
Human Lectins and Their Roles in Viral Infections

Abstract
:1. Introduction
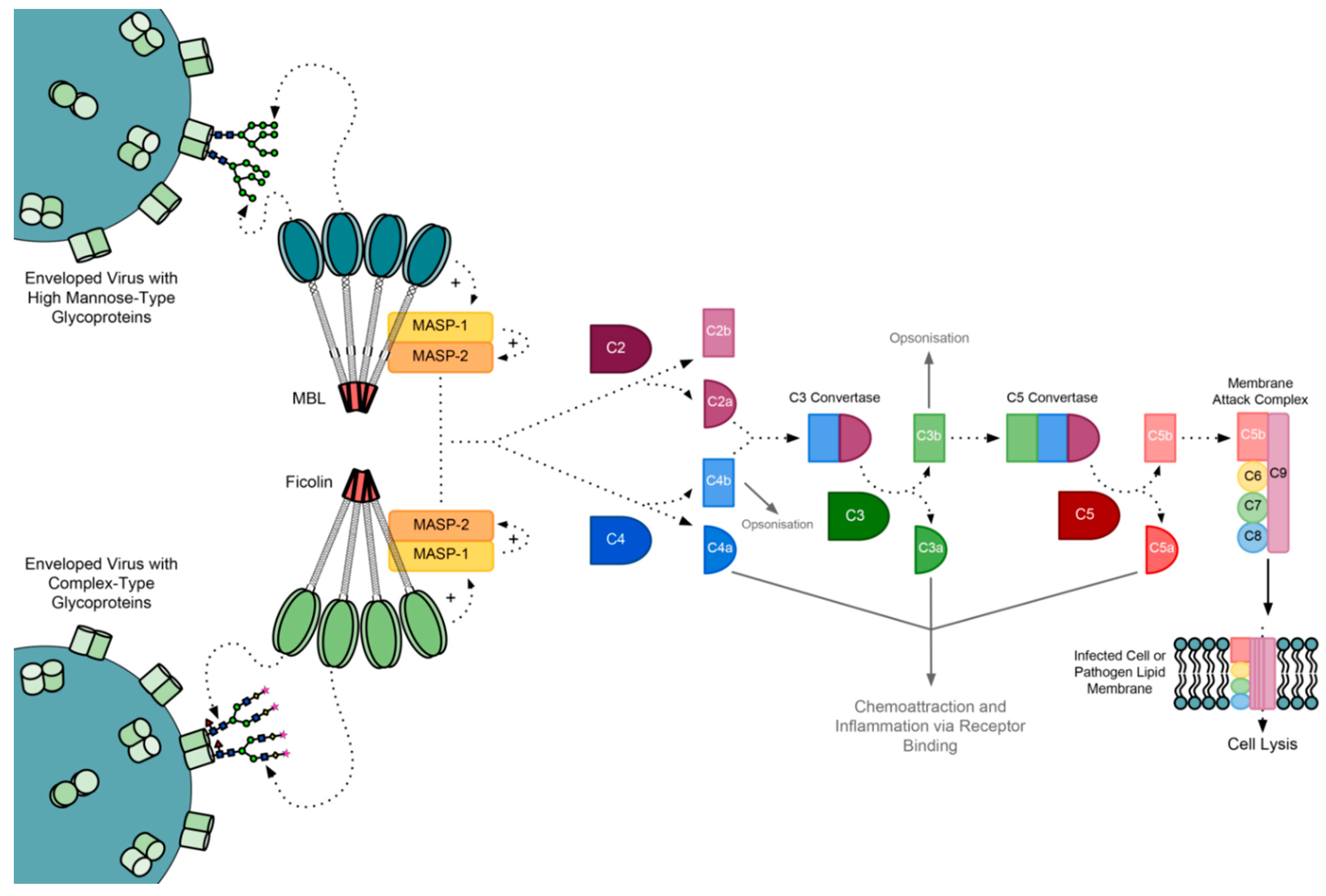
2. The Complement Cascade
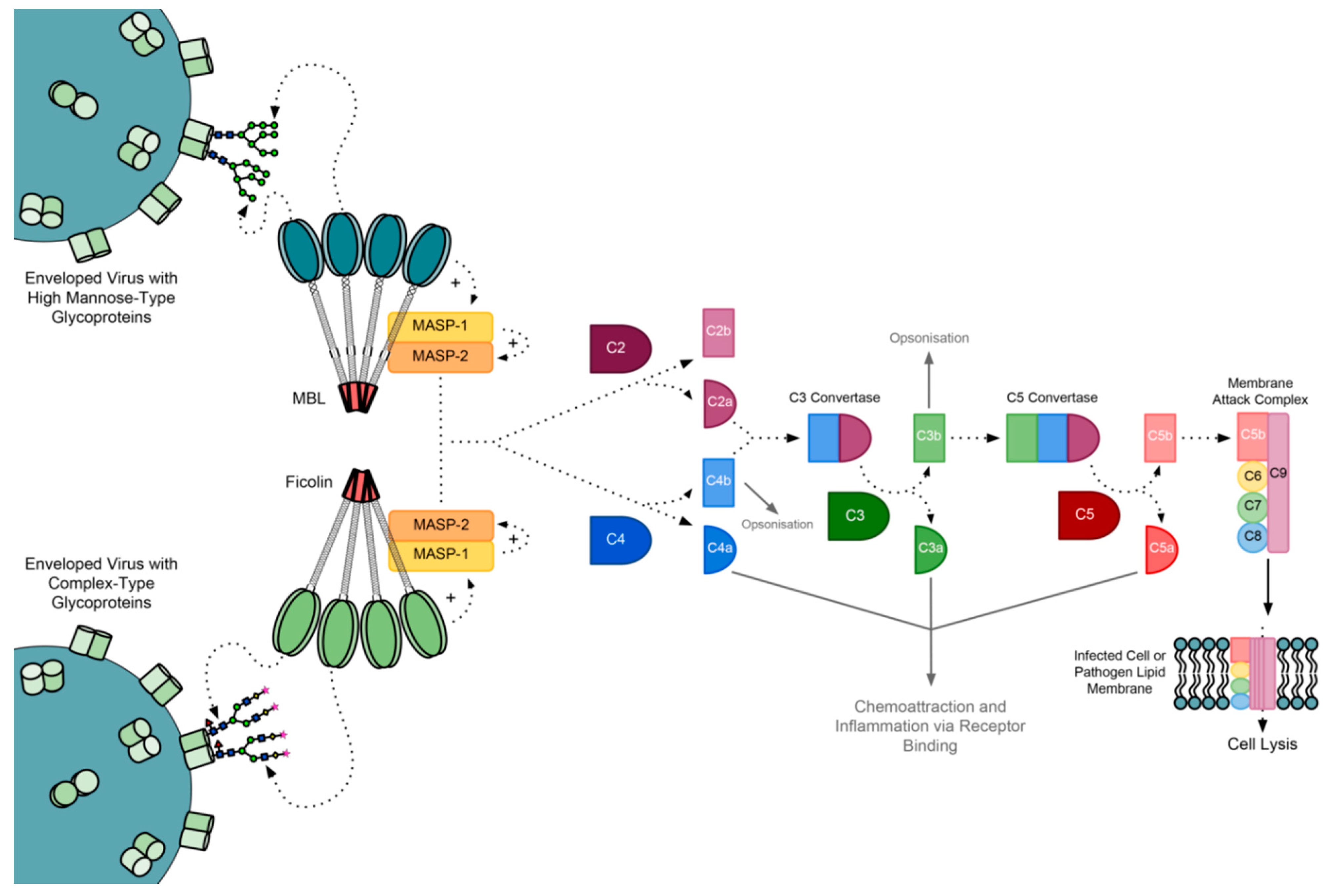
The MBL-Associated Serine Proteases
3. Mannose-Binding Lectin (MBL)
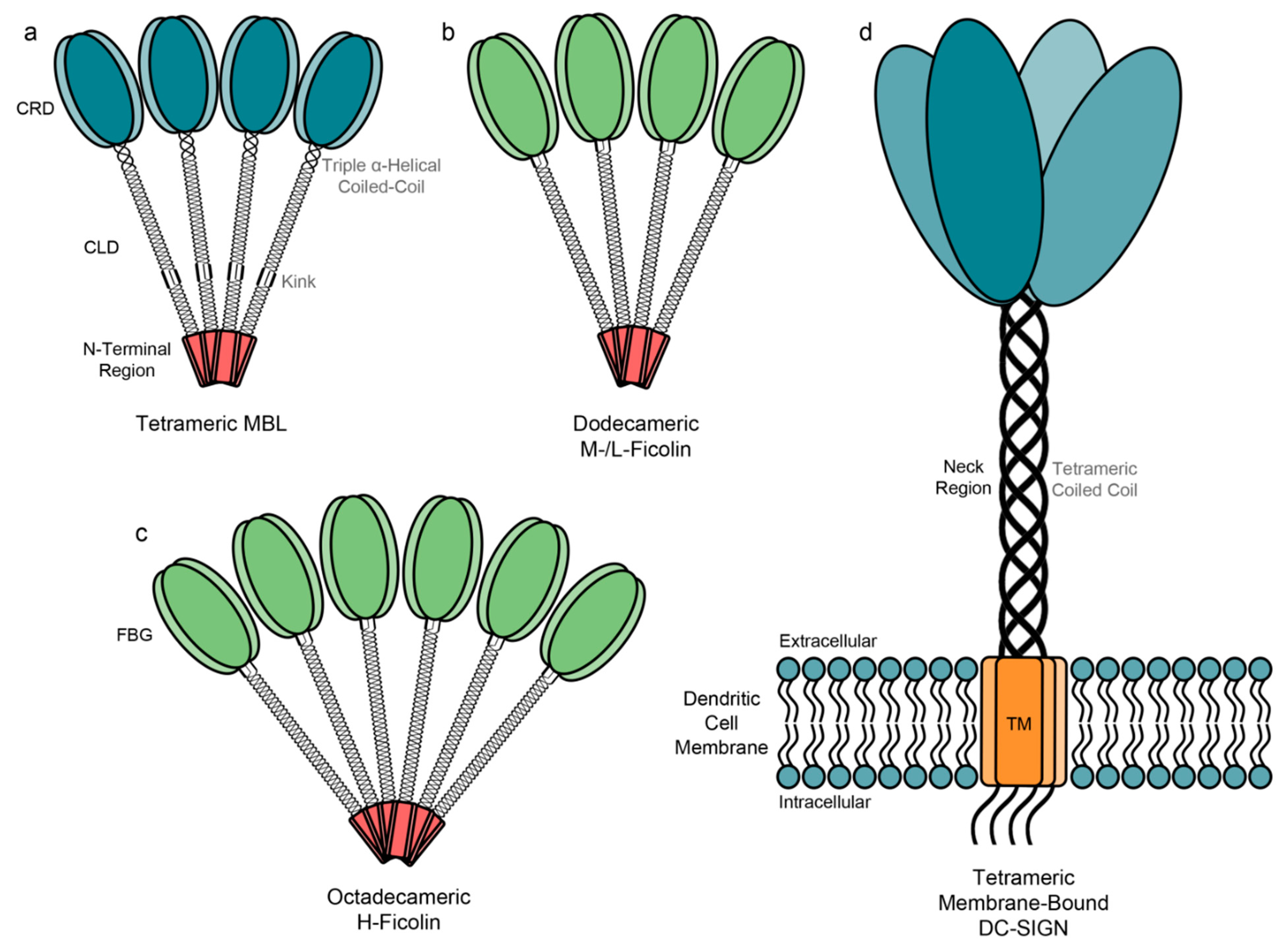
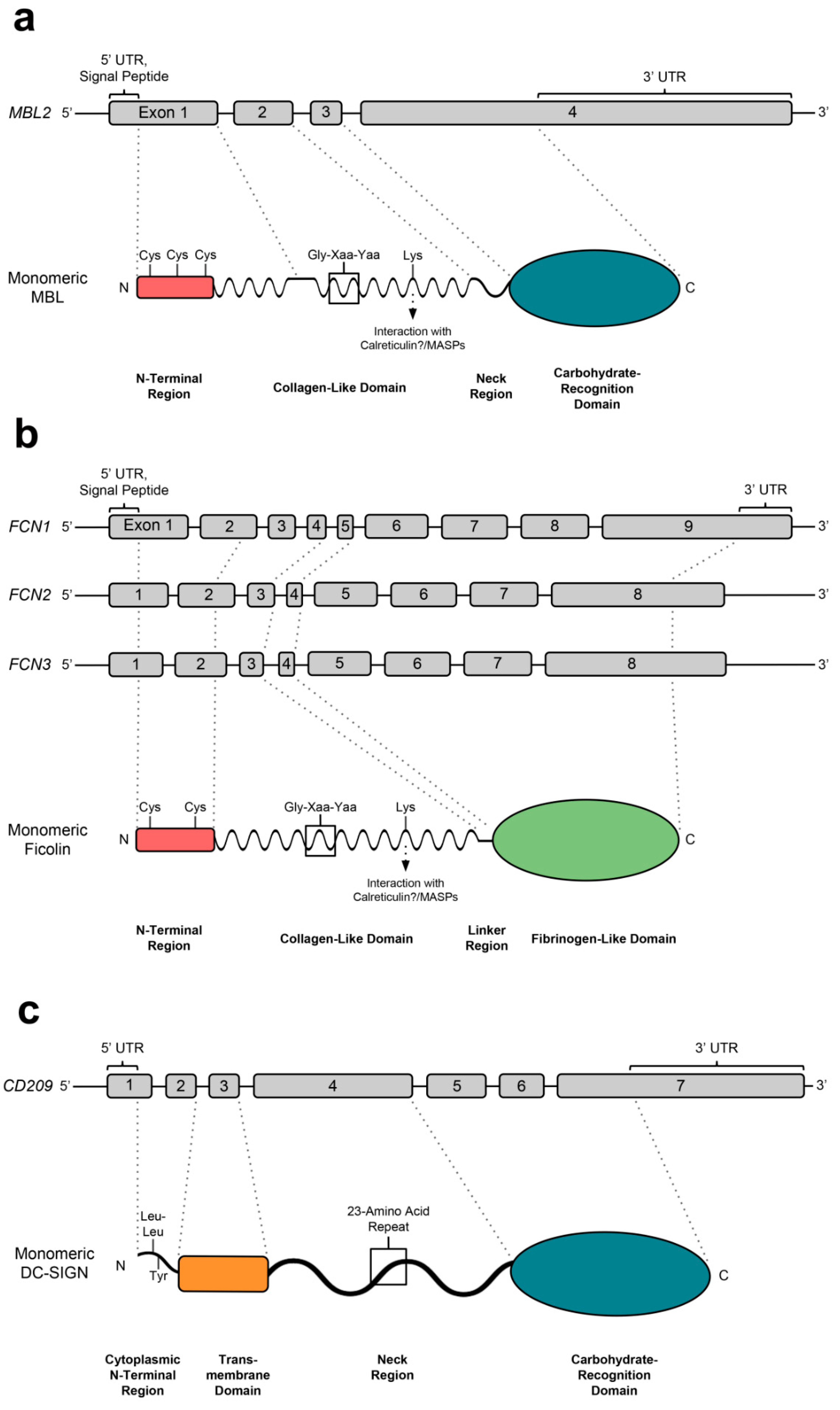
3.1. Genetics, Structure, Expression and Binding Specificities of MBL

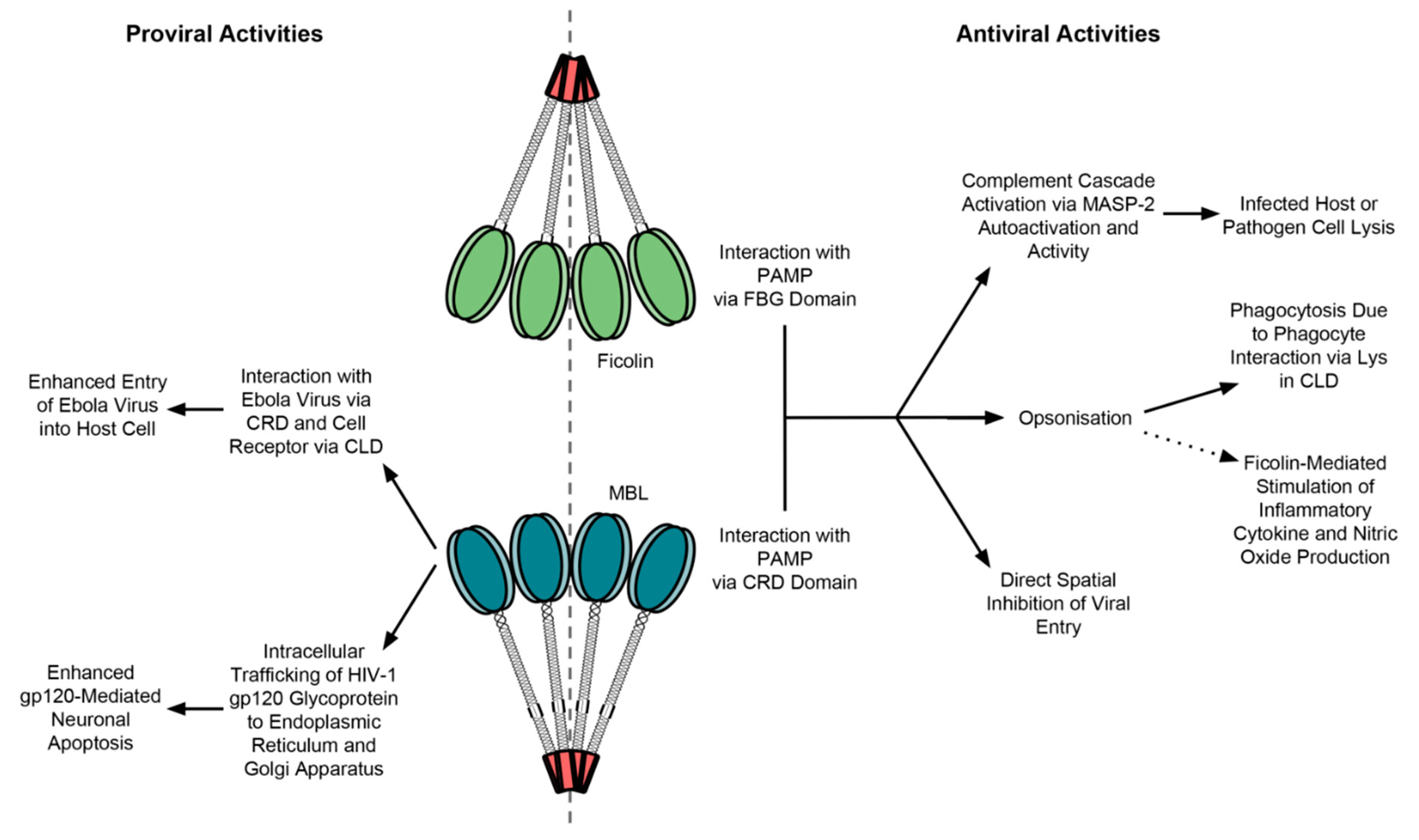
3.2. MBL Interaction with Viruses
Viral Exploitation of MBL
3.3. MBL Variants
The Effect of MBL Variant Alleles on Viral Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | dbSNP (Alternative Name) | Nucleotide Position | Major Allele | Minor Allele | Region | Amino Acid Mutation | Relevance to Specific Virus Infections |
|---|---|---|---|---|---|---|---|
| MBL2 | rs11003125 (H/L) | −550 | G | C | Promoter | - | HIV [74,75] |
| rs7096206 (X/Y) | −221 | C | G | Promoter | - | HBV [81], HCV [82,83], HIV [72,75], HTLV [84], SARS-CoV [48] | |
| rs7095891 (P/Q) | +4 | C | T | 5’ UTR | - | ||
| rs5030737 (MBL-D) | +223 | C | T | Exon 1 | Arg52Cys | CMV [85,86], DV[50], HBV [87,88], HCV [82,83,89,90], HIV [70,72,80], | |
| rs1800450 (MBL-B) | +230 | G | A | Exon 1 | Gly54Asp | CMV [85,86], DV [50], HBV [81,88,91], HCV [82,83,89,90], HIV [70,72,73,80], HTLV [92], SARS-CoV [48] | |
| rs1800451 (MBL-C) | +239 | G | A | Exon 1 | Gly57Glu | CMV [85,86], DV [50], HBV [88], HCV [82,83,89,90], HIV [70,72,80] | |
| FCN1 | rs2989727 | −1981 | G | A | Promoter | - | |
| rs10120023 | −542 | G | A | Promoter | - | ||
| rs28909976 | −271 | - | InsT | Promoter | - | ||
| rs10117466 | −144 | C | A | Promoter | - | Increased serum concentration [93] | |
| rs10441778 | +1435 | G | A | Exon 2 | Gly43Asp | Likely affects structure and oligomerisation [94] | |
| ss76901539 | +3458 | G | A | Exon 4 | Arg93Gln | Likely affects structure and oligomerisation [94] | |
| rs148649884 | +6658 | G | A | Exon 8 | Ala218Thr | Reduced serum concentration, reduced ligand binding [93] | |
| rs150625869 | +7895 | T | C | Exon 9 | Ser268Pro | Abolished serum concentration [93] | |
| rs1071583 | +7918 | G | A | Exon 9 | - | ||
| ss76901546 | +7929 | G | A | Exon 9 | Trp279STOP | Likely affects structure and oligomerisation [94] | |
| rs138055828 | +7959 | A | G | Exon 9 | Ala289Ser | Reduced serum concentration, reduced ligand binding [93] | |
| ss76901547 | +8000 | G | A | Exon 9 | Gly303Ser | Likely affects function [94] | |
| FCN2 | rs3124952 | −986 | G | A | Promoter | - | Reduced serum concentration [94]; HBV [95] |
| rs3124953 | −602 | G | A | Promoter | - | Increased serum concentration [94]; HBV [95] | |
| rs17514136 | −4 | A | G | Promoter | - | Increased serum concentration [94]; HBV [95] | |
| ss76901565 | +4423 | C | T | Exon 5 | Arg103Cys | Likely affects chemical and structural properties [94] | |
| ss76901566 | +4526 | C | T | Exon 5 | Thr137Met | Likely affects chemical and structural properties [94] | |
| ss76901570 | +4957 | G | A | Exon 6 | Arg147Gln | Likely affects ligand binding [94] | |
| ss76901571 | +4987 | G | A | Exon 6 | Arg157Gln | Likely affects ligand binding [94] | |
| rs17549193 | +6359 | C | T | Exon 8 | Thr236Met | Reduced binding to GlcNAc [94] and PTX3 [96]; | |
| rs7851696 | +6424 | G | T | Exon 8 | Ala258Ser | Increased binding to GlcNAc [94]; CMV [86], HBV [95] | |
| rs28357091 | +6443_44 | CT | A | Exon 8 | Ala264fs | Truncated protein [94] | |
| FCN3 | rs28357092 | +1637 | C | delC | Exon 5 | Leu117fs | Truncated protein [94]; Severe, recurrent respiratory and gastrointestinal infections [97,98,99] |
| ss76901551 | +1663 | A | G | Exon 5 | Thr125Ala | Likely affects function [94] | |
| ss76901555 | +5543 | T | C | Exon 8 | Val287Ala | Likely affects function [94] | |
| CD209 | rs4804803 | −336 | A | G | Promoter | - | DV [100,101,102], HCV [103], HIV [104,105,106], SARS-CoV [107] |
| rs11465366 | −201 | C | A | Promoter | - | HIV [104,106] | |
| rs2287886 | −139 | T | C | Promoter | - | HIV [106,108] | |
| rs41374747 | +660 | G | A | Exon 4 | Arg198Gln | HIV [104] | |
| rs11465380 | +791 | C | G | Exon 4 | Leu242Val | HIV [104] |
4. Ficolins
4.1. Genetics, Structure, Expression and Binding Specificities of Ficolins
4.2. The Roles of Ficolins in the Immune Response
4.3. Ficolin Interaction with Viruses
4.4. Single Nucleotide Polymorphisms in FCN Genes
4.4.1. The Significance of FCN Gene Single Nucleotide Polymorphisms in Viral Infections
4.4.2. The Significance of Ficolin Serum Concentrations in Viral Infections
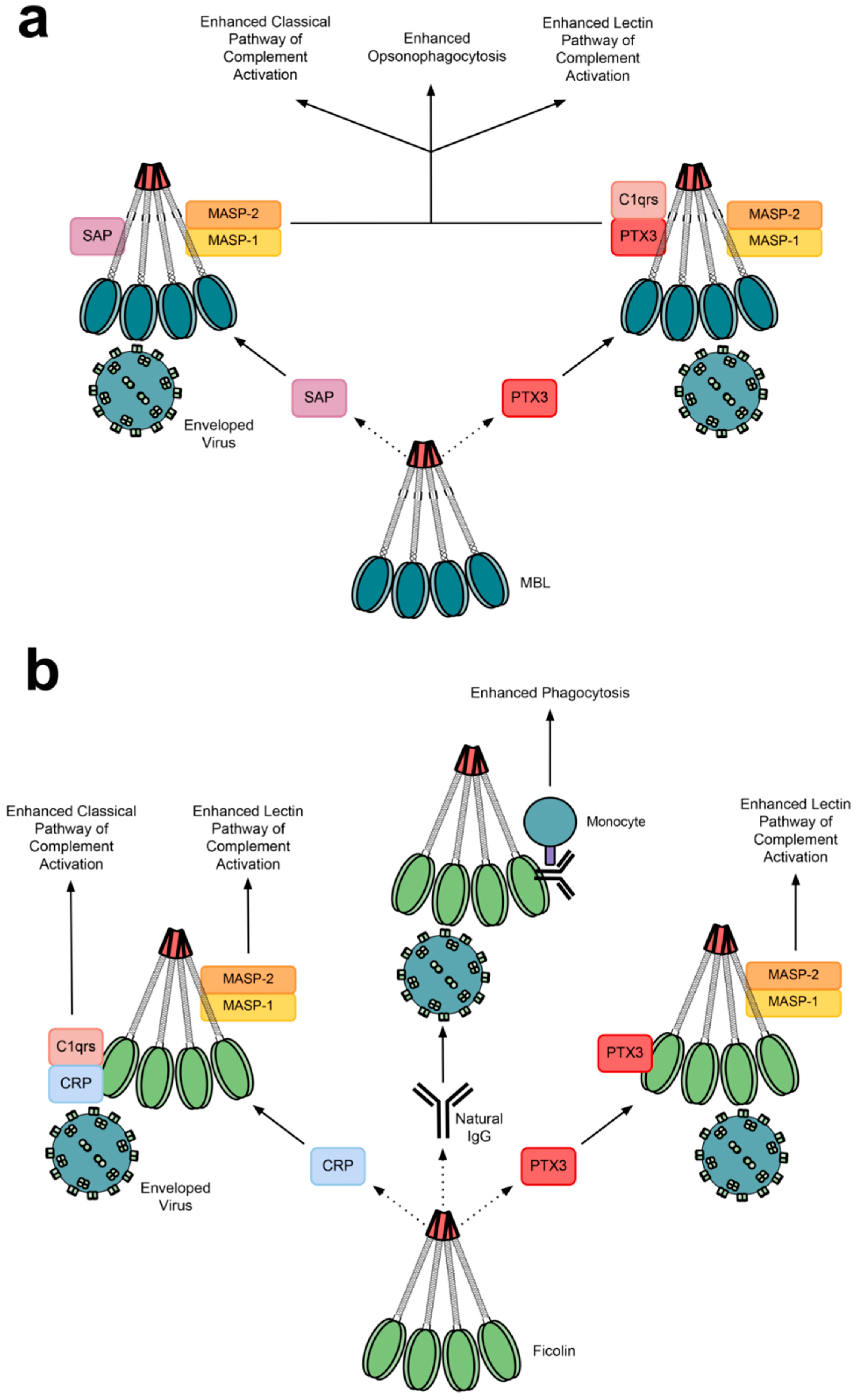
4.5. Cooperative Relationships between Lectins and Other Immune Proteins

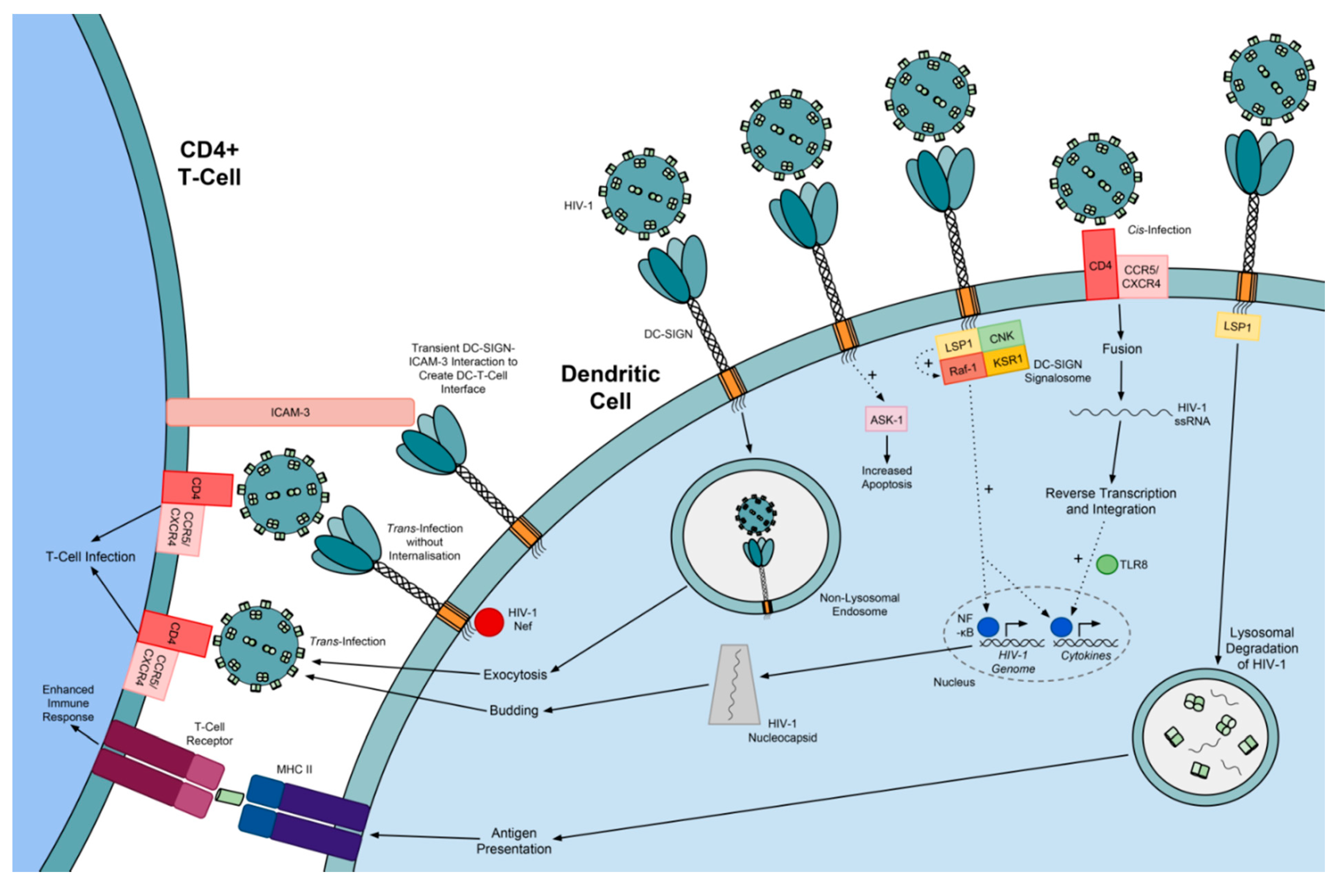
5. DC-SIGN
5.1. Genetics, Structure, Expression and Binding Specificities of DC-SIGN
5.2. Exploitation of DC-SIGN by Viruses
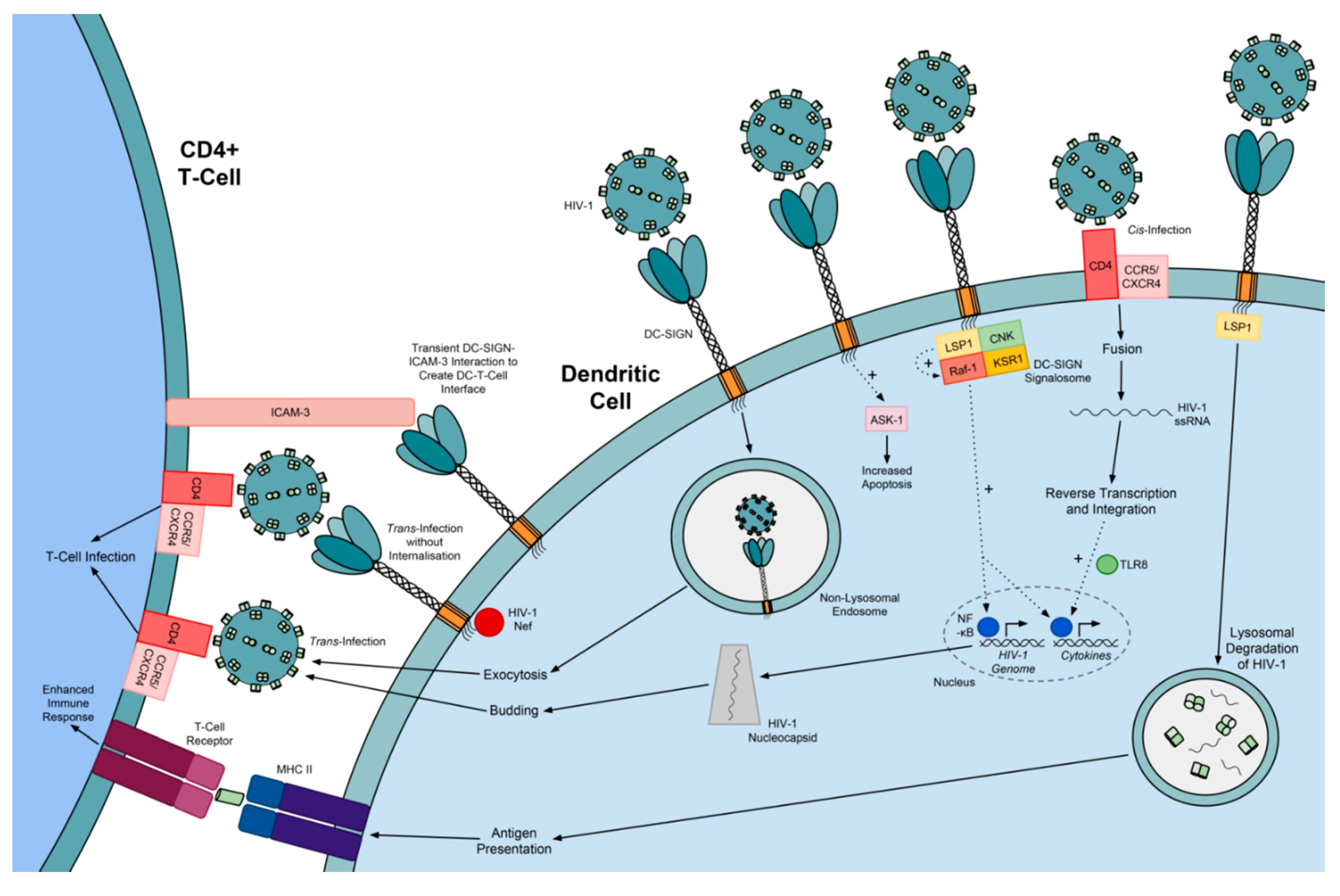
5.3. DC-SIGN Variants
5.4. The Significance of DC-SIGN Variants in Viral Infection
7. Lectin Therapy for Viral Infections
7.1. Soluble Lectin Therapy
7.2. Therapy Using Xenogeneic Lectins
8. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| CRD | carbohydrate-recognition domain |
| PRR | pattern-recognition receptor |
| PAMP | pathogen-associated molecular pattern |
| MBL | mannose-binding lectin |
| DC-SIGN | dendritic cell-specific ICAM-3 grabbing non-integrin |
| MAC | membrane attack complex |
| MASP | MBL-associated serine protease |
| MAP | MBL-associated protein |
| CLD | collagen-like domain |
| GlcNAc | N-acetylglucosamine |
| SARS-CoV | severe acute respiratory syndrome coronavirus |
| DV | Dengue virus |
| WNV | West Nile virus |
| EBOV | Ebola virus |
| HAART | highly active antiretroviral therapy |
| CMV | cytomegalovirus |
| HTLV | human T-cell lymphotropic virus |
| FBG | fibrinogen-like |
| GalNAc | N-acetylgalactosamine |
| GPCR43 | G-protein-coupled receptor 43 |
| PTX3 | pentraxin 3 |
| SAP | serum amyloid P component |
| nIgG | natural immunoglobulin G |
| CRP | C-reactive protein |
| DC | dendritic cell |
| MV | measles virus |
| Siglec-1 | sialic acid-binding Ig-like lectin 1 |
| sDC-SIGN | soluble DC-SIGN |
| RCL | recombinant chimaeric lectin |
| CBA | carbohydrate-binding agent |
| GRFT | Griffithsin |
| CV-N | Cyanovirin |
| SVN | Scytovirin |
| OAA | Oscillatoria agardhii agglutinin |
Conflicts of Interest
References
- Loris, R. Principles of structures of animal and plant lectins. Biochim. Biophys. Acta 2002, 1572, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Sekine, H.; Kenjo, A.; Azumi, K.; Ohi, G.; Takahashi, M.; Kasukawa, R.; Ichikawa, N.; Nakata, M.; Mizuochi, T.; Matsushita, M.; et al. An ancient lectin-dependent complement system in an ascidian: Novel lectin isolated from the plasma of the solitary ascidian, Halocynthia roretzi. J. Immunol. 2001, 167, 4504–4510. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Etzler, M.E.; Cummings, R.D.; Esko, J.D. Discovery and Classification of Glycan-Binding Proteins. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor: Laurel Hollow, NY, USA, 2009. [Google Scholar]
- Ghazarian, H.; Idoni, B.; Oppenheimer, S.B. A glycobiology review: Carbohydrates, lectins and implications in cancer therapeutics. Acta Histochem. 2011, 113, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Vigerust, D.J.; Shepherd, V.L. Virus glycosylation: Role in virulence and immune interactions. Trends Microbiol. 2007, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Sarma, J.V.; Ward, P.A. The complement system. Cell Tissue Res. 2011, 343, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, M.; Crispin, M.; Scanlan, C.N.; Zitzmann, N.; Dwek, R.A. Emerging principles for the therapeutic exploitation of glycosylation. Science 2014, 343, 1235681. [Google Scholar] [CrossRef] [PubMed]
- Dahl, M.R.; Thiel, S.; Matsushita, M.; Fujita, T.; Willis, A.C.; Christensen, T.; Vorup-Jensen, T.; Jensenius, J.C. MASP-3 and its association with distinct complexes of the mannan-binding lectin complement activation pathway. Immunity 2001, 15, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Skjoedt, M.O.; Hummelshoj, T.; Palarasah, Y.; Honore, C.; Koch, C.; Skjodt, K.; Garred, P. A novel mannose-binding lectin/ficolin-associated protein is highly expressed in heart and skeletal muscle tissues and inhibits complement activation. J. Biol. Chem. 2010, 285, 8234–8243. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.M.; Thiel, S.; Thelen, M.; Lynch, N.J.; Vorup-Jensen, T.; Jensenius, J.C.; Schwaeble, W.J. Two constituents of the initiation complex of the mannan-binding lectin activation pathway of complement are encoded by a single structural gene. J. Immunol. 1999, 162, 3481–3490. [Google Scholar] [PubMed]
- Matsushita, M.; Thiel, S.; Jensenius, J.C.; Terai, I.; Fujita, T. Proteolytic activities of two types of mannose-binding lectin-associated serine protease. J. Immunol. 2000, 165, 2637–2642. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Xu, S.; Oliveira, C.L.; Pedersen, J.S.; Thiel, S.; Besenbacher, F.; Vorup-Jensen, T. Conformational changes in mannan-binding lectin bound to ligand surfaces. J. Immunol. 2007, 178, 3016–3022. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, H.; Uitdehaag, J.C.; Davies, J.M.; Wallis, R.; Drickamer, K.; Weis, W.I. Crystal structure of the CUB1-EGF-CUB2 region of mannose-binding protein associated serine protease-2. EMBO J. 2003, 22, 2348–2359. [Google Scholar] [CrossRef] [PubMed]
- Gál, P.; Harmat, V.; Kocsis, A.; Bian, T.; Barna, L.; Ambrus, G.; Vegh, B.; Balczer, J.; Sim, R.B.; Naray-Szabo, G.; et al. A true autoactivating enzyme. Structural insight into mannose-binding lectin-associated serine protease-2 activations. J. Biol. Chem. 2005, 280, 33435–33444. [Google Scholar] [CrossRef]
- Gingras, A.R.; Girija, U.V.; Keeble, A.H.; Panchal, R.; Mitchell, D.A.; Moody, P.C.; Wallis, R. Structural basis of mannan-binding lectin recognition by its associated serine protease MASP-1: Implications for complement activation. Structure 2011, 19, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, G.; Gal, P.; Kojima, M.; Szilagyi, K.; Balczer, J.; Antal, J.; Graf, L.; Laich, A.; Moffatt, B.E.; Schwaeble, W.; et al. Natural substrates and inhibitors of mannan-binding lectin-associated serine protease-1 and -2: A study on recombinant catalytic fragments. J. Immunol. 2003, 170, 1374–1382. [Google Scholar] [CrossRef]
- Megyeri, M.; Harmat, V.; Major, B.; Vegh, A.; Balczer, J.; Heja, D.; Szilagyi, K.; Datz, D.; Pal, G.; Zavodszky, P.; et al. Quantitative characterization of the activation steps of mannan-binding lectin (MBL)-associated serine proteases (MASPs) points to the central role of MASP-1 in the initiation of the complement lectin pathway. J. Biol. Chem. 2013, 288, 8922–8934. [Google Scholar] [CrossRef]
- Degn, S.E.; Jensen, L.; Olszowski, T.; Jensenius, J.C.; Thiel, S. Co-complexes of MASP-1 and MASP-2 associated with the soluble pattern-recognition molecules drive lectin pathway activation in a manner inhibitable by MAp44. J. Immunol. 2013, 191, 1334–1345. [Google Scholar] [CrossRef] [PubMed]
- Parej, K.; Hermann, A.; Donath, N.; Zavodszky, P.; Gal, P.; Dobo, J. Dissociation and re-association studies on the interaction domains of mannan-binding lectin (MBL)-associated serine proteases, MASP-1 and MASP-2, provide evidence for heterodimer formation. Mol. Immunol. 2014, 59, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wallis, R.; Shaw, J.M.; Uitdehaag, J.; Chen, C.B.; Torgersen, D.; Drickamer, K. Localization of the serine protease-binding sites in the collagen-like domain of mannose-binding protein: Indirect effects of naturally occurring mutations on protease binding and activation. J. Biol. Chem. 2004, 279, 14065–14073. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, D.; Kanno, K.; Takahashi, M.; Endo, Y.; Lynch, N.J.; Schwaeble, W.J.; Matsushita, M.; Okabe, M.; Fujita, T. Small mannose-binding lectin-associated protein plays a regulatory role in the lectin complement pathway. J. Immunol. 2006, 177, 8626–8632. [Google Scholar] [CrossRef] [PubMed]
- Rosbjerg, A.; Munthe-Fog, L.; Garred, P.; Skjoedt, M.O. Heterocomplex formation between MBL/ficolin/CL-11-associated serine protease-1 and -3 and MBL/ficolin/CL-11-associated protein-1. J. Immunol. 2014, 192, 4352–4360. [Google Scholar] [CrossRef] [PubMed]
- Gulla, K.C.; Gupta, K.; Krarup, A.; Gal, P.; Schwaeble, W.J.; Sim, R.B.; O’Connor, C.D.; Hajela, K. Activation of mannan-binding lectin-associated serine proteases leads to generation of a fibrin clot. Immunology 2010, 129, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Chang, W.C.; Takahashi, M.; Pavlov, V.; Ishida, Y.; la Bonte, L.; Shi, L.; Fujita, T.; Stahl, G.L.; van Cott, E.M. Mannose-binding lectin and its associated proteases (MASPs) mediate coagulation and its deficiency is a risk factor in developing complications from infection, including disseminated intravascular coagulation. Immunobiology 2011, 216, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Sirmaci, A.; Walsh, T.; Akay, H.; Spiliopoulos, M.; Sakalar, Y.B.; Hasanefendioglu-Bayrak, A.; Duman, D.; Farooq, A.; King, M.C.; Tekin, M. MASP1 mutations in patients with facial, umbilical, coccygeal, and auditory findings of Carnevale, Malpuech, OSA, and Michels syndromes. Am. J. Hum. Genet. 2010, 87, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Sastry, K.; Herman, G.A.; Day, L.; Deignan, E.; Bruns, G.; Morton, C.C.; Ezekowitz, R.A. The human mannose-binding protein gene. Exon structure reveals its evolutionary relationship to a human pulmonary surfactant gene and localization to chromosome 10. J. Exp. Med. 1989, 170, 1175–1189. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Nathamu, S.; Adame, A.; Alire, T.U.; Dumaop, W.; Gouaux, B.; Moore, D.J.; Masliah, E.; HIV Neurobehavioral Research Center Group. Expression of mannose binding lectin in HIV-1-infected brain: Implications for HIV-related neuronal damage and neuroAIDS. Neurobehav HIV Med. 2011, 3, 41–52. [Google Scholar] [CrossRef]
- Jensen, P.H.; Weilguny, D.; Matthiesen, F.; McGuire, K.A.; Shi, L.; Hojrup, P. Characterization of the oligomer structure of recombinant human mannan-binding lectin. J. Biol. Chem. 2005, 280, 11043–11051. [Google Scholar] [CrossRef] [PubMed]
- Yokota, Y.; Arai, T.; Kawasaki, T. Oligomeric structures required for complement activation of serum mannan-binding proteins. J. Biochem. 1995, 117, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Jensenius, H.; Klein, D.C.; van Hecke, M.; Oosterkamp, T.H.; Schmidt, T.; Jensenius, J.C. Mannan-binding lectin: Structure, oligomerization, and flexibility studied by atomic force microscopy. J. Mol. Biol. 2009, 391, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Sheriff, S.; Chang, C.Y.; Ezekowitz, R.A. Human mannose-binding protein carbohydrate recognition domain trimerizes through a triple alpha-helical coiled-coil. Nat. Struct. Biol. 1994, 1, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Phillips, A.; Gor, J.; Wallis, R.; Perkins, S.J. Near-planar solution structures of mannose-binding lectin oligomers provide insight on activation of lectin pathway of complement. J. Biol. Chem. 2012, 287, 3930–3945. [Google Scholar] [CrossRef] [PubMed]
- Teillet, F.; Lacroix, M.; Thiel, S.; Weilguny, D.; Agger, T.; Arlaud, G.J.; Thielens, N.M. Identification of the site of human mannan-binding lectin involved in the interaction with its partner serine proteases: The essential role of Lys55. J. Immunol. 2007, 178, 5710–5716. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Drickamer, K.; Hendrickson, W.A. Structure of a C-type mannose-binding protein complexed with an oligosaccharide. Nature 1992, 360, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.C. Phospholipid-binding activity of human mannan-binding lectin. Immunol. Lett. 1998, 61, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Palaniyar, N.; Nadesalingam, J.; Clark, H.; Shih, M.J.; Dodds, A.W.; Reid, K.B. Nucleic acid is a novel ligand for innate, immune pattern recognition collectins surfactant proteins A and D and mannose-binding lectin. J. Biol. Chem. 2004, 279, 32728–32736. [Google Scholar] [CrossRef] [PubMed]
- Teillet, F.; Dublet, B.; Andrieu, J.P.; Gaboriaud, C.; Arlaud, G.J.; Thielens, N.M. The two major oligomeric forms of human mannan-binding lectin: Chemical characterization, carbohydrate-binding properties, and interaction with MBL-associated serine proteases. J. Immunol. 2005, 174, 2870–2877. [Google Scholar] [CrossRef] [PubMed]
- Saifuddin, M.; Hart, M.L.; Gewurz, H.; Zhang, Y.; Spear, G.T. Interaction of mannose-binding lectin with primary isolates of human immunodeficiency virus type 1. J. Gen. Virol. 2000, 81, 949–955. [Google Scholar] [PubMed]
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody neutralization and escape by HIV-1. Nature 2003, 422, 307–312. [Google Scholar]
- Ying, H.; Ji, X.; Hart, M.L.; Gupta, K.; Saifuddin, M.; Zariffard, M.R.; Spear, G.T. Interaction of mannose-binding lectin with HIV type 1 is sufficient for virus opsonization but not neutralization. AIDS Res. Hum. Retrovir. 2004, 20, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Senaldi, G.; Davies, E.T.; Mahalingam, M.; Lu, J.; Pozniak, A.; Peakman, M.; Reid, K.B.; Vergani, D. Circulating levels of mannose binding protein in human immunodeficiency virus infection. J. Infect. 1995, 31, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Jack, D.L.; Lee, M.E.; Turner, M.W.; Klein, N.J.; Read, R.C. Mannose-binding lectin enhances phagocytosis and killing of Neisseria meningitidis by human macrophages. J. Leukoc. Biol. 2005, 77, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Pagh, R.; Duus, K.; Laursen, I.; Hansen, P.R.; Mangor, J.; Thielens, N.; Arlaud, G.J.; Kongerslev, L.; Hojrup, P.; Houen, G. The chaperone and potential mannan-binding lectin (MBL) co-receptor calreticulin interacts with MBL through the binding site for MBL-associated serine proteases. FEBS J. 2008, 275, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Spear, G.T.; Zariffard, M.R.; Xin, J.; Saifuddin, M. Inhibition of DC-SIGN-mediated trans infection of T cells by mannose-binding lectin. Immunology 2003, 110, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; White, M.R.; Moyo, P.; McClear, S.; Thiel, S.; Hartshorn, K.L.; Takahashi, K. Lack of the pattern recognition molecule mannose-binding lectin increases susceptibility to influenza A virus infection. BMC Immunol. 2010, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Kase, T.; Suzuki, Y.; Kawai, T.; Sakamoto, T.; Ohtani, K.; Eda, S.; Maeda, A.; Okuno, Y.; Kurimura, T.; Wakamiya, N. Human mannan-binding lectin inhibits the infection of influenza A virus without complement. Immunology 1999, 97, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.S.; Keogh, M.J.; Owsianka, A.M.; Adair, R.; Patel, A.H.; Arnold, J.N.; Ball, J.K.; Sim, R.B.; Tarr, A.W.; Hickling, T.P. Specific interaction of hepatitis C virus glycoproteins with mannan binding lectin inhibits virus entry. Protein Cell 2010, 1, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Ip, W.K.; Chan, K.H.; Law, H.K.; Tso, G.H.; Kong, E.K.; Wong, W.H.; To, Y.F.; Yung, R.W.; Chow, E.Y.; Au, K.L.; et al. Mannose-binding lectin in severe acute respiratory syndrome coronavirus infection. J. Infect. Dis. 2005, 191, 1697–1704. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lu, K.; Pfefferle, S.; Bertram, S.; Glowacka, I.; Drosten, C.; Pohlmann, S.; Simmons, G. A single asparagine-linked glycosylation site of the severe acute respiratory syndrome coronavirus spike glycoprotein facilitates inhibition by mannose-binding lectin through multiple mechanisms. J. Virol. 2010, 84, 8753–8764. [Google Scholar] [CrossRef] [PubMed]
- Avirutnan, P.; Hauhart, R.E.; Marovich, M.A.; Garred, P.; Atkinson, J.P.; Diamond, M.S. Complement-mediated neutralization of dengue virus requires mannose-binding lectin. MBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Lin, T.Y.; Beasley, D.W.; Stover, C.M.; Schwaeble, W.J.; Pierson, T.C.; Diamond, M.S. Direct complement restriction of flavivirus infection requires glycan recognition by mannose-binding lectin. Cell Host Microbe 2010, 8, 186–195. [Google Scholar] [CrossRef] [PubMed]
- El Saadany, S.A.; Ziada, D.H.; Farrag, W.; Hazaa, S. Fibrosis severity and mannan-binding lectin (MBL)/MBL-associated serine protease 1 (MASP-1) complex in HCV-infected patients. Arab. J. Gastroenterol. 2011, 12, 68–73. [Google Scholar]
- Michelow, I.C.; Lear, C.; Scully, C.; Prugar, L.I.; Longley, C.B.; Yantosca, L.M.; Ji, X.; Karpel, M.; Brudner, M.; Takahashi, K.; et al. High-dose mannose-binding lectin therapy for Ebola virus infection. J. Infect. Dis. 2011, 203, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Olinger, G.G.; Aris, S.; Chen, Y.; Gewurz, H.; Spear, G.T. Mannose-binding lectin binds to Ebola and Marburg envelope glycoproteins, resulting in blocking of virus interaction with DC-SIGN and complement-mediated virus neutralization. J. Gen. Virol. 2005, 86, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Brudner, M.; Karpel, M.; Lear, C.; Chen, L.; Yantosca, L.M.; Scully, C.; Sarraju, A.; Sokolovska, A.; Zariffard, M.R.; Eisen, D.P.; et al. Lectin-dependent enhancement of Ebola virus infection via soluble and transmembrane C-type lectin receptors. PLoS One 2013, 8, e60838. [Google Scholar] [CrossRef] [PubMed]
- Bachis, A.; Aden, S.A.; Nosheny, R.L.; Andrews, P.M.; Mocchetti, I. Axonal transport of human immunodeficiency virus type 1 envelope protein glycoprotein 120 is found in association with neuronal apoptosis. J. Neurosci. 2006, 26, 6771–6780. [Google Scholar] [CrossRef] [PubMed]
- Teodorof, C.; Divakar, S.; Soontornniyomkij, B.; Achim, C.L.; Kaul, M.; Singh, K.K. Intracellular mannose binding lectin mediates subcellular trafficking of HIV-1 gp120 in neurons. Neurobiol. Dis. 2014, 69, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Sumiya, M.; Super, M.; Tabona, P.; Levinsky, R.J.; Arai, T.; Turner, M.W.; Summerfield, J.A. Molecular basis of opsonic defect in immunodeficient children. Lancet 1991, 337, 1569–1570. [Google Scholar] [CrossRef] [PubMed]
- Lipscombe, R.J.; Sumiya, M.; Hill, A.V.; Lau, Y.L.; Levinsky, R.J.; Summerfield, J.A.; Turner, M.W. High frequencies in African and non-African populations of independent mutations in the mannose binding protein gene. Hum. Mol. Genet. 1992, 1, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.O.; Garred, P.; Kurtzhals, J.A.; Lamm, L.U.; Ryder, L.P.; Thiel, S.; Svejgaard, A. A new frequent allele is the missing link in the structural polymorphism of the human mannan-binding protein. Immunogenetics 1994, 40, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.O.; Satz, M.L.; Hogh, B.; Svejgaard, A.; Garred, P. Different molecular events result in low protein levels of mannan-binding lectin in populations from southeast Africa and South America. J. Immunol. 1998, 161, 3169–3175. [Google Scholar] [PubMed]
- Chalmers, J.D.; McHugh, B.J.; Doherty, C.; Smith, M.P.; Govan, J.R.; Kilpatrick, D.C.; Hill, A.T. Mannose-binding lectin deficiency and disease severity in non-cystic fibrosis bronchiectasis: A prospective study. Lancet Respir. Med. 2013, 1, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Lipscombe, R.J.; Sumiya, M.; Summerfield, J.A.; Turner, M.W. Distinct physicochemical characteristics of human mannose binding protein expressed by individuals of differing genotype. Immunology 1995, 85, 660–667. [Google Scholar] [PubMed]
- Verga Falzacappa, M.V.; Segat, L.; Puppini, B.; Amoroso, A.; Crovella, S. Evolution of the mannose-binding lectin gene in primates. Genes Immun. 2004, 5, 653–661. [Google Scholar]
- Garred, P.; Larsen, F.; Seyfarth, J.; Fujita, R.; Madsen, H.O. Mannose-binding lectin and its genetic variants. Genes Immun. 2006, 7, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Soltani, A.; RahmatiRad, S.; Pourpak, Z.; Alizadeh, Z.; Saghafi, S.; HajiBeigi, B.; Zeidi, M.; Farazmand, A. Polymorphisms and serum level of mannose-binding lectin: An Iranian survey. Iran. J. Allergy Asthma Immunol. 2014, 13, 428–432. [Google Scholar] [PubMed]
- Lee, S.G.; Yum, J.S.; Moon, H.M.; Kim, H.J.; Yang, Y.J.; Kim, H.L.; Yoon, Y.; Lee, S.; Song, K. Analysis of mannose-binding lectin 2 (MBL2) genotype and the serum protein levels in the Korean population. Mol. Immunol. 2005, 42, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Thiel, S.; Bjerke, T.; Hansen, D.; Poulsen, L.K.; Schiotz, P.O.; Jensenius, J.C. Ontogeny of human mannan-binding protein, a lectin of the innate immune system. Pediatr. Allergy Immunol. 1995, 6, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Roos, A.; Garred, P.; Wildenberg, M.E.; Lynch, N.J.; Munoz, J.R.; Zuiverloon, T.C.; Bouwman, L.H.; Schlagwein, N.; Fallaux van den Houten, F.C.; Faber-Krol, M.C.; et al. Antibody-mediated activation of the classical pathway of complement may compensate for mannose-binding lectin deficiency. Eur. J. Immunol. 2004, 34, 2589–2598. [Google Scholar] [CrossRef] [PubMed]
- Garred, P.; Madsen, H.O.; Balslev, U.; Hofmann, B.; Pedersen, C.; Gerstoft, J.; Svejgaard, A. Susceptibility to HIV infection and progression of AIDS in relation to variant alleles of mannose-binding lectin. Lancet 1997, 349, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Prohaszka, Z.; Thiel, S.; Ujhelyi, E.; Szlavik, J.; Banhegyi, D.; Fust, G. Mannan-binding lectin serum concentrations in HIV-infected patients are influenced by the stage of disease. Immunol. Lett. 1997, 58, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Catano, G.; Agan, B.K.; Kulkarni, H.; Telles, V.; Marconi, V.C.; Dolan, M.J.; Ahuja, S.K. Independent effects of genetic variations in mannose-binding lectin influence the course of HIV disease: The advantage of heterozygosity for coding mutations. J. Infect. Dis. 2008, 198, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Vallinoto, A.C.; Menezes-Costa, M.R.; Alves, A.E.; Machado, L.F.; de Azevedo, V.N.; Souza, L.L.; Ishak Mde, O.; Ishak, R. Mannose-binding lectin gene polymorphism and its impact on human immunodeficiency virus 1 infection. Mol. Immunol. 2006, 43, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Boniotto, M.; Crovella, S.; Pirulli, D.; Scarlatti, G.; Spano, A.; Vatta, L.; Zezlina, S.; Tovo, P.A.; Palomba, E.; Amoroso, A. Polymorphisms in the MBL2 promoter correlated with risk of HIV-1 vertical transmission and AIDS progression. Genes Immun. 2000, 1, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Vallinoto, A.C.; Muto, N.A.; Alves, A.E.; Machado, L.F.; Azevedo, V.N.; Souza, L.L.; Ishak, M.O.; Ishak, R. Characterization of polymorphisms in the mannose-binding lectin gene promoter among human immunodeficiency virus 1 infected subjects. Mem. Inst. Oswaldo Cruz. 2008, 103, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Arias, M.; di Flumeri, C.; Garcia, L.F.; Schurr, E. Absence of association between mannose-binding lectin gene polymorphisms and HIV-1 infection in a Colombian population. Immunogenetics 2003, 55, 49–52. [Google Scholar] [PubMed]
- Lian, Y.C.; Della-Negra, M.; Rutz, R.; Ferriani, V.; de Moraes Vasconcelos, D.; da Silva Duarte, A.J.; Kirschfink, M.; Grumach, A.S. Immunological analysis in paediatric HIV patients at different stages of the disease. Scand. J. Immunol. 2004, 60, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.L.; Andersen, P.L.; Koch, C.; Jensenius, J.C.; Thiel, S. The level of the serum opsonin, mannan-binding protein in HIV-1 antibody-positive patients. Clin. Exp. Immunol. 1995, 100, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Heggelund, L.; Mollnes, T.E.; Ueland, T.; Christophersen, B.; Aukrust, P.; Froland, S.S. Mannose-binding lectin in HIV infection: Relation to disease progression and highly active antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2003, 32, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Maas, J.; de Roda Husman, A.M.; Brouwer, M.; Krol, A.; Coutinho, R.; Keet, I.; van Leeuwen, R.; Schuitemaker, H. Presence of the variant mannose-binding lectin alleles associated with slower progression to AIDS. Amsterdam Cohort Study. AIDS 1998, 12, 2275–2280. [Google Scholar] [CrossRef] [PubMed]
- Chong, W.P.; To, Y.F.; Ip, W.K.; Yuen, M.F.; Poon, T.P.; Wong, W.H.; Lai, C.L.; Lau, Y.L. Mannose-binding lectin in chronic hepatitis B virus infection. Hepatology 2005, 42, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Halla, M.C.; do Carmo, R.F.; Silva Vasconcelos, L.R.; Pereira, L.B.; Moura, P.; de Siqueira, E.R.; Pereira, L.M.; Mendonca Cavalcanti Mdo, S. Association of hepatitis C virus infection and liver fibrosis severity with the variants alleles of MBL2 gene in a Brazilian population. Hum. Immunol. 2010, 71, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Alves Pedroso, M.L.; Boldt, A.B.; Pereira-Ferrari, L.; Steffensen, R.; Strauss, E.; Jensenius, J.C.; Ioshii, S.O.; Messias-Reason, I. Mannan-binding lectin MBL2 gene polymorphism in chronic hepatitis C: Association with the severity of liver fibrosis and response to interferon therapy. Clin. Exp. Immunol. 2008, 152, 258–264. [Google Scholar]
- Alves, A.E.; Hermes, R.B.; Tamegao-Lopes, B.; Machado, L.F.; Azevedo, V.N.; Ishak, M.O.; Ishak, R.; Lemos, J.A.; Vallinoto, A.C. Polymorphism in the promoter region of the mannose-binding lectin gene among human T-cell lymphotropic virus infected subjects. Mem. Inst. Oswaldo Cruz. 2007, 102, 991–994. [Google Scholar] [CrossRef]
- Kwakkel-van Erp, J.M.; Paantjens, A.W.; van Kessel, D.A.; Grutters, J.C.; van den Bosch, J.M.; van de Graaf, E.A.; Otten, H.G. Mannose-binding lectin deficiency linked to cytomegalovirus (CMV) reactivation and survival in lung transplantation. Clin. Exp. Immunol. 2011, 165, 410–416. [Google Scholar]
- De Rooij, B.J.; van der Beek, M.T.; van Hoek, B.; Vossen, A.C.; Rogier Ten Hove, W.; Roos, A.; Schaapherder, A.F.; Porte, R.J.; van der Reijden, J.J.; Coenraad, M.J.; et al. Mannose-binding lectin and ficolin-2 gene polymorphisms predispose to cytomegalovirus (re)infection after orthotopic liver transplantation. J. Hepatol. 2011, 55, 800–807. [Google Scholar]
- Thomas, H.C.; Foster, G.R.; Sumiya, M.; McIntosh, D.; Jack, D.L.; Turner, M.W.; Summerfield, J.A. Mutation of gene of mannose-binding protein associated with chronic hepatitis B viral infection. Lancet 1996, 348, 1417–1419. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.D.; Zhao, M.F.; Wan, T.H.; Song, G.Z.; He, J.L.; Chen, Z. Association between Mannose-binding lectin gene polymorphisms and hepatitis B virus infection: A meta-analysis. PLoS One 2013, 8, e75371. [Google Scholar] [CrossRef] [PubMed]
- Koutsounaki, E.; Goulielmos, G.N.; Koulentaki, M.; Choulaki, C.; Kouroumalis, E.; Galanakis, E. Mannose-binding lectin MBL2 gene polymorphisms and outcome of hepatitis C virus-infected patients. J. Clin. Immunol. 2008, 28, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, A.; Mossad, N.; Abdel Hafez, H.; Elsayed, N. Mannose-binding lectin exon 1 polymorphisms in Egyptian patients with chronic hepatitis C virus infection. Comp. Clin. Pathol. 2014, 23, 1339–1342. [Google Scholar] [CrossRef]
- Yuen, M.F.; Lau, C.S.; Lau, Y.L.; Wong, W.M.; Cheng, C.C.; Lai, C.L. Mannose binding lectin gene mutations are associated with progression of liver disease in chronic hepatitis B infection. Hepatology 1999, 29, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Pontes, G.S.; Tamegao-Lopes, B.; Machado, L.F.; Azevedo, V.N.; Ishak, M.O.; Ishak, R.; Lemos, J.A.; Vallinoto, A.C. Characterization of mannose-binding lectin gene polymorphism among human T-cell lymphotropic virus 1 and 2-infected asymptomatic subjects. Hum. Immunol. 2005, 66, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Ammitzbøll, C.G.; Kjaer, T.R.; Steffensen, R.; Stengaard-Pedersen, K.; Nielsen, H.J.; Thiel, S.; Bogsted, M.; Jensenius, J.C. Non-synonymous polymorphisms in the FCN1 gene determine ligand-binding ability and serum levels of M-ficolin. PLoS One 2012, 7, e50585. [Google Scholar] [CrossRef] [PubMed]
- Hummelshøj, T.; Munthe-Fog, L.; Madsen, H.O.; Garred, P. Functional SNPs in the human ficolin (FCN) genes reveal distinct geographical patterns. Mol. Immunol. 2008, 45, 2508–2520. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.V.; Toan, N.L.; Song le, H.; Ouf, E.A.; Bock, C.T.; Kremsner, P.G.; Kun, J.F.; Velavan, T.P. Ficolin-2 levels and FCN2 haplotypes influence hepatitis B infection outcome in Vietnamese patients. PLoS One 2011, 6, e28113. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Doni, A.; Hummelshoj, T.; Honore, C.; Bastone, A.; Mantovani, A.; Thielens, N.M.; Garred, P. Synergy between ficolin-2 and pentraxin 3 boosts innate immune recognition and complement deposition. J. Biol. Chem. 2009, 284, 28263–28275. [Google Scholar] [CrossRef] [PubMed]
- Munthe-Fog, L.; Hummelshoj, T.; Ma, Y.J.; Hansen, B.E.; Koch, C.; Madsen, H.O.; Skjodt, K.; Garred, P. Characterization of a polymorphism in the coding sequence of FCN3 resulting in a Ficolin-3 (Hakata antigen) deficiency state. Mol. Immunol. 2008, 45, 2660–2666. [Google Scholar] [CrossRef] [PubMed]
- Munthe-Fog, L.; Hummelshoj, T.; Honore, C.; Madsen, H.O.; Permin, H.; Garred, P. Immunodeficiency associated with FCN3 mutation and ficolin-3 deficiency. N. Engl. J. Med. 2009, 360, 2637–2644. [Google Scholar] [CrossRef] [PubMed]
- Schlapbach, L.J.; Thiel, S.; Kessler, U.; Ammann, R.A.; Aebi, C.; Jensenius, J.C. Congenital H-ficolin deficiency in premature infants with severe necrotising enterocolitis. Gut 2011, 60, 1438–1439. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, R.F.; Liu, J.W.; Lee, I.K.; Lee, C.P.; Kuo, H.C.; Huang, S.K.; Yang, K.D. DC-SIGN (CD209) Promoter -336 A/G polymorphism is associated with dengue hemorrhagic fever and correlated to DC-SIGN expression and immune augmentation. PLoS Negl. Trop. Dis. 2011, 5, e934. [Google Scholar] [CrossRef] [PubMed]
- Sakuntabhai, A.; Turbpaiboon, C.; Casademont, I.; Chuansumrit, A.; Lowhnoo, T.; Kajaste-Rudnitski, A.; Kalayanarooj, S.M.; Tangnararatchakit, K.; Tangthawornchaikul, N.; Vasanawathana, S.; et al. A variant in the CD209 promoter is associated with severity of dengue disease. Nat. Genet. 2005, 37, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.F.; Lima, C.P.; Azevedo Rdo, S.; Mendonca, D.S.; Rodrigues, S.G.; Carvalho, V.L.; Pinto, E.V.; Maia, A.L.; Maia, M.H.; Vasconcelos, J.M.; et al. Polymorphism of DC-SIGN (CD209) promoter in association with clinical symptoms of dengue fever. Viral Immunol. 2014, 27, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.J.; Dring, M.; Ryan, C.M.; McNulty, C.; Stevenson, N.J.; Lawless, M.W.; Crowe, J.; Nolan, N.; Hegarty, J.E.; O’Farrelly, C. Variant in CD209 promoter is associated with severity of liver disease in chronic hepatitis C virus infection. Hum. Immunol. 2010, 71, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Boily-Larouche, G.; Milev, M.P.; Zijenah, L.S.; Labbe, A.C.; Zannou, D.M.; Humphrey, J.H.; Ward, B.J.; Poudrier, J.; Mouland, A.J.; Cohen, E.A.; et al. Naturally-occurring genetic variants in human DC-SIGN increase HIV-1 capture, cell-transfer and risk of mother-to-child transmission. PLoS One 2012, 7, e40706. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.P.; Lederman, M.M.; Hutcheson, H.B.; Goedert, J.J.; Nelson, G.W.; van Kooyk, Y.; Detels, R.; Buchbinder, S.; Hoots, K.; Vlahov, D.; et al. Association of DC-SIGN promoter polymorphism with increased risk for parenteral, but not mucosal, acquisition of human immunodeficiency virus type 1 infection. J. Virol. 2004, 78, 14053–14056. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.C.; Segat, L.; Zanin, V.; Arraes, L.C.; Crovella, S. Polymorphisms in DC-SIGN and L-SIGN genes are associated with HIV-1 vertical transmission in a Northeastern Brazilian population. Hum. Immunol. 2012, 73, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.Y.; Xu, M.S.; Ching, J.C.; So, T.M.; Lai, S.T.; Chu, C.M.; Yam, L.Y.; Wong, A.T.; Chung, P.H.; Chan, V.S.; et al. CD209 (DC-SIGN) -336A>G promoter polymorphism and severe acute respiratory syndrome in Hong Kong Chinese. Hum. Immunol. 2010, 71, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Kagone, T.S.; Bisseye, C.; Meda, N.; Testa, J.; Pietra, V.; Kania, D.; Yonli, A.T.; Compaore, T.R.; Nikiema, J.B.; de Souza, C.; et al. A variant of DC-SIGN gene promoter associated with resistance to HIV-1 in serodiscordant couples in Burkina Faso. Asian Pac. J. Trop. Med. 2014, 7S1, S93–S96. [Google Scholar] [CrossRef]
- Eisen, D.P.; Marshall, C.; Dean, M.M.; Sasadeusz, J.; Richards, M.; Buising, K.; Cheng, A.; Johnson, P.D.; Barr, I.G.; McBryde, E.S. No association between mannose-binding lectin deficiency and H1N1 2009 infection observed during the first season of this novel pandemic influenza virus. Hum. Immunol. 2011, 72, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.E.; Zhang, M.; Zarin-Pass, R.; Bernig, T.; Tseng, F.C.; Xiao, N.; Yeager, M.; Edlin, B.R.; Chanock, S.J.; O’Brien, T.R. MBL2 and hepatitis C virus infection among injection drug users. BMC Infect. Dis. 2008, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Vallinoto, A.C.; da Silva, R.F.; Hermes, R.B.; Amaral, I.S.; Miranda, E.C.; Barbosa, M.S.; Moia Lde, J.; Conde, S.R.; Soares Mdo, C.; Lemos, J.A.; et al. Mannose-binding lectin gene polymorphisms are not associated with susceptibility to hepatitis C virus infection in the Brazilian Amazon region. Hum. Immunol. 2009, 70, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Esmat, S.; Omran, D.; Sleem, G.A.; Rashed, L. Serum mannan-binding lectin in egyptian patients with chronic hepatitis C: Its relation to disease progression and response to treatment. Hepat. Mon. 2012, 12, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Q.; Wang, H.; Tian, T.; Zhu, Q.; Wang, H.; Bai, X.; Yang, X.; Wang, Z.; Dong, M. Potential role of mannose-binding lectin in intrauterine transmission of hepatitis B virus. Jpn. J. Infect. Dis. 2013, 66, 391–393. [Google Scholar] [PubMed]
- Hӧhler, T.; Wunschel, M.; Gerken, G.; Schneider, P.M.; Meyer zum Buschenfelde, K.H.; Rittner, C. No association between mannose-binding lectin alleles and susceptibility to chronic hepatitis B virus infection in German patients. Exp. Clin. Immunogenet. 1998, 15, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Sato, Y.; Matsushita, M.; Fujita, T. Cloning and characterization of the human lectin P35 gene and its related gene. Genomics 1996, 36, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, H.; Ronnstrand, L.; Miyagawa, K.; Ohashi, H.; Heldin, C.H.; Miyazono, K. Purification of transforming growth factor-beta 1 binding proteins from porcine uterus membranes. J. Biol. Chem. 1991, 266, 22459–22464. [Google Scholar] [PubMed]
- Lu, J.; Le, Y.; Kon, O.L.; Chan, J.; Lee, S.H. Biosynthesis of human ficolin, an Escherichia coli-binding protein, by monocytes: Comparison with the synthesis of two macrophage-specific proteins, C1q and the mannose receptor. Immunology 1996, 89, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, R.; Yae, Y.; Akaiwa, M.; Kitajima, S.; Shibata, Y.; Sato, H.; Hirata, J.; Okochi, K.; Izuhara, K.; Hamasaki, N. Cloning and characterization of the Hakata antigen, a member of the ficolin/opsonin p35 lectin family. J. Biol. Chem. 1998, 273, 20721–20727. [Google Scholar] [CrossRef] [PubMed]
- Kenjo, A.; Takahashi, M.; Matsushita, M.; Endo, Y.; Nakata, M.; Mizuochi, T.; Fujita, T. Cloning and characterization of novel ficolins from the solitary ascidian, Halocynthia roretzi. J. Biol. Chem. 2001, 276, 19959–19965. [Google Scholar] [CrossRef] [PubMed]
- Lynch, N.J.; Khan, S.U.; Stover, C.M.; Sandrini, S.M.; Marston, D.; Presanis, J.S.; Schwaeble, W.J. Composition of the lectin pathway of complement in Gallus gallus: Absence of mannan-binding lectin-associated serine protease-1 in birds. J. Immunol. 2005, 174, 4998–5006. [Google Scholar] [CrossRef] [PubMed]
- Hummelshøj, T.; Nissen, J.; Munthe-Fog, L.; Koch, C.; Bertelsen, M.F.; Garred, P. Allelic lineages of the ficolin genes (FCNs) are passed from ancestral to descendant primates. PLoS One 2011, 6, e28187. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, H.; Hellman, U.; Wernstedt, C.; Gonez, L.J.; Claesson-Welsh, L.; Heldin, C.H.; Miyazono, K. Molecular cloning and characterization of ficolin, a multimeric protein with fibrinogen- and collagen-like domains. J. Biol. Chem. 1993, 268, 14505–14513. [Google Scholar] [PubMed]
- Garred, P.; Honore, C.; Ma, Y.J.; Rorvig, S.; Cowland, J.; Borregaard, N.; Hummelshoj, T. The genetics of ficolins. J. Innate Immun. 2010, 2, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Hummelshøj, T.; Thielens, N.M.; Madsen, H.O.; Arlaud, G.J.; Sim, R.B.; Garred, P. Molecular organization of human Ficolin-2. Mol. Immunol. 2007, 44, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Endo, Y.; Taira, S.; Sato, Y.; Fujita, T.; Ichikawa, N.; Nakata, M.; Mizuochi, T. A novel human serum lectin with collagen- and fibrinogen-like domains that functions as an opsonin. J. Biol. Chem. 1996, 271, 2448–2454. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Dumestre-Perard, C.; Schoehn, G.; Houen, G.; Cesbron, J.Y.; Arlaud, G.J.; Thielens, N.M. Residue Lys57 in the collagen-like region of human L-ficolin and its counterpart Lys47 in H-ficolin play a key role in the interaction with the mannan-binding lectin-associated serine proteases and the collectin receptor calreticulin. J. Immunol. 2009, 182, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Drickamer, K. Trimeric structure of a C-type mannose-binding protein. Structure 1994, 2, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, T.; Erickson, H.P. The disulfide bonding pattern in ficolin multimers. J. Biol. Chem. 2004, 279, 6534–6539. [Google Scholar] [CrossRef] [PubMed]
- Akaiwa, M.; Yae, Y.; Sugimoto, R.; Suzuki, S.O.; Iwaki, T.; Izuhara, K.; Hamasaki, N. Hakata antigen, a new member of the ficolin/opsonin p35 family, is a novel human lectin secreted into bronchus/alveolus and bile. J. Histochem. Cytochem. 1999, 47, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Endo, Y.; Iwaki, D.; Nakata, M.; Matsushita, M.; Wada, I.; Inoue, K.; Munakata, M.; Fujita, T. Human M-ficolin is a secretory protein that activates the lectin complement pathway. J. Immunol. 2005, 175, 3150–3156. [Google Scholar] [CrossRef] [PubMed]
- Schlapbach, L.J.; Aebi, C.; Hansen, A.G.; Hirt, A.; Jensenius, J.C.; Ammann, R.A. H-ficolin serum concentration and susceptibility to fever and neutropenia in paediatric cancer patients. Clin. Exp. Immunol. 2009, 157, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.C.; Fujita, T.; Matsushita, M. P35, an opsonic lectin of the ficolin family, in human blood from neonates, normal adults, and recurrent miscarriage patients. Immunol. Lett. 1999, 67, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Munthe-Fog, L.; Hummelshoj, T.; Hansen, B.E.; Koch, C.; Madsen, H.O.; Skjodt, K.; Garred, P. The impact of FCN2 polymorphisms and haplotypes on the Ficolin-2 serum levels. Scand. J. Immunol. 2007, 65, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Wittenborn, T.; Thiel, S.; Jensen, L.; Nielsen, H.J.; Jensenius, J.C. Characteristics and biological variations of M-ficolin, a pattern recognition molecule, in plasma. J. Innate Immun. 2010, 2, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Garlatti, V.; Belloy, N.; Martin, L.; Lacroix, M.; Matsushita, M.; Endo, Y.; Fujita, T.; Fontecilla-Camps, J.C.; Arlaud, G.J.; Thielens, N.M.; et al. Structural insights into the innate immune recognition specificities of L- and H-ficolins. EMBO J. 2007, 26, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Vassal-Stermann, E.; Lacroix, M.; Gout, E.; Laffly, E.; Pedersen, C.M.; Martin, L.; Amoroso, A.; Schmidt, R.R.; Zahringer, U.; Gaboriaud, C.; et al. Human L-Ficolin Recognizes Phosphocholine Moieties of Pneumococcal Teichoic Acid. J. Immunol. 2014. [Google Scholar] [CrossRef]
- Luo, F.; Sun, X.; Wang, Y.; Wang, Q.; Wu, Y.; Pan, Q.; Fang, C.; Zhang, X.L. Ficolin-2 Defends against Virulent Mycobacteria Tuberculosis Infection in Vivo, and Its Insufficiency Is Associated with Infection in Humans. PLoS One 2013, 8, e73859. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.L.; Honore, C.; Hummelshoj, T.; Hansen, B.E.; Madsen, H.O.; Garred, P. Ficolin-2 recognizes DNA and participates in the clearance of dying host cells. Mol. Immunol. 2007, 44, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.R.; Brown, R.J.; Zothner, C.; Urbanowicz, R.A.; Mason, C.P.; Krarup, A.; McClure, C.P.; Irving, W.L.; Ball, J.K.; Harris, M.; et al. Recombinant human L-ficolin directly neutralizes hepatitis C virus entry. J. Innate Immun. 2014, 6, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Keirstead, N.D.; Lee, C.; Yoo, D.; Brooks, A.S.; Hayes, M.A. Porcine plasma ficolin binds and reduces infectivity of porcine reproductive and respiratory syndrome virus (PRRSV) in vitro. Antivir. Res. 2008, 77, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ali, M.A.; Shi, Y.; Zhao, Y.; Luo, F.; Yu, J.; Xiang, T.; Tang, J.; Li, D.; Hu, Q.; et al. Specifically binding of L-ficolin to N-glycans of HCV envelope glycoproteins E1 and E2 leads to complement activation. Cell. Mol. Immunol. 2009, 6, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Chen, H.; Wang, F.; Jeza, V.T.; Hou, W.; Zhao, Y.; Xiang, T.; Zhu, Y.; Endo, Y.; Fujita, T.; et al. L-ficolin binds to the glycoproteins hemagglutinin and neuraminidase and inhibits influenza A virus infection both in vitro and in vivo. J. Innate Immun. 2012, 4, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ren, Y.; Zhang, X.; Zhao, P.; Tao, W.; Zhong, J.; Li, Q.; Zhang, X.L. Ficolin-2 inhibits hepatitis C virus infection, whereas apolipoprotein E3 mediates viral immune escape. J. Immunol. 2014, 193, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; White, M.; Vathipadiekal, V.; Tripathi, S.; Mbianda, J.; Ieong, M.; Qi, L.; Taubenberger, J.K.; Takahashi, K.; Jensenius, J.C.; et al. Human H-ficolin inhibits replication of seasonal and pandemic influenza A viruses. J. Immunol. 2012, 189, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Ding, Q.; Zhang, X. Ficolins and infectious diseases. Virol. Sin. 2014, 29, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Honore, C.; Rorvig, S.; Hummelshoj, T.; Skjoedt, M.O.; Borregaard, N.; Garred, P. Tethering of Ficolin-1 to cell surfaces through recognition of sialic acid by the fibrinogen-like domain. J. Leukoc. Biol. 2010, 88, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, L.; Ang, Z.; Yoong, S.L.; Tran, T.T.; Anand, G.S.; Tan, N.S.; Ho, B.; Ding, J.L. Secreted M-ficolin anchors onto monocyte transmembrane G protein-coupled receptor 43 and cross talks with plasma C-reactive protein to mediate immune signaling and regulate host defense. J. Immunol. 2010, 185, 6899–6910. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, T.R.; Hansen, A.G.; Sorensen, U.B.; Nielsen, O.; Thiel, S.; Jensenius, J.C. Investigations on the pattern recognition molecule M-ficolin: Quantitative aspects of bacterial binding and leukocyte association. J. Leukoc. Biol. 2011, 90, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Hummelshøj, T.; Munthe-Fog, L.; Madsen, H.O.; Fujita, T.; Matsushita, M.; Garred, P. Polymorphisms in the FCN2 gene determine serum variation and function of Ficolin-2. Hum. Mol. Genet. 2005, 14, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Herpers, B.L.; Immink, M.M.; de Jong, B.A.; van Velzen-Blad, H.; de Jongh, B.M.; van Hannen, E.J. Coding and non-coding polymorphisms in the lectin pathway activator L-ficolin gene in 188 Dutch blood bank donors. Mol. Immunol. 2006, 43, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Garred, P.; Honore, C.; Ma, Y.J.; Munthe-Fog, L.; Hummelshoj, T. MBL2, FCN1, FCN2 and FCN3-The genes behind the initiation of the lectin pathway of complement. Mol. Immunol. 2009, 46, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Assaf, A.; Hoang, T.V.; Faik, I.; Aebischer, T.; Kremsner, P.G.; Kun, J.F.; Velavan, T.P. Genetic evidence of functional ficolin-2 haplotype as susceptibility factor in cutaneous leishmaniasis. PLoS One 2012, 7, e34113. [Google Scholar] [CrossRef] [PubMed]
- De Messias-Reason, I.; Kremsner, P.G.; Kun, J.F. Functional haplotypes that produce normal ficolin-2 levels protect against clinical leprosy. J. Infect. Dis. 2009, 199, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, J.D.; Kilpatrick, D.C.; McHugh, B.J.; Smith, M.P.; Govan, J.R.W.; Doherty, C.; Matsushita, M.; Hart, S.P.; Sethi, T.; Hill, A.T. T2 Single nucleotide polymorphisms in the ficolin-2 gene predispose to Pseudomonas aeruginosa infection and disease severity in non-cystic fibrosis bronchiectasis. Thorax 2011, 66, A1–A2. [Google Scholar] [CrossRef] [PubMed]
- Messias-Reason, I.J.; Schafranski, M.D.; Kremsner, P.G.; Kun, J.F. Ficolin 2 (FCN2) functional polymorphisms and the risk of rheumatic fever and rheumatic heart disease. Clin. Exp. Immunol. 2009, 157, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.J.; Vannberg, F.O.; Khor, C.C.; Segal, S.; Moore, C.E.; Knox, K.; Day, N.P.; Davies, R.J.; Crook, D.W.; Hill, A.V. Functional polymorphisms in the FCN2 gene are not associated with invasive pneumococcal disease. Mol. Immunol. 2007, 44, 3267–3270. [Google Scholar] [CrossRef] [PubMed]
- Ruskamp, J.M.; Hoekstra, M.O.; Postma, D.S.; Kerkhof, M.; Bottema, R.W.; Koppelman, G.H.; Rovers, M.M.; Wijga, A.H.; de Jongste, J.C.; Brunekreef, B.; et al. Exploring the role of polymorphisms in ficolin genes in respiratory tract infections in children. Clin. Exp. Immunol. 2009, 155, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Boldt, A.B.; Sanchez, M.I.; Stahlke, E.R.; Steffensen, R.; Thiel, S.; Jensenius, J.C.; Prevedello, F.C.; Mira, M.T.; Kun, J.F.; Messias-Reason, I.J. Susceptibility to leprosy is associated with M-ficolin polymorphisms. J. Clin. Immunol. 2013, 33, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Vander Cruyssen, B.; Nuytinck, L.; Boullart, L.; Elewaut, D.; Waegeman, W.; van Thielen, M.; de Meester, E.; Lebeer, K.; Rossau, R.; de Keyser, F. Polymorphisms in the ficolin 1 gene (FCN1) are associated with susceptibility to the development of rheumatoid arthritis. Rheumatology (Oxford) 2007, 46, 1792–1795. [Google Scholar] [CrossRef]
- Urban, T.J.; Thompson, A.J.; Bradrick, S.S.; Fellay, J.; Schuppan, D.; Cronin, K.D.; Hong, L.; McKenzie, A.; Patel, K.; Shianna, K.V.; et al. IL28B genotype is associated with differential expression of intrahepatic interferon-stimulated genes in patients with chronic hepatitis C. Hepatology 2010, 52, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J.; Li, J.H.; Thompson, A.; Suchindran, S.; Lao, X.Q.; Patel, K.; Tillmann, H.L.; Muir, A.J.; McHutchison, J.G. Replicated association between an IL28B gene variant and a sustained response to pegylated interferon and ribavirin. Gastroenterology 2010, 138, 2307–2314. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Luo, F.L.; Fu, J.L.; Chen, T.L.; Wu, S.M.; Zhou, Y.D.; Zhang, X.L. Early increased ficolin-2 concentrations are associated with severity of liver inflammation and efficacy of anti-viral therapy in chronic hepatitis C patients. Scand. J. Immunol. 2013, 77, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Cedzynski, M.; Atkinson, A.P.; St Swierzko, A.; MacDonald, S.L.; Szala, A.; Zeman, K.; Buczylko, K.; Bak-Romaniszyn, L.; Wiszniewska, M.; Matsushita, M.; et al. L-ficolin (ficolin-2) insufficiency is associated with combined allergic and infectious respiratory disease in children. Mol. Immunol. 2009, 47, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Faik, I.; Oyedeji, S.I.; Idris, Z.; de Messias-Reason, I.J.; Lell, B.; Kremsner, P.G.; Kun, J.F. Ficolin-2 levels and genetic polymorphisms of FCN2 in malaria. Hum. Immunol. 2011, 72, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Doni, A.; Skjoedt, M.O.; Honore, C.; Arendrup, M.; Mantovani, A.; Garred, P. Heterocomplexes of mannose-binding lectin and the pentraxins PTX3 or serum amyloid P component trigger cross-activation of the complement system. J. Biol. Chem. 2011, 286, 3405–3417. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Zhang, J.; Tan, N.S.; Ho, B.; Ding, J.L. Natural IgG antibodies provide innate protection against ficolin-opsonized bacteria. EMBO J. 2013, 32, 2905–2919. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Zhang, J.; Yang, L.; Anand, G.S.; Ding, J.L. Molecular interaction between natural IgG and ficolin—Mechanistic insights on adaptive-innate immune crosstalk. Sci. Rep. 2014, 4, 3675. [Google Scholar] [CrossRef] [PubMed]
- Gout, E.; Moriscot, C.; Doni, A.; Dumestre-Perard, C.; Lacroix, M.; Perard, J.; Schoehn, G.; Mantovani, A.; Arlaud, G.J.; Thielens, N.M. M-ficolin interacts with the long pentraxin PTX3: A novel case of cross-talk between soluble pattern-recognition molecules. J. Immunol. 2011, 186, 5815–5822. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Doni, A.; Romani, L.; Jurgensen, H.J.; Behrendt, N.; Mantovani, A.; Garred, P. Ficolin-1-PTX3 complex formation promotes clearance of altered self-cells and modulates IL-8 production. J. Immunol. 2013, 191, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Koh, J.; Lu, J.; Thiel, S.; Leong, B.S.; Sethi, S.; He, C.Y.; Ho, B.; Ding, J.L. Local inflammation induces complement crosstalk which amplifies the antimicrobial response. PLoS Pathog. 2009, 5, e1000282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, L.; Anand, G.S.; Ho, B.; Ding, J.L. Pathophysiological condition changes the conformation of a flexible FBG-related protein, switching it from pathogen-recognition to host-interaction. Biochimie 2011, 93, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, D.; Haak, S.; Sisirak, V.; Reizis, B. The role of dendritic cells in autoimmunity. Nat. Rev. Immunol. 2013, 13, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Adema, G.J.; van Kooyk, Y.; Figdor, C.G. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell 2000, 100, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Fadden, A.J.; Drickamer, K. A novel mechanism of carbohydrate recognition by the C-type lectins DC-SIGN and DC-SIGNR. Subunit organization and binding to multivalent ligands. J. Biol. Chem. 2001, 276, 28939–28945. [Google Scholar] [CrossRef] [PubMed]
- Manzo, C.; Torreno-Pina, J.A.; Joosten, B.; Reinieren-Beeren, I.; Gualda, E.J.; Loza-Alvarez, P.; Figdor, C.G.; Garcia-Parajo, M.F.; Cambi, A. The neck region of the C-type lectin DC-SIGN regulates its surface spatiotemporal organization and virus-binding capacity on antigen-presenting cells. J. Biol. Chem. 2012, 287, 38946–38955. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.S.; Gregorio, G.; Bitton, N.; Hendrickson, W.A.; Littman, D.R. DC-SIGN-mediated internalization of HIV is required for trans-enhancement of T cell infection. Immunity 2002, 16, 135–144. [Google Scholar] [CrossRef]
- Soilleux, E.J.; Barten, R.; Trowsdale, J. DC-SIGN; a related gene, DC-SIGNR; and CD23 form a cluster on 19p13. J. Immunol. 2000, 165, 2937–2942. [Google Scholar] [CrossRef] [PubMed]
- Baribaud, F.; Pohlmann, S.; Leslie, G.; Mortari, F.; Doms, R.W. Quantitative expression and virus transmission analysis of DC-SIGN on monocyte-derived dendritic cells. J. Virol. 2002, 76, 9135–9142. [Google Scholar] [CrossRef] [PubMed]
- Pӧhlmann, S.; Baribaud, F.; Lee, B.; Leslie, G.J.; Sanchez, M.D.; Hiebenthal-Millow, K.; Munch, J.; Kirchhoff, F.; Doms, R.W. DC-SIGN interactions with human immunodeficiency virus type 1 and 2 and simian immunodeficiency virus. J. Virol. 2001, 75, 4664–4672. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Simmons, G.; Pohlmann, S.; Baribaud, F.; Ni, H.; Leslie, G.J.; Haggarty, B.S.; Bates, P.; Weissman, D.; Hoxie, J.A.; et al. Differential N-linked glycosylation of human immunodeficiency virus and Ebola virus envelope glycoproteins modulates interactions with DC-SIGN and DC-SIGNR. J. Virol. 2003, 77, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.T. Targeting early infection to prevent HIV-1 mucosal transmission. Nature 2010, 464, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Kinter, A.; Broder, C.C.; Dimitrov, D.S. Interactions of CCR5 and CXCR4 with CD4 and gp120 in human blood monocyte-derived dendritic cells. Exp. Mol. Pathol. 2000, 68, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Moris, A.; Nobile, C.; Buseyne, F.; Porrot, F.; Abastado, J.P.; Schwartz, O. DC-SIGN promotes exogenous MHC-I-restricted HIV-1 antigen presentation. Blood 2004, 103, 2648–2654. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.L.; Ganesh, L.; Leung, K.; Jongstra-Bilen, J.; Jongstra, J.; Nabel, G.J. Leukocyte-specific protein 1 interacts with DC-SIGN and mediates transport of HIV to the proteasome in dendritic cells. J. Exp. Med. 2007, 204, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Gringhuis, S.I.; van der Vlist, M.; van den Berg, L.M.; den Dunnen, J.; Litjens, M.; Geijtenbeek, T.B. HIV-1 exploits innate signaling by TLR8 and DC-SIGN for productive infection of dendritic cells. Nat. Immunol. 2010, 11, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, K.; Wang, Y.; Scala, C.; Jeffs, S.; Longstaff, C.; Stieh, D.; Haggarty, B.; Vanham, G.; Schols, D.; Balzarini, J.; et al. DC-SIGN increases the affinity of HIV-1 envelope glycoprotein interaction with CD4. PLoS One 2011, 6, e28307. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Leslie, G.; Soilleux, E.; O’Doherty, U.; Baik, S.; Levroney, E.; Flummerfelt, K.; Swiggard, W.; Coleman, N.; Malim, M.; et al. cis Expression of DC-SIGN allows for more efficient entry of human and simian immunodeficiency viruses via CD4 and a coreceptor. J. Virol. 2001, 75, 12028–12038. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, K.; Kim, Y.; Latinovic, O.; Morozov, V.; Melikyan, G.B. HIV enters cells via endocytosis and dynamin-dependent fusion with endosomes. Cell 2009, 137, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Van Montfort, T.; Eggink, D.; Boot, M.; Tuen, M.; Hioe, C.E.; Berkhout, B.; Sanders, R.W. HIV-1 N-glycan composition governs a balance between dendritic cell-mediated viral transmission and antigen presentation. J. Immunol. 2011, 187, 4676–4685. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Wu, L.; Bohks, S.M.; KewalRamani, V.N.; Unutmaz, D.; Hope, T.J. Recruitment of HIV and its receptors to dendritic cell-T cell junctions. Science 2003, 300, 1295–1297. [Google Scholar] [CrossRef] [PubMed]
- Sol-Foulon, N.; Moris, A.; Nobile, C.; Boccaccio, C.; Engering, A.; Abastado, J.P.; Heard, J.M.; van Kooyk, Y.; Schwartz, O. HIV-1 Nef-induced upregulation of DC-SIGN in dendritic cells promotes lymphocyte clustering and viral spread. Immunity 2002, 16, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Cavrois, M.; Neidleman, J.; Kreisberg, J.F.; Greene, W.C. In vitro derived dendritic cells trans-infect CD4 T cells primarily with surface-bound HIV-1 virions. PLoS Pathog. 2007, 3, e4. [Google Scholar] [CrossRef] [PubMed]
- Hodges, A.; Sharrocks, K.; Edelmann, M.; Baban, D.; Moris, A.; Schwartz, O.; Drakesmith, H.; Davies, K.; Kessler, B.; McMichael, A.; et al. Activation of the lectin DC-SIGN induces an immature dendritic cell phenotype triggering Rho-GTPase activity required for HIV-1 replication. Nat. Immunol. 2007, 8, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Useros, N.; Lorizate, M.; Puertas, M.C.; Rodriguez-Plata, M.T.; Zangger, N.; Erikson, E.; Pino, M.; Erkizia, I.; Glass, B.; Clotet, B.; et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLoS Biol. 2012, 10, e1001448. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hwang, S.L.; Chan, V.S.; Chung, N.P.; Wang, S.R.; Li, Z.; Ma, J.; Lin, C.W.; Hsieh, Y.J.; Chang, K.P.; et al. Binding of HIV-1 gp120 to DC-SIGN promotes ASK-1-dependent activation-induced apoptosis of human dendritic cells. PLoS Pathog. 2013, 9, e1003100. [Google Scholar] [CrossRef] [PubMed]
- De Witte, L.; de Vries, R.D.; van der Vlist, M.; Yuksel, S.; Litjens, M.; de Swart, R.L.; Geijtenbeek, T.B. DC-SIGN and CD150 have distinct roles in transmission of measles virus from dendritic cells to T-lymphocytes. PLoS Pathog. 2008, 4, e1000049. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.P.; Lasala, F.; Carrillo, J.; Muniz, O.; Corbi, A.L.; Delgado, R. C-type lectins DC-SIGN and L-SIGN mediate cellular entry by Ebola virus in cis and in trans. J. Virol. 2002, 76, 6841–6844. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Huang, Y.; Ganesh, L.; Leung, K.; Kong, W.P.; Schwartz, O.; Subbarao, K.; Nabel, G.J. pH-dependent entry of severe acute respiratory syndrome coronavirus is mediated by the spike glycoprotein and enhanced by dendritic cell transfer through DC-SIGN. J. Virol. 2004, 78, 5642–5650. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.F.; Huang, J.C.; Lee, Y.M.; Liu, S.J.; Chan, Y.J.; Chau, Y.P.; Chong, P.; Chen, Y.M. DC-SIGN mediates avian H5N1 influenza virus infection in cis and in trans. Biochem. Biophys. Res. Commun. 2008, 373, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Pӧhlmann, S.; Zhang, J.; Baribaud, F.; Chen, Z.; Leslie, G.J.; Lin, G.; Granelli-Piperno, A.; Doms, R.W.; Rice, C.M.; McKeating, J.A. Hepatitis C virus glycoproteins interact with DC-SIGN and DC-SIGNR. J. Virol. 2003, 77, 4070–4080. [Google Scholar] [CrossRef] [PubMed]
- Halary, F.; Amara, A.; Lortat-Jacob, H.; Messerle, M.; Delaunay, T.; Houles, C.; Fieschi, F.; Arenzana-Seisdedos, F.; Moreau, J.F.; Dechanet-Merville, J. Human cytomegalovirus binding to DC-SIGN is required for dendritic cell infection and target cell trans-infection. Immunity 2002, 17, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Rappocciolo, G.; Hensler, H.R.; Jais, M.; Reinhart, T.A.; Pegu, A.; Jenkins, F.J.; Rinaldo, C.R. Human herpesvirus 8 infects and replicates in primary cultures of activated B lymphocytes through DC-SIGN. J. Virol. 2008, 82, 4793–4806. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, I.S.; Lekkerkerker, A.N.; Depla, E.; Bosman, F.; Musters, R.J.; Depraetere, S.; van Kooyk, Y.; Geijtenbeek, T.B. Hepatitis C virus targets DC-SIGN and L-SIGN to escape lysosomal degradation. J. Virol. 2004, 78, 8322–8332. [Google Scholar] [CrossRef] [PubMed]
- Pӧhlmann, S.; Soilleux, E.J.; Baribaud, F.; Leslie, G.J.; Morris, L.S.; Trowsdale, J.; Lee, B.; Coleman, N.; Doms, R.W. DC-SIGNR, a DC-SIGN homologue expressed in endothelial cells, binds to human and simian immunodeficiency viruses and activates infection in trans. Proc. Natl. Acad. Sci. USA 2001, 98, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Marzi, A.; Gramberg, T.; Simmons, G.; Moller, P.; Rennekamp, A.J.; Krumbiegel, M.; Geier, M.; Eisemann, J.; Turza, N.; Saunier, B.; et al. DC-SIGN and DC-SIGNR interact with the glycoprotein of Marburg virus and the S protein of severe acute respiratory syndrome coronavirus. J. Virol. 2004, 78, 12090–12095. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, L.B.; Patin, E.; Neyrolles, O.; Cann, H.M.; Gicquel, B.; Quintana-Murci, L. The heritage of pathogen pressures and ancient demography in the human innate-immunity CD209/CD209L region. Am. J. Hum. Genet. 2005, 77, 869–886. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.W.; Nguyen, H.Y.; Hanna, S.L.; Sanchez, M.D.; Doms, R.W.; Pierson, T.C. West Nile virus discriminates between DC-SIGN and DC-SIGNR for cellular attachment and infection. J. Virol 2006, 80, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Mummidi, S.; Catano, G.; Lam, L.; Hoefle, A.; Telles, V.; Begum, K.; Jimenez, F.; Ahuja, S.S.; Ahuja, S.K. Extensive repertoire of membrane-bound and soluble dendritic cell-specific ICAM-3-grabbing nonintegrin 1 (DC-SIGN1) and DC-SIGN2 isoforms. Inter-individual variation in expression of DC-SIGN transcripts. J. Biol. Chem. 2001, 276, 33196–33212. [Google Scholar] [CrossRef] [PubMed]
- Plazolles, N.; Humbert, J.M.; Vachot, L.; Verrier, B.; Hocke, C.; Halary, F. Pivotal advance: The promotion of soluble DC-SIGN release by inflammatory signals and its enhancement of cytomegalovirus-mediated cis-infection of myeloid dendritic cells. J. Leukoc. Biol. 2011, 89, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Gómez, D.; Sierra-Filardi, E.; Martinez-Nunez, R.T.; Caparros, E.; Delgado, R.; Munoz-Fernandez, M.A.; Abad, M.A.; Jimenez-Barbero, J.; Leal, M.; Corbi, A.L. Structural requirements for multimerization of the pathogen receptor dendritic cell-specific ICAM3-grabbing non-integrin (CD209) on the cell surface. J. Biol. Chem. 2008, 283, 3889–3903. [Google Scholar] [CrossRef] [PubMed]
- Alagarasu, K.; Selvaraj, P.; Swaminathan, S.; Raghavan, S.; Narendran, G.; Narayanan, P.R. CCR2, MCP-1, SDF-1a & DC-SIGN gene polymorphisms in HIV-1 infected patients with & without tuberculosis. Indian J. Med. Res. 2009, 130, 444–450. [Google Scholar] [PubMed]
- Zhang, J.; Zhang, X.; Fu, J.; Bi, Z.; Arheart, K.L.; Barreiro, L.B.; Quintana-Murci, L.; Pahwa, S.; Liu, H. Protective role of DC-SIGN (CD209) neck-region alleles with <5 repeat units in HIV-1 transmission. J. Infect. Dis. 2008, 198, 68–71. [Google Scholar]
- Liu, H.; Hwangbo, Y.; Holte, S.; Lee, J.; Wang, C.; Kaupp, N.; Zhu, H.; Celum, C.; Corey, L.; McElrath, M.J.; et al. Analysis of genetic polymorphisms in CCR5, CCR2, stromal cell-derived factor-1, RANTES, and dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin in seronegative individuals repeatedly exposed to HIV-1. J. Infect. Dis. 2004, 190, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.G.; Hildreth, J.E. Involvement of macrophage mannose receptor in the binding and transmission of HIV by macrophages. Eur. J. Immunol. 2003, 33, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, J.R.; Rogers, R.; Molina, R.M.; Dangond, F.; McLane, M.F.; Essex, M.; Brain, J.D. Noninfectious entry of HIV-1 into peripheral and brain macrophages mediated by the mannose receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 5097–5102. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, K.L.; Sastry, K.N.; Chang, D.; White, M.R.; Crouch, E.C. Enhanced anti-influenza activity of a surfactant protein D and serum conglutinin fusion protein. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, L90–L98. [Google Scholar] [PubMed]
- Yang, L.; Hu, S.; Chroneos, Z. Targeting of the surfactant protein A receptor SP-R210L variant by influenza A virus in macrophages. (INM8P.440). J. Immunol. 2014, 192, 124–126. [Google Scholar]
- Pandit, H.; Gopal, S.; Sonawani, A.; Yadav, A.K.; Qaseem, A.S.; Warke, H.; Patil, A.; Gajbhiye, R.; Kulkarni, V.; Al-Mozaini, M.A.; et al. Surfactant protein D inhibits HIV-1 infection of target cells via interference with gp120-CD4 interaction and modulates pro-inflammatory cytokine production. PLoS One 2014, 9, e102395. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.; Gaiha, G.D.; Palaniyar, N.; Dong, T.; Mitchell, D.A.; Clark, H.W. Surfactant Protein D modulates HIV infection of both T-cells and dendritic cells. PLoS One 2013, 8, e59047. [Google Scholar] [CrossRef] [PubMed]
- Gaiha, G.D.; Dong, T.; Palaniyar, N.; Mitchell, D.A.; Reid, K.B.; Clark, H.W. Surfactant protein A binds to HIV and inhibits direct infection of CD4+ cells, but enhances dendritic cell-mediated viral transfer. J. Immunol. 2008, 181, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nagai, K.; Tsutsumi, H.; Kuroki, Y. Lactoferrin and surfactant protein A exhibit distinct binding specificity to F protein and differently modulate respiratory syncytial virus infection. Eur. J. Immunol. 2003, 33, 2894–2902. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, C.; Manya, H.; Ouellet, M.; Clark, G.F.; Endo, T.; Tremblay, M.J.; Sato, S. Host-soluble galectin-1 promotes HIV-1 replication through a direct interaction with glycans of viral gp120 and host CD4. J. Virol. 2011, 85, 11742–11751. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Hong, P.W.; Lee, B.; Baum, L.G. Galectin-9 binding to cell surface protein disulfide isomerase regulates the redox environment to enhance T-cell migration and HIV entry. Proc. Natl. Acad. Sci. USA 2011, 108, 10650–10655. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.F.; Tsao, C.H.; Lin, Y.T.; Hsu, D.K.; Chiang, M.L.; Lo, C.H.; Chien, F.C.; Chen, P.; Arthur Chen, Y.M.; Chen, H.Y.; et al. Galectin-3 promotes HIV-1 budding via association with Alix and Gag p6. Glycobiology 2014, 24, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- De Witte, L.; Nabatov, A.; Pion, M.; Fluitsma, D.; de Jong, M.A.; de Gruijl, T.; Piguet, V.; van Kooyk, Y.; Geijtenbeek, T.B. Langerin is a natural barrier to HIV-1 transmission by Langerhans cells. Nat. Med. 2007, 13, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Van der Vlist, M.; de Witte, L.; de Vries, R.D.; Litjens, M.; de Jong, M.A.; Fluitsma, D.; de Swart, R.L.; Geijtenbeek, T.B. Human Langerhans cells capture measles virus through Langerin and present viral antigens to CD4(+) T cells but are incapable of cross-presentation. Eur. J. Immunol. 2011, 41, 2619–2631. [Google Scholar] [CrossRef] [PubMed]
- Valdimarsson, H. Infusion of plasma-derived mannan-binding lectin (MBL) into MBL-deficient humans. Biochem. Soc. Trans. 2003, 31, 768–769. [Google Scholar] [CrossRef] [PubMed]
- Laursen, I. Mannan-binding lectin (MBL) production from human plasma. Biochem. Soc. Trans. 2003, 31, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Jensenius, J.C.; Jensen, P.H.; McGuire, K.; Larsen, J.L.; Thiel, S. Recombinant mannan-binding lectin (MBL) for therapy. Biochem. Soc. Trans. 2003, 31, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.A.; Matthiesen, F.; Agger, T.; Kongerslev, L.; Thiel, S.; Cornelissen, K.; Axelsen, M. Phase I safety, tolerability, and pharmacokinetic study of recombinant human mannan-binding lectin. J. Clin. Immunol. 2006, 26, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Michelow, I.C.; Dong, M.; Mungall, B.A.; Yantosca, L.M.; Lear, C.; Ji, X.; Karpel, M.; Rootes, C.L.; Brudner, M.; Houen, G.; et al. A novel L-ficolin/mannose-binding lectin chimeric molecule with enhanced activity against Ebola virus. J. Biol. Chem. 2010, 285, 24729–24739. [Google Scholar] [CrossRef] [PubMed]
- White, M.R.; Crouch, E.; Chang, D.; Sastry, K.; Guo, N.; Engelich, G.; Takahashi, K.; Ezekowitz, R.A.; Hartshorn, K.L. Enhanced antiviral and opsonic activity of a human mannose-binding lectin and surfactant protein D chimera. J. Immunol. 2000, 165, 2108–2115. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Moyo, P.; Chigweshe, L.; Chang, W.C.; White, M.R.; Hartshorn, K.L. Efficacy of recombinant chimeric lectins, consisting of mannose binding lectin and L-ficolin, against influenza A viral infection in mouse model study. Virus Res. 2013, 178, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Hartshorn, K.L.; White, M.R.; Moyo, P.; Michelow, I.C.; Koziel, H.; Kinane, B.T.; Schmidt, E.V.; Fujita, T.; Takahashi, K. Recombinant chimeric lectins consisting of mannose-binding lectin and L-ficolin are potent inhibitors of influenza A virus compared with mannose-binding lectin. Biochem. Pharmacol. 2011, 81, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Crouch, E.; Nikolaidis, N.; McCormack, F.X.; McDonald, B.; Allen, K.; Rynkiewicz, M.J.; Cafarella, T.M.; White, M.; Lewnard, K.; Leymarie, N.; et al. Mutagenesis of surfactant protein D informed by evolution and x-ray crystallography enhances defenses against influenza A virus in vivo. J. Biol. Chem. 2011, 286, 40681–40692. [Google Scholar] [CrossRef] [PubMed]
- Hillaire, M.L.; van Eijk, M.; Vogelzang-van Trierum, S.E.; Fouchier, R.A.; Osterhaus, A.D.; Haagsman, H.P.; Rimmelzwaan, G.F. Recombinant porcine surfactant protein D inhibits influenza A virus replication ex vivo. Virus Res. 2014, 181, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Tarr, A.W.; Urbanowicz, R.A.; Ball, J.K. The role of humoral innate immunity in hepatitis C virus infection. Viruses 2012, 4, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Marzi, A.; Mitchell, D.A.; Chaipan, C.; Fisch, T.; Doms, R.W.; Carrington, M.; Desrosiers, R.C.; Pohlmann, S. Modulation of HIV and SIV neutralization sensitivity by DC-SIGN and mannose-binding lectin. Virology 2007, 368, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Boross, P.; Leusen, J.H. Boosting antibody therapy with complement. Blood 2012, 119, 5945–5947. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, K.B.; Gray, E.S.; Mufhandu, H.; McMahon, J.B.; Chakauya, E.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. The lectins griffithsin, cyanovirin-N and scytovirin inhibit HIV-1 binding to the DC-SIGN receptor and transfer to CD4(+) cells. Virology 2012, 423, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Francois, K.O.; van Laethem, K.; Hoorelbeke, B.; Renders, M.; Auwerx, J.; Liekens, S.; Oki, T.; Igarashi, Y.; Schols, D. Pradimicin S, a highly soluble nonpeptidic small-size carbohydrate-binding antibiotic, is an anti-HIV drug lead for both microbicidal and systemic use. Antimicrob. Agents Chemother. 2010, 54, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Buffa, V.; Stieh, D.; Mamhood, N.; Hu, Q.; Fletcher, P.; Shattock, R.J. Cyanovirin-N potently inhibits human immunodeficiency virus type 1 infection in cellular and cervical explant models. J. Gen. Virol. 2009, 90, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Emau, P.; Jiang, Y.; Agy, M.B.; Shattock, R.J.; Schmidt, A.; Morton, W.R.; Gustafson, K.R.; Boyd, M.R. Cyanovirin-N inhibits AIDS virus infections in vaginal transmission models. AIDS Res. Hum. Retrovir. 2004, 20, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Whitley, M.J.; Furey, W.; Kollipara, S.; Gronenborn, A.M. Burkholderia oklahomensis agglutinin is a canonical two-domain OAA-family lectin: Structures, carbohydrate binding and anti-HIV activity. FEBS J. 2013, 280, 2056–2067. [Google Scholar] [CrossRef] [PubMed]
- Ferir, G.; Huskens, D.; Noppen, S.; Koharudin, L.M.; Gronenborn, A.M.; Schols, D. Broad anti-HIV activity of the Oscillatoria agardhii agglutinin homologue lectin family. J. Antimicrob. Chemother. 2014, 69, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lagenaur, L.A.; Simpson, D.A.; Essenmacher, K.P.; Frazier-Parker, C.L.; Liu, Y.; Tsai, D.; Rao, S.S.; Hamer, D.H.; Parks, T.P.; et al. Engineered vaginal lactobacillus strain for mucosal delivery of the human immunodeficiency virus inhibitor cyanovirin-N. Antimicrob. Agents Chemother. 2006, 50, 3250–3259. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Furey, W.; Gronenborn, A.M. Novel fold and carbohydrate specificity of the potent anti-HIV cyanobacterial lectin from Oscillatoria agardhii. J. Biol. Chem. 2011, 286, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Verhoye, L.; Wychowski, C.; Leroux-Roels, G.; Palmer, K.E.; Dubuisson, J. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-spectrum in vitro activity and in vivo efficacy of the antiviral protein griffithsin against emerging viruses of the family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Ishag, H.Z.; Li, C.; Huang, L.; Sun, M.X.; Wang, F.; Ni, B.; Malik, T.; Chen, P.Y.; Mao, X. Griffithsin inhibits Japanese encephalitis virus infection in vitro and in vivo. Arch. Virol. 2013, 158, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.; Shirakura, M.; et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis C virus. PLoS One 2013, 8, e64449. [Google Scholar] [CrossRef] [PubMed]
- Helle, F.; Wychowski, C.; Vu-Dac, N.; Gustafson, K.R.; Voisset, C.; Dubuisson, J. Cyanovirin-N inhibits hepatitis C virus entry by binding to envelope protein glycans. J. Biol. Chem. 2006, 281, 25177–25183. [Google Scholar] [CrossRef] [PubMed]
- Dey, B.; Lerner, D.L.; Lusso, P.; Boyd, M.R.; Elder, J.H.; Berger, E.A. Multiple antiviral activities of cyanovirin-N: Blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Bailey, K.W.; Wong, M.H.; O’Keefe, B.R.; Gustafson, K.R.; Mishin, V.P.; Gubareva, L.V. Treatment of influenza A (H1N1) virus infections in mice and ferrets with cyanovirin-N. Antivir. Res. 2008, 80, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Bertaux, C.; Daelemans, D.; Meertens, L.; Cormier, E.G.; Reinus, J.F.; Peumans, W.J.; van Damme, E.J.; Igarashi, Y.; Oki, T.; Schols, D.; et al. Entry of hepatitis C virus and human immunodeficiency virus is selectively inhibited by carbohydrate-binding agents but not by polyanions. Virology 2007, 366, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [PubMed]
- Chen, J.; Huang, D.; Chen, W.; Guo, C.; Wei, B.; Wu, C.; Peng, Z.; Fan, J.; Hou, Z.; Fang, Y.; et al. Linker-extended native cyanovirin-N facilitates PEGylation and potently inhibits HIV-1 by targeting the glycan ligand. PLoS One 2014, 9, e86455. [Google Scholar] [CrossRef] [PubMed]
- Barton, C.; Kouokam, J.C.; Lasnik, A.B.; Foreman, O.; Cambon, A.; Brock, G.; Montefiori, D.C.; Vojdani, F.; McCormick, A.A.; O’Keefe, B.R.; et al. Activity of and effect of subcutaneous treatment with the broad-spectrum antiviral lectin griffithsin in two laboratory rodent models. Antimicrob. Agents Chemother. 2014, 58, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Fan, J.; Kitazato, K. The antiviral protein cyanovirin-N: The current state of its production and applications. Appl. Microbiol. Biotechnol. 2010, 86, 805–812. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mason, C.P.; Tarr, A.W. Human Lectins and Their Roles in Viral Infections. Molecules 2015, 20, 2229-2271. https://doi.org/10.3390/molecules20022229
Mason CP, Tarr AW. Human Lectins and Their Roles in Viral Infections. Molecules. 2015; 20(2):2229-2271. https://doi.org/10.3390/molecules20022229
Chicago/Turabian StyleMason, Christopher P., and Alexander W. Tarr. 2015. "Human Lectins and Their Roles in Viral Infections" Molecules 20, no. 2: 2229-2271. https://doi.org/10.3390/molecules20022229
APA StyleMason, C. P., & Tarr, A. W. (2015). Human Lectins and Their Roles in Viral Infections. Molecules, 20(2), 2229-2271. https://doi.org/10.3390/molecules20022229