
Effect of Gedunin on Acute Articular Inflammation and Hypernociception in Mice

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
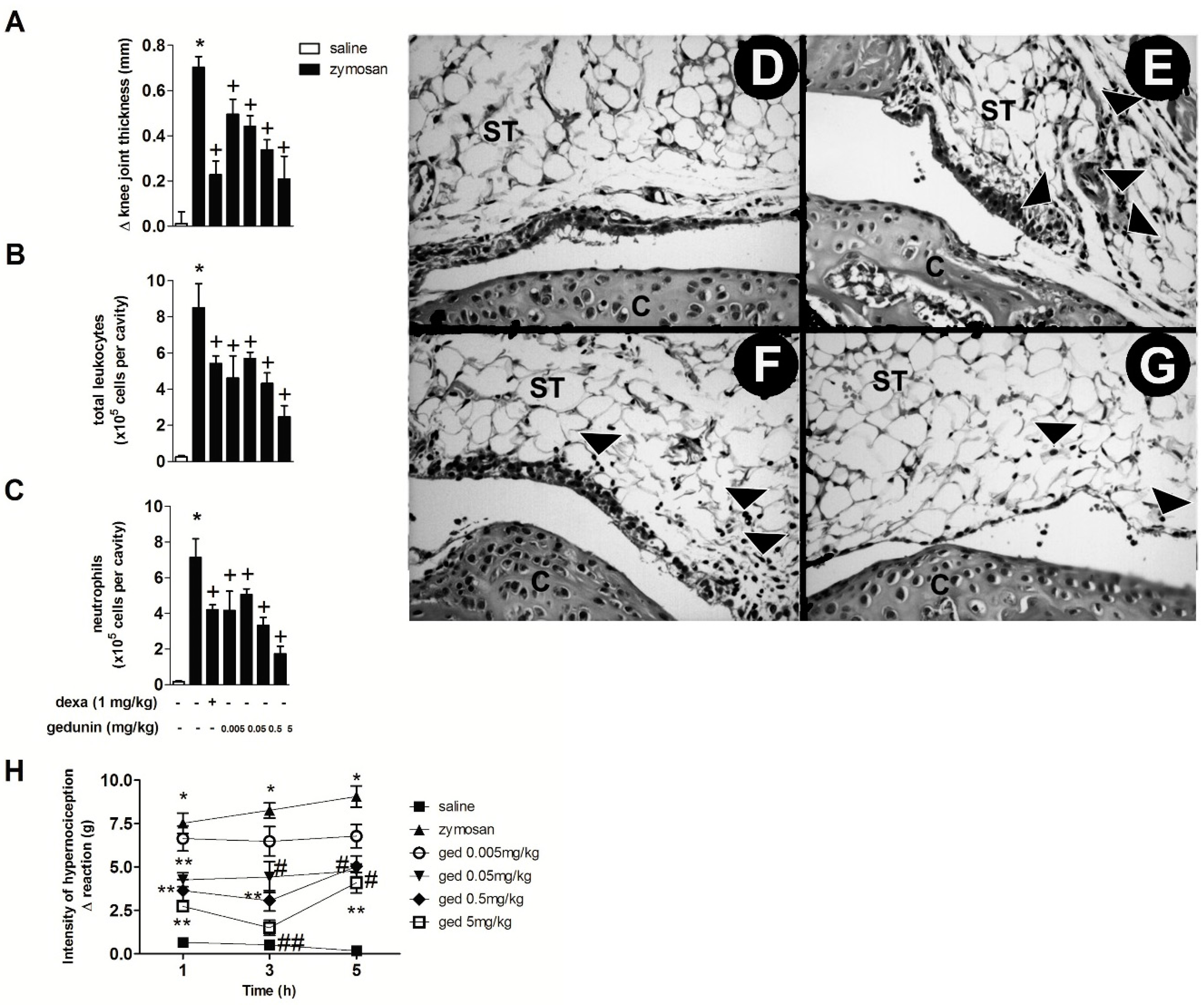
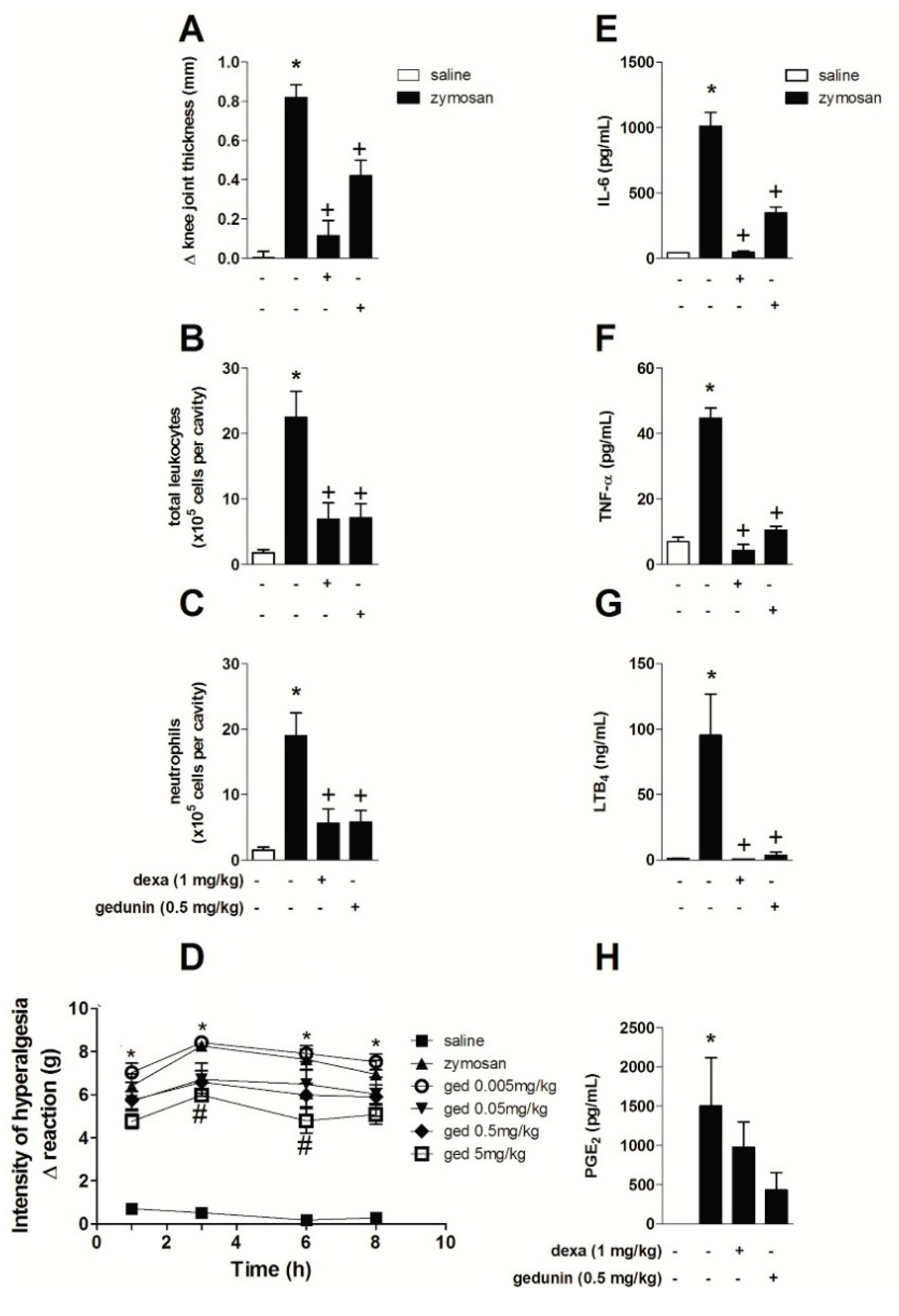
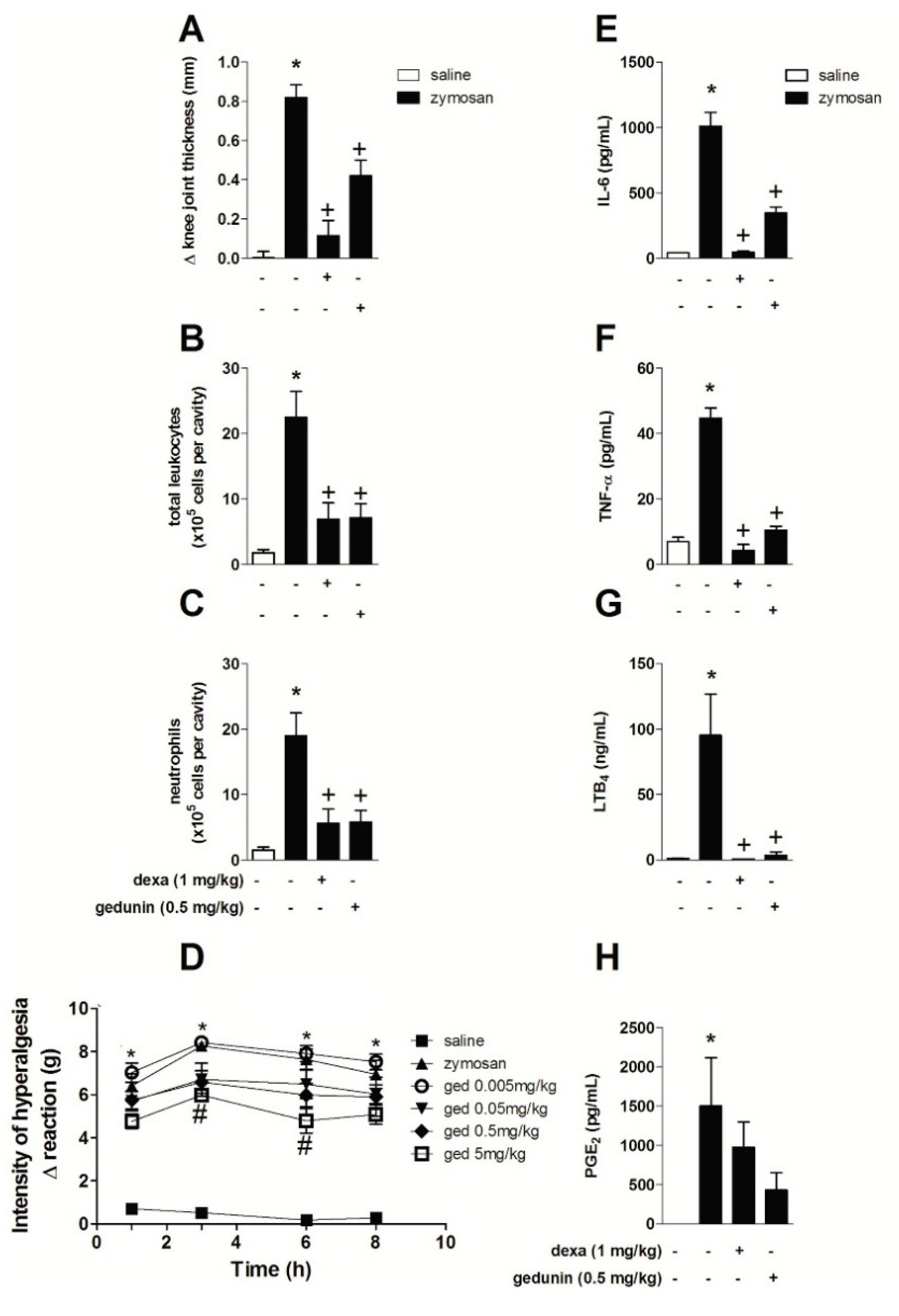
2.1. Gedunin Pre-Treatment Impairs 6 h Zymosan-Induced Articular Edema, Neutrophil Accumulation and Hypernociception
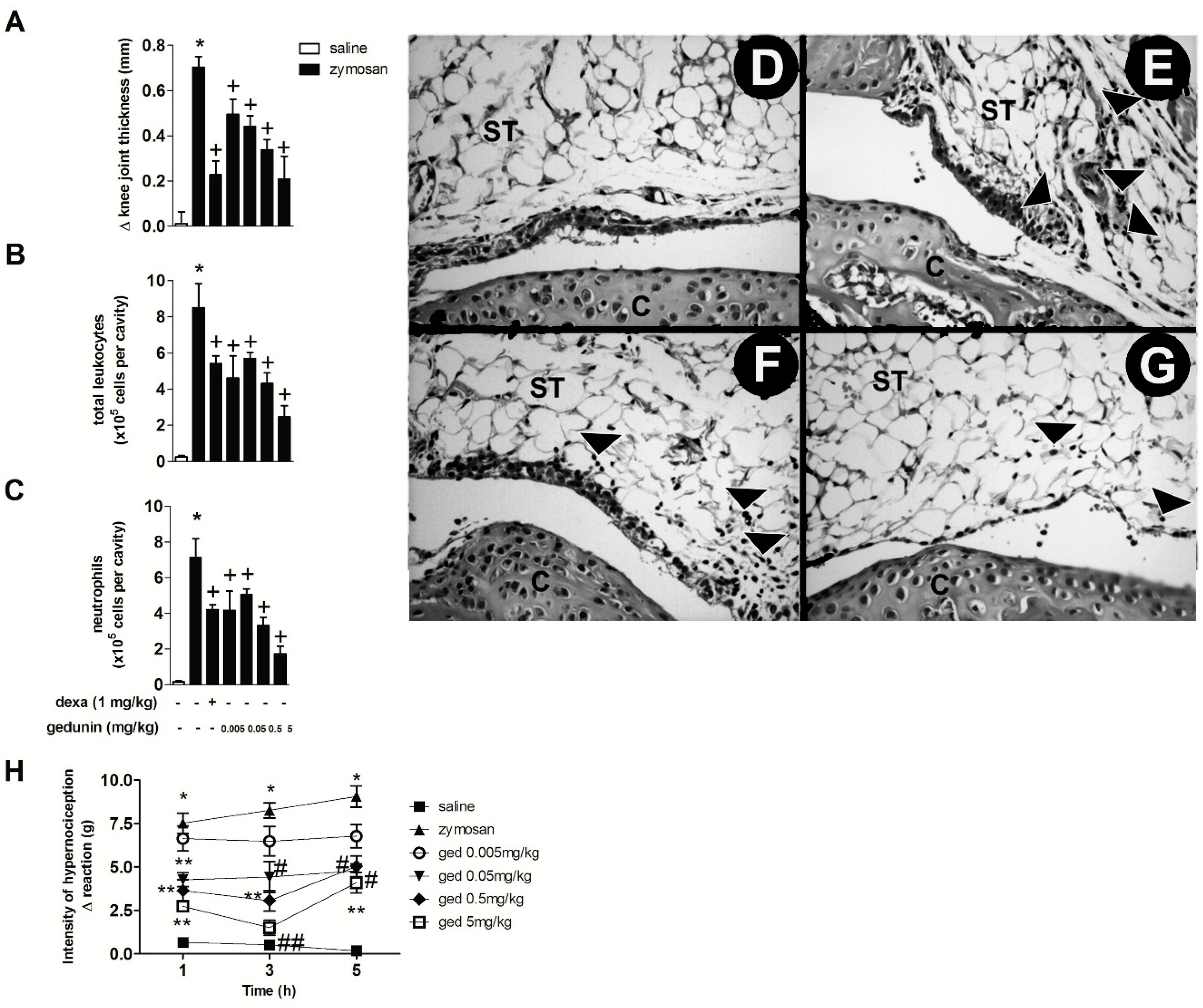
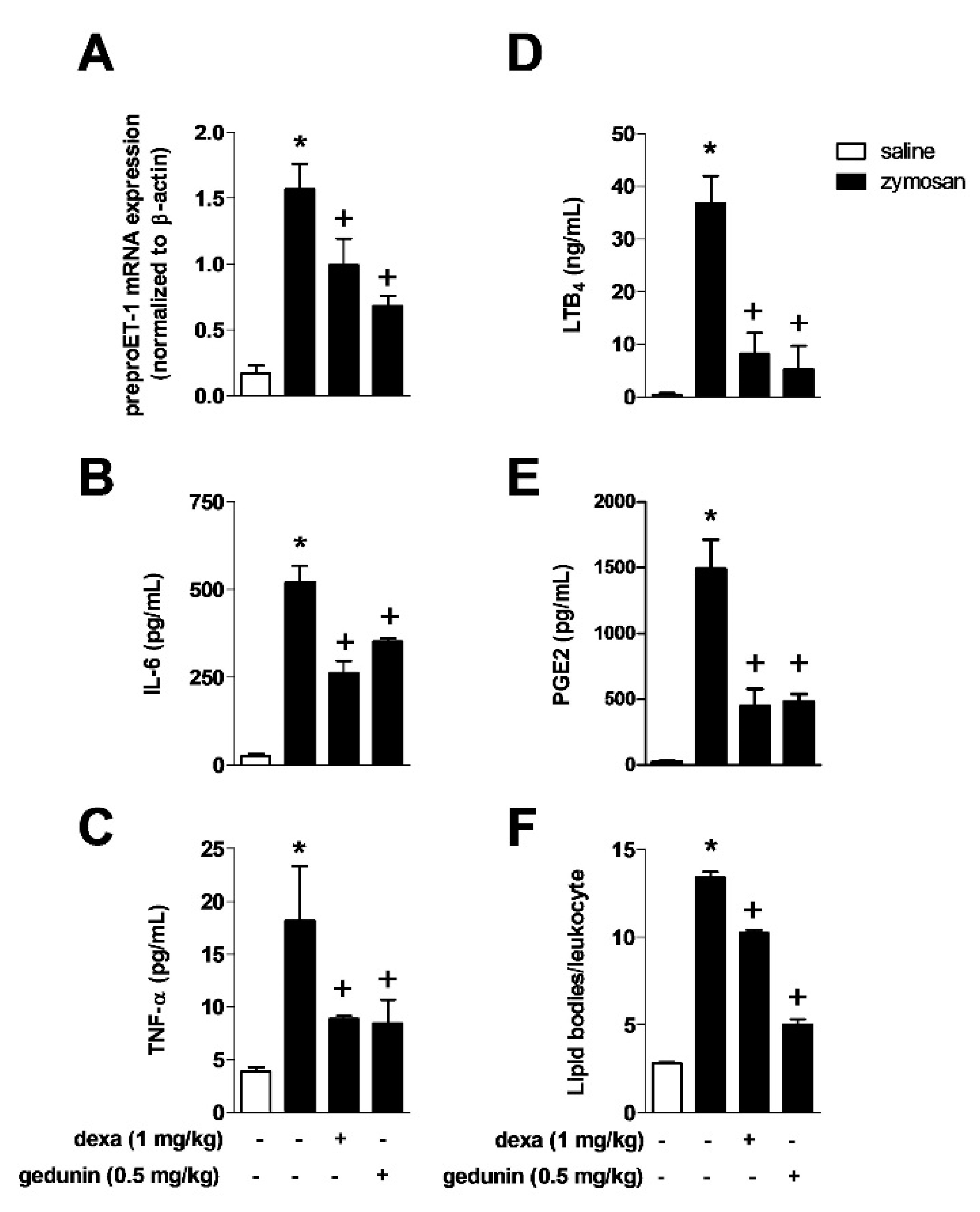
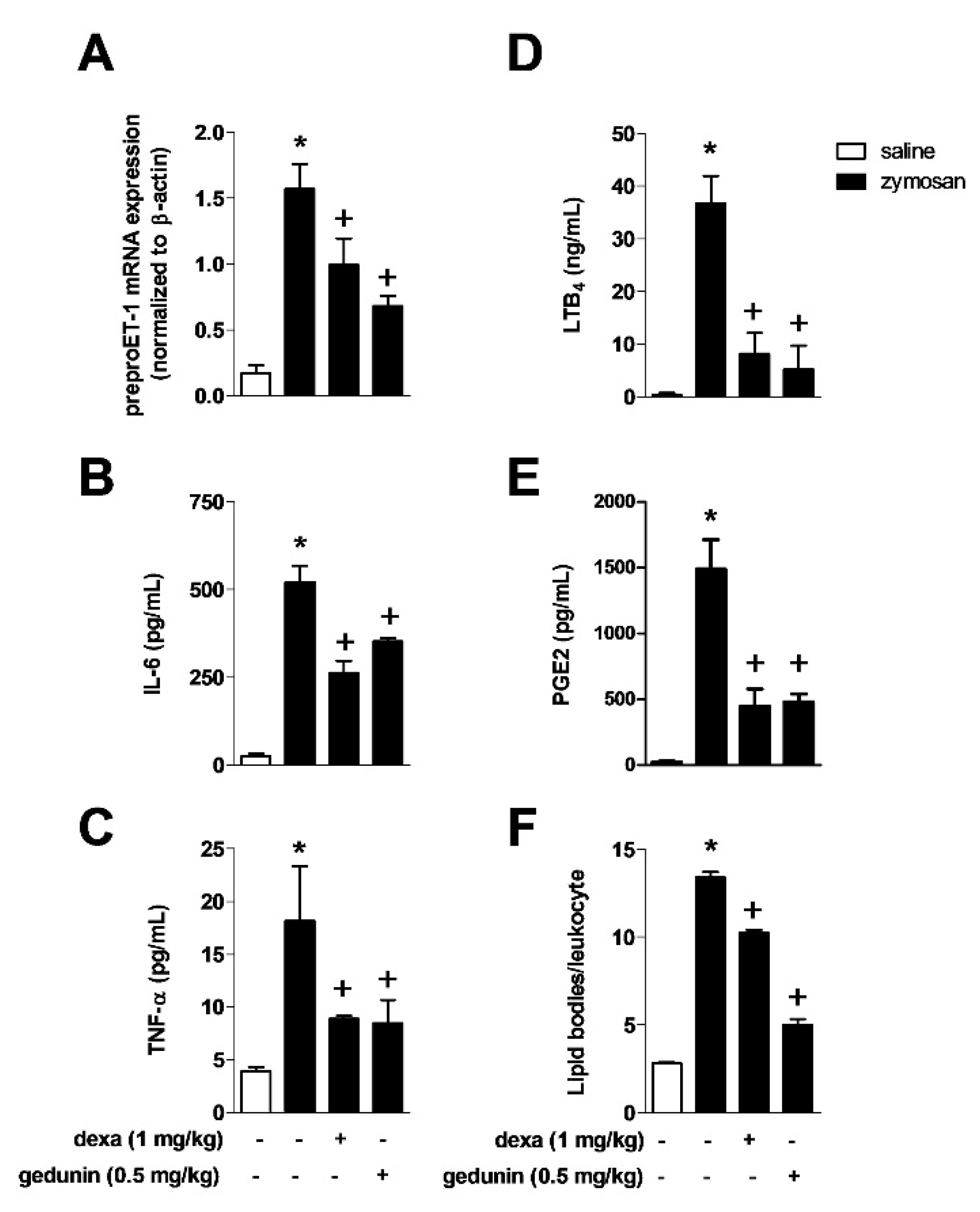
2.2. Gedunin Pre-Treatment Modulates Expression and Production of Inflammatory Mediators after Zymosan i.a. Stimulation
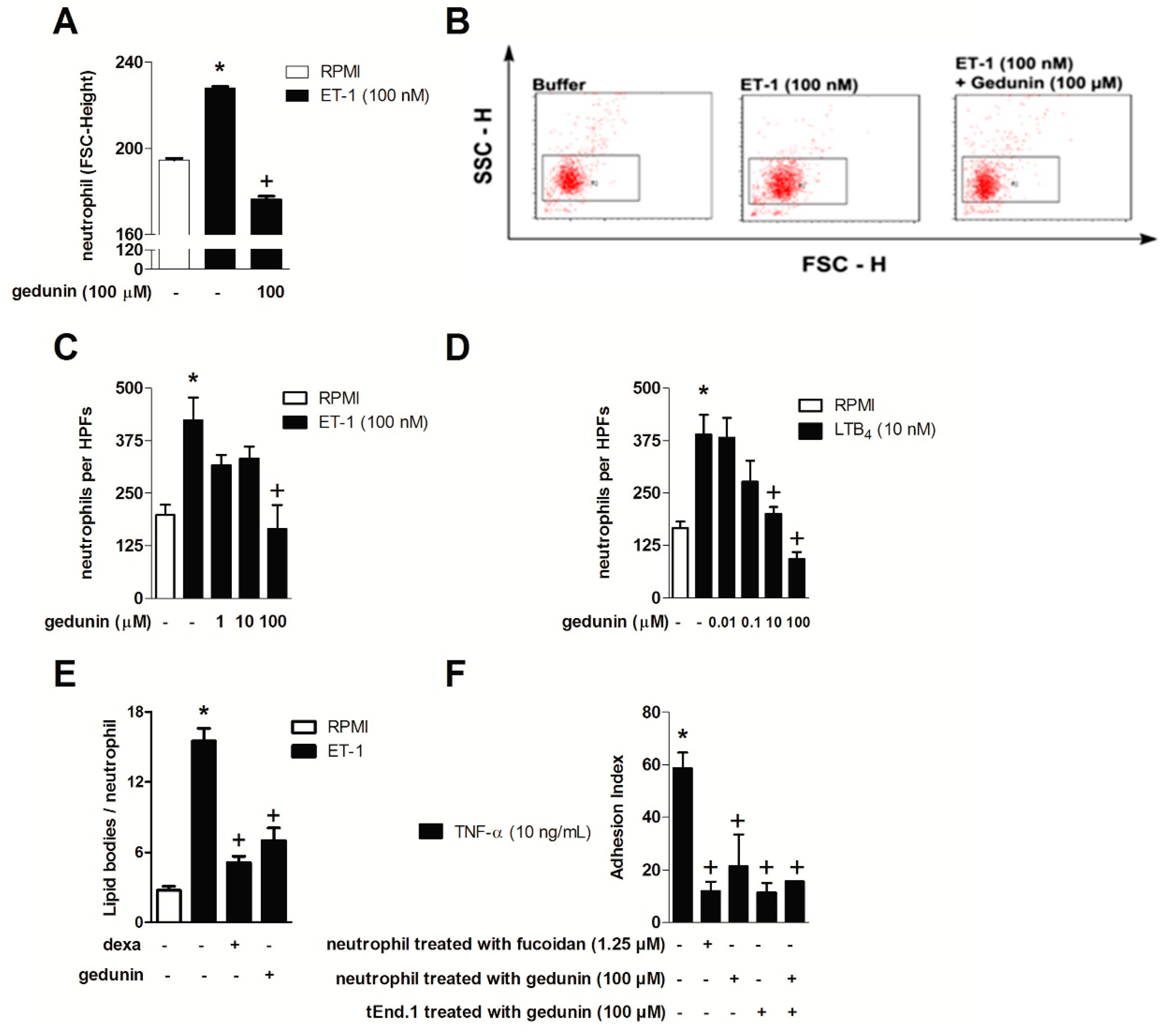
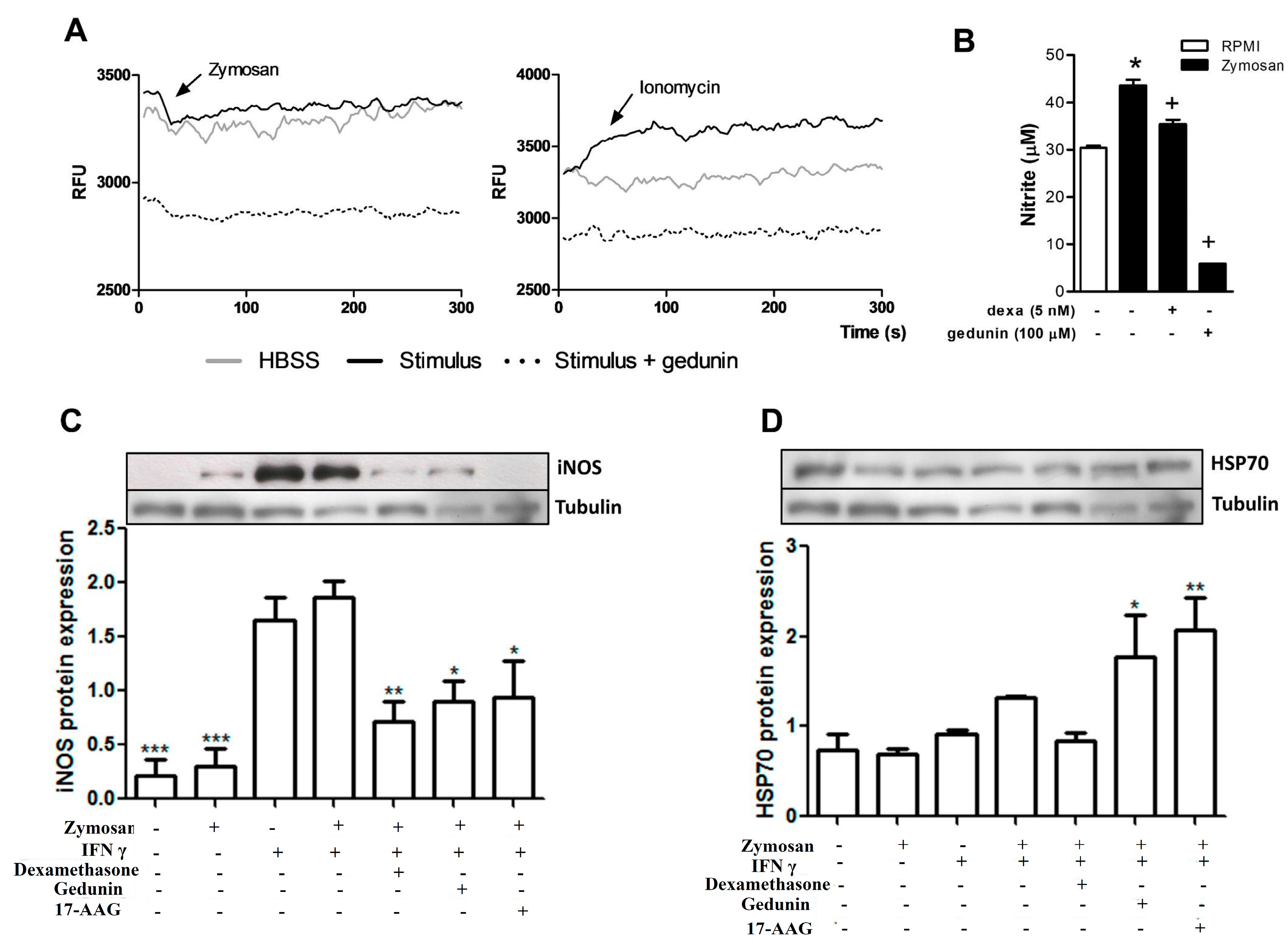
2.3. Gedunin Impairs Neutrophil Activation in Vitro


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Dose | Viability (%) |
|---|---|---|
| Gedunin (μM) | 1000 | 20 |
| 400 | 40 | |
| 200 | 100 | |
| 100 | 100 | |
| 50 | 100 | |
| 20 | 100 | |
| 10 | 100 | |
| 1 | 100 | |
| 0.1 | 100 | |
| DMSO | 50 | 100 |
| 5 | 100 | |
| 0.5 | 100 | |
| Tween 2% | - | 0 |
2.4. Gedunin Post-Treatment Impairs Zymosan-Induced Edema Formation and Neutrophil Influx to Inflamed Knee Joints
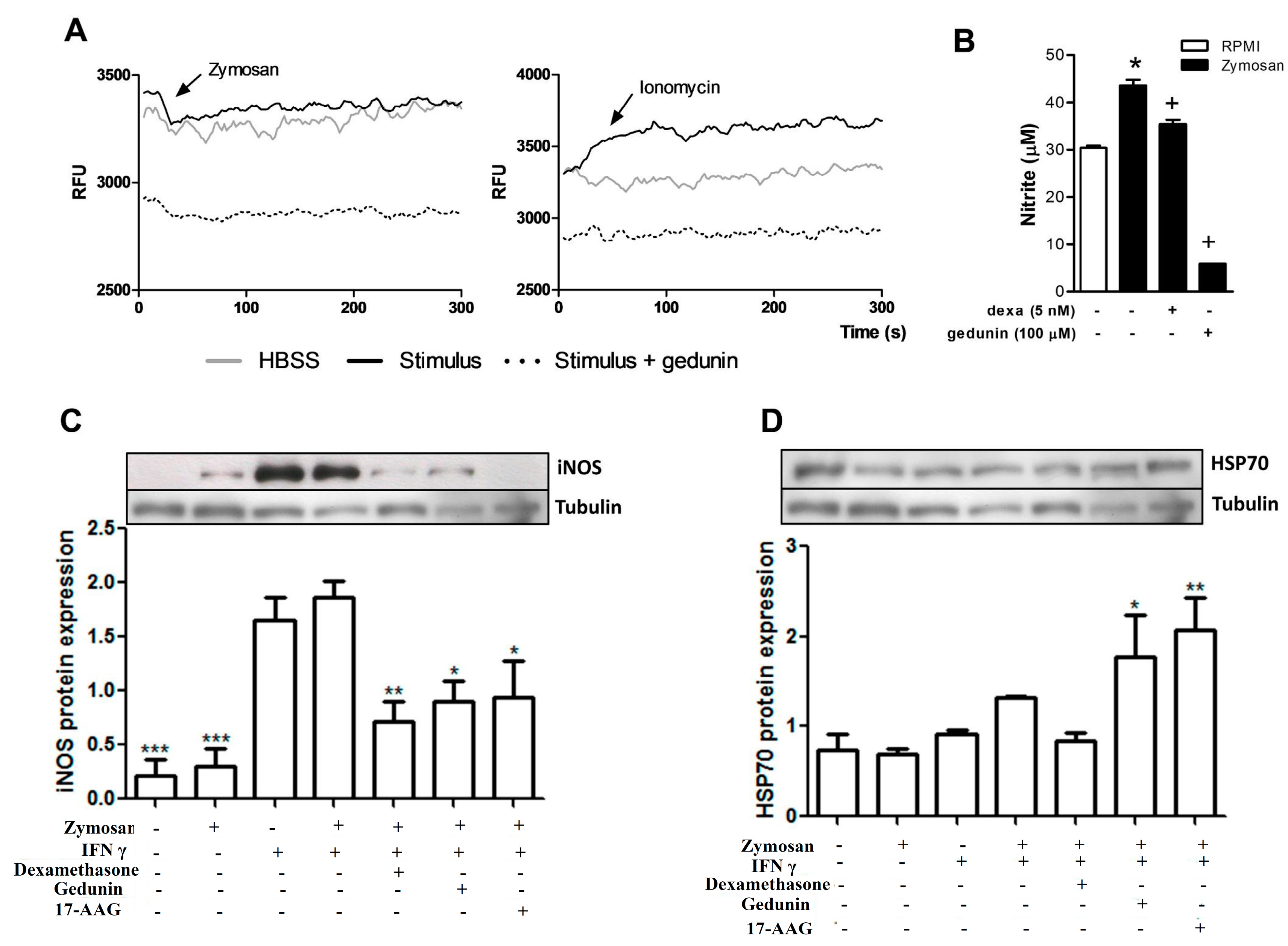
2.5. Gedunin Modulates Macrophage Activation in Vitro
3. Experimental Section
3.1. Animals
3.2. Induction of Acute Joint Inflammation
3.3. In Vivo Pretreatments
3.4. In Vivo Post-Treatments
3.5. Measurement of Knee Joint Swelling
3.6. Collection of Synovial Fluid and Leukocyte Counts
3.7. Histology
3.8. Evaluation of Articular Hypernociception
3.9. Real-Time RT–PCR for Evaluation of Preproet-1 Expression
3.10. Determination of Inflammatory Mediators in Synovial Washes
3.11. Lipid Body Induction and Staining
3.12. Shape Change Assay
3.13. Neutrophil Chemotaxis
3.14. Cell Adhesion Assay
3.15. Cytotoxicity Assay
3.16. Calcium Mobilization Assay
3.17. Determination of Nitrite Production
3.18. Western Blotting
3.19. Statistical Analysis
3.20. Drugs and Reagents
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sweeney, S.E.; Firestein, G.S. Rheumatoid arthritis: Regulation of synovial inflammation. Int. J. Biochem. Cell Biol. 2004, 36, 372–378. [Google Scholar]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar]
- Firestein, G.S. Pathogenesis of rheumatoid arthritis: How early is early? Arthritis Res. Ther. 2005, 7, 157–159. [Google Scholar]
- Gegout, P.; Gillet, P.; Chevrier, D.; Guingamp, C.; Terlain, B.; Netter, P. Characterization of zymosan-induced arthritis in the rat: Effects on joint inflammation and cartilage metabolism. Life Sci. 1994, 55, PL321–PL326. [Google Scholar]
- Keystone, E.C.; Schorlemmer, H.U.; Pope, C.; Allison, A.C. Zymosan-induced arthritis: A model of chronic proliferative arthritis following activation of the alternative pathway of complement. Arthritis Rheum. 1977, 20, 1396–1401. [Google Scholar]
- Conte, F.P.; Menezes-de-Lima, O.; Verri, W.A.; Cunha, F.Q.; Penido, C.; Henriques, M.G. Lipoxin A(4) attenuates zymosan-induced arthritis by modulating endothelin-1 and its effects. Br. J. Pharmacol. 2010, 161, 911–924. [Google Scholar]
- Penido, C.; Conte, F.P.; Chagas, M.S.; Rodrigues, C.A.; Pereira, J.F.; Henriques, M.G. Antiinflammatory effects of natural tetranortriterpenoids isolated from Carapa guianensis Aublet on zymosan-induced arthritis in mice. Inflamm. Res. 2006, 55, 457–464. [Google Scholar]
- Conte, F.E.P.; Barja-Fidalgo, C.; Verri, W.A.; Cunha, F.Q.; Rae, G.A.; Penido, C.; Henriques, M. Endothelins modulate inflammatory reaction in zymosan-induced arthritis: Participation of LTB4, TNF-alpha, and CXCL-1. J. Leukoc Biol. 2008, 84, 652–660. [Google Scholar]
- Eggleton, P.; Wang, L.; Penhallow, J.; Crawford, N.; Brown, K.A. Differences in oxidative response of subpopulations of neutrophils from healthy subjects and patients with rheumatoid arthritis. Ann. Rheum. Dis. 1995, 54, 916–923. [Google Scholar]
- Baici, A.; Salgam, P.; Cohen, G.; Fehr, K.; Böni, A. Action of collagenase and elastase from human polymorphonuclear leukocytes on human articular cartilage. Rheumatol. Int. 1982, 2, 11–16. [Google Scholar]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5. [Google Scholar] [CrossRef]
- Wright, H.L.; Moots, R.J.; Bucknall, R.C.; Edwards, S.W. Neutrophil function in inflammation and inflammatory diseases. Rheumatology (Oxf.) 2010, 49, 1618–1631. [Google Scholar]
- Wright, H.L.; Moots, R.J.; Edwards, S.W. The multifactorial role of neutrophils in rheumatoid arthritis. Nat. Rev. Rheumatol. 2014, 10, 593–601. [Google Scholar]
- Wipke, B.T.; Allen, P.M. Essential role of neutrophils in the initiation and progression of a murine model of rheumatoid arthritis. J. Immunol. 2001, 167, 1601–1608. [Google Scholar]
- Tanaka, D.; Kagari, T.; Doi, H.; Shimozato, T. Essential role of neutrophils in anti-type II collagen antibody and lipopolysaccharide-induced arthritis. Immunology 2006, 119, 195–202. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar]
- Patwardhan, C.A.; Fauq, A.; Peterson, L.B.; Miller, C.; Blagg, B.S.; Chadli, A. Gedunin inactivates the co-chaperone p23 protein causing cancer cell death by apoptosis. J. Biol. Chem. 2013, 288, 7313–7325. [Google Scholar]
- Wax, S.; Piecyk, M.; Maritim, B.; Anderson, P. Geldanamycin inhibits the production of inflammatory cytokines in activated macrophages by reducing the stability and translation of cytokine transcripts. Arthritis Rheum. 2003, 48, 541–550. [Google Scholar]
- Bucci, M.; Roviezzo, F.; Cicala, C.; Sessa, W.C.; Cirino, G. Geldanamycin, an inhibitor of heat shock protein 90 (Hsp90) mediated signal transduction has anti-inflammatory effects and interacts with glucocorticoid receptor in vivo. Br. J. Pharmacol. 2000, 131, 13–16. [Google Scholar]
- Madrigal-Matute, J.; López-Franco, O.; Blanco-Colio, L.M.; Muñoz-García, B.; Ramos-Mozo, P.; Ortega, L.; Egido, J.; Martín-Ventura, J.L. Heat shock protein 90 inhibitors attenuate inflammatory responses in atherosclerosis. Cardiovasc. Res. 2010, 86, 330–337. [Google Scholar]
- Ferraris, F.K.; Moret, K.H.; Figueiredo, A.B.; Penido, C.; Henriques, M. Gedunin, a natural tetranortriterpenoid, modulates T lymphocyte responses and ameliorates allergic inflammation. Int. Immunopharmacol. 2012, 14, 82–93. [Google Scholar]
- Poulaki, V.; Iliaki, E.; Mitsiades, N.; Mitsiades, C.S.; Paulus, Y.N.; Bula, D.V.; Gragoudas, E.S.; Miller, J.W. Inhibition of Hsp90 attenuates inflammation in endotoxin-induced uveitis. FASEB J. 2007, 21, 2113–2123. [Google Scholar]
- Penido, C.; Costa, K.A.; Pennaforte, R.J.; Costa, M.F.; Pereira, J.F.; Siani, A.C.; Henriques, M.G. Anti-allergic effects of natural tetranortriterpenoids isolated from Carapa guianensis Aublet on allergen-induced vascular permeability and hyperalgesia. Inflamm. Res. 2005, 54, 295–303. [Google Scholar]
- Carey, M.A.; Germolec, D.R.; Langenbach, R.; Zeldin, D.C. Cyclooxygenase enzymes in allergic inflammation and asthma. Prostaglandins Leukot. Essent. Fatty Acids 2003, 69, 157–162. [Google Scholar]
- Coelho, F.M.; Pinho, V.; Amaral, F.A.; Sachs, D.; Costa, V.V.; Rodrigues, D.H.; Vieira, A.T.; Silva, T.A.; Souza, D.G.; Bertini, R.; et al. The chemokine receptors CXCR1/CXCR2 modulate antigen-induced arthritis by regulating adhesion of neutrophils to the synovial microvasculature. Arthritis Rheum. 2008, 58, 2329–2337. [Google Scholar]
- Liu, H.; Pope, R.M. Phagocytes: Mechanisms of inflammation and tissue destruction. Rheum. Dis. Clin. N. Am. 2004, 30, 19–39. [Google Scholar]
- Edwards, S.W.; Hallett, M.B. Seeing the wood for the trees: The forgotten role of neutrophils in rheumatoid arthritis. Immunol. Today 1997, 18, 320–324. [Google Scholar]
- Paulino, E.C.; Steil, A.A.; Jancar, S. Effect of endothelins on human neutrophil activation by immune complexes. Int. Immunopharmacol. 2006, 6, 1119–1125. [Google Scholar]
- Verri, W.A.; Cunha, T.M.; Magro, D.A.; Guerrero, A.T.; Vieira, S.M.; Carregaro, V.; Souza, G.R.; Henriques, M.; Ferreira, S.H.; Cunha, F.Q. Targeting endothelin ETA and ETB receptors inhibits antigen-induced neutrophil migration and mechanical hypernociception in mice. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 379, 271–279. [Google Scholar]
- Donate, P.B.; Cunha, T.M.; Verri, W.A.; Junta, C.M.; Lima, F.O.; Vieira, S.M.; Peres, R.S.; Bombonato-Prado, K.F.; Louzada, P.; Ferreira, S.H.; et al. Bosentan, an endothelin receptor antagonist, ameliorates collagen-induced arthritis: The role of TNF-α in the induction of endothelin system genes. Inflamm. Res. 2012, 61, 337–348. [Google Scholar]
- Imhof, A.K.; Glück, L.; Gajda, M.; Bräuer, R.; Schaible, H.G.; Schulz, S. Potent anti-inflammatory and antinociceptive activity of the endothelin receptor antagonist bosentan in monoarthritic mice. Arthritis Res. Ther. 2011, 13, R97. [Google Scholar] [CrossRef]
- Buckland, J. Experimental arthritis: Endothelin blockade: A new therapy for inflammatory arthritis? Nat. Rev. Rheumatol. 2011, 7, 497. [Google Scholar] [CrossRef]
- Griffiths, R.J.; Smith, M.A.; Roach, M.L.; Stock, J.L.; Stam, E.J.; Milici, A.J.; Scampoli, D.N.; Eskra, J.D.; Byrum, R.S.; Koller, B.H.; et al. Collagen-induced arthritis is reduced in 5-lipoxygenase-activating protein-deficient mice. J. Exp. Med. 1997, 185, 1123–1129. [Google Scholar]
- Guerrero, A.T.; Verri, W.A.; Cunha, T.M.; Silva, T.A.; Schivo, I.R.; Dal-Secco, D.; Canetti, C.; Rocha, F.A.; Parada, C.A.; Cunha, F.Q.; et al. Involvement of LTB4 in zymosan-induced joint nociception in mice: Participation of neutrophils and PGE2. J. Leukoc. Biol. 2008, 83, 122–130. [Google Scholar]
- Davidson, E.M.; Rae, S.A.; Smith, M.J. Leukotriene B4, a mediator of inflammation present in synovial fluid in rheumatoid arthritis. Ann. Rheum. Dis. 1983, 42, 677–679. [Google Scholar]
- Gürsel, T.; Firat, S.; Ercan, Z.S. Increased serum leukotriene B4 level in the active stage of rheumatoid arthritis in children. Prostaglandins Leukot. Essent. Fatty Acids 1997, 56, 205–207. [Google Scholar]
- Canetti, C.A.; Leung, B.P.; Culshaw, S.; McInnes, I.B.; Cunha, F.Q.; Liew, F.Y.; Cannetti, C.A. IL-18 enhances collagen-induced arthritis by recruiting neutrophils via TNF-alpha and leukotriene B4. J. Immunol. 2003, 171, 1009–1015. [Google Scholar]
- Grespan, R.; Fukada, S.Y.; Lemos, H.P.; Vieira, S.M.; Napimoga, M.H.; Teixeira, M.M.; Fraser, A.R.; Liew, F.Y.; McInnes, I.B.; Cunha, F.Q. CXCR2-specific chemokines mediate leukotriene B4-dependent recruitment of neutrophils to inflamed joints in mice with antigen-induced arthritis. Arthritis Rheum. 2008, 58, 2030–2040. [Google Scholar]
- Carreira, E.U.; Carregaro, V.; Teixeira, M.M.; Moriconi, A.; Aramini, A.; Verri, W.A.; Ferreira, S.H.; Cunha, F.Q.; Cunha, T.M. Neutrophils recruited by CXCR1/2 signalling mediate post-incisional pain. Eur. J. Pain 2013, 17, 654–663. [Google Scholar]
- Pinto, L.G.; Cunha, T.M.; Vieira, S.M.; Lemos, H.P.; Verri, W.A.; Cunha, F.Q.; Ferreira, S.H. IL-17 mediates articular hypernociception in antigen-induced arthritis in mice. Pain 2010, 148, 247–256. [Google Scholar]
- Cunha, T.M.; Verri, W.A.; Schivo, I.R.; Napimoga, M.H.; Parada, C.A.; Poole, S.; Teixeira, M.M.; Ferreira, S.H.; Cunha, F.Q. Crucial role of neutrophils in the development of mechanical inflammatory hypernociception. J. Leukoc. Biol. 2008, 83, 824–832. [Google Scholar]
- Cunha, T.M.; Barsante, M.M.; Guerrero, A.T.; Verri, W.A.; Ferreira, S.H.; Coelho, F.M.; Bertini, R.; di Giacinto, C.; Allegretti, M.; Cunha, F.Q.; et al. Treatment with DF 2162, a non-competitive allosteric inhibitor of CXCR1/2, diminishes neutrophil influx and inflammatory hypernociception in mice. Br. J. Pharmacol. 2008, 154, 460–470. [Google Scholar]
- Lavich, T.R.; Siqueira, R.E.A.; Farias-Filho, F.A.; Cordeiro, R.S.; Rodrigues e Silva, P.M.; Martins, M.A. Neutrophil infiltration is implicated in the sustained thermal hyperalgesic response evoked by allergen provocation in actively sensitized rats. Pain 2006, 125, 180–187. [Google Scholar]
- Ting, E.; Guerrero, A.T.; Cunha, T.M.; Verri, W.A.; Taylor, S.M.; Woodruff, T.M.; Cunha, F.Q.; Ferreira, S.H. Role of complement C5a in mechanical inflammatory hypernociception: Potential use of C5a receptor antagonists to control inflammatory pain. Br. J. Pharmacol. 2008, 153, 1043–1053. [Google Scholar]
- Brennan, F.M.; McInnes, I.B. Evidence that cytokines play a role in rheumatoid arthritis. J. Clin. Investig. 2008, 118, 3537–3545. [Google Scholar]
- Willrich, M.A.; Murray, D.L.; Snyder, M.R. Tumor necrosis factor inhibitors: Clinical utility in autoimmune diseases. Transl. Res. 2014, 165, 270–282. [Google Scholar]
- Karampetsou, M.P.; Liossis, S.N.; Sfikakis, P.P. TNF-α antagonists beyond approved indications: Stories of success and prospects for the future. QJM 2010, 103, 917–928. [Google Scholar]
- Feldmann, M.; Maini, R.N. Anti-TNF alpha therapy of rheumatoid arthritis: What have we learned? Annu. Rev. Immunol. 2001, 19, 163–196. [Google Scholar]
- Ferraccioli, G.; Bracci-Laudiero, L.; Alivernini, S.; Gremese, E.; Tolusso, B.; de Benedetti, F. Interleukin-1β and interleukin-6 in arthritis animal models: Roles in the early phase of transition from acute to chronic inflammation and relevance for human rheumatoid arthritis. Mol. Med. 2010, 16, 552–557. [Google Scholar]
- Park, J.Y.; Pillinger, M.H. Interleukin-6 in the pathogenesis of rheumatoid arthritis. Bull. NYU Hosp. Jt. Dis. 2007, 65 (Suppl. 1), S4–S10. [Google Scholar]
- Boe, A.; Baiocchi, M.; Carbonatto, M.; Papoian, R.; Serlupi-Crescenzi, O. Interleukin 6 knock-out mice are resistant to antigen-induced experimental arthritis. Cytokine 1999, 11, 1057–1064. [Google Scholar]
- Genovese, M.C.; Fleischmann, R.; Furst, D.; Janssen, N.; Carter, J.; Dasgupta, B.; Bryson, J.; Duncan, B.; Zhu, W.; Pitzalis, C.; et al. Efficacy and safety of olokizumab in patients with rheumatoid arthritis with an inadequate response to TNF inhibitor therapy: Outcomes of a randomised Phase IIb study. Ann. Rheum. Dis. 2014, 73, 1607–1615. [Google Scholar]
- Huizinga, T.W.; Fleischmann, R.M.; Jasson, M.; Radin, A.R.; van Adelsberg, J.; Fiore, S.; Huang, X.; Yancopoulos, G.D.; Stahl, N.; Genovese, M.C. Sarilumab, a fully human monoclonal antibody against IL-6Rα in patients with rheumatoid arthritis and an inadequate response to methotrexate: Efficacy and safety results from the randomised SARIL-RA-MOBILITY Part A trial. Ann. Rheum. Dis. 2014, 73, 1626–1634. [Google Scholar]
- Smolen, J.S.; Beaulieu, A.; Rubbert-Roth, A.; Ramos-Remus, C.; Rovensky, J.; Alecock, E.; Woodworth, T.; Alten, R.; Investigators, O. Effect of interleukin-6 receptor inhibition with tocilizumab in patients with rheumatoid arthritis (OPTION study): A double-blind, placebo-controlled, randomised trial. Lancet 2008, 371, 987–997. [Google Scholar]
- Nishimoto, N.; Kishimoto, T. Interleukin 6: From bench to bedside. Nat. Clin. Pract. Rheumatol. 2006, 2, 619–626. [Google Scholar]
- Mathis, S.P.; Jala, V.R.; Lee, D.M.; Haribabu, B. Nonredundant roles for leukotriene B4 receptors BLT1 and BLT2 in inflammatory arthritis. J. Immunol. 2010, 185, 3049–3056. [Google Scholar]
- Mencarelli, M.; Pecorelli, A.; Carbotti, P.; Valacchi, G.; Grasso, G.; Muscettola, M. Endothelin receptor A expression in human inflammatory cells. Regul. Pept. 2009, 158, 1–5. [Google Scholar]
- Toffoli, M.C.; Gabra, B.H.; Teixeira, C.F.; Sirois, P.; Jancar, S. Endothelins mediate neutrophil activation, ProMMP-9 release and endothelial cell detachment. Inflammation 2007, 30, 28–37. [Google Scholar]
- Bozza, P.T.; Bakker-Abreu, I.; Navarro-Xavier, R.A.; Bandeira-Melo, C. Lipid body function in eicosanoid synthesis: An update. Prostaglandins Leukot. Essent. Fatty Acids 2011, 85, 205–213. [Google Scholar]
- Brandt, G.E.; Schmidt, M.D.; Prisinzano, T.E.; Blagg, B.S. Gedunin, a novel hsp90 inhibitor: Semisynthesis of derivatives and preliminary structure-activity relationships. J. Med. Chem. 2008, 51, 6495–6502. [Google Scholar]
- Zhang, B.; Au, Q.; Yoon, I.S.; Tremblay, M.H.; Yip, G.; Zhou, Y.; Barber, J.R.; Ng, S.C. Identification of small-molecule HSF1 amplifiers by high content screening in protection of cells from stress induced injury. Biochem. Biophys. Res. Commun. 2009, 390, 925–930. [Google Scholar]
- Kim, N.; Kim, J.Y.; Yenari, M.A. Anti-inflammatory properties and pharmacological induction of Hsp70 after brain injury. Inflammopharmacology 2012, 20, 177–185. [Google Scholar]
- Jones, Q.; Voegeli, T.S.; Li, G.; Chen, Y.; Currie, R.W. Heat shock proteins protect against ischemia and inflammation through multiple mechanisms. Inflamm. Allergy Drug Targets 2011, 10, 247–259. [Google Scholar]
- Schett, G.; Redlich, K.; Xu, Q.; Bizan, P.; Gröger, M.; Tohidast-Akrad, M.; Kiener, H.; Smolen, J.; Steiner, G. Enhanced expression of heat shock protein 70 (hsp70) and heat shock factor 1 (HSF1) activation in rheumatoid arthritis synovial tissue. Differential regulation of hsp70 expression and hsf1 activation in synovial fibroblasts by proinflammatory cytokines, shear stress, and antiinflammatory drugs. J. Clin. Investig. 1998, 102, 302–311. [Google Scholar]
- Marković, M.; Majkić-Singh, N.; Ignjatović, S. Beneficial effects of cellular stress response in traditional spa treatment of rheumatoid arthritis. Clin. Lab. 2009, 55, 235–241. [Google Scholar]
- Echeverría, P.C.; Bernthaler, A.; Dupuis, P.; Mayer, B.; Picard, D. An interaction network predicted from public data as a discovery tool: Application to the Hsp90 molecular chaperone machine. PLoS One 2011, 6, e26044. [Google Scholar]
- Chen, G.; Cao, P.; Goeddel, D.V. TNF-induced recruitment and activation of the IKK complex require Cdc37 and Hsp90. Mol. Cell 2002, 9, 401–410. [Google Scholar]
- Lewis, J.; Devin, A.; Miller, A.; Lin, Y.; Rodriguez, Y.; Neckers, L.; Liu, Z.G. Disruption of hsp90 function results in degradation of the death domain kinase, receptor-interacting protein (RIP), and blockage of tumor necrosis factor-induced nuclear factor-kappaB activation. J. Biol. Chem. 2000, 275, 10519–10526. [Google Scholar]
- Tsatsanis, C.; Androulidaki, A.; Venihaki, M.; Margioris, A.N. Signalling networks regulating cyclooxygenase-2. Int. J. Biochem. Cell Biol. 2006, 38, 1654–1661. [Google Scholar]
- Ramsay, R.G.; Ciznadija, D.; Vanevski, M.; Mantamadiotis, T. Transcriptional regulation of cyclo-oxygenase expression: Three pillars of control. Int. J. Immunopathol. Pharmacol. 2003, 16 (Suppl. 2), S59–S67. [Google Scholar]
- Frasnelli, M.E.; Tarussio, D.; Chobaz-Péclat, V.; Busso, N.; So, A. TLR2 modulates inflammation in zymosan-induced arthritis in mice. Arthritis Res. Ther. 2005, 7, R370–R379. [Google Scholar]
- Lotito, A.P.; Muscará, M.N.; Kiss, M.H.; Teixeira, S.A.; Novaes, G.S.; Laurindo, I.M.; Silva, C.A.; Mello, S.B. Nitric oxide-derived species in synovial fluid from patients with juvenile idiopathic arthritis. J. Rheumatol. 2004, 31, 992–997. [Google Scholar]
- Grabowski, P.S.; Macpherson, H.; Ralston, S.H. Nitric oxide production in cells derived from the human joint. Br. J. Rheumatol. 1996, 35, 207–212. [Google Scholar]
- Sakurai, H.; Kohsaka, H.; Liu, M.F.; Higashiyama, H.; Hirata, Y.; Kanno, K.; Saito, I.; Miyasaka, N. Nitric oxide production and inducible nitric oxide synthase expression in inflammatory arthritides. J. Clin. Investig. 1995, 96, 2357–2363. [Google Scholar]
- Taskiran, D.; Stefanovic-Racic, M.; Georgescu, H.; Evans, C. Nitric oxide mediates suppression of cartilage proteoglycan synthesis by interleukin-1. Biochem. Biophys. Res. Commun. 1994, 200, 142–148. [Google Scholar]
- Kaur, H.; Halliwell, B. Evidence for nitric oxide-mediated oxidative damage in chronic inflammation. Nitrotyrosine in serum and synovial fluid from rheumatoid patients. FEBS Lett. 1994, 350, 9–12. [Google Scholar]
- Siebelt, M.; Jahr, H.; Groen, H.C.; Sandker, M.; Waarsing, J.H.; Kops, N.; Müller, C.; van Eden, W.; de Jong, M.; Weinans, H. Hsp90 inhibition protects against biomechanically induced osteoarthritis in rats. Arthritis Rheum. 2013, 65, 2102–2112. [Google Scholar]
- Verri, W.A.; Guerrero, A.T.; Fukada, S.Y.; Valerio, D.A.; Cunha, T.M.; Xu, D.; Ferreira, S.H.; Liew, F.Y.; Cunha, F.Q. IL-33 mediates antigen-induced cutaneous and articular hypernociception in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 2723–2728. [Google Scholar]
- Ferraris, F.K.; Rodrigues, R.; da Silva, V.P.; Figueiredo, R.; Penido, C.; Henriques, M. Modulation of T lymphocyte and eosinophil functions in vitro by natural tetranortriterpenoids isolated from Carapa guianensis Aublet. Int. Immunopharmacol. 2011, 11, 1–11. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conte, F.P.; Ferraris, F.K.; Costa, T.E.M.M.; Pacheco, P.; Seito, L.N.; Verri, W.A., Jr.; Cunha, F.Q.; Penido, C.; Henriques, M.G. Effect of Gedunin on Acute Articular Inflammation and Hypernociception in Mice. Molecules 2015, 20, 2636-2657. https://doi.org/10.3390/molecules20022636
Conte FP, Ferraris FK, Costa TEMM, Pacheco P, Seito LN, Verri WA Jr., Cunha FQ, Penido C, Henriques MG. Effect of Gedunin on Acute Articular Inflammation and Hypernociception in Mice. Molecules. 2015; 20(2):2636-2657. https://doi.org/10.3390/molecules20022636
Chicago/Turabian StyleConte, Fernando P., Fausto K. Ferraris, Thadeu E. M. M. Costa, Patricia Pacheco, Leonardo N. Seito, Waldiceu A. Verri, Jr., Fernando Q. Cunha, Carmen Penido, and Maria G. Henriques. 2015. "Effect of Gedunin on Acute Articular Inflammation and Hypernociception in Mice" Molecules 20, no. 2: 2636-2657. https://doi.org/10.3390/molecules20022636
APA StyleConte, F. P., Ferraris, F. K., Costa, T. E. M. M., Pacheco, P., Seito, L. N., Verri, W. A., Jr., Cunha, F. Q., Penido, C., & Henriques, M. G. (2015). Effect of Gedunin on Acute Articular Inflammation and Hypernociception in Mice. Molecules, 20(2), 2636-2657. https://doi.org/10.3390/molecules20022636