moreno, J.J.; Medina, M.; Mauricio, J.C. Potential application of a glucose-transport-deficient mutant of Schizosaccharomyces pombe for removing gluconic acid from grape must. J. Agric. Food Chem. 2005, 53, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Moreno, J.J.; Maestre, O. Removing gluconic acid by using different treatments with a Schizosaccharomyces pombe mutant: Effect on fermentation by products. Food Chem. 2007, 104, 457–465. [Google Scholar] [CrossRef]
- Peinado, R.A.; Maestre, O.; Mauricio, J.C. Use of a Schizosaccharomyces pombe mutant to reduce the content in gluconic acid of must obtained from rotten grapes. J. Agric. Food Chem. 2009, 57, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Palomero, P.; Morata, A.; Calderón, F.; Suárez-Lépe, J.A. New applications for Schizosaccharomyces pombe in the alcoholic fermentation of red wines. Int. J. Food Sci. Technol. 2012, 47, 2101–2108. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, P.; Gálvez, L.; Morata, A.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Quality and Composition of Red Wine Fermented with Schizosaccharomyces pombe as Sole Fermentative Yeast, and in Mixed and Sequential Fermentations with Saccharomyces cerevisiae. Food Technol. Biotechnol. 2014, 52, 376–382. [Google Scholar]
- Jeffares, D.C. The genomic and phenotypic diversity of Schizosaccharomyces pombe. Nat. Genet. 2015, 47, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Gálvez, L.; Palomero, F.; Calderón, F.; Morata, A.; Suárez-Lepe, J.A. Schizosaccharomyces selective differential media. Afr. J. Microbiol. Res. 2013, 7, 3026–3036. [Google Scholar]
- Benito., S.; Palomero, P.; Calderón, F.; Palmero, D.; Suárez-Lépe, J.A. Selection of Appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 2014, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeast in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with winemaking. World J. Microbiol. Biotechnol. 1998, 14, 199–203. [Google Scholar] [CrossRef]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernández-Orte, P.; Lagarrigue, L.; Lonvaud-Funel, A.; Bely, M. Genetic characterization and phenotypic variability in Torulaspora delbrueckii species: Potential applications in the wine industry. Int. J. Food. Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Merico, A.; Sulo, P.; Piskur, J.; Compagno, C. Fermentative lifestyle in yeasts belonging to the Saccharomyces complex. FEBS J. 2007, 274, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Kutyna, D.R.; Varela, C.; Henschke, P.A.; Chambers, P.J.; Stanley, G.A. Microbiological approaches to lowering ethanol concentration in wine. Trends Food Sci. Technol. 2010, 21, 293–302. [Google Scholar] [CrossRef]
- Blanco, P. Guia sobre la utilización de levaduras no-Saccharomyces en la elaboración de vino, 1st ed.Ministerio de Agricultura, Alimentación y Medio Ambiente (Gobierno de España): Madrid, España, 2014; pp. 9–18.
- Lehtonen, P. Determination of amines and amino acids in wine: A review. Am. J. Enol. Vitic. 1996, 47, 127–133. [Google Scholar]
- Alcaide-Hidalgo, J.M.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Polo, M.C. Influence of malolactic fermentation, postfermentative treatments and ageing with lees on nitrogen compounds of red wines. Food. Chem. 2007, 103, 572–581. [Google Scholar] [CrossRef]
- Tegmo-Larsson, I.M.; Spittler, T.D.; Rodriguez, S.B. Effect of malolactic fermentation on ethyl carbamate formation in Chardonnay. Am. J. Enol. Vitic. 1989, 40, 106–108. [Google Scholar]
- Lubbers, M.W.; Rodriguez, S.B.; Honey, N.K.; Thornton, R.J. Purification and characterization of urease from Schizosaccharomyces pombe. Can. J. Microbiol. 1996, 42, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, T.L.; Kennedy, A.; Vasconcelos, M.C. Use of Microscale Fermentations in Grape and Wine Research. Am. J. Enol. Vitic. 2007, 58, 534–539. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W. Methods for the isolation, maintenance and identification of yeasts. In The Yeast A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; pp. 79–80. [Google Scholar]
- Vaughnan-Martini, A.; Martini, A. Determination of ethanol production. In The Yeast A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; p. 107. [Google Scholar]
- Compendium of International Methods of Wine and Must Analysis; The International Organisation of Vine and Wine: Paris, France, 2015.
- Morris, E.O.; Eddy, A.A. Method for the measurement of wild yeast infection in pitching yeast. J. Inst. Brew. 1957, 63, 34–35. [Google Scholar] [CrossRef]
- International Organization for Standardization. Sensory analysis—Apparatus—Wine-Tasting Glass. 2010. Available online: http://www.iso.org/iso/catalogue_detail?csnumber=9002 (accessed on 11 May 2015).
- Sample Availability: Samples are available from authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).