Sesamin Enhances Cholesterol Efflux in RAW264.7 Macrophages

Abstract
:1. Introduction
2. Results and Discussion
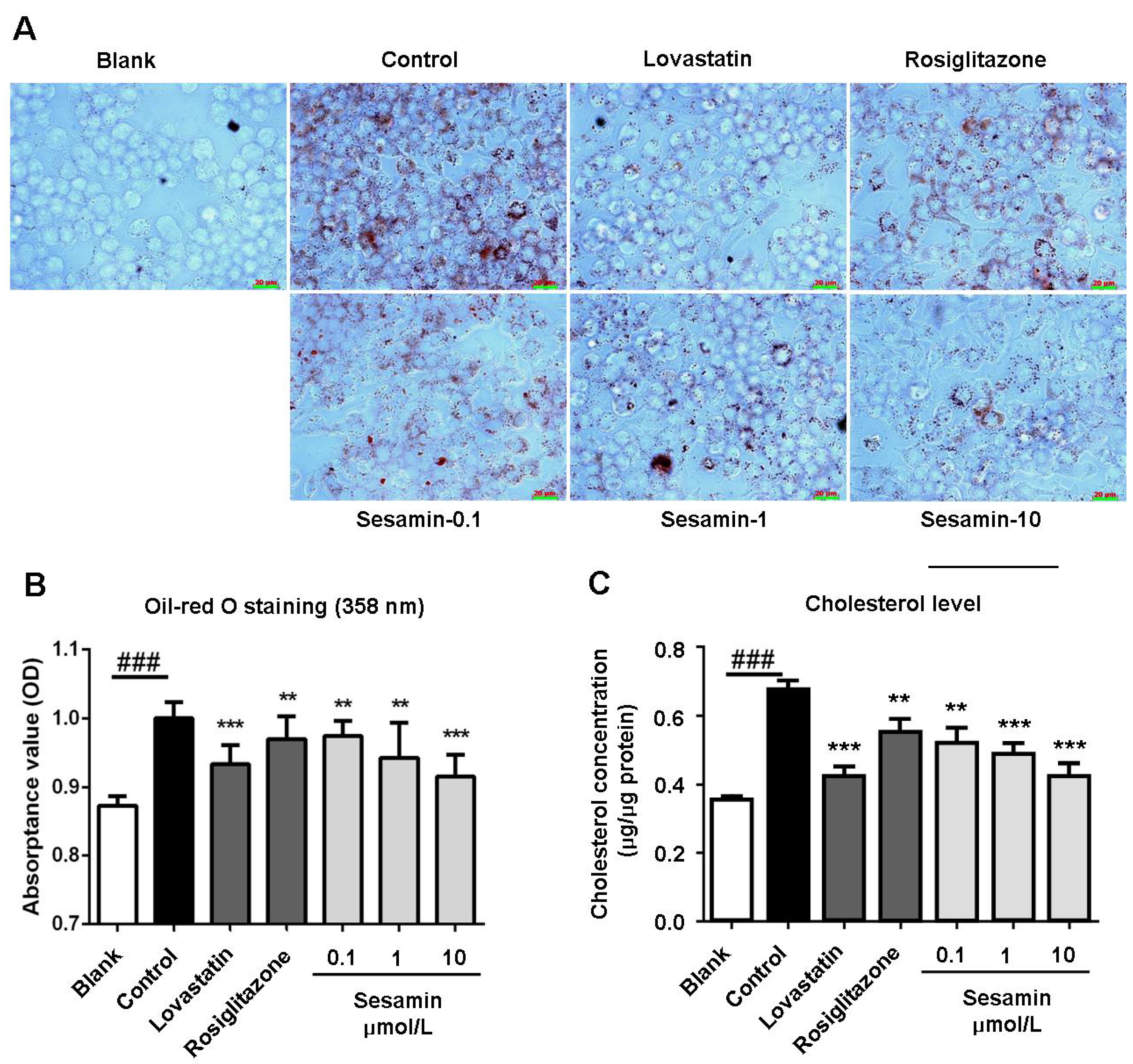
2.1. Sesamin Inhibits oxLDL-Induced Cholesterol Accumulation in RAW264.7 Cells
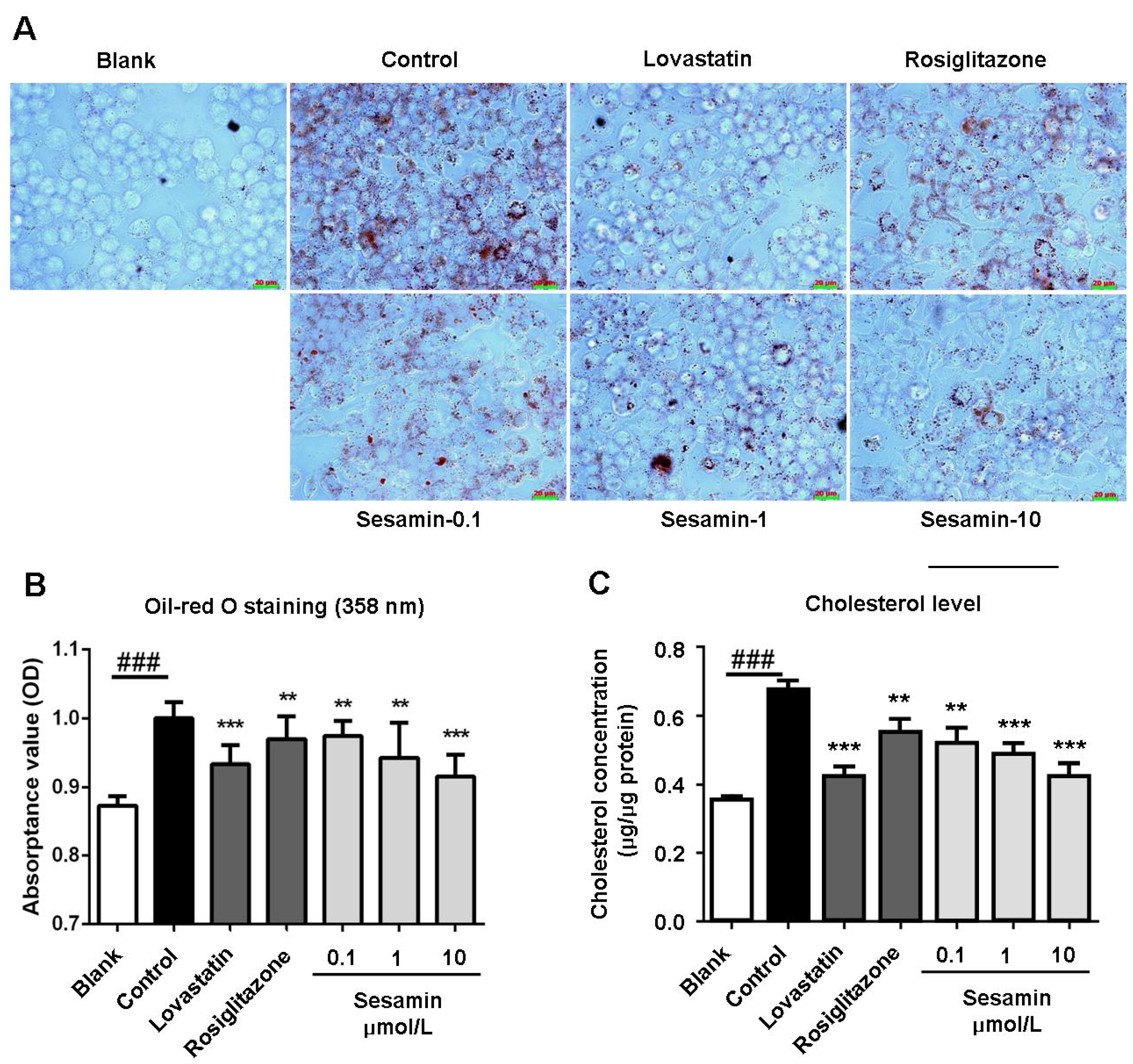
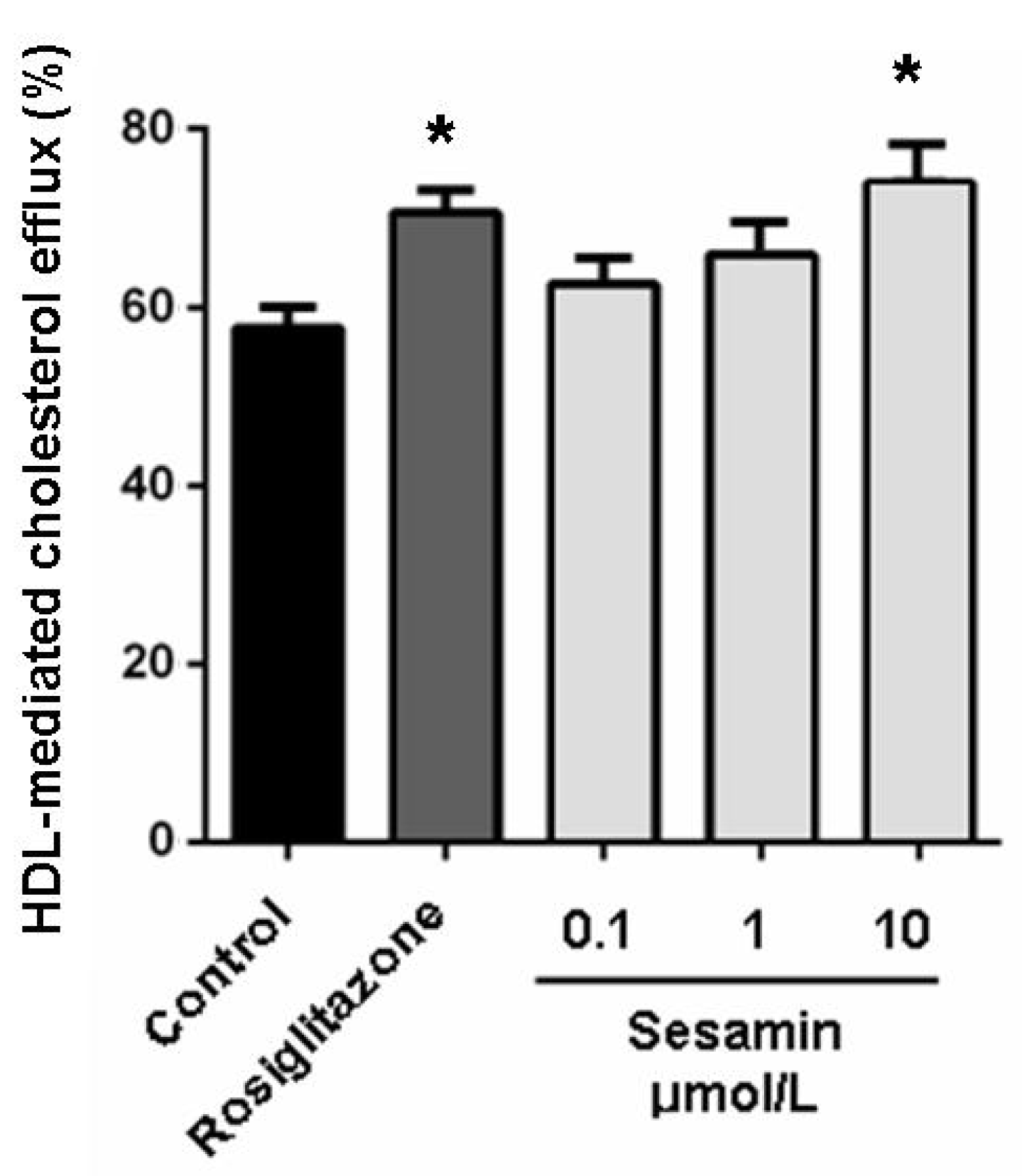
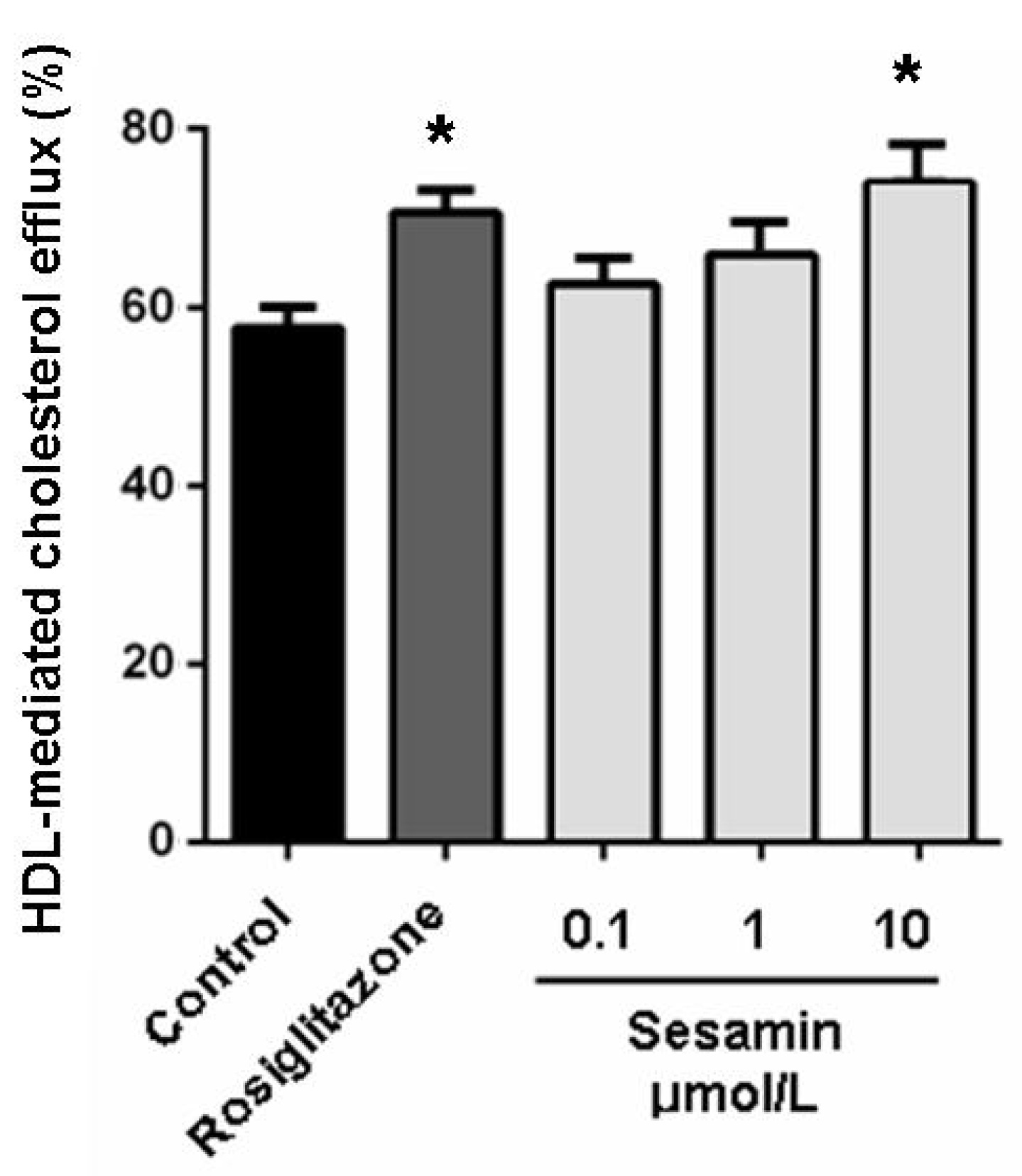
2.2. Sesamin Enhances Cholesterol Efflux from RAW264.7 Macrophages
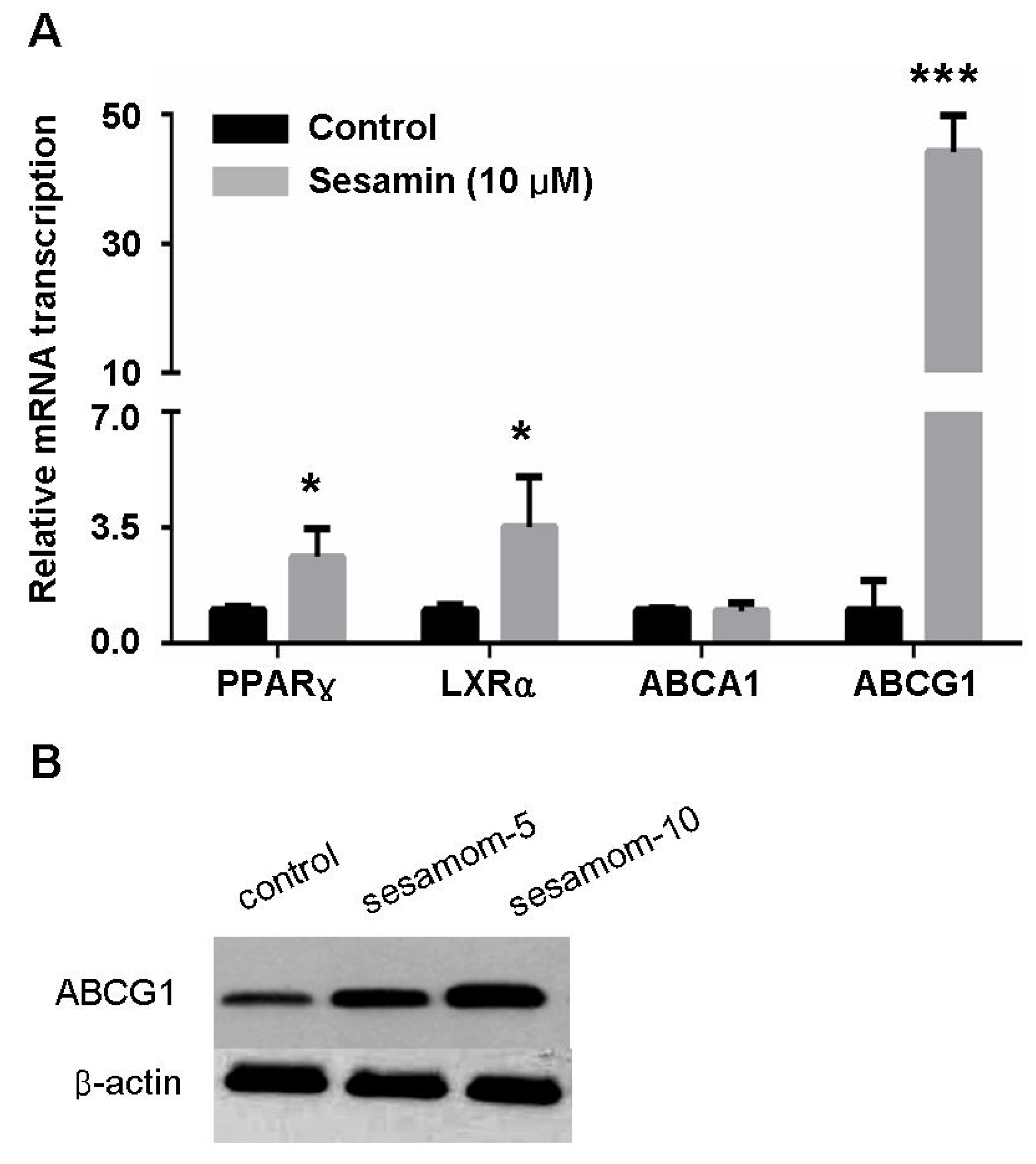
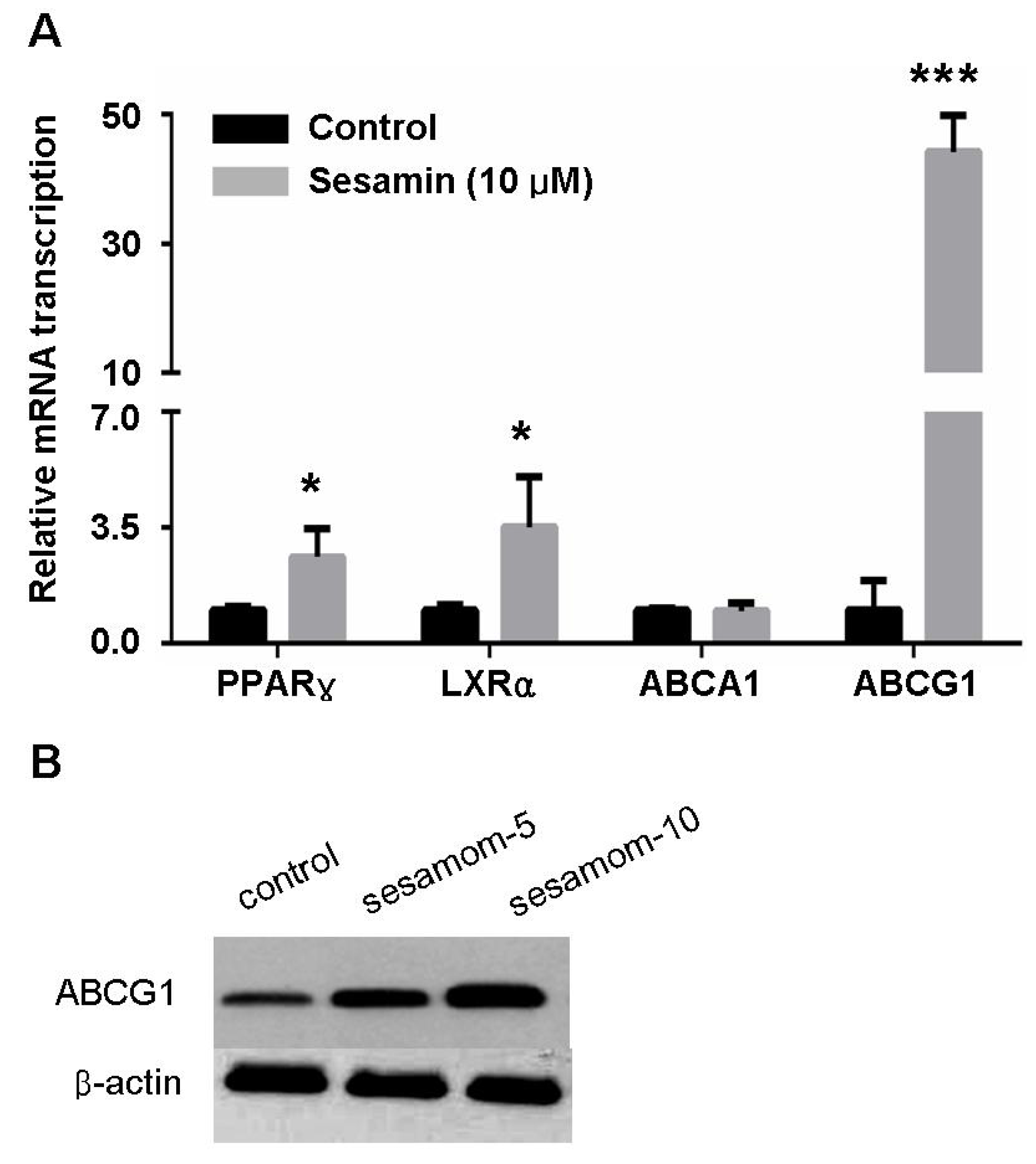
2.3. Sesamin Increases Transcription Of Cholesterol Efflux-Associated Genes

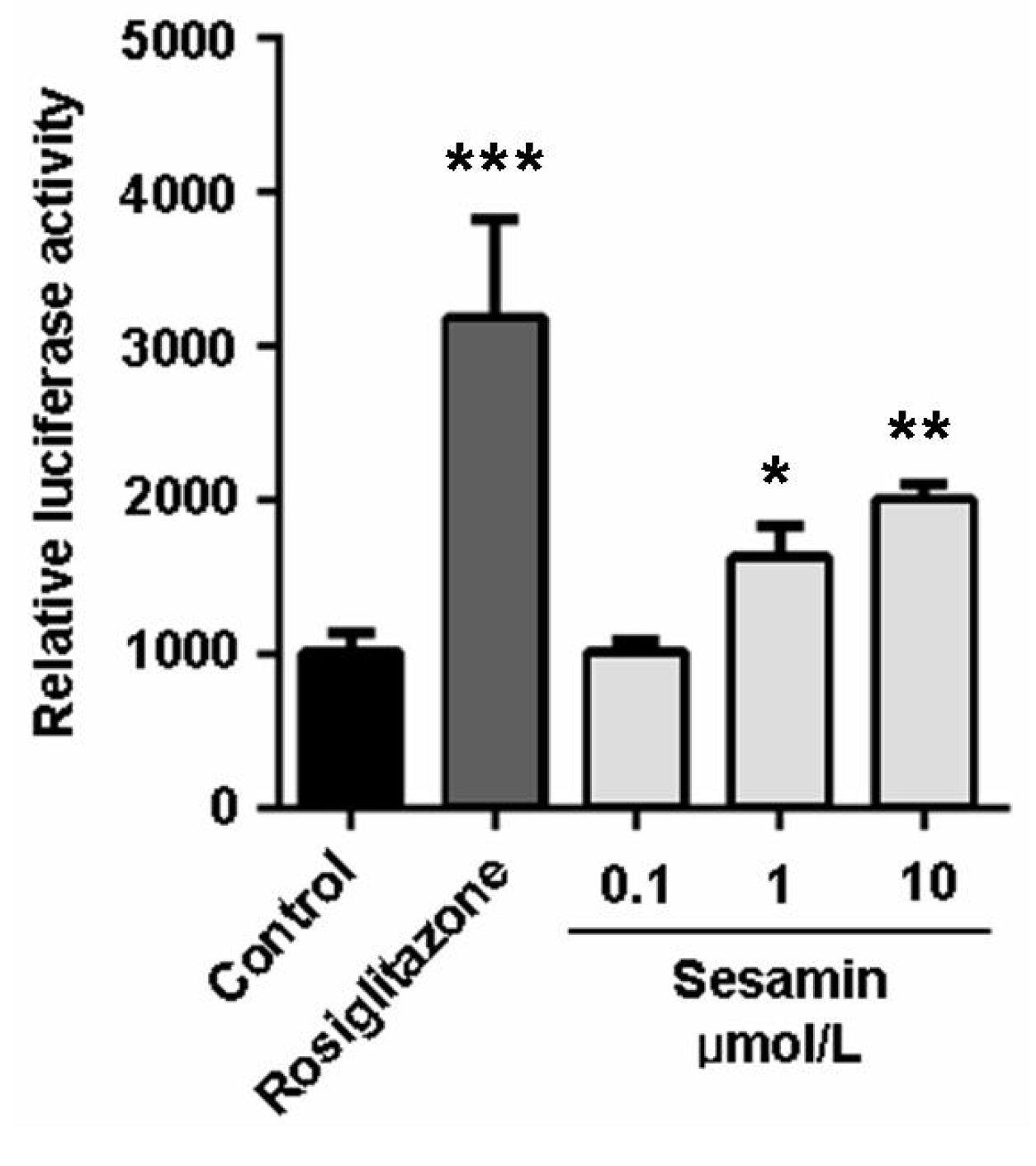
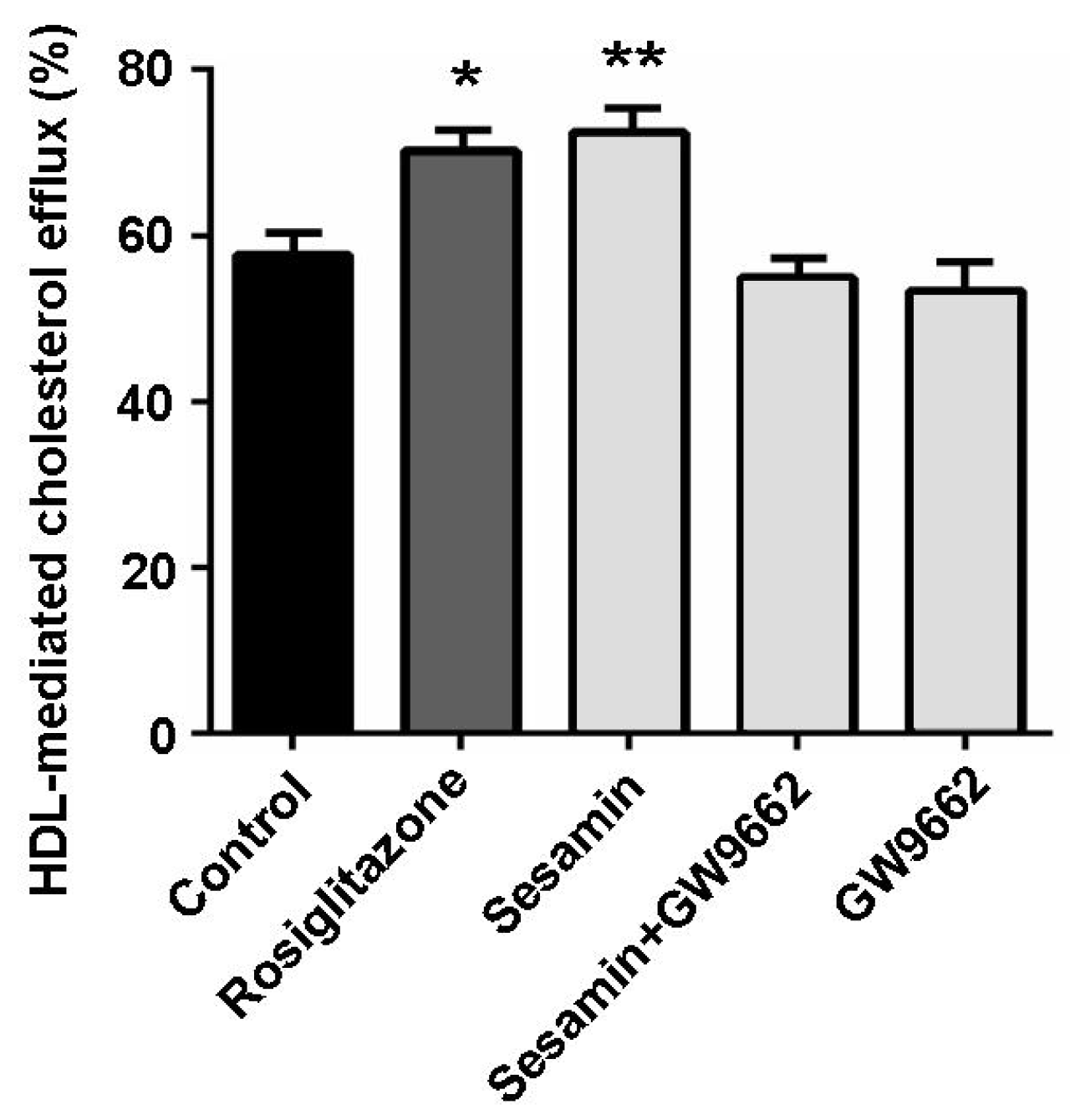
2.4. The Cholesterol Efflux-Promoting Effect of Sesamin is Abolished by Pparγ Antagonist Gw9662

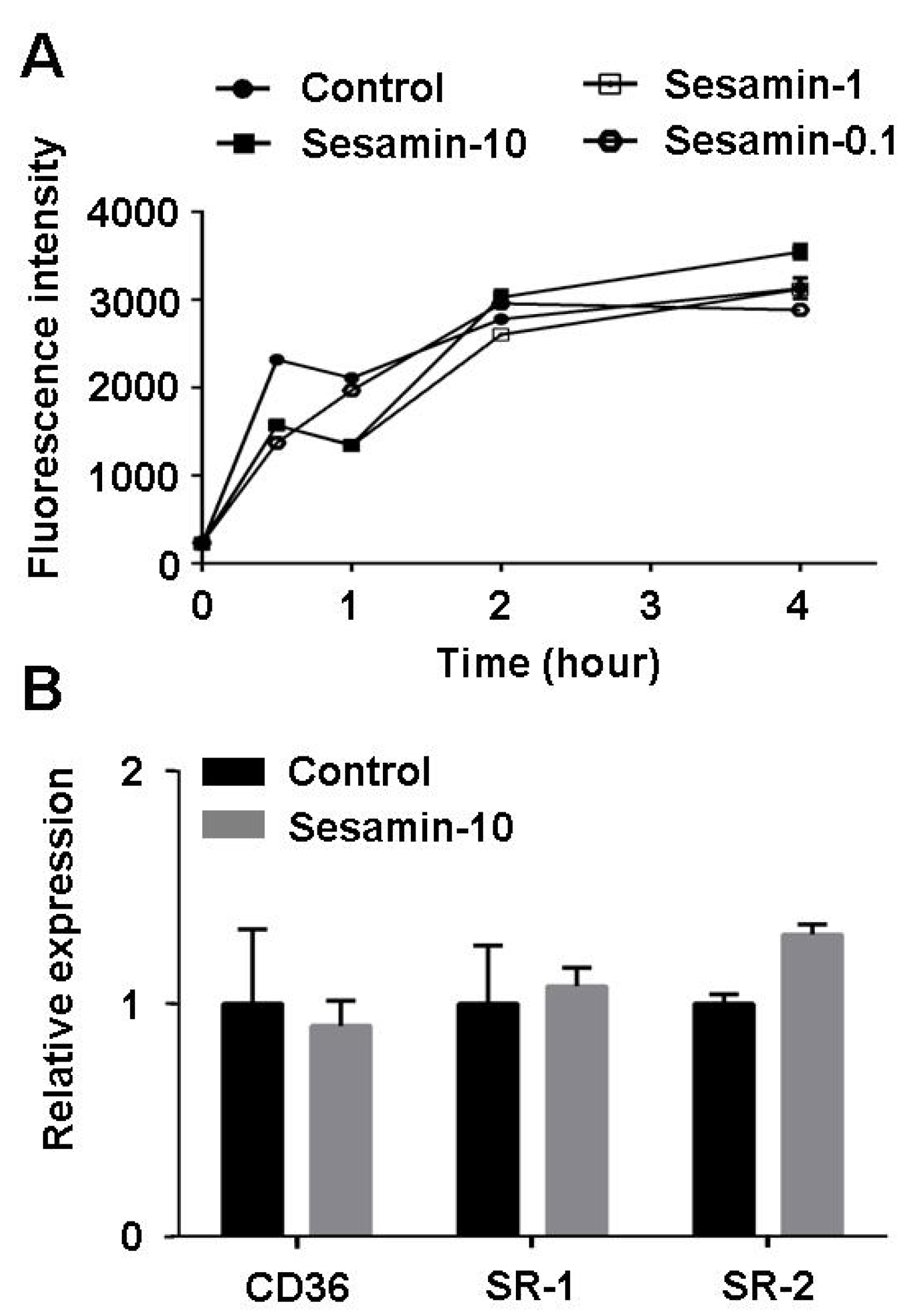
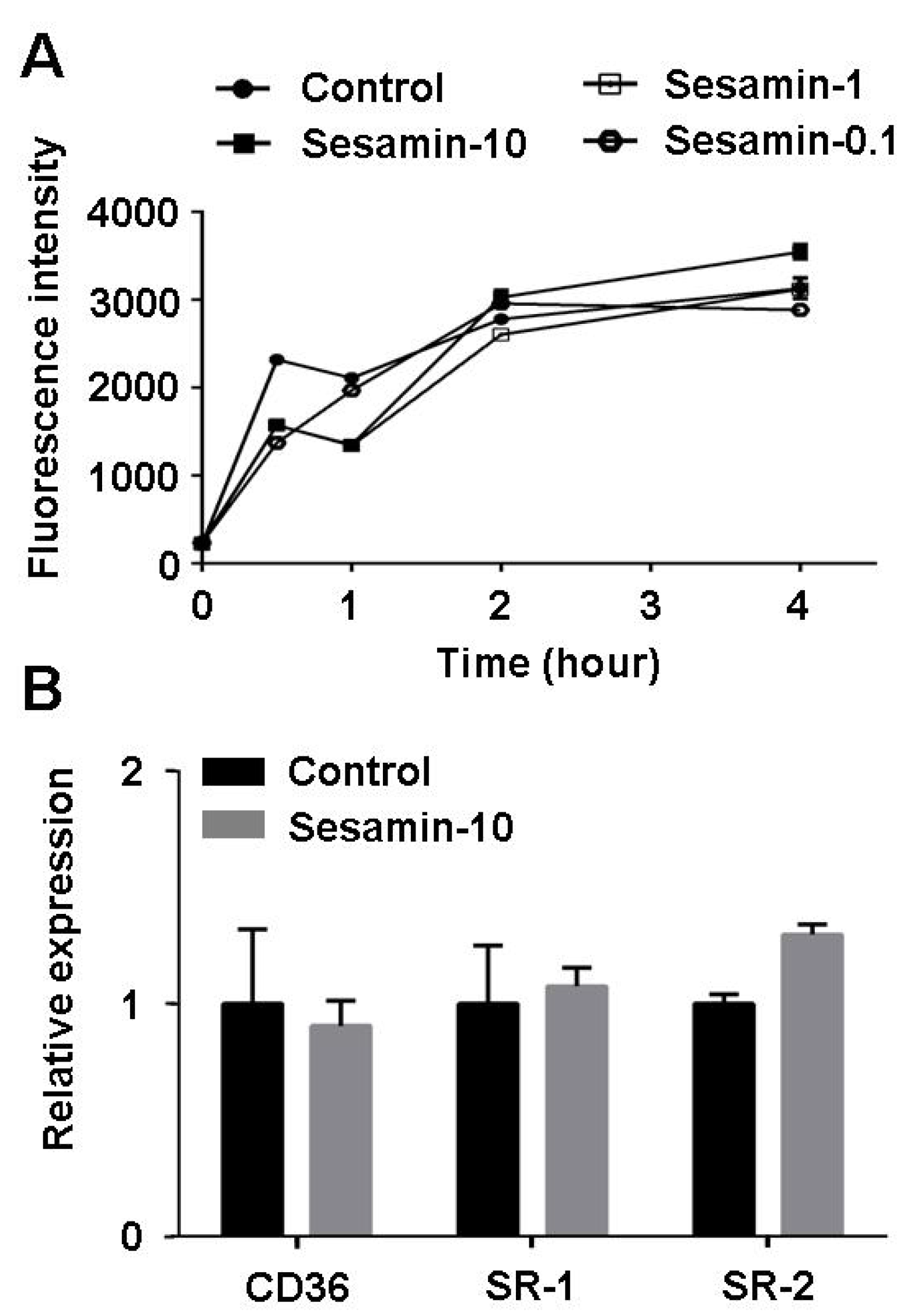
2.5. Sesamin Shows no Effect on Oxldl Uptake in Raw264.7 Macrophages
3. Experimental
3.1. Materials
3.2. Cell Culture and Cholesterol Accumulation Assay
3.3. Cellular Cholesterol Efflux Experiments
3.4. Quantitative Real-Time PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward (5'–3') | Reverse (5'–3') |
|---|---|---|
| PPARγ | GCAGCTACTGCATGTGATCAAGA | GTCAGCGGGTGGGACTTTC |
| LXRα | AGGAGTGTCGACTTCGCAAA | CTCTTCTTGCCGCTTCAGTTT |
| ABCA1 | CCCAGAGCAAAAAGGGACTC | GGTCATCATCACTTTGGTCCTTG |
| ABCG1 | CAAGACCCTTTTGAAAGGGATCTC | GCCAGAATATTCATGAGTGTGGAC |
| CD36 | CAAGCTCCTTGGCATGGTAGA | TGGATTTGCAAGCACAATATGAA |
| SR-1 | TTAAAGGTGATCGGGGACAAA | CAACCAGTCGAACTGTCTTAAG |
| SR-2 | TTAAAGGTGATCGGGGACAAA | AGCTGATCTTAAAAGGGTCTTG |
| β-actin | CCTGGCACCCAGCACAAT | GCCGATCCACACACGGAGTACT |
3.5. 25-NBD Cholesterol Uptake Assay
3.6. Luciferase Assay
3.7. Statistics Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yu, X.H.; Fu, Y.C.; Zhang, D.W.; Yin, K.; Tang, C.K. Foam cells in atherosclerosis. Clin. Chim. Acta 2013, 424, 245–252. [Google Scholar]
- Reiss, A.B.; Cronstein, B.N. Regulation of foam cells by adenosine. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 879–886. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.F.; Chen, W.J.; Tang, S.L.; Mo, Z.C.; Tang, Y.Y.; Li, Y.; Wang, J.L.; Liu, X.Y.; Peng, J.; et al. MicroRNA-27a/b regulates cellular cholesterol efflux, influx and esterification/hydrolysis in THP-1 macrophages. Atherosclerosis 2014, 234, 54–64. [Google Scholar] [CrossRef]
- Yvan-Charvet, L.; Wang, N.; Tall, A.R. Role of HDL, ABCA1, and ABCG1 transporters in cholesterol efflux and immune responses. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 139–143. [Google Scholar] [CrossRef]
- Tall, A.R.; Yvan-Charvet, L.; Terasaka, N.; Pagler, T.; Wang, N. HDL, ABC transporters, and cholesterol efflux: Implications for the treatment of atherosclerosis. Cell. Metab. 2008, 7, 365–375. [Google Scholar] [CrossRef]
- Meurs, I.; van Eck, M.; van Berkel, T.J. High-density lipoprotein: Key molecule in cholesterol efflux and the prevention of atherosclerosis. Curr. Pharm. Des. 2010, 16, 1445–1467. [Google Scholar] [CrossRef]
- Rohrer, L.; Ohnsorg, P.M.; Lehner, M.; Landolt, F.; Rinninger, F.; von Eckardstein, A. High-density lipoprotein transport through aortic endothelial cells involves scavenger receptor BI and ATP-binding cassette transporter G1. Circ. Res. 2009, 104, 1142–1150. [Google Scholar] [CrossRef]
- Chinetti, G.; Lestavel, S.; Bocher, V.; Remaley, A.T.; Neve, B.; Torra, I.P.; Teissier, E.; Minnich, A.; Jaye, M.; Duverger, N.; et al. PPAR-alpha and PPAR-gamma activators induce cholesterol removal from human macrophage foam cells through stimulation of the ABCA1 pathway. Nat. Med. 2001, 7, 53–58. [Google Scholar]
- Kiso, Y. Antioxidative roles of sesamin, a functional lignan in sesame seed, and it’s effect on lipid- and alcohol-metabolism in the liver: A DNA microarray study. Biofactors 2004, 21, 191–196. [Google Scholar] [CrossRef]
- Lee, W.J.; Ou, H.C.; Wu, C.M.; Lee, I.T.; Lin, S.Y.; Lin, L.Y.; Tsai, K.L.; Lee, S.D.; Sheu, W.H. Sesamin mitigates inflammation and oxidative stress in endothelial cells exposed to oxidized low-density lipoprotein. J. Agric. Food Chem. 2009, 57, 11406–11417. [Google Scholar] [CrossRef]
- Miyawaki, T.; Aono, H.; Toyoda-Ono, Y.; Maeda, H.; Kiso, Y.; Moriyama, K. Antihypertensive effects of sesamin in humans. J. Nutr. Sci. Vitaminol. 2009, 55, 87–91. [Google Scholar] [CrossRef]
- Saeed, M.; Khalid, H.; Sugimoto, Y.; Efferth, T. The lignan, (−)-sesamin reveals cytotoxicity toward cancer cells: Pharmacogenomic determination of genes associated with sensitivity or resistance. Phytomedicine 2014, 21, 689–696. [Google Scholar] [CrossRef]
- Rogi, T.; Tomimori, N.; Ono, Y.; Kiso, Y. The mechanism underlying the synergetic hypocholesterolemic effect of sesamin and alpha-tocopherol in rats fed a high-cholesterol diet. J. Pharmacol. Sci. 2011, 115, 408–416. [Google Scholar] [CrossRef]
- Li, W.X.; Kong, X.; Zhang, J.X.; Yang, J.R. Long-term intake of sesamin improves left ventricular remodelling in spontaneously hypertensive rats. Food Funct. 2013, 4, 453–460. [Google Scholar] [CrossRef]
- Wu, X.Q.; Kong, X.; Zhou, Y.; Huang, K.; Yang, J.R.; Li, X.L. Sesamin exerts renoprotective effects by enhancing NO bioactivity in renovascular hypertensive rats fed with high-fat-sucrose diet. Eur. J. Pharmacol. 2012, 683, 231–237. [Google Scholar] [CrossRef]
- Hong, L.; Yi, W.; Liangliang, C.; Juncheng, H.; Qin, W.; Xiaoxiang, Z. Hypoglycaemic and hypolipidaemic activities of sesamin from sesame meal and its ability to ameliorate insulin resistance in KK-Ay mice. J. Sci. Food Agric. 2013, 93, 1833–1838. [Google Scholar] [CrossRef]
- Ma, J.Q.; Ding, J.; Zhang, L.; Liu, C.M. Hepatoprotective properties of sesamin against CCl4 induced oxidative stress-mediated apoptosis in mice via JNK pathway. Food Chem. Toxicol. 2014, 64, 41–48. [Google Scholar] [CrossRef]
- Ide, T.; Azechi, A.; Kitade, S.; Kunimatsu, Y.; Suzuki, N.; Nakajima, C. Combined effect of sesamin and alpha-lipoic acid on hepatic fatty acid metabolism in rats. Eur. J. Nutr. 2013, 52, 1015–1027. [Google Scholar] [CrossRef]
- Ashakumary, L.; Rouyer, I.; Takahashi, Y.; Ide, T.; Fukuda, N.; Aoyama, T.; Hashimoto, T.; Mizugaki, M.; Sugano, M. Sesamin, a sesame lignan, is a potent inducer of hepatic fatty acid oxidation in the rat. Metabolism 1999, 48, 1303–1313. [Google Scholar] [CrossRef]
- Yaguchi, Y.; Komura, T.; Kashima, N.; Tamura, M.; Kage-Nakadai, E.; Saeki, S.; Terao, K.; Nishikawa, Y. Influence of oral supplementation with sesamin on longevity of Caenorhabditis elegans and the host defense. Eur. J. Nutr. 2014. [Google Scholar] [CrossRef]
- Deng, P.; Wang, C.; Chen, L.; Du, Y.; Yan, X.; Chen, M.; Yang, G.; He, G. Sesamin Induces cell cycle arrest and apoptosis through the inhibition of signal transducer and activator of transcription 3 signalling in human hepatocellular carcinoma cell line HepG2. Biol. Pharm. Bull. 2013, 36, 1540–1548. [Google Scholar] [CrossRef]
- Periasamy, S.; Hsu, D.Z.; Chang, P.C.; Liu, M.Y. Sesame oil attenuates nutritional fibrosing steatohepatitis by modulating matrix metalloproteinases-2, 9 and PPAR-gamma. J. Nutr. Biochem. 2014, 25, 337–344. [Google Scholar] [CrossRef]
- Dandona, P.; Chaudhuri, A.; Ghanim, H. Rosiglitazone, thiazolidinediones and atherosclerosis. Atherosclerosis 2010, 211, 384–385. [Google Scholar] [CrossRef]
- Loke, W.M.; Proudfoot, J.M.; Hodgson, J.M.; McKinley, A.J.; Hime, N.; Magat, M.; Stocker, R.; Croft, K.D. Specific dietary polyphenols attenuate atherosclerosis in apolipoprotein E-knockout mice by alleviating inflammation and endothelial dysfunction. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 749–757. [Google Scholar] [CrossRef]
- Ozasa, H.; Ayaori, M.; Iizuka, M.; Terao, Y.; Uto-Kondo, H.; Yakushiji, E.; Takiguchi, S.; Nakaya, K.; Hisada, T.; Uehara, Y.; et al. Pioglitazone enhances cholesterol efflux from macrophages by increasing ABCA1/ABCG1 expressions via PPARgamma/LXRalpha pathway: Findings from in vitro and ex vivo studies. Atherosclerosis 2011, 219, 141–150. [Google Scholar] [CrossRef]
- Wojtowicz, A.K.; Szychowski, K.A.; Kajta, M. PPAR-gamma agonist GW1929 but not antagonist GW9662 reduces TBBPA-induced neurotoxicity in primary neocortical cells. Neurotox. Res. 2014, 25, 311–322. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, X.; Luan, H.; Sun, G.; Sun, X.; Wang, X.; Guo, P.; Xu, X. The caffeoylquinic acid-rich Pandanus tectorius fruit extract increases insulin sensitivity and regulates hepatic glucose and lipid metabolism in diabetic db/db mice. J. Nutr. Biochem. 2014, 25, 412–419. [Google Scholar] [CrossRef]
- Wu, C.; Guo, Y.; Su, Y.; Zhang, X.; Luan, H.; Zhu, H.; He, H.; Wang, X.; Sun, G.; Sun, X.; et al. Cordycepin activates AMP-activated protein kinase (AMPK) via interaction with the gamma1 subunit. J. Cell. Mol. Med. 2014, 18, 293–304. [Google Scholar] [CrossRef]
- Wu, C.; Luan, H.; Wang, S.; Zhang, X.; Wang, R.; Jin, L.; Guo, P.; Chen, X. Modulation of lipogenesis and glucose consumption in HepG2 cells and C2C12 myotubes by sophoricoside. Molecules 2013, 18, 15624–15635. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, N.; Wu, C.; Sun, L.; Zheng, J.; Guo, P. Sesamin Enhances Cholesterol Efflux in RAW264.7 Macrophages. Molecules 2014, 19, 7516-7527. https://doi.org/10.3390/molecules19067516
Liu N, Wu C, Sun L, Zheng J, Guo P. Sesamin Enhances Cholesterol Efflux in RAW264.7 Macrophages. Molecules. 2014; 19(6):7516-7527. https://doi.org/10.3390/molecules19067516
Chicago/Turabian StyleLiu, Nan, Chongming Wu, Lizhong Sun, Jun Zheng, and Peng Guo. 2014. "Sesamin Enhances Cholesterol Efflux in RAW264.7 Macrophages" Molecules 19, no. 6: 7516-7527. https://doi.org/10.3390/molecules19067516
APA StyleLiu, N., Wu, C., Sun, L., Zheng, J., & Guo, P. (2014). Sesamin Enhances Cholesterol Efflux in RAW264.7 Macrophages. Molecules, 19(6), 7516-7527. https://doi.org/10.3390/molecules19067516