Effectiveness of Phenolic Compounds against Citrus Green Mould




Abstract
:1. Introduction
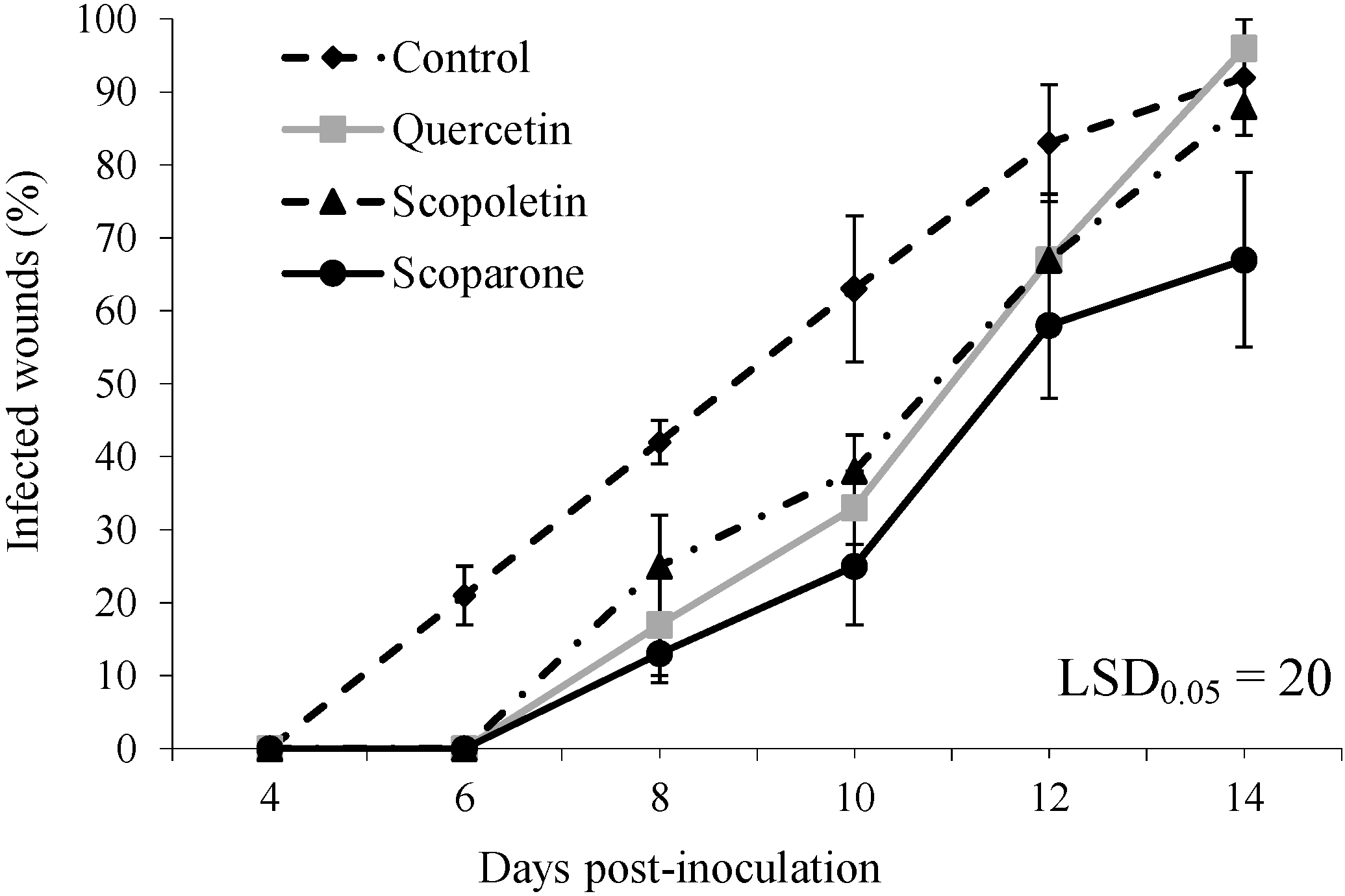
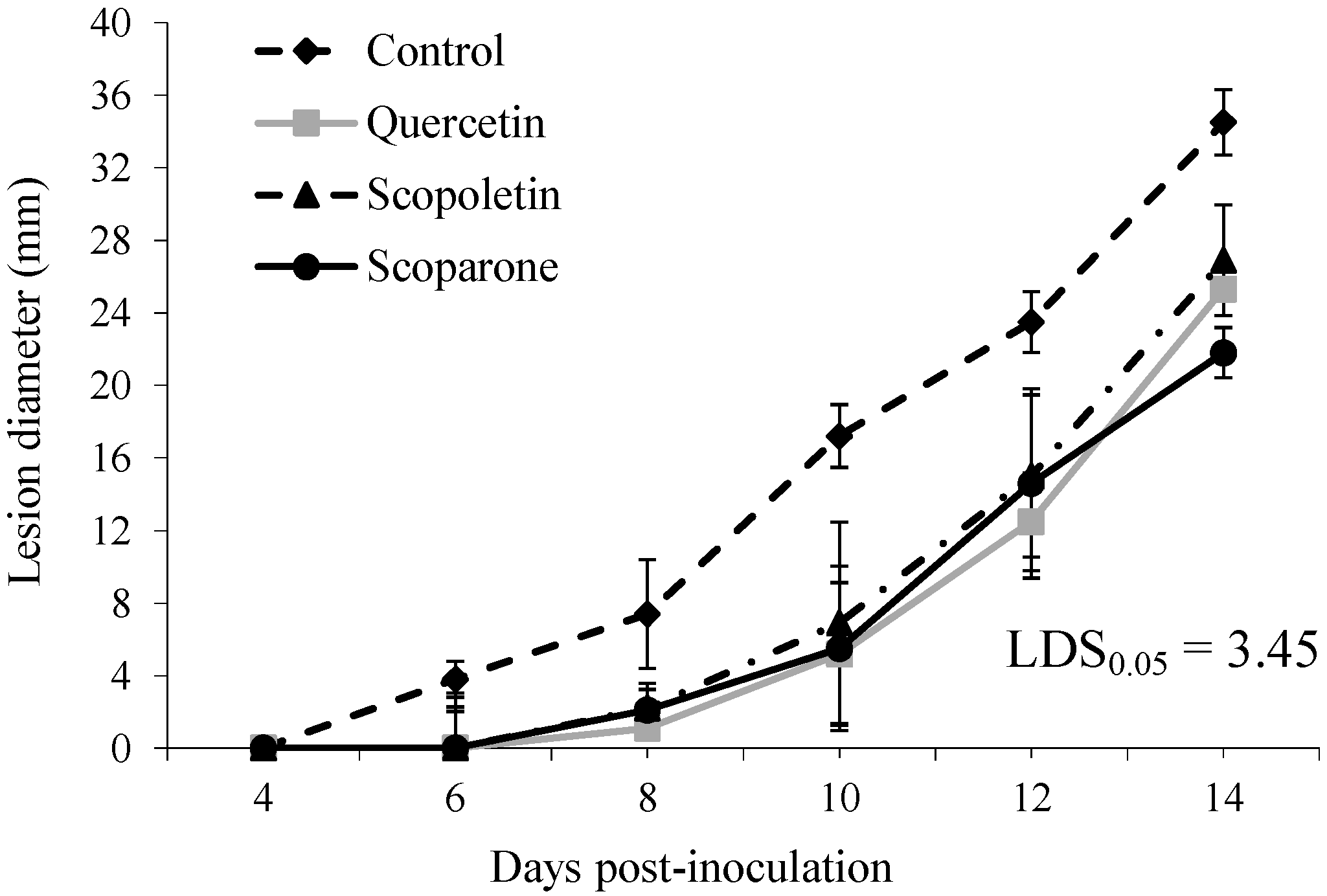
2. Results and Discussion

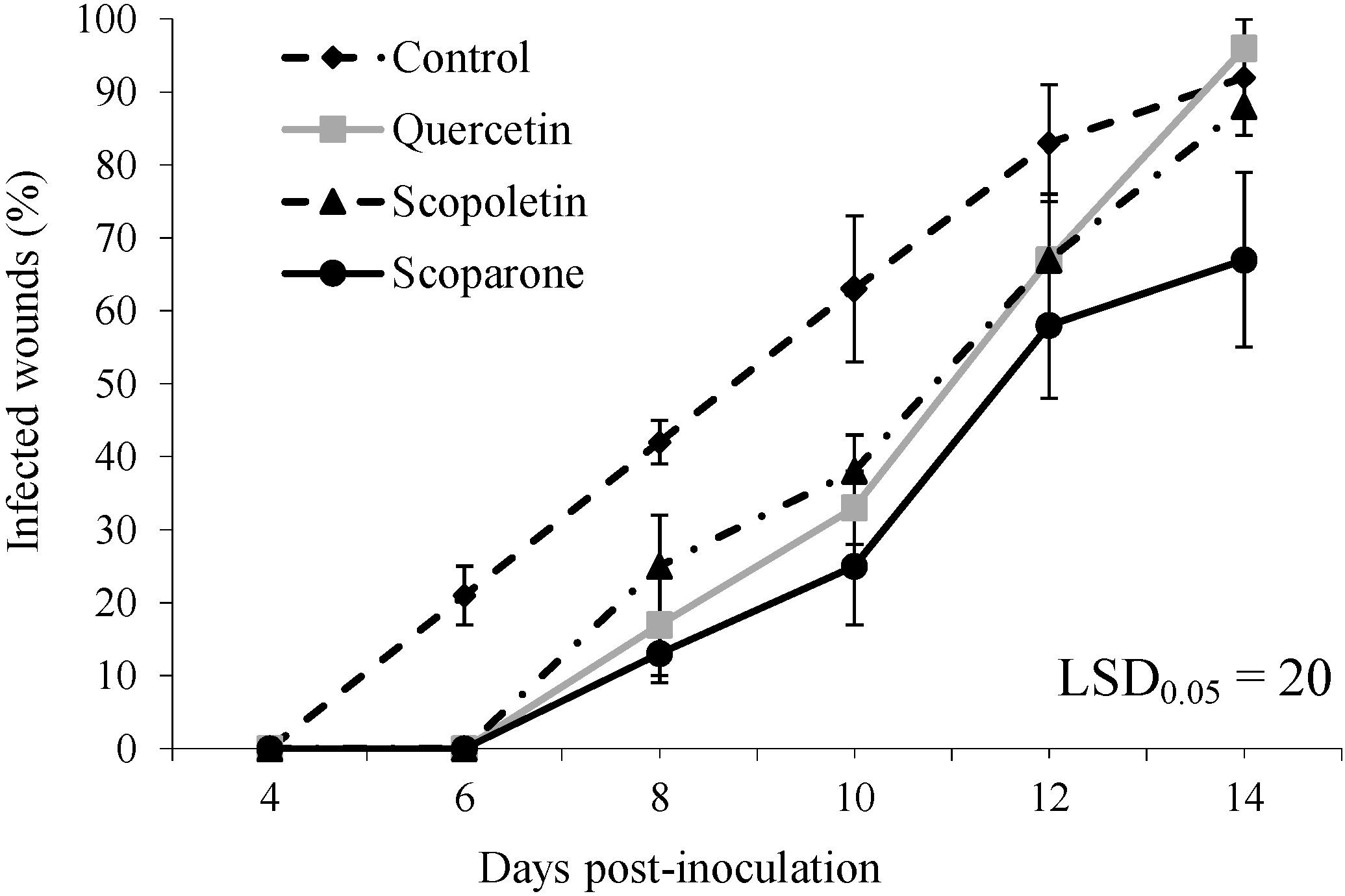{kind=link}
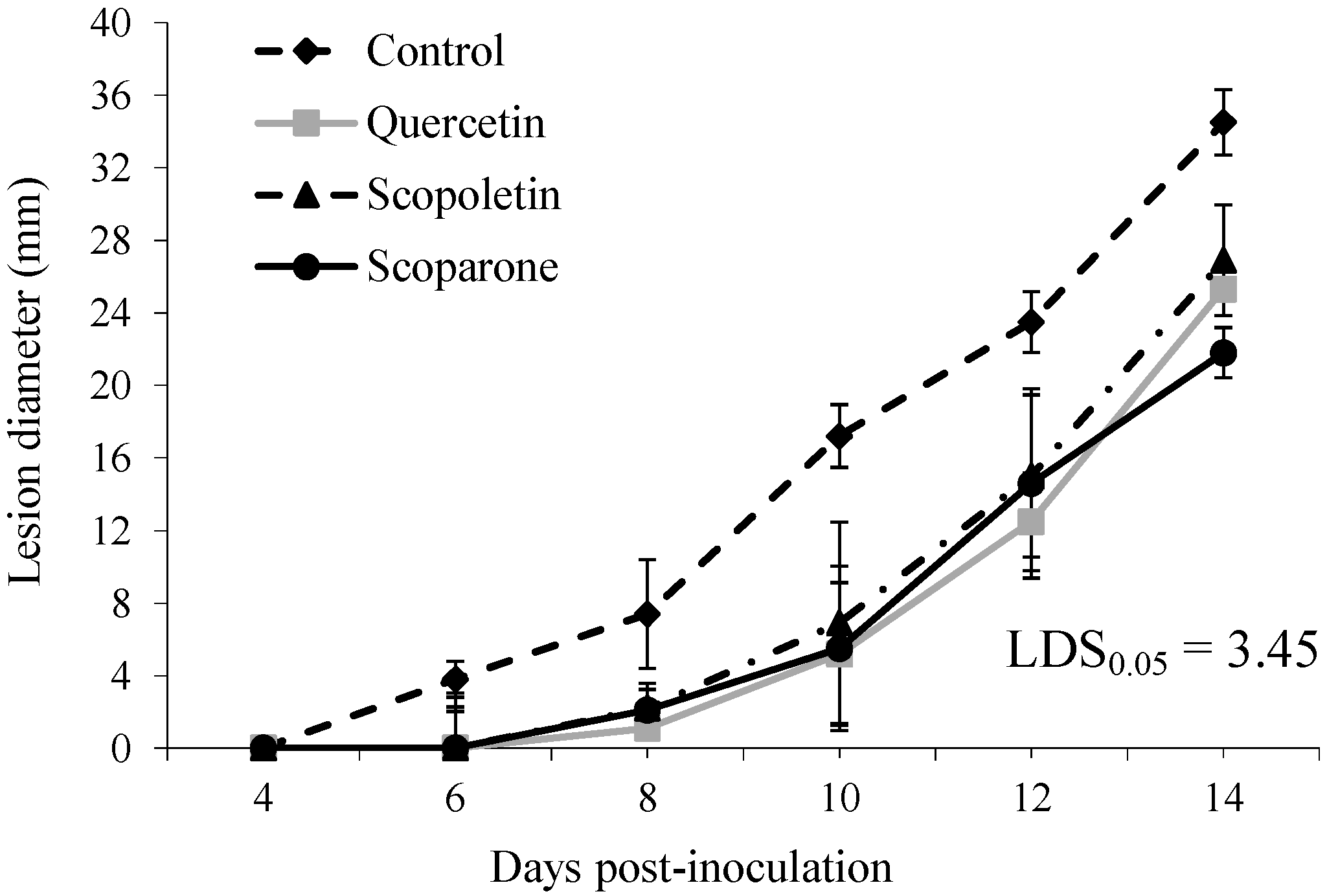{kind=link}
| Treatment | Colony Diameter (mm) | |
|---|---|---|
| 3 DPI | 6 DPI | |
| Control [no phenolics] | 14.7 ± 1.4 b,c | 54.2 ± 0.5 a,b |
| Quercetin 10 µg/mL | 16.3 ± 0.6 a,b | 56.5 ± 0.4 a |
| Quercetin 100 µg/mL | 17.8 ± 0.2 a | 47.7 ± 1.2 d |
| Scopoletin 10 µg/mL | 14.0 ± 0.4 c,d | 52.5 ± 1.1 b,c |
| Scopoletin 100 µg/mL | 13.0 ± 0.8 c,d | 51.5 ± 1.8 c |
| Scoparone 10 µg/mL | 14.7 ± 0.5 b,c | 54.7 ± 1.2 a,b |
| Scoparone 100 µg/mL | 12.8 ± 0.6 d | 50.3 ± 0.9 c |
3. Experimental Section
3.1. Chemicals
3.2. Preparation of Phenolic Compounds Solutions
3.3. Penicillium Conidial Suspension
3.4. In Vitro Trials
3.5. In Vivo Trials
3.6. Statistical Analysis
3.7. Control Index Calculation
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Holmes, G.J.; Eckert, J.W. Sensitivity of Penicillium digitatum and P. italicum to postharvest citrus fungicides in California. Phytopathology 2001, 89, 716–721. [Google Scholar] [CrossRef]
- Plaza, P.; Usall, J.; Teixidó, N.; Viňas, I. Effect of water activity and temperature on germination and growth of Penicillium digitatum, P. italicum and Geotrichum candidum. J. Appl. Microbiol. 2003, 94, 549–554. [Google Scholar] [CrossRef]
- Youssef, K.; Ligorio, A.; Sanzani, S.M.; Nigro, F.; Ippolito, A. Investigations of Penicillium spp. population dynamic in citrus packinghouse. J. Plant Pathol. 2011, 93 (Suppl. 4), 61–62. [Google Scholar]
- Daferera, D.J.; Ziogas, B.N.; Polissiou, M.G. GC-MS analysis of essential oils from some Greek aromatic plants and their fungitoxicity on Penicillium digitatum. J. Agric. Food Chem. 2000, 48, 2576–2581. [Google Scholar] [CrossRef]
- Ippolito, A.; Sanzani, S.M. Control of postharvest decay by the integration of pre and postharvest application of nonchemical compounds. Acta Hortic. 2011, 905, 135–143. [Google Scholar]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Ismail, M.A.; Rouself, R.L.; Brown, G.E. Wound healing in citrus: Isolation and identification of 7-hydroxycoumarin [umbelliferone] from grapefruit flavedo and its effects on Penicillium digitatum Sacc. Hortic. Sci. 1978, 13, 358. [Google Scholar]
- Jeandet, P.; Clément, C.; Courot, E.; Cordelier, S. Modulation of phytoalexin biosynthesis in engineered plants for disease resistance. Int. J. Mol. Sci. 2013, 14, 14136–14170. [Google Scholar] [CrossRef]
- Youssef, K.; Sanzani, S.M.; Ligorio, A.; Ippolito, A.; Terry, L.A. Sodium carbonate and bicarbonate treatments induce resistance to postharvest green mould on citrus fruit. Postharvest Biol. Technol. 2014, 87, 61–69. [Google Scholar] [CrossRef]
- Ben-Yehoshoua, S.; Kim, J.J.; Shapiro, B. Elicitation of resistance to the development of decay in citrus fruit by curing of sealed fruit. Acta Hortic. 1989, 258, 623–630. [Google Scholar]
- Dubery, I.A.; Holzapfel, C.W.; Kruger, G.J.; Schabor, J.C.; Dyk, M.V. Characterization of a γ-radiation-induced antifungal stress metabolite in citrus peel. Phytochemistry 1988, 27, 2769–2772. [Google Scholar] [CrossRef]
- Kim, J.J.; Ben-Yehoshua, S.; Shapiro, B.; Henis, Y.; Carmeli, S. Accumulation of scoparone in heat-treated lemon fruit inoculated with Penicillium digitatum Sacc. Plant Physiol. 1991, 97, 880–885. [Google Scholar] [CrossRef]
- Gottstein, D.; Gross, D. Phytoalexins of woody plants. Trees 1992, 6, 55–68. [Google Scholar]
- Mayr, U.; Treutter, D.; Santos-Buelga, C.; Bauer, H.; Feucht, W. Developmental changes in the phenol concentrations of Golden Delicious apple fruits and leaves. Phytochemistry 1995, 38, 1151–1155. [Google Scholar] [CrossRef]
- Sanzani, S.M.; de Girolamo, A.; Schena, L.; Solfrizzo, M.; Ippolito, A.; Visconti, A. Control of Penicillium expansum and patulin accumulation on apples by quercetin and umbelliferone. Eur. Food Res. Technol. 2009, 228, 381–389. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.C.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A.M. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar]
- Shaw, C.Y.; Chen, C.H.; Hsu, C.C.; Chen, C.C.; Tsai, Y.C. Antioxidant properties of scopoletin isolated from Sinomonium acutum. Phytother. Res. 2003, 17, 823–825. [Google Scholar] [CrossRef]
- Sourivong, P.; Schronerová, K.; Babincova, M. Scoparone inhibits ultraviolet radiation-induced lipid peroxidation. Z. Naturforschung 2007, 62, 61–64. [Google Scholar]
- Rodov, V.; Ben-Yehoshua, S.; D’hallewin, G.; Castina, T.; Farg, D. Accumulation of the phytoalexins, scoparone and scopoletin in citrus fruit subjected to various postharvest treatments. Acta Hortic. 1994, 381, 517–524. [Google Scholar]
- Sanzani, S.M.; Montemurro, C.; di Rienzo, V.; Solfrizzo, M.; Ippolito, A. Genetic structure and natural variation associated with host of origin in Penicillium expansum strains causing blue mould. Int. J. Food Microbiol. 2013, 165, 111–120. [Google Scholar] [CrossRef]
- Afek, U.; Sztejnberg, A. Accumulation of scoparone, a phytoalexin associated with resistance of citrus to Phytophthora citrophthora. Phytopathology 1988, 78, 1678–1682. [Google Scholar] [CrossRef]
- Garcia, D.; Sanier, C.; Macheix, J.J.; D’Auzac, J. Accumulation of scopoletin in Hevea brasiliensis infected by Microcyclus ulei [P. Henn.] V. ARX and evaluation of its fungitoxcity for three leaf pathogens of rubber tree. Physiol. Mol. Plant Pathol. 1995, 47, 213–223. [Google Scholar] [CrossRef]
- Rodov, V.; Ben-Yehoshua, S.; Kim, J.J.; Shapiro, B.; Ittah, Y. Ultraviolet illumination induces scoparone production in kumquat and orange fruit and improve decay resistance. J. Am. Soc. Hortic. Sci. 1992, 117, 788–792. [Google Scholar]
- Rodov, V.; Burns, P.; Ben-Shalom, N.; Fluhr, R.; Ben-Yehoshua, S. Induced local disease resistance in Citrus mesocarp [albedo]: Accumulation of phytoalexins and PR-proteins. Proc. Int. Soc. Citric. 1996, 2, 1101–1104. [Google Scholar]
- Ortuño, A.; Díaz, L.; Alvarez, N.; Porras, I.; García-Lidón, A.; del Río, J.A. Comparative study of flavonoid and scoparone accumulation in different Citrus species and their susceptibility to Penicillium digitatum. Food Chem. 2011, 125, 232–239. [Google Scholar] [CrossRef]
- McRae, K.B.; Lidster, P.D.; Demarco, A.C.; Dick, A.J. Comparison of the polyphenol profiles of apple fruit cultivars by correspondence analysis. J. Sci. Food Agric. 1990, 50, 329–342. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chuang, Y.C.; Hsu, H.W. The flavonoid, carotenoid and pectin content in peels of citrus cultivated in Taiwan. Food Chem. 2008, 106, 277–284. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Schena, L.; de Girolamo, A.; Ippolito, A.; González-Candelas, L. Characterization of genes associated to induced resistance against Penicillium expansum in apple fruit treated with quercetin. Postharvest Biol. Technol. 2010, 56, 1–11. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Schena, L.; Nigro, F.; de Girolamo, A.; Ippolito, A. Effect of quercetin and umbelliferone on the transcript level of Penicillium expansum genes involved in patulin biosynthesis. Eur. J. Plant Pathol. 2009, 125, 223–233. [Google Scholar]
- Sanzani, S.M.; Reverberi, M.; Punelli, M.; Ippolito, A.; Fanelli, C. Study on the role of patulin on pathogenicity and virulence of Penicillium expansum. Int. J. Food Microbiol. 2012, 153, 323–331. [Google Scholar] [CrossRef]
- Fallanaj, F.; Sanzani, S.M.; Zavanella, C.; Ippolito, A. Salt addition improves the control of citrus postharvest diseases using electrolysis with conductive diamond electrodes. J. Plant Pathol. 2013, 95, 373–383. [Google Scholar]
- Mari, M.; Leoni, O.; Iori, R.; Cembali, T. Antifungal vapour-phase activity of ally-isothiocyanate against Penicillium expansum on pears. Plant Pathol. 2002, 51, 231–236. [Google Scholar] [CrossRef]
- Feliziani, E.; Santini, M.; Landi, L.; Romanazzi, G. Pre-and postharvest treatment with alternatives to synthetic fungicides to control postharvest decay of sweet cherry. Postharvest Biol. Technol. 2013, 78, 133–138. [Google Scholar] [CrossRef]
- Gatto, M.A.; Sanzani, S.M.; Tardia, P.; Linsalata, V.; Pieralice, M.; Sergio, L.; di Venere, D. Antifungal activity of total and fractionated phenolic extracts from two wild edible herbs. Nat. Sci. 2013, 5, 895–902. [Google Scholar]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sanzani, S.M.; Schena, L.; Ippolito, A. Effectiveness of Phenolic Compounds against Citrus Green Mould. Molecules 2014, 19, 12500-12508. https://doi.org/10.3390/molecules190812500
Sanzani SM, Schena L, Ippolito A. Effectiveness of Phenolic Compounds against Citrus Green Mould. Molecules. 2014; 19(8):12500-12508. https://doi.org/10.3390/molecules190812500
Chicago/Turabian StyleSanzani, Simona M., Leonardo Schena, and Antonio Ippolito. 2014. "Effectiveness of Phenolic Compounds against Citrus Green Mould" Molecules 19, no. 8: 12500-12508. https://doi.org/10.3390/molecules190812500
APA StyleSanzani, S. M., Schena, L., & Ippolito, A. (2014). Effectiveness of Phenolic Compounds against Citrus Green Mould. Molecules, 19(8), 12500-12508. https://doi.org/10.3390/molecules190812500