Entrapment of Probiotics in Water Extractable Arabinoxylan Gels: Rheological and Microstructural Characterization


 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
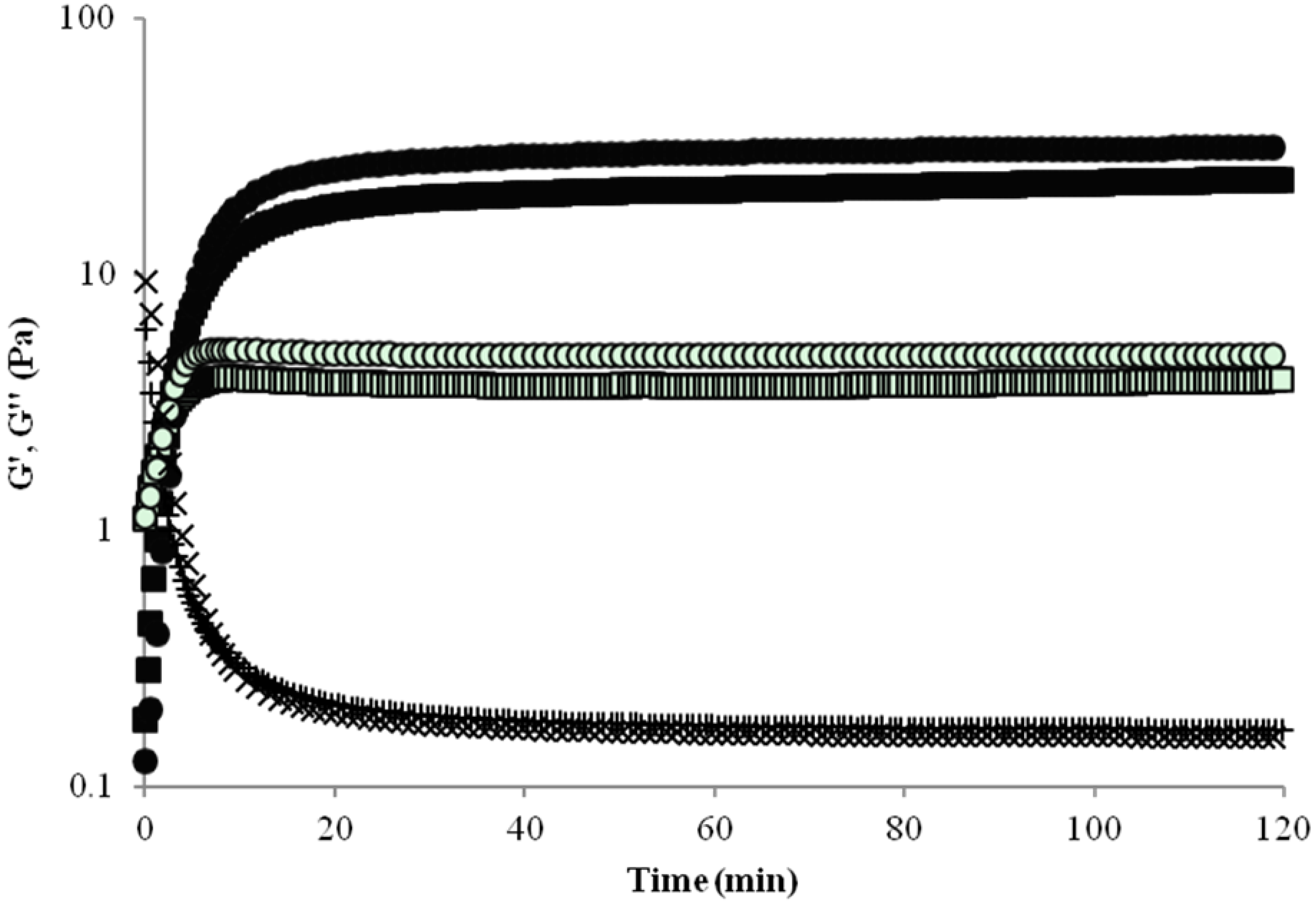
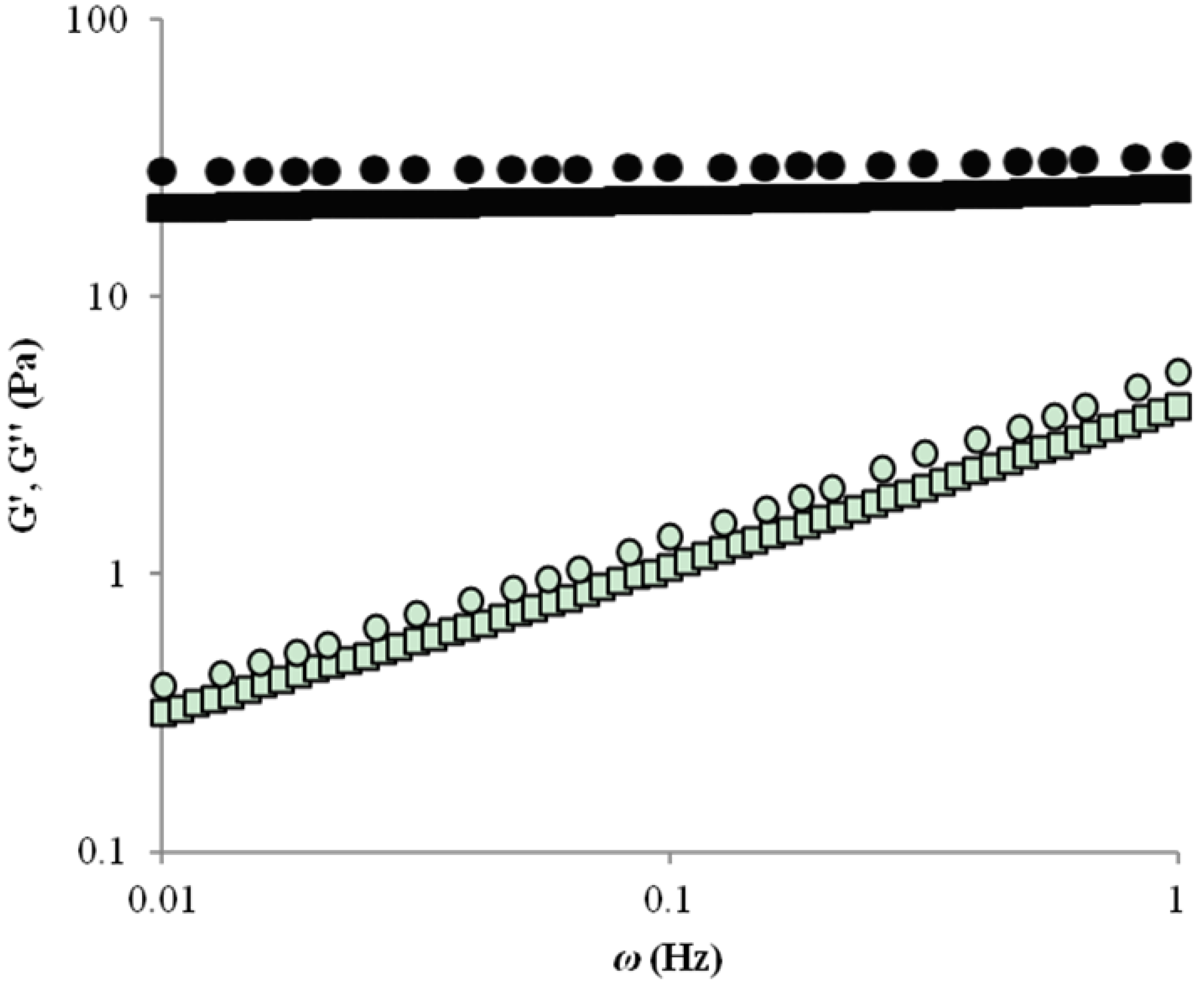
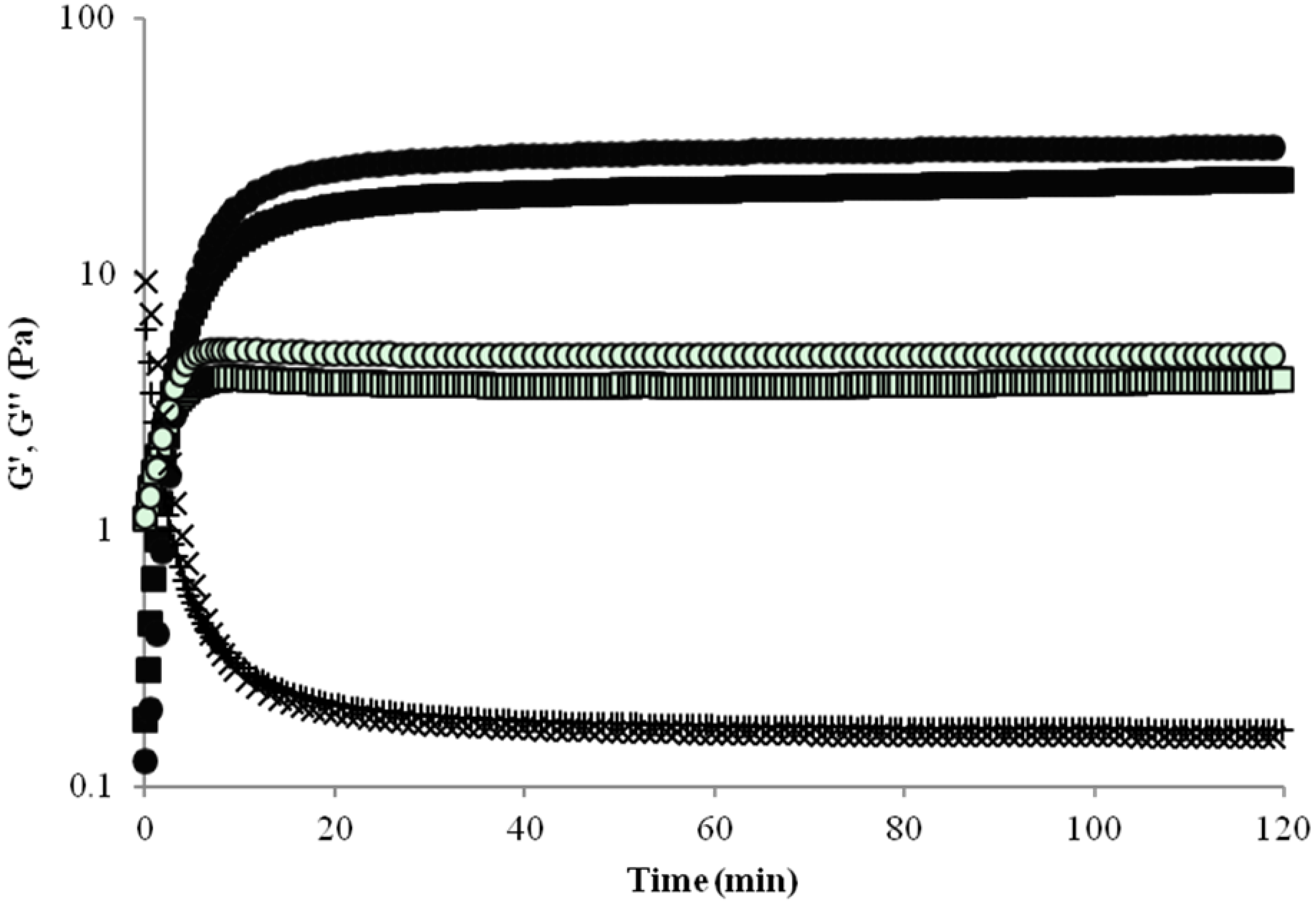
2.1. Rheology
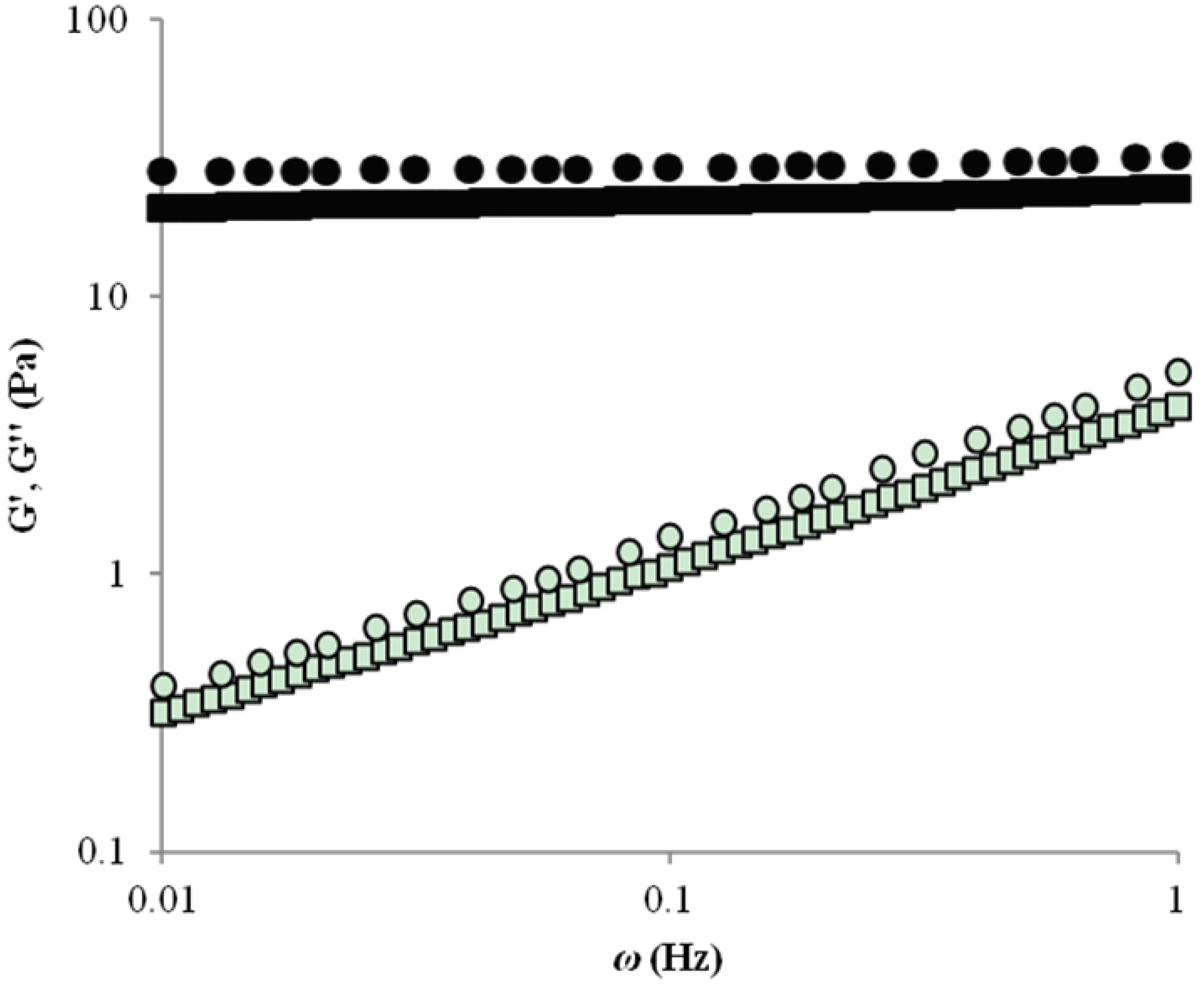
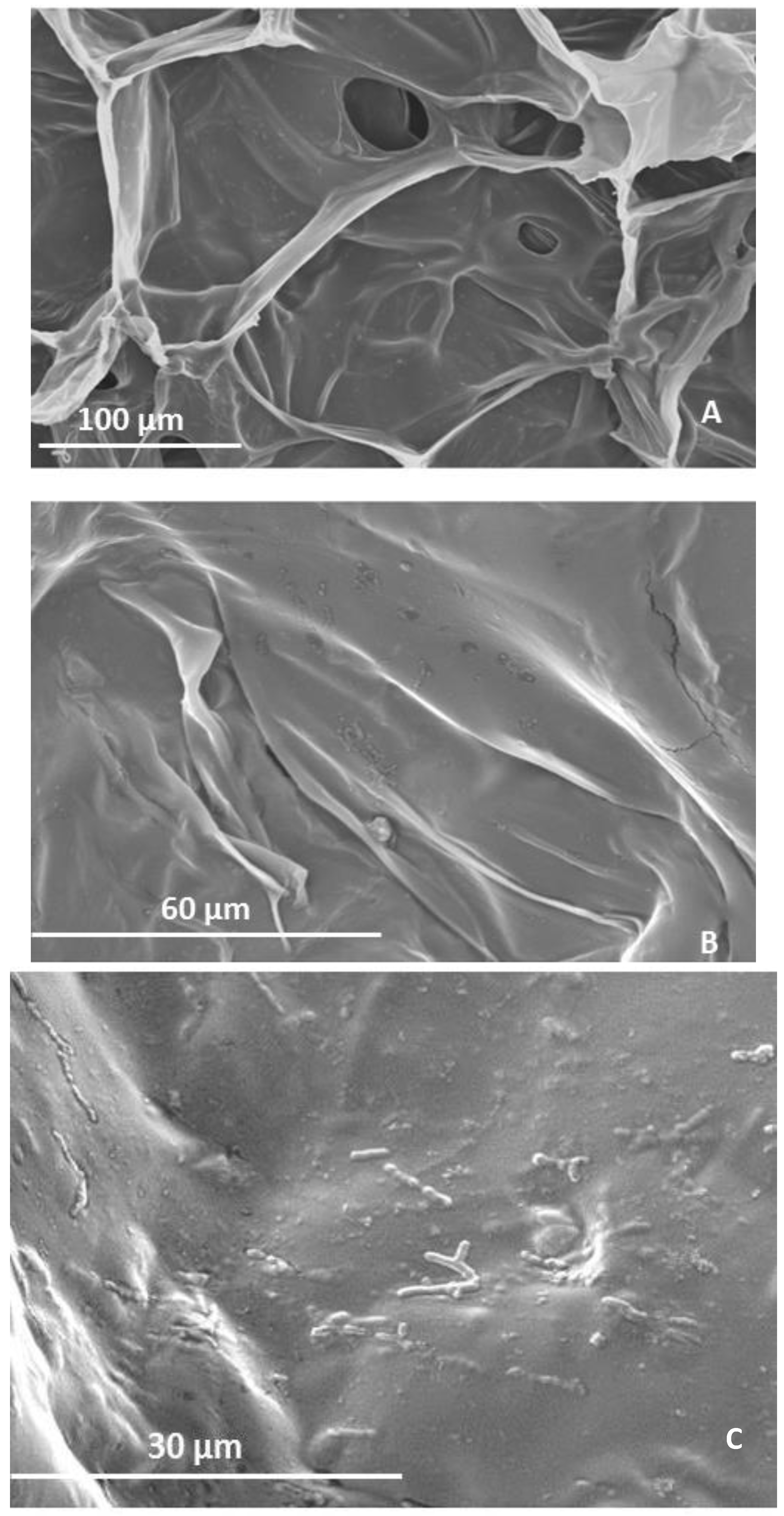
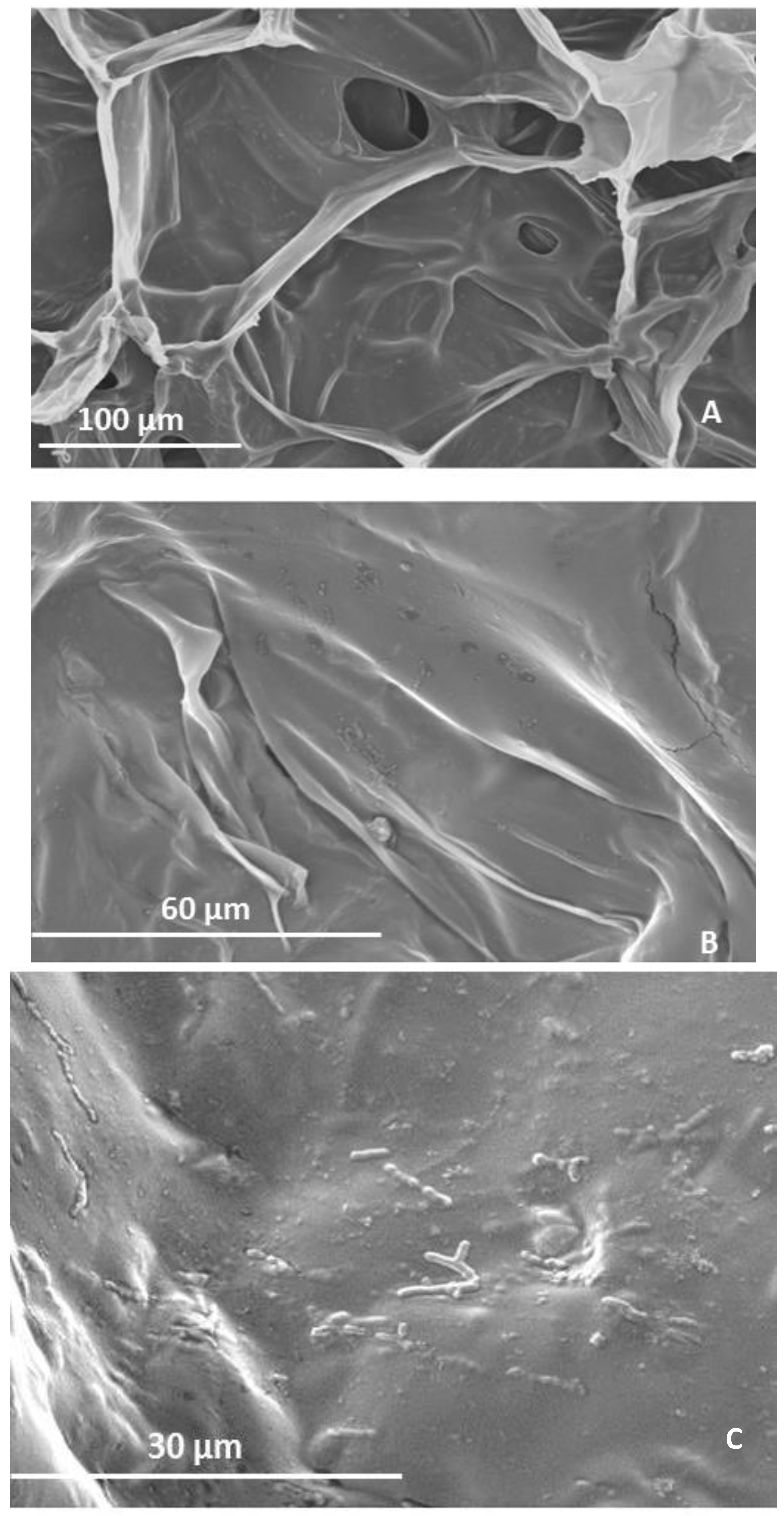
2.2. Microstructure

3. Experimental
3.1. Materials
3.2. WEAX and WEAX/B. Longum Gelation
3.3. Rheology
3.4. Microstructure
4. Conclusions
Acknowledgments
Author Contributions
Conflictts of Interest
References
- Gentile, F.T.; Doherty, E.J.; Rein, D.H.; Shoichet, M.S.; Winn, S.R. Polymer science for microencapsulation of cells for central nervous system transplantation. React. Polym. 1995, 25, 207–227. [Google Scholar] [CrossRef]
- Gbassi, G.K.; Vandamme, T. Probiotic Encapsulation Technology: From Microencapsulation to Release into the Gut. Pharmaceutics 2012, 4, 149–163. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Biliaderis, C.G. Cereal arabinoxylans: Advances in structure and physicochemical properties. Carbohydr. Polym. 1995, 28, 33–48. [Google Scholar] [CrossRef]
- Carvajal-Millan, E.; Landillon, V.; Morel, M.H.; Rouau, X.; Doublier, J.L.; Micard, V. Arabinoxylan gels: Impact of the feruloylation degree on their structure and properties. Biomacromolecules 2005, 6, 309–317. [Google Scholar] [CrossRef]
- Hopkins, M.J.; Englyst, H.N.; Macfarlane, S.; Furrie, E.; Macfarlane, G.T.; McBain, A.J. Degradation of cross-linked and non-cross-linked arabinoxylans by the intestinal microbiota in children. Appl. Environ. Microbiol. 2003, 69, 6354–6360. [Google Scholar] [CrossRef]
- Rascón-Chu, A.; Martínez-López, A.L.; Berlanga-Reyes, C.; Carvajal-Millan, E.; Campa-Mada, A.C.; Gardea, A.A.; Orozco-Avitia, A. Arabinoxylans Gels as Lycopene Carriers: In vitro Degradation by Colonic Bacteria. MRS Online Proc. Libr. 2012, 1487. imrc12-s4b-p035. [Google Scholar] [CrossRef]
- Van Laere, K.M.J.; Hartemink, R.; Bosveld, M.; Shols, H.A.; Voragen, A.G.J. Fermentation of plant cell wall derived polysaccharides by intestinal bacteria. J. Agric. Food Chem. 2000, 48, 1644–1652. [Google Scholar] [CrossRef]
- Broekaert, W.; Courtin, C.; Verbeke, K.; van de Wiele, T.; Delcour, J. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef]
- Silva1, A.M.; Barbosa, F.H.F.; Duarte, R.; Vieira, L.Q.; Arantes, R.M.E.; Nicoli, J.R. Effect of Bifidobacterium longum ingestion on experimental salmonellosis in mice. J. Appl. Microbiol. 2004, 97, 29–37. [Google Scholar] [CrossRef]
- Doublier, J.L.; Cuvelier, G. Gums and Hydrocolloids: Functional Aspects. In Carbohydrates in Food; Eliasson, A.C., Ed.; Marcel Dekker: New York, NY, USA, 1996; pp. 283–318. [Google Scholar]
- Ross-Murphy, S.B. Rheological Methods. In Biophysical Methods in Food Research; Chan, H.W.S., Ed.; Blackwell Scientific Publications: Oxford, UK, 1984; pp. 138–199. [Google Scholar]
- Paes, G.; Chabbert, B. Characterization of Arabinoxylan/cellulose nanocrystals gels to investigate fluorescent probes mobility in bioinspired models of plant secondary cell wall. Biomacromolecules 2012, 13, 206–214. [Google Scholar] [CrossRef]
- Iravani, S.; Fitchett, C.S.; Georget, D.M.R. Physical characterization of arabinoxylan powder and its hydrogel containing a methyl xanthine. Carbohydr. Polym. 2011, 85, 201–207. [Google Scholar] [CrossRef]
- Martínez-López, A.L.; Carvajal-Millan, E.; Miki-Yoshida, M.; Alvarez-Contreras, L.; Rascón-Chu, A.; Lizardi-Mendoza, J.; López-Franco, Y. Arabinoxylan microspheres: Structural and textural characteristics. Molecules 2013, 18, 4640–4650. [Google Scholar] [CrossRef]
- Martínez-López, A.L.; Carvajal-Millan, E.; Rascón-Chu, A.; Márquez-Escalante, J.; Martínez-Robinson, K. Gels of ferulated arabinoxylans extracted from nixtamalizedand non-nixtamalized maize bran: Rheological and structural characteristics. CyTA-J. Food 2013, 11, 22–28. [Google Scholar] [CrossRef]
- Zhu, S.; Ramaswamy, H.S.; le Bail, A. Ice-crystal formation in gelatine gel during pressure shift versus conventional freezing. J. Food Eng. 2005, 66, 69–76. [Google Scholar] [CrossRef]
- Wu, Z. Bacterial Low Temperature Survival, Ice Nucleation Protein and Ice Associated Polymers. Ph.D. Thesis, Queen’s University, Kingston, ON, Canada, 2010. [Google Scholar]
- Sousa, S.; Gomes, A.M.; Pintado, M.M.; Silva, J.P.; Costa, P.; Amaral, M.H.; Duarte, A.C.; Rodrigues, D.; Rocha-Santos, T.A.P.; Freitas, A.C. Characterization of freezing effect upon stability of probiotic loaded, calcium-alginate microparticles. Food Bioprod. Process. 2013, in press. [Google Scholar]
- Sathyabama, S.; Ranjith kumar, M.; Bruntha devi, P.; Vijayabharathi, R.; Brindha priyadharisini, V. Co-encapsulation of probiotics with prebiotics on alginate matrix and its effect on viability in simulated gastric environment. LWT- Food Sci. Technol. 2014, 57, 419–425. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, L.; Lobato-Calleros, C.; Pimentel-González, D.J.; Vernon-Carter, E.J. Lactobacillus plantarum protection by entrapment in whey protein isolate: κ-carrageenan complex coacervates. Food Hydrocolloid. 2014, 36, 181–188. [Google Scholar] [CrossRef]
- Figueroa-Espinoza, M.C.; Rouau, X. Oxidative crosslinking of pentosans by a fungal laccase and horseradish peroxidase: Mechanism of linkage between feruloylated arabinoxylans. Cereal Chem. 1998, 75, 259–265. [Google Scholar] [CrossRef]
- AACC. Approved Methods of the American Association of Cereal Chemists, 10th ed.; AACC: St. Paul, MN, USA, 2000. [Google Scholar]
- Carvajal-Millan, E.; Guigliarelli, B.; Belle, V.; Rouau, X.; Micard, V. Storage stability of arabinoxylan gels. Carbohydr. Polym. 2005, 59, 181–188. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the arabinoxylans are not available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Morales-Ortega, A.; Carvajal-Millan, E.; Brown-Bojorquez, F.; Rascón-Chu, A.; Torres-Chavez, P.; López-Franco, Y.L.; Lizardi-Mendoza, J.; Martínez-López, A.L.; Campa-Mada, A.C. Entrapment of Probiotics in Water Extractable Arabinoxylan Gels: Rheological and Microstructural Characterization. Molecules 2014, 19, 3628-3637. https://doi.org/10.3390/molecules19033628
Morales-Ortega A, Carvajal-Millan E, Brown-Bojorquez F, Rascón-Chu A, Torres-Chavez P, López-Franco YL, Lizardi-Mendoza J, Martínez-López AL, Campa-Mada AC. Entrapment of Probiotics in Water Extractable Arabinoxylan Gels: Rheological and Microstructural Characterization. Molecules. 2014; 19(3):3628-3637. https://doi.org/10.3390/molecules19033628
Chicago/Turabian StyleMorales-Ortega, Adriana, Elizabeth Carvajal-Millan, Francisco Brown-Bojorquez, Agustín Rascón-Chu, Patricia Torres-Chavez, Yolanda L. López-Franco, Jaime Lizardi-Mendoza, Ana L. Martínez-López, and Alma C. Campa-Mada. 2014. "Entrapment of Probiotics in Water Extractable Arabinoxylan Gels: Rheological and Microstructural Characterization" Molecules 19, no. 3: 3628-3637. https://doi.org/10.3390/molecules19033628
APA StyleMorales-Ortega, A., Carvajal-Millan, E., Brown-Bojorquez, F., Rascón-Chu, A., Torres-Chavez, P., López-Franco, Y. L., Lizardi-Mendoza, J., Martínez-López, A. L., & Campa-Mada, A. C. (2014). Entrapment of Probiotics in Water Extractable Arabinoxylan Gels: Rheological and Microstructural Characterization. Molecules, 19(3), 3628-3637. https://doi.org/10.3390/molecules19033628