Antibacterial and Antioxidant Activity of Essential Oil Terpenes against Pathogenic and Spoilage-Forming Bacteria and Cell Structure-Activity Relationships Evaluated by SEM Microscopy

Abstract
:1. Introduction
2. Results and Discussion
2.1. Determination of Antioxidant Acitivity by FRAP and DPPH Method

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
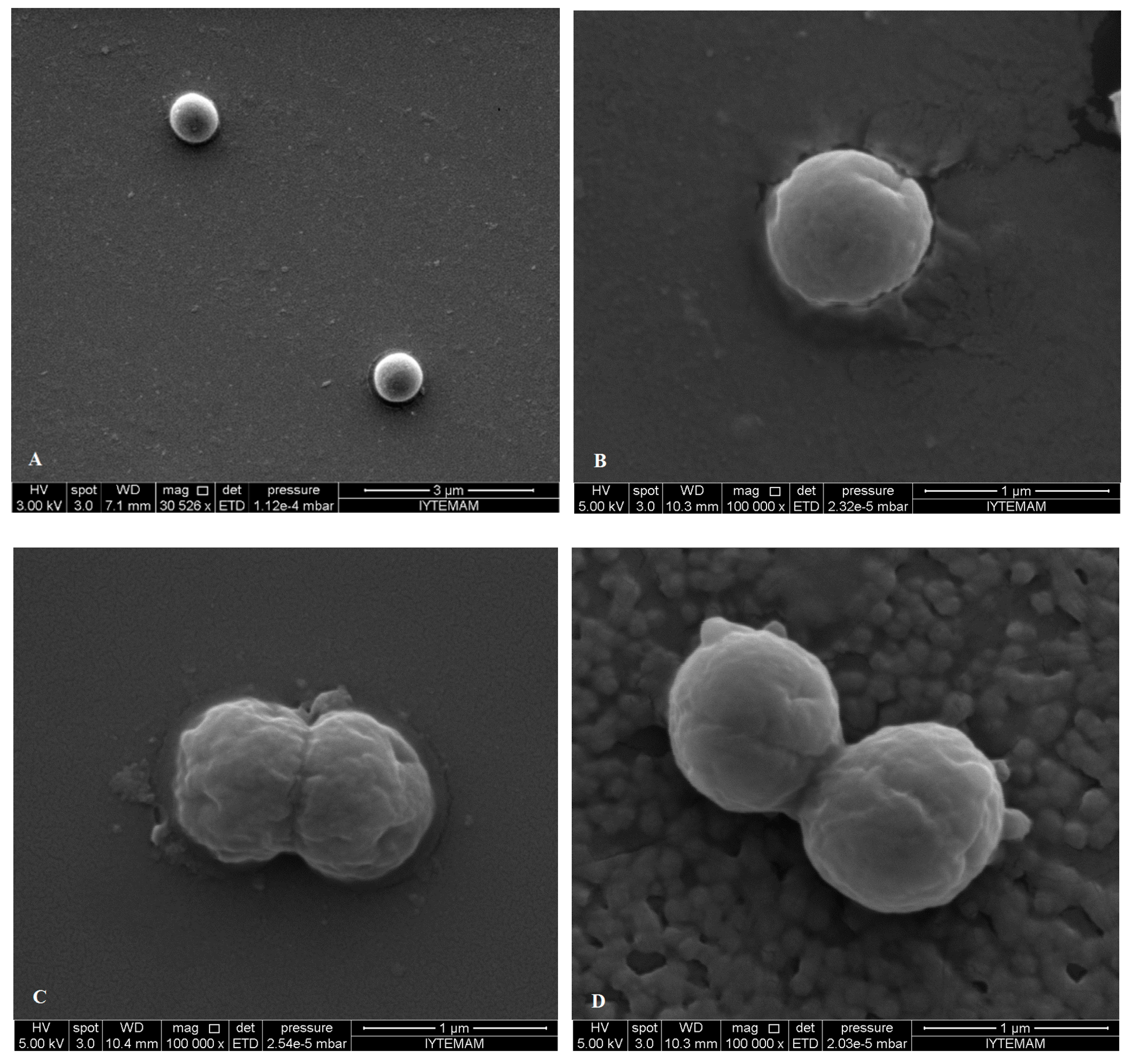{kind=link}
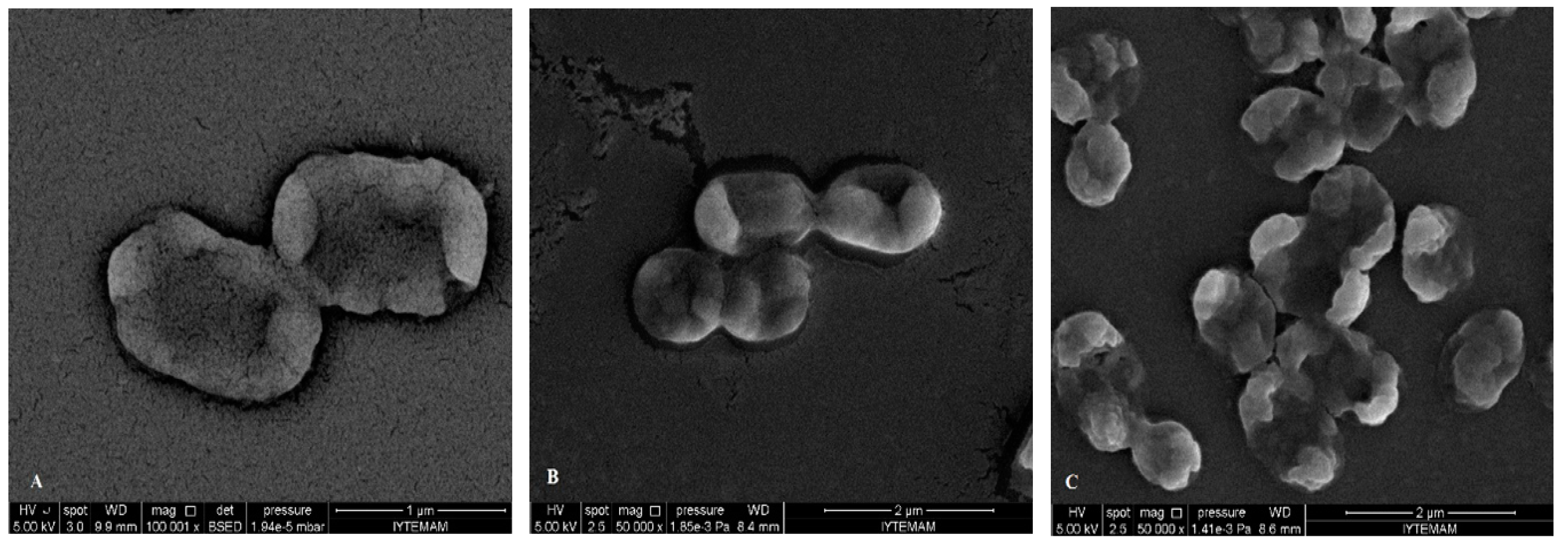{kind=link}
{kind=link}
| EO Component | FRAP (mmol Trolox mL−1) | DPPH (IC50 µL·mL−1) |
|---|---|---|
| Eucalyptol | 0.58 ± 0.22 | NA |
| α-terpineol | 1.23 ± 0.56 | 433.97 ± 13.69 |
| Linalool | 1.20 ± 0.27 | 325.05 ± 20.19 |
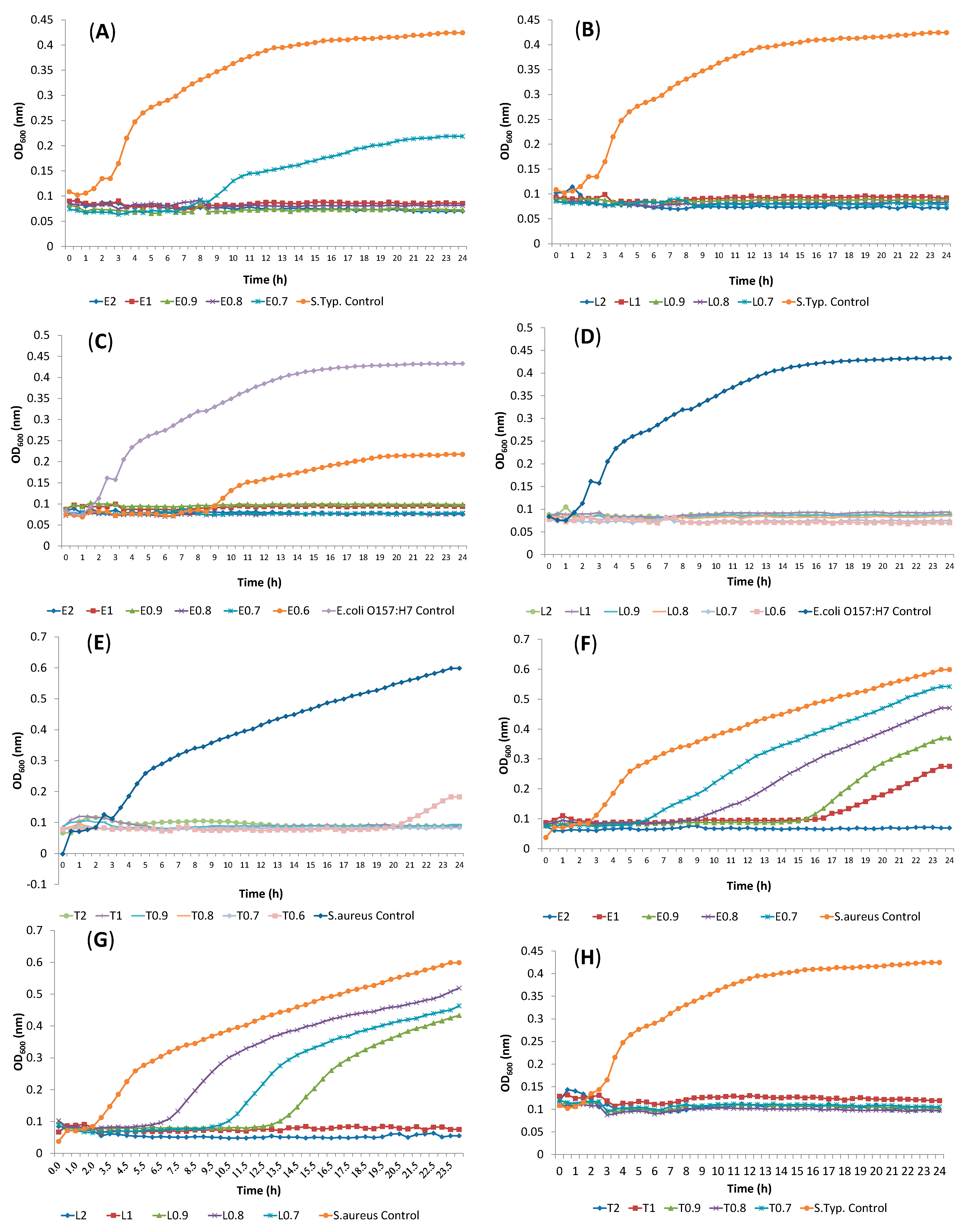
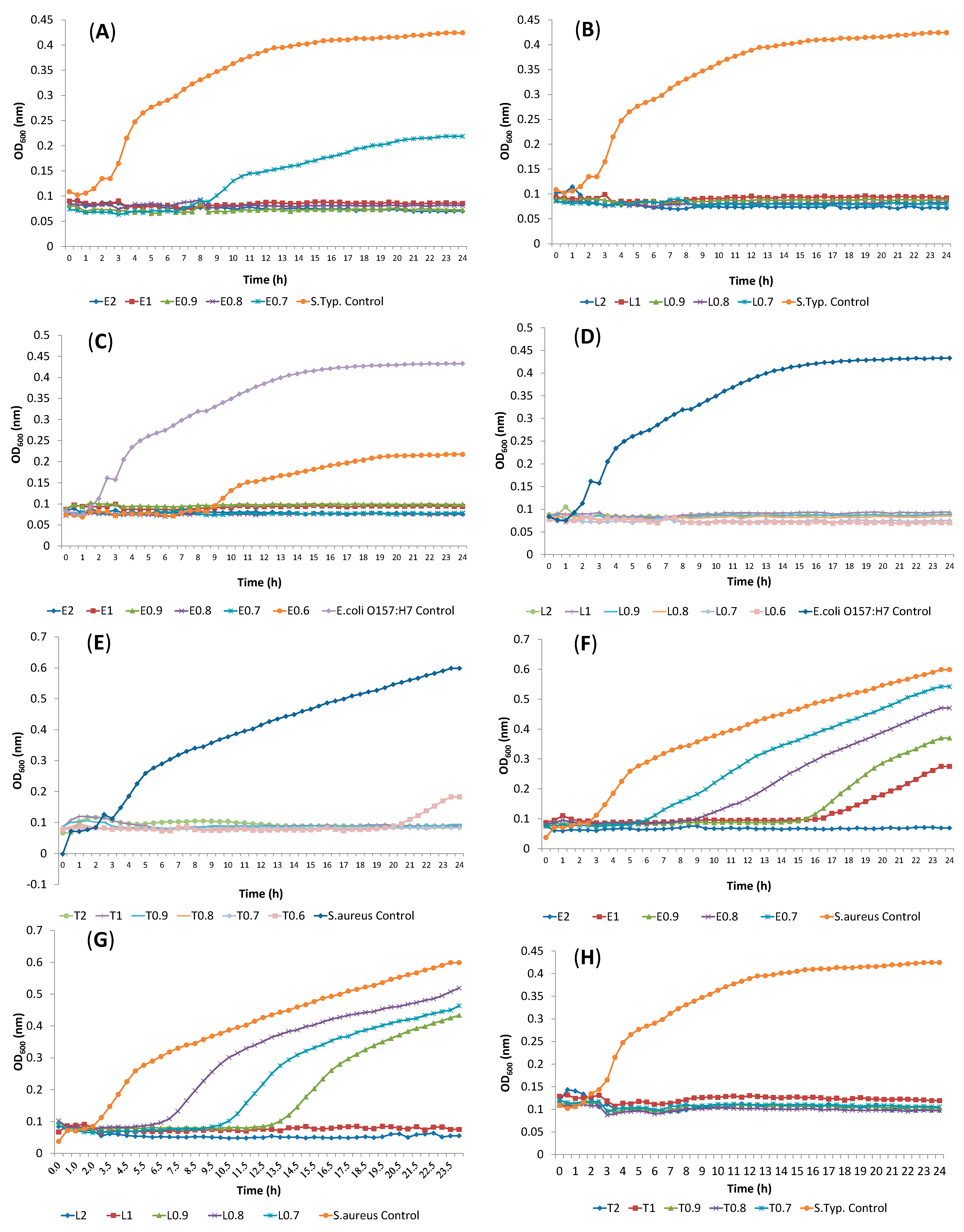
2.2. Determination of Antibacterial Activity and Minimal Inhibitory Concentration (MIC)
| Bacteria | MIC (%) for EO Components (v/v) * | ||
|---|---|---|---|
| α-Terpineol (T) | Linalool (L) % | Eucalyptol (E) % | |
| Gram Positive Bacteria | |||
| Staphylococcus aureus | 0.7 | 1 | 2 |
| Carnobacterium divergens | 0.6 | 2 | 2 |
| Listeria innocua | 0.6 | 1 | >2 |
| Gram Negative Bacteria | |||
| Shewanella putrefaciens | 2 | >2 | >2 |
| Serratia liquefaciens | 0.6 | 1 | >2 |
| E.coli O157:H7 | 0.6 | 0.6 | 0.7 |
| Salmonella Typhimurium | 0.7 | 0.7 | 0.8 |
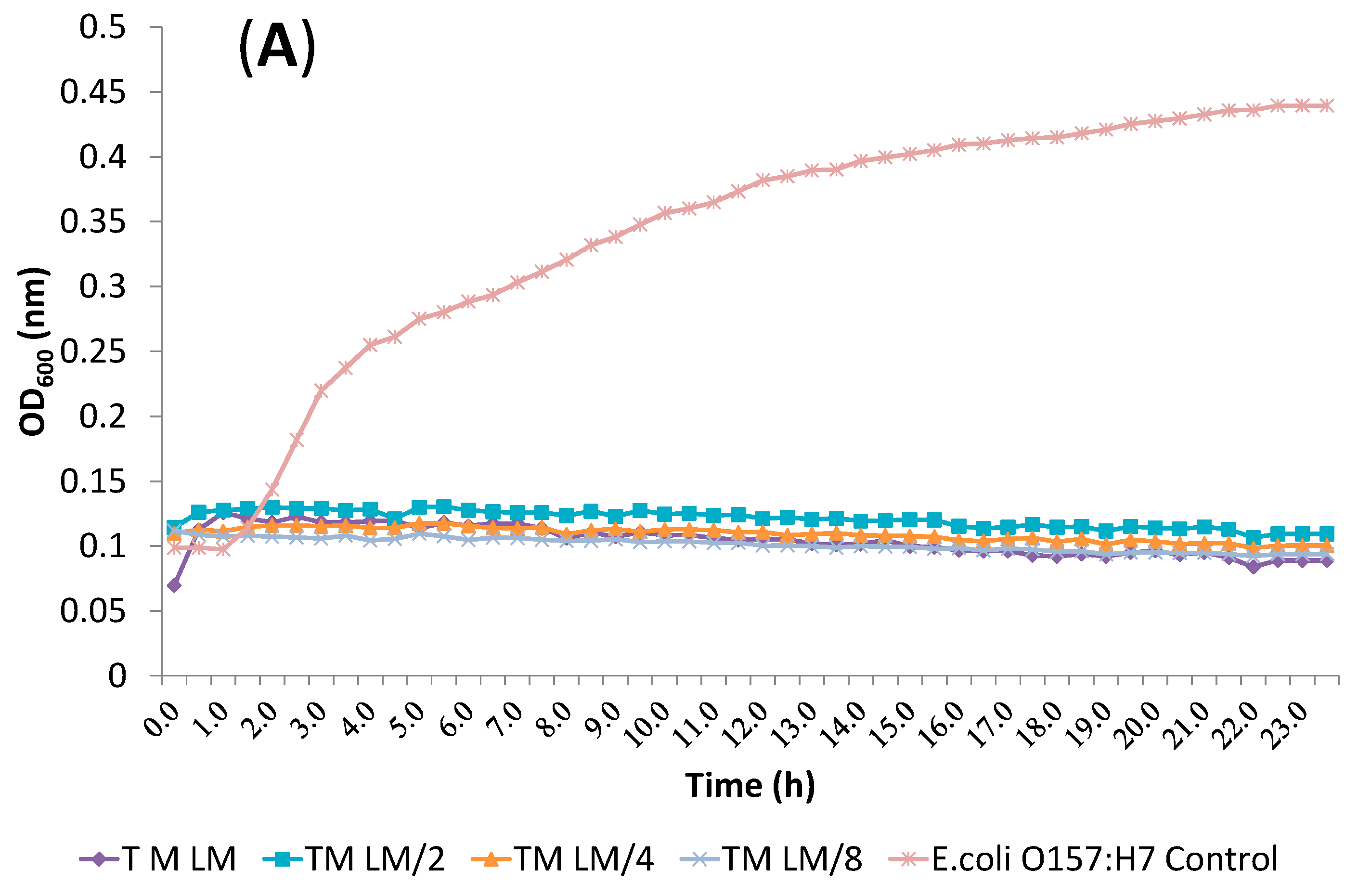
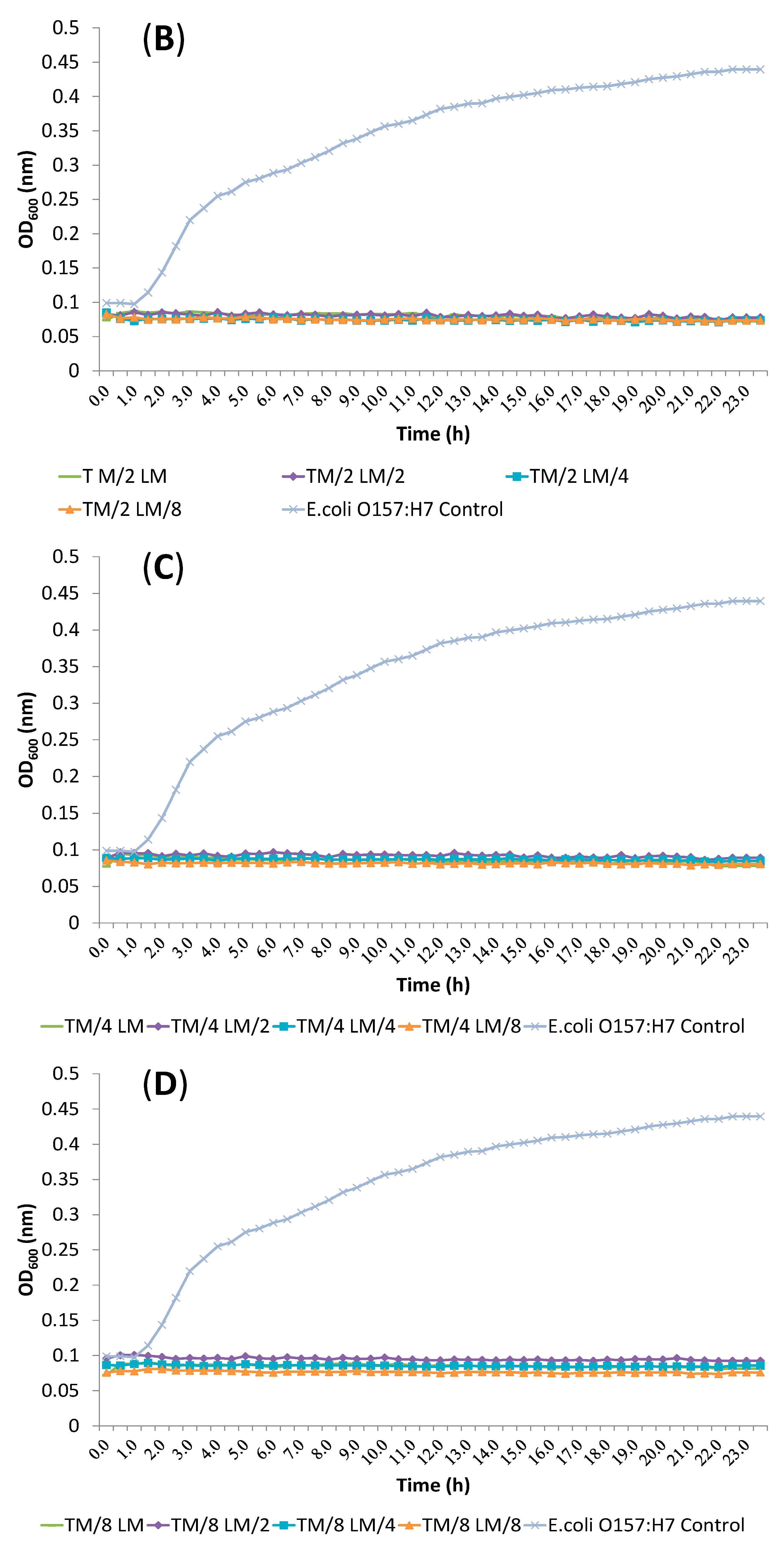
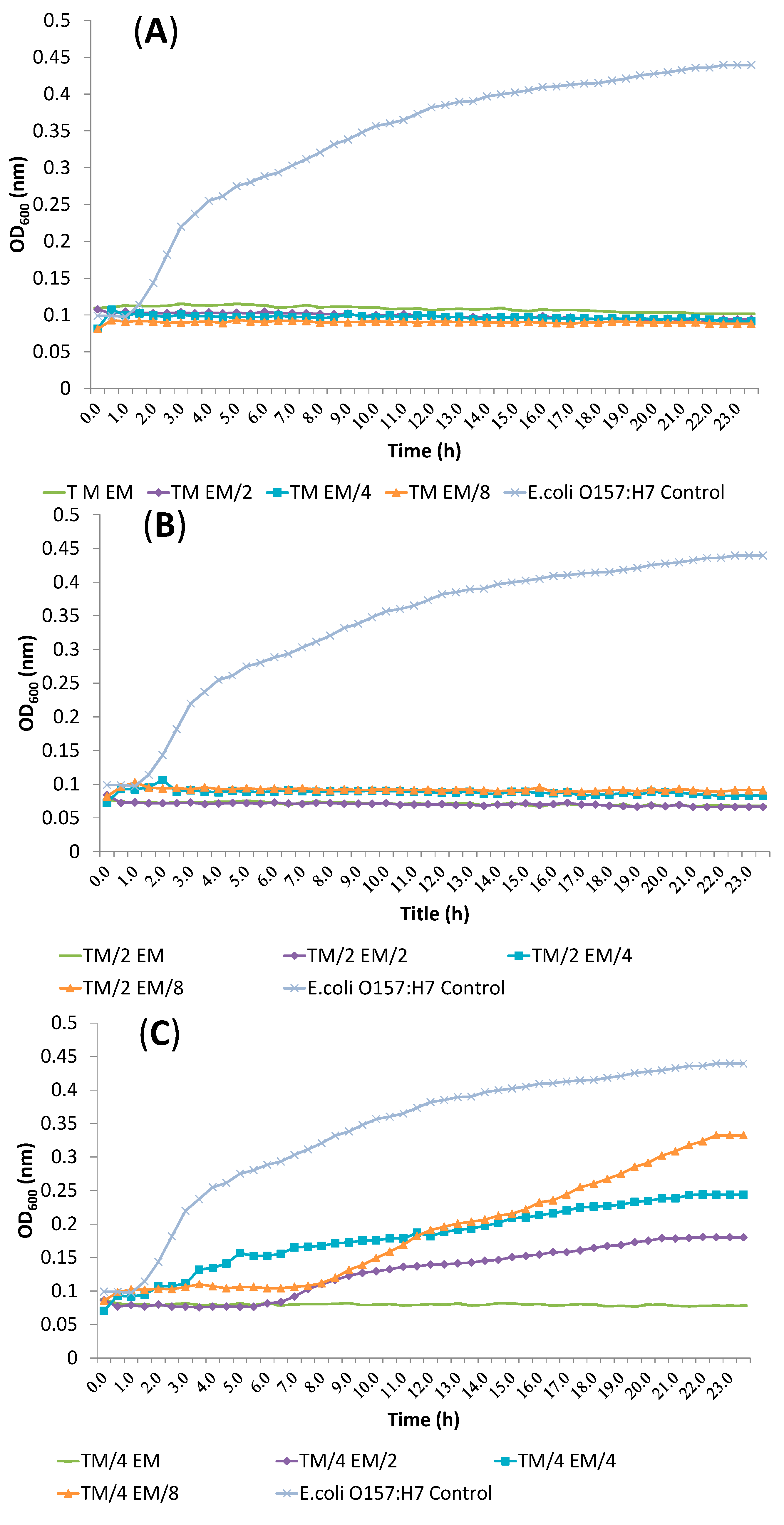
2.3. Synergism Testing of Constituents of Essential Oils
| Bacterium | Components | MIC (%) | Combined MIC (%) | FIC | Effect | |||
|---|---|---|---|---|---|---|---|---|
| A | B | A | B | A | B | |||
| E. coli O157:H7 | α-terpineol | linalool | 0.6 | 0.6 | 0.3 | 0.3 | 1 | Additive |
| α-terpineol | linalool | 0.6 | 0.6 | 0.15 | 0.15 | 0.5 | Synergy | |
| α-terpineol | eucalyptol | 0.6 | 0.7 | 0.3 | 0.35 | 1 | Additive | |
| linalool | eucalyptol | 0.6 | 0.7 | 0.3 | 0.35 | 1 | Additive | |
| S. Typhimurium | α-terpineol | linalool | 0.7 | 0.7 | 0.35 | 0.35 | 1 | Additive |
| α-terpineol | linalool | 0.7 | 0.7 | 0.175 | 0.175 | 0.5 | Synergy | |
| α-terpineol | eucalyptol | 0.7 | 0.8 | 0.35 | 0.4 | 1 | Additive | |
| linalool | eucalyptol | 0.7 | 0.8 | 0.35 | 0.4 | 1 | Additive | |
| S. aureus | α-terpineol | linalool | 0.7 | 1 | 0.35 | 0.5 | 1 | Additive |
| α-terpineol | linalool | 0.7 | 1 | 0.175 | 0.25 | 0.5 | Synergy | |
| α-terpineol | eucalyptol | 0.7 | 2 | 0.35 | 1 | 1 | Additive | |
| linalool | eucalyptol | 1 | 2 | 0.35 | 1 | 1 | Additive | |
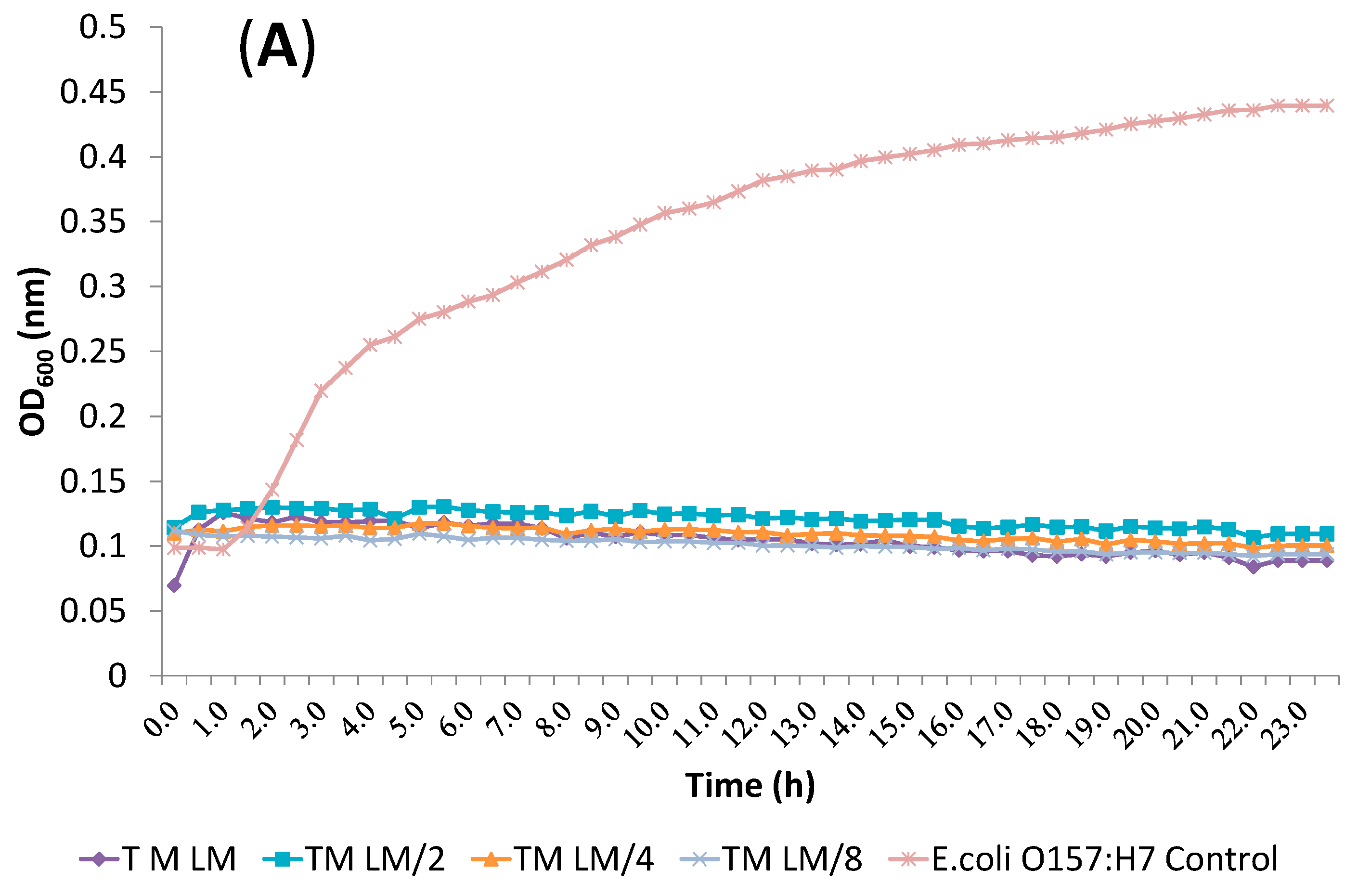

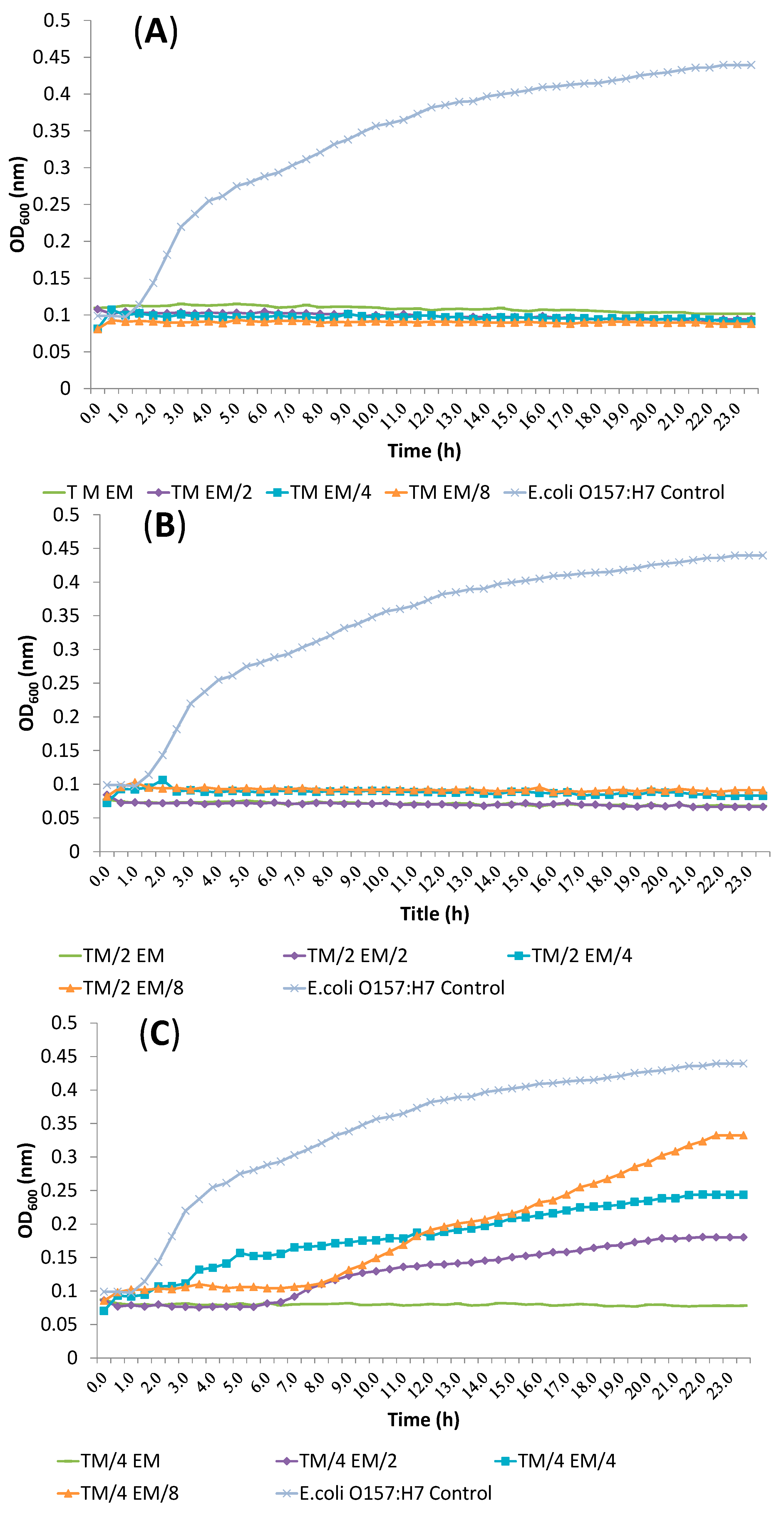
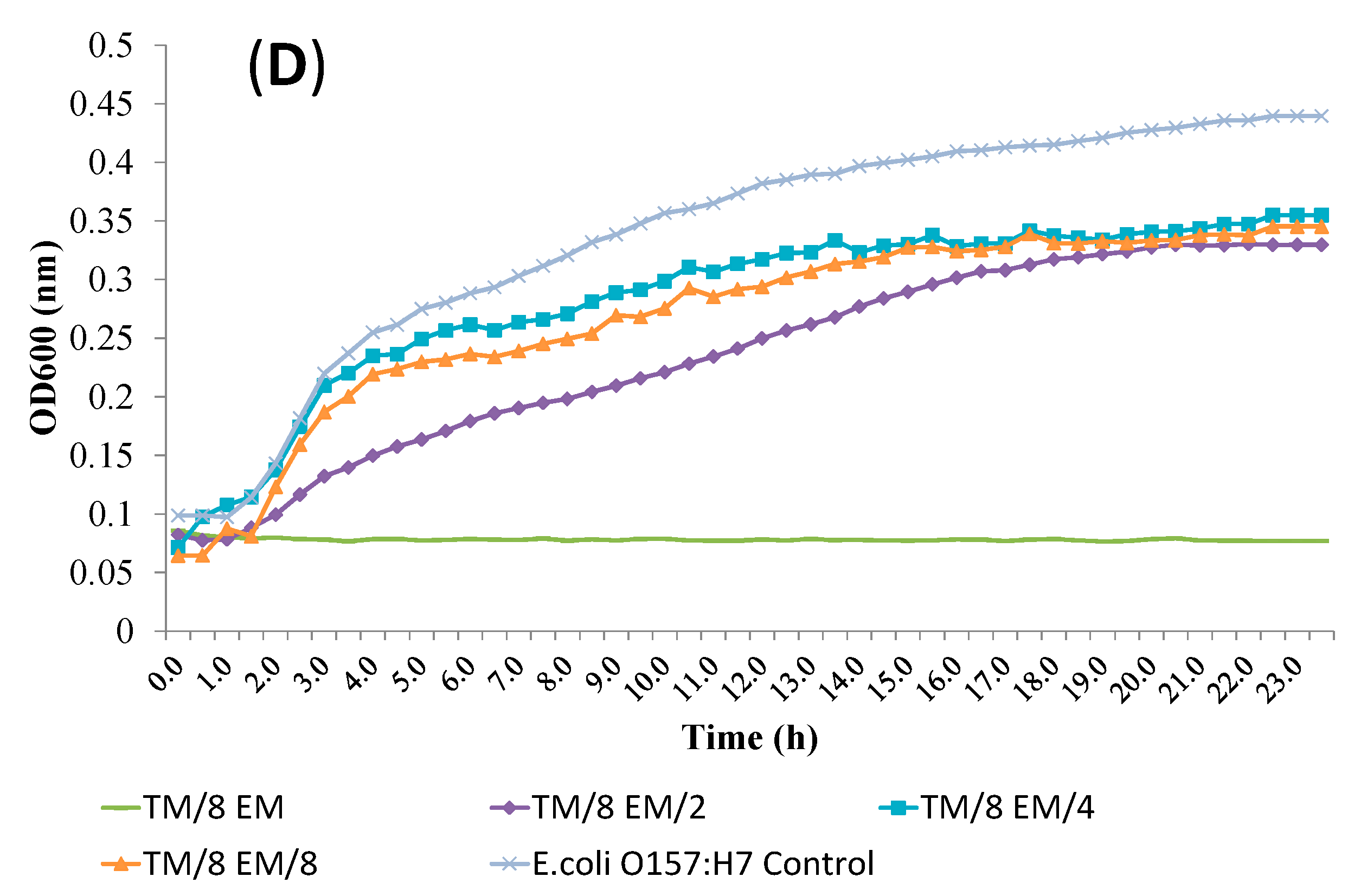
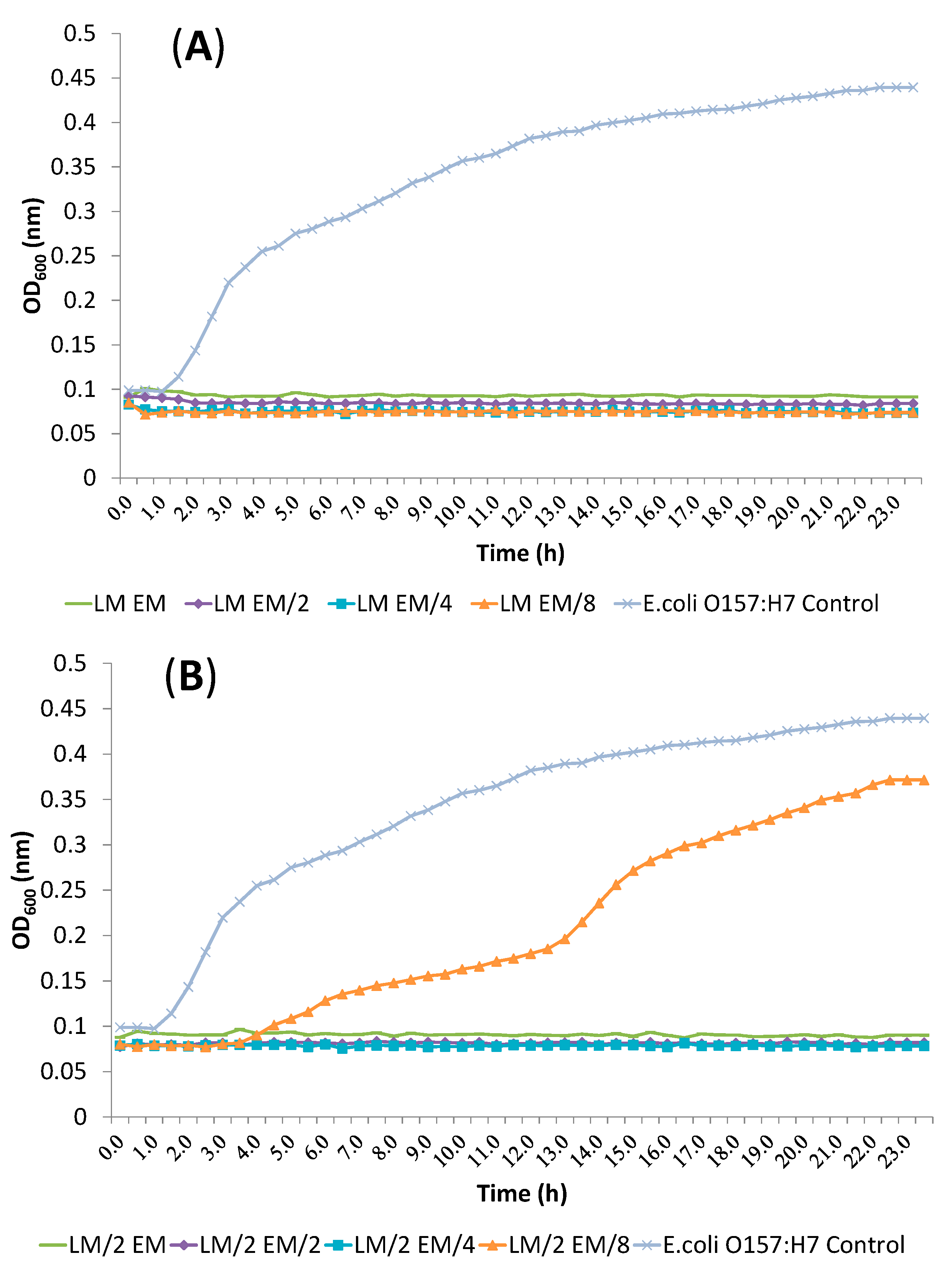
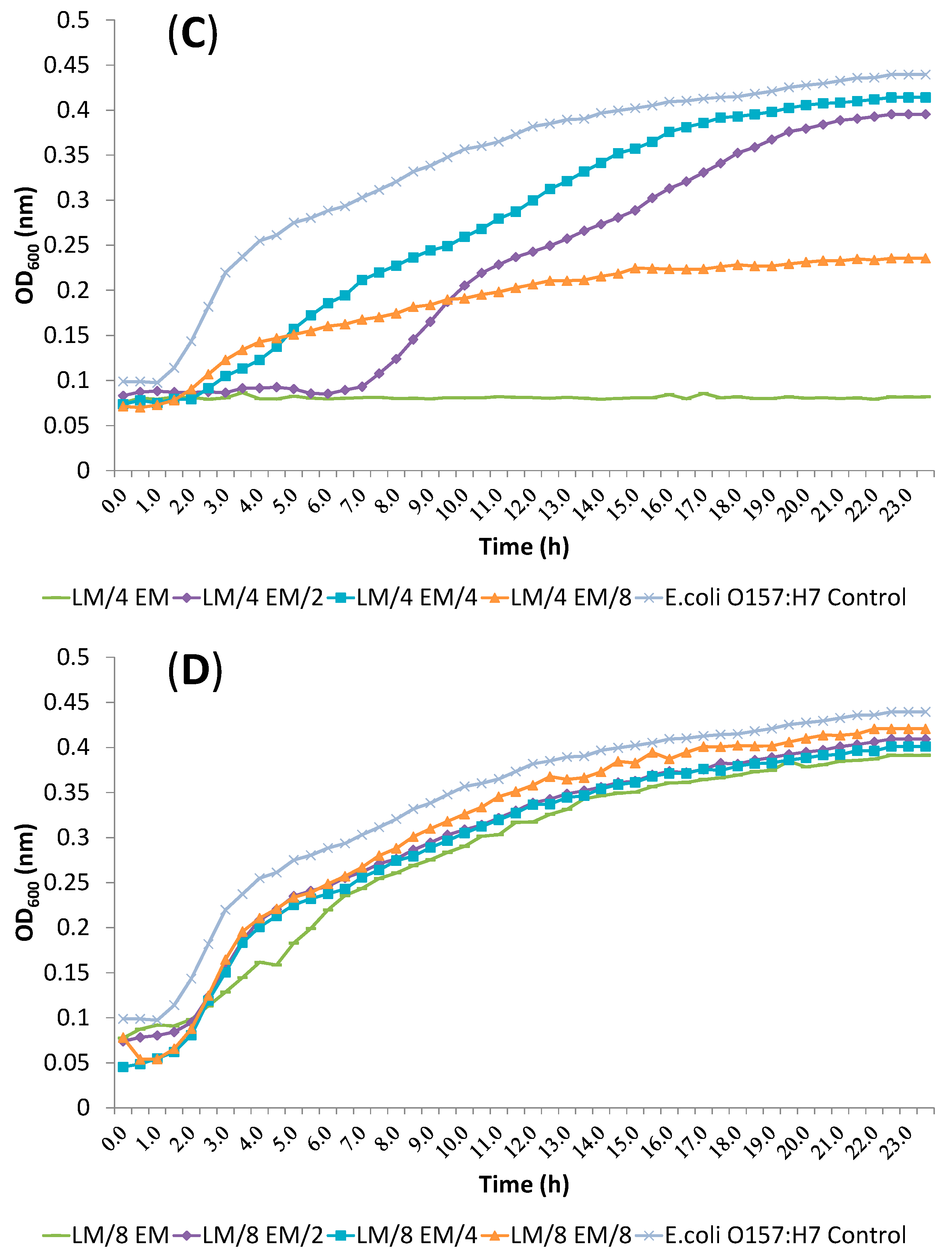
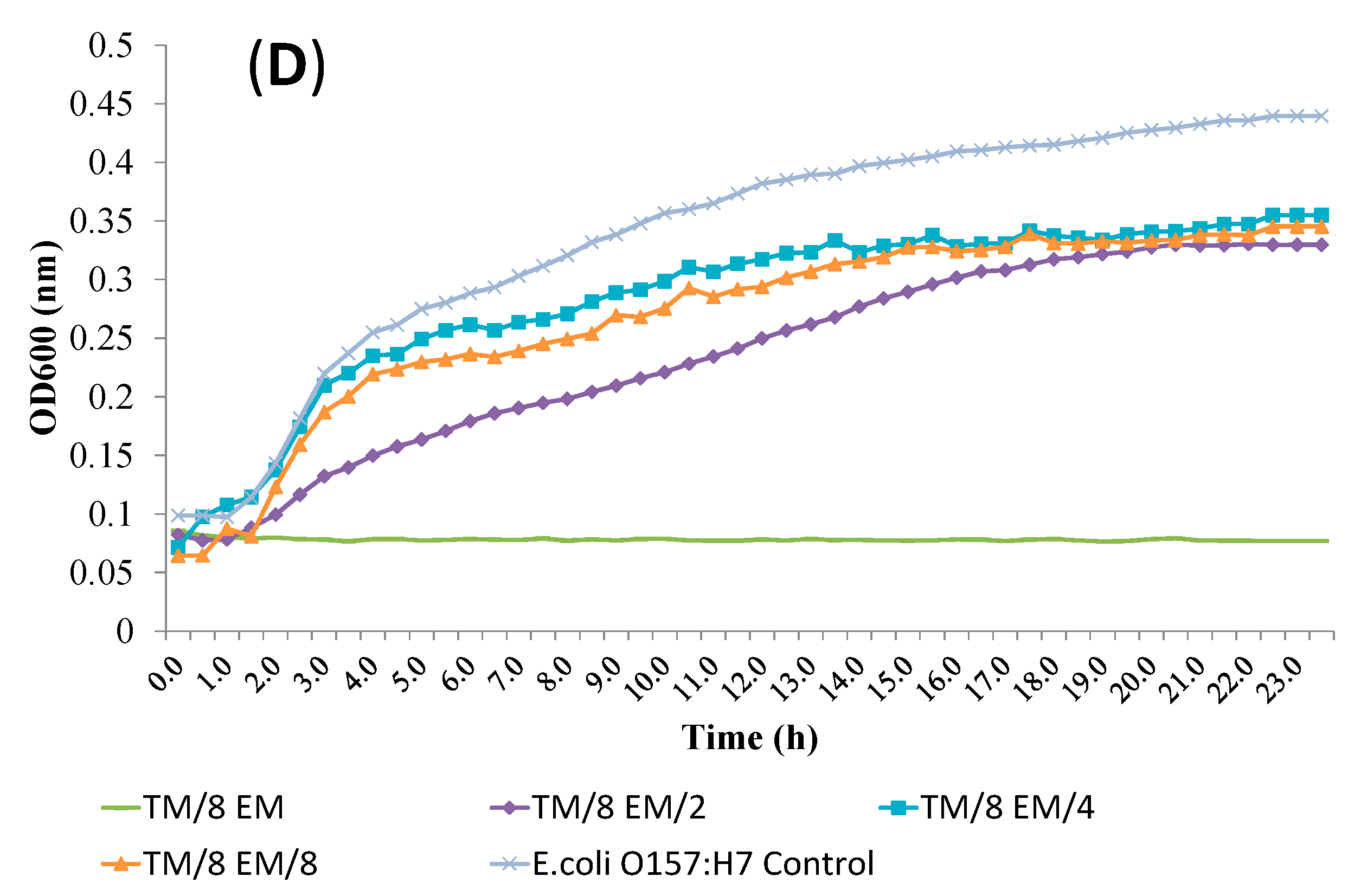
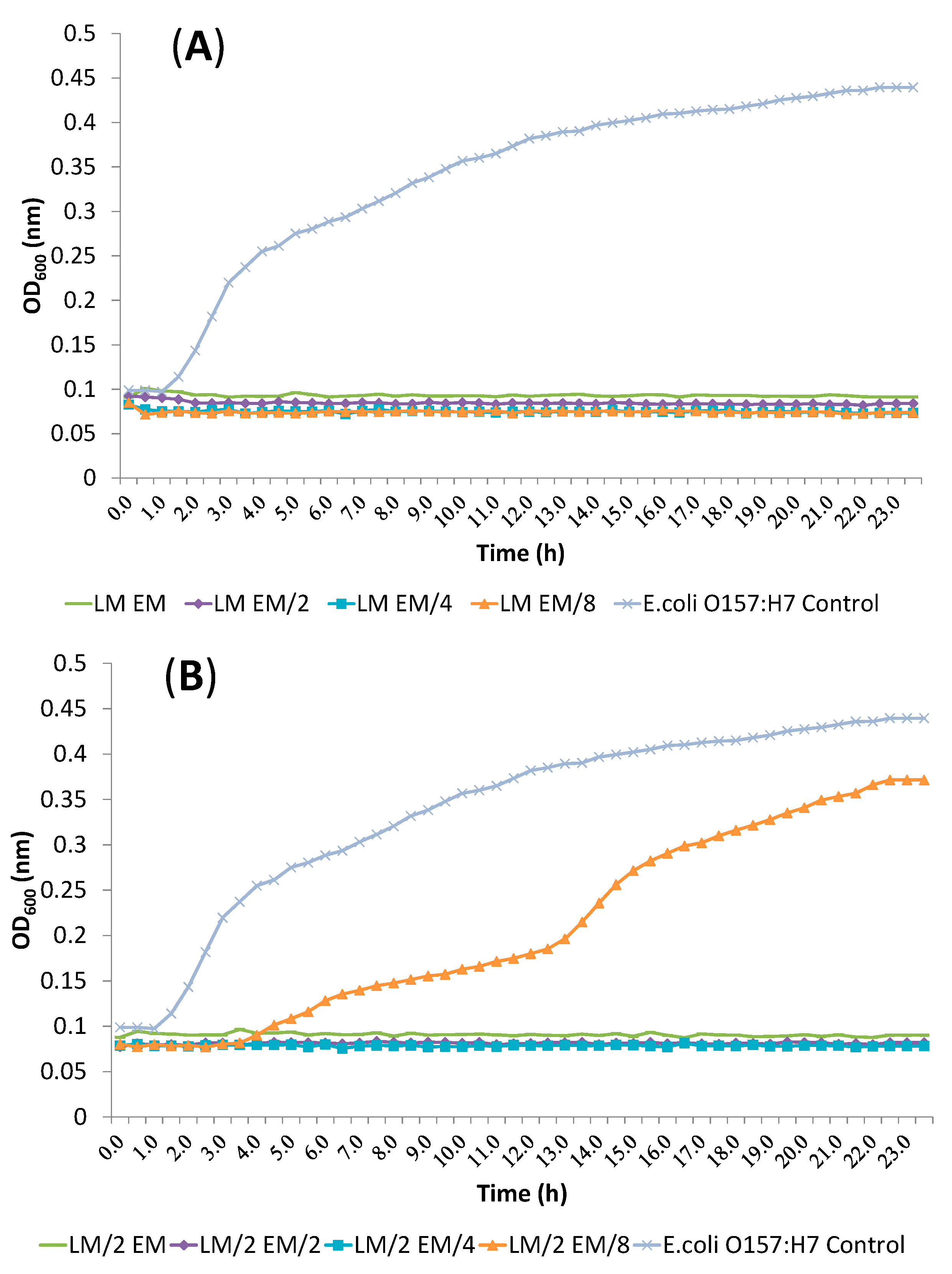
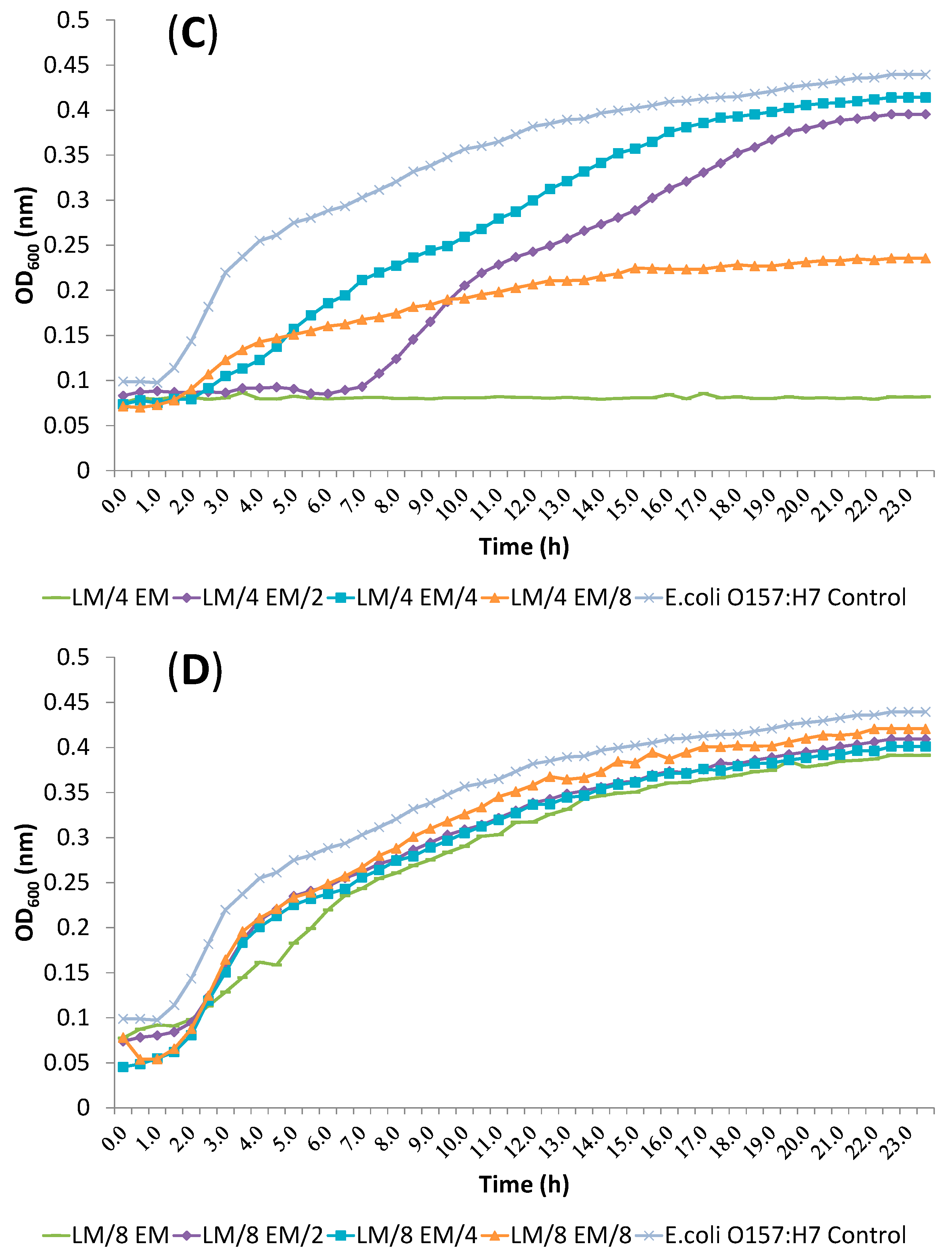
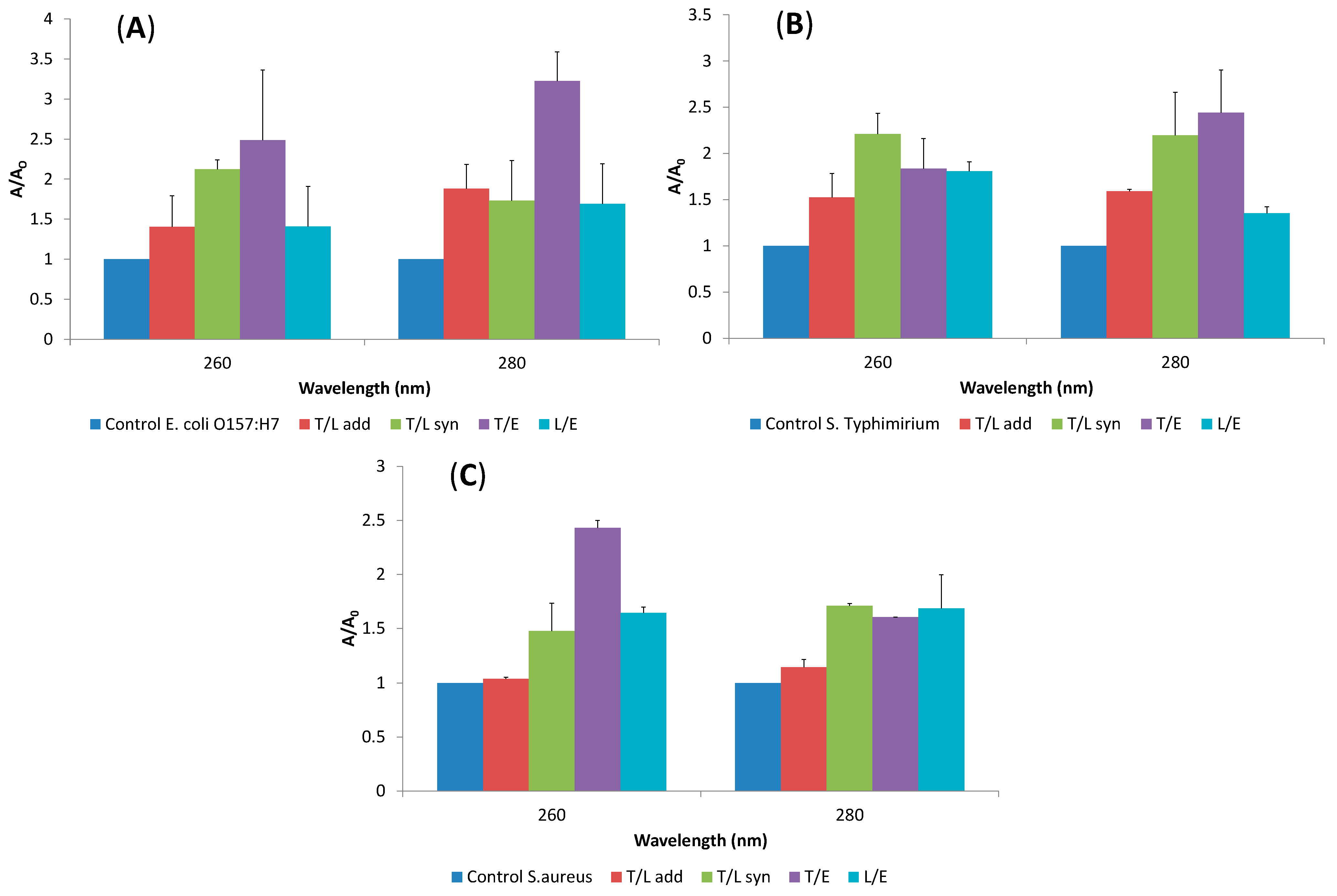
2.4. Release of Cellular Material after Treatment with Essential Oil Components
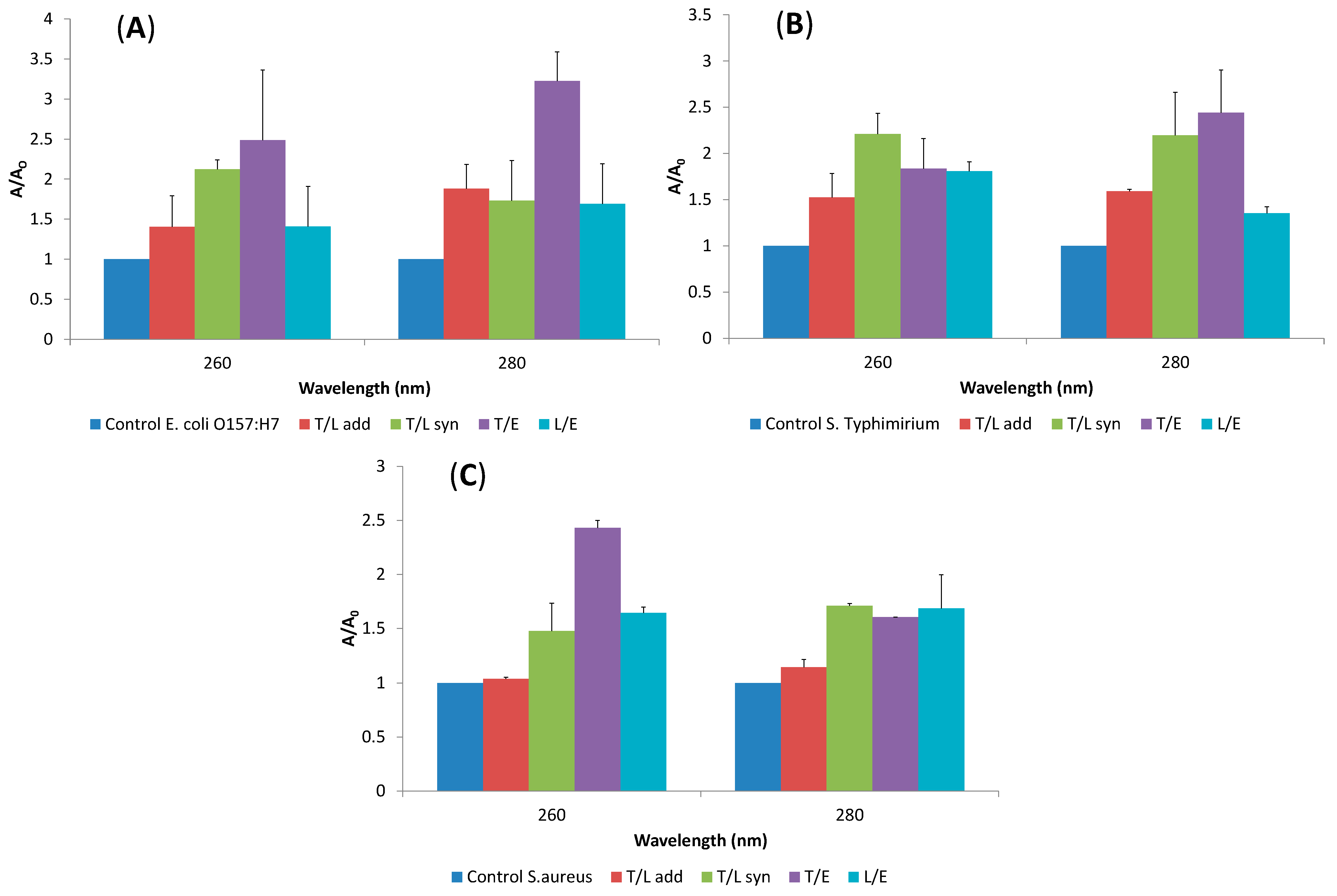
2.5. SEM Microscopy

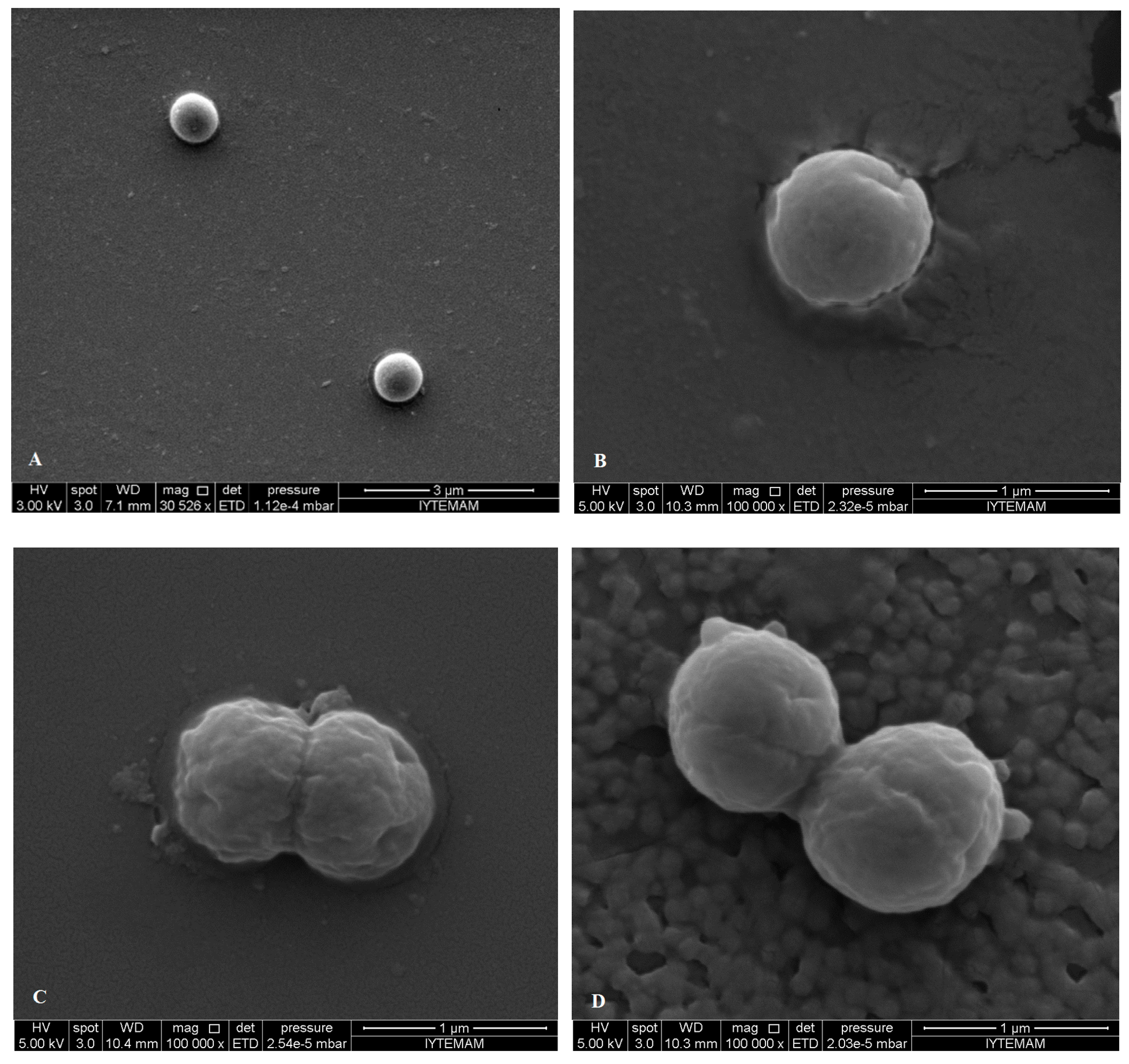
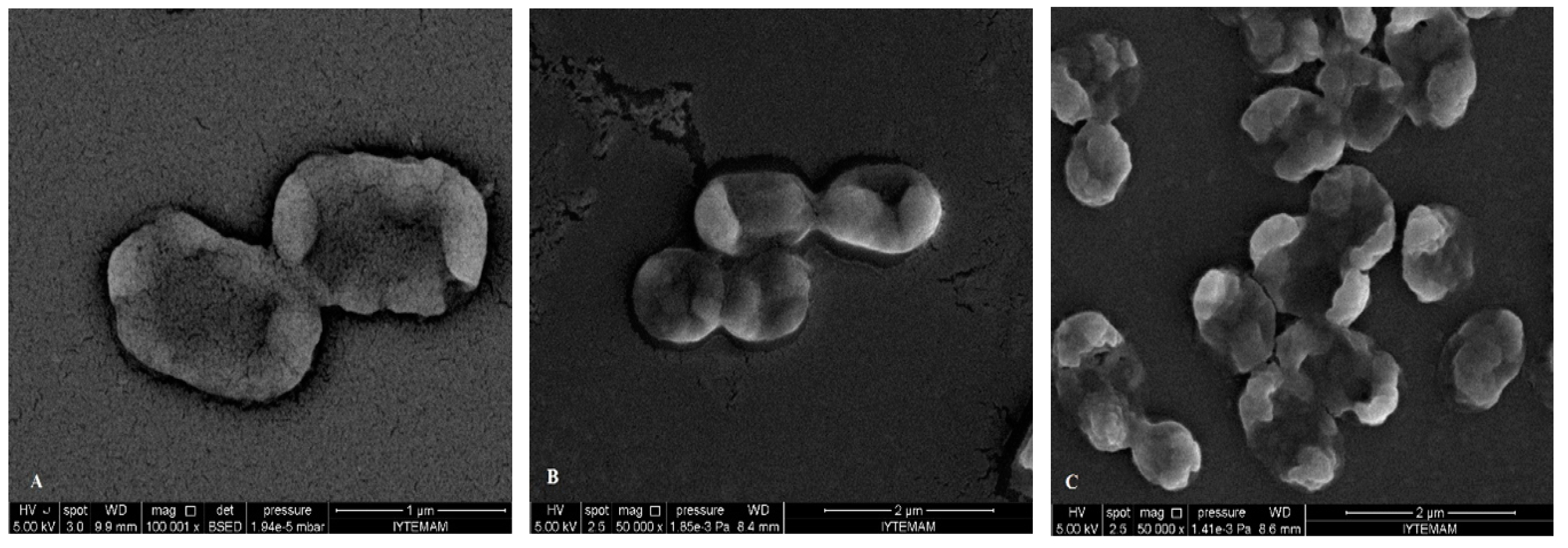

3. Experimental Section
3.1. Essential Oil Components
3.2. Bacterial Strains
3.3. Preparation of Bacterial Suspensions
3.4. Determination of Antioxidant Activity by the FRAP Method
3.5. Determination of Antioxidant Activity by the DPPH Method
3.6. Minimal Inhibitory Concentration (MIC) by Broth Micro- and Macro-Dilution Method
3.7. Synergism Testing of Terpenes by Checkerboard Method
3.8. Release of Cellular Material
3.9. SEM Microscopy
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Burt, S.A.; Reinders, R.D. Antibacterial activity of selected plant essential oils against Escherichia coli O157:H7. Lett. Appl. Microbiol. 2003, 36, 162–167. [Google Scholar] [PubMed]
- Dadalioğlu, I.; Evrendilek, G.A. Chemical compositions and antibacterial effects of essential oils of Turkish oregano (Origanum minutiflorum), bay laurel (Laurus nobilis), Spanish lavender (Lavandula stoechas L.), and fennel (Foeniculum vulgare) on common foodborne pathogens. J. Agric. Food Chem. 2004, 52, 8255–8260. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Henika, P.R.; Mandrell, R.E. Bactericidal activities of plant essential oils and some of their isolated constituents against Campylobacter jejuni, Escherichia coli, Listeria monocytogenes, and Salmonella enterica. J. Food Prot. 2002, 65, 1545–1560. [Google Scholar] [PubMed]
- Carson, C.F.; Hammer, K.A. Chemistry and Bioactivity of Essential Oils. In Lipids and Essential Oils as Antimicrobial Agents; Thormar, C., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2011; Chapter 9. [Google Scholar]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.; Barry-Ryan, C.; Bourke, P. Antimicrobial activity of plant essential oils using food model media: Efficacy, synergistic potential and interactions with food components. Food Microbiol. 2009, 26, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.; Liang, H.; Yuan, Q.; Li, C. In vitro antimicrobial effects and mechanism of action of selected plant essential oil combinations against four food-related microorganisms. Food Res. Int. 2011, 44, 3057–3064. [Google Scholar] [CrossRef]
- Farag, R.S.; Daw, Z.Y.; Hewedi, F.M.; El-Baroty, G.S.A. Antimicrobial activity of some Egyptian spice essential oils. J. Food Prot. 1989, 52, 665–667. [Google Scholar]
- Moleyar, V.; Narasimham, P. Antibacterial activity of essential components. Int. J. Food Microbiol. 1992, 16, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Marshall, M.R.; Wei, C.-I. Antibacterial activity of some essential oil components against five foodborne pathogens. J. Agric. Food Chem. 1995, 3, 2839–2845. [Google Scholar] [CrossRef]
- Tsao, R.; Zhou, T. Antifungal activity of monoterpenoids against postharvest pathogens Botrytis cinerea and Monilinia fructicola. J. Essent. Oil Res. 2000, 12, 113–121. [Google Scholar] [CrossRef]
- Lambert, R.J.W.; Skandamis, P.N.; Coote, P.J.; Nychas, G.J.E. A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar]
- Ben Arfa, A.; Combes, S.; Preziosi-Belloy, L.; Gontard, N.; Chalier, P. Antimicrobial activity of carvacrol related to its chemical structure. Lett. Appl. Microbiol. 2006, 43, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Kapros, T.; McDaniel, S.C. Cytotoxicity of tea tree oil in tobacco cells. Allelopath. J. 2009, 23, 185–192. [Google Scholar]
- Di Pasqua, R.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane toxicity of antimicrobial compounds from essential oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.Z.; Sun, M.; Corke, H. Antioxidant capacity of 26 spice extracts and characterization of their phenolic constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar] [CrossRef] [PubMed]
- Gourine, N.; Yousfi, M.; Bombarda, I.; Nadjemi, B.; Stocker, P.; Gaydou, E.M. Antioxidant activities and chemical composition of essential oil of Pistacia atlantica from Algeria. Ind. Crop Prod. 2010, 31, 203–208. [Google Scholar] [CrossRef]
- Politeo, O.; Jukic, M.; Milos, M. Comparison of chemical composition and antioxidant activity of glycosidically bound and free volatiles from clove (Eugenia caryophyllata Thunb.). J. Food Biochem. 2010, 34, 129–141. [Google Scholar] [CrossRef]
- Tepe, B.; Sokmen, M.; Akpulat, H.A.; Daferera, D.; Polissiou, M.; Sokmen, A. Antioxidative activity of the essential oils of Thymus sipyleus subsp. sipyleus var. sipyleus and Thymus sipyleus subsp. sipyleus var. rosulans. J. Food Eng. 2005, 66, 447–454. [Google Scholar]
- Candan, F.; Unlu, M.; Tepe, B.; Daferera, D.; Polissiou, M.; Sökmen, A.; Akpulat, H.A. Antioxidant and antimicrobial activity of the essential oil and methanol extracts of Achillea millefolium subsp. millefolium Afan. (Asteraceae). J. Ethnopharmacol. 2003, 87, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Viuda-Martos, M.; López-Marcos, M.C.; Fernández-López, J.; Sendra, E.; López-Vargas, J.H.; Pérez-Álvarez, J.A. Role of fiber in cardiovascular diseases: A review. Compr. Rev. Food Sci. Food Saf. 2010, 9, 240–258. [Google Scholar] [CrossRef]
- Lee, K.G.; Shibamoto, T. Antioxidant activities of volatile components isolated from Eucalyptus species. J. Sci. Food Agric. 2001, 81, 1573–1579. [Google Scholar] [CrossRef]
- Ruberto, G.; Baratta, M.T. Antioxidant activity of selected essential oil components in two lipid model systems. Food Chem. 2000, 69, 167–174. [Google Scholar] [CrossRef]
- De Sousa, J.P.; de Azeredo, G.A.; de Araujo Torres, R.; da Silva Vasconcelos, M.A.; da Conceicao, M.L.; de Souza, E.L. Synergies of carvacrol and 1,8-cineole to inhibit bacteria associated with minimally processed vegetables. Int. J. Food Microbiol. 2012, 154, 145–151. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, J.P.; Torres Rde, A.; de Azeredo, G.A.; Figueiredo, R.C.; Vasconcelos, M.A.; de Souza, E.L. Carvacrol and 1,8-cineole alone or in combination at sublethal concentrations induce changes in the cell morphology and membrane permeability of Pseudomonas fluorescens in a vegetable-based broth. Int. J. Food Microbiol. 2012, 158, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Gomes Neto, N.J.; da Silva Luz, I.; Honório, V.G.; da Conceição, M.L.; de Souza, E.L. Pseudomonas aeruginosa cells adapted to Rosmarinus officinalis L. essential oil and 1,8-cineole acquire no direct and cross protection in a meat-based broth. Food Res. Int. 2012, 49, 143–146. [Google Scholar] [CrossRef]
- Park, S.N.; Lim, Y.K.; Freire, M.O.; Cho, E.; Jin, D.; Kook, J.K. Antimicrobial effect of linalool and alpha-terpineol against periodontopathic and cariogenic bacteria. Anaerobe 2012, 18, 369–372. [Google Scholar] [CrossRef]
- Aboul Ela, M.A.; El-Shaer, N.S.; Ghanem, N.B. Antimicrobial evaluation and chromatographic analysis of some essential and fixed oils. Pharmazie 1996, 51, 993–994. [Google Scholar] [PubMed]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and antifungal activity of aromatic constituents of essential oils. Microbios 1997, 89, 39–46. [Google Scholar] [PubMed]
- Wan, J.; Wilcock, A.; Coventry, M.J. The effect of essential oils of basil on the growth of Aeromonas hydrophila and Pseudomonas fluorescens. J. Appl. Microbiol. 1998, 84, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Ojeda-Sana, A.M.; van Baren, C.M.; Elechosa, M.A.; Juárez, M.A.; Moreno, S. New insights into antibacterial and antioxidant activities of rosemary essential oils and their main components. Food Cont. 2013, 31, 189–195. [Google Scholar] [CrossRef]
- Ait-Ouazzou, A.; Cherrat, L.; Espina, L.; Lorán, S.; Rota, C.; Pagán, R. The antimicrobial activity of hydrophobic essential oil constituents acting alone or in combined processes of food preservation. Innov. Food Sci. Emerg. Technol. 2011, 12, 320–329. [Google Scholar] [CrossRef]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of antibacterial action of three monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [PubMed]
- Sikkema, J.; de Bont, J.A.M.; Poolman, B. Interactions of cyclic hydrocarbons with biological membranes. J. Biol. Chem. 1994, 269, 8022–8028. [Google Scholar] [PubMed]
- Sikkema, J.; de Bont, J.A.M.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [PubMed]
- Helander, I.M.; Alakomi, H.-L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Bassolé, I.H.N.; Lamien-Meda, A.; Bayala, B.; Tirogo, S.; Franz, C.; Novak, J.; Nebié, R.C.; Dicko, M.H. Composition and antimicrobial activities of Lippia multiflora Moldenke, Mentha piperita L. and Ocimum basilicum L. essential oils and their major monoterpene alcohols alone and in combination. Molecules 2010, 15, 7825–7839. [Google Scholar] [CrossRef]
- Gallucci, M.N.; Oliva, M.; Casero, C.; Dambolena, J.; Luna, A.; Zygadlo, J.; Demo, M. Antimicrobial combined action of terpenes against the food-borne microorganisms Escherichia coli, Staphylococcus aureus and Bacillus cereus. Flavour. Fragr. J. 2009, 24, 348–354. [Google Scholar] [CrossRef]
- Pei, R.S.; Zhou, F.; Ji, B.P.; Xu, J. Evaluation of combined antibacterial effects of eugenol, cinnamaldehyde, thymol, and carvacrol against E. coli with an improved method. J. Food Sci. 2009, 74, M379–M383. [Google Scholar] [CrossRef] [PubMed]
- De Azeredo, G.A.; Stamford, T.L.M.; Nunes, P.C.; Gomes Neto, N.J.; de Oliveira, M.E.G.; de Souza, E.L. Combined application of essential oils from Origanum vulgare L. and Rosmarinus officinalis L. to inhibit bacteria and autochthonous microflora associated with minimally processed vegetables. Food Res. Int. 2011, 44, 1541–1548. [Google Scholar]
- Goñi, P.; López, P.; Sánchez, C.; Gómez-Lus, R.; Becerril, R.; Nerín, C. Antimicrobial activity in the vapour phase of a combination of cinnamon and clove essential oils. Food Chem. 2009, 116, 982–989. [Google Scholar] [CrossRef]
- Gutierrez, J.; Rodriguez, G.; Barry-Ryan, C.; Bourke, P. Efficacy of plant essential oils against food-borne pathogens and spoilage bacteria associated with ready to eat vegetables: Antimicrobial and sensory screening. J. Food Prot. 2008, 71, 1846–1854. [Google Scholar]
- Moosavy, M.H.; Basti, A.A.; Misaghi, A.; Salehi, T.Z.; Abbasifar, R.; Mousavi, H.A.E.; Alipour, M.; Razavi, N.E.; Gandomi, H.; Noori, N. Effect of Zataria multiflora Boiss. essential oil and nisin on Salmonella typhimurium and Staphylococcus aureus in a food model system and on the bacterial cell membranes. Food Res. Int. 2008, 41, 1050–1057. [Google Scholar] [CrossRef]
- Turgis, M.; Vu, K.D.; Dupont, C.; Lacroix, M. Combined antimicrobial effect of essential oils and bacteriocins against foodborne pathogens and food spoilage bacteria. Food Res. Int. 2012, 48, 696–702. [Google Scholar] [CrossRef]
- Karatzas, A.K.; Kets, E.P.; Smid, E.J.; Bennik, M.H. The combined action of carvacrol and high hydrostatic pressure on Listeria monocytogenes Scott A. J. Appl. Microbiol. 2001, 90, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.N.; Juliani, H.R. Essential oils in combination and their antimicrobial properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Delaquis, P.J.; Stanich, K.; Girard, B.; Mazza, G. Antimicrobial activity of individual and mixed fractions of dill, cilantro, coriander and eucalyptus essential oils. Int. J. Food Microbiol. 2002, 74, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Juliani, H.R.; Koroch, A.R.; Simon, J.E. Chemical Diversity of Essential Oils of Ocimum species and Their Associated Antioxidant and Antimicrobial Activity. In Essential Oils and Aromas: Green Extractions and Applications; Chemat, F., Varshney, V.K., Allaf, K., Eds.; Har Krishan Bhalla & Sons: Dehradun, India, 2009. [Google Scholar]
- Lis-Balchin, M.; Deans, S.G. Bioactivity of selected plant essential oil against Listeria monocytogenes. J. Appl. Microbiol. 1997, 82, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Bennik, M.H.J.; Moezelaar, R. The phenolic hydroxyl group of carvacrol is essential for action against the foodborne pathogen Bacillus cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef]
- De Billerbeck, V.G.; Roques, C.G.; Bessière, J.M.; Fonvieille, J.L.; Dargent, R. Effects of Cymbopogon nardus (L.) W. Watson essential oil on the growth and morphogenesis of Aspergillus niger. Can. J. Microbiol. 2001, 47, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, N.; Kamiński, M.; Kurlenda, J.; Asztemborska, M.; Ochocka, J.R. Antibacterial and antifungal activity of juniper berry oil and its selected components. Phytother. Res. 2003, 17, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Packiyasothy, V.E.; Kyle, S. Antimicrobial properties of some herb essential oils. Food Aust. 2002, 54, 384–387. [Google Scholar]
- Shan, B.; Cai, Y.-Z.; Brooks, J.D.; Corke, H. Antibacterial properties and major bioactive components of cinnamon stick (Cinnamomum burmannii): Activity against foodborne pathogenic bacteria. J. Agric. Food Chem. 2007, 55, 5484–5490. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Denyer, S.P. Mechanisms of action of biocides. Int. Biodeterior. 1990, 26, 89–100. [Google Scholar] [CrossRef]
- Ultee, A.; Slump, R.A.; Steging, G.; Smid, E.J. Antimicrobial activity of carvacrol toward Bacillus cereus on rice. J. Food Prot. 2000, 63, 620–624. [Google Scholar] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Comp. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Ramos, C.; Teixeira, B.; Batista, I.; Matos, O.; Serrano, C.; Neng, N.R.; Nogueira, J.M.F.; Nunes, M.L.; Marques, A. Antioxidant and antibacterial activity of essential oil and extracts of bay laurel Laurus nobilis Linnaeus (Lauraceae) from Portugal. Nat. Prod. Res. 2011, 1–12. [Google Scholar]
- Schwarz, K.; Bertelsen, G.; Nissen, L.R.; Gardner, P.T.; Heinonen, M.I.; Hopia, A.; Huynh-Ba, T.; Lambelet, P.; McPhail, D.; Skibsted, L.H.; et al. Investigation of plant extracts for the protection of processed foods against lipid oxidation. Comparison of antioxidant assays based on radical scavenging, lipid oxidation and analysis of the principal antioxidant compounds. Eur. Food Res. Technol. 2001, 212, 319–328. [Google Scholar] [CrossRef]
- Klančnik, A.; Piskernik, S.; Jeršek, B.; Možina, S.S. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J. Microbiol. Methods 2010, 81, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Selection of yeasts as starter cultures for table olives: A step-by-step procedure. Front. Microbiol. 2012, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.E.V.; Stamford, T.L.M.; Neto, N.J.G.; de Souza, E.L. Inhibition of Staphylococcus aureus in broth and meat broth using synergies of phenolics and organic acids. Int. J. Food Microbiol. 2010, 137, 312–316. [Google Scholar] [PubMed]
- Mackay, M.L.; Milne, K.; Gould, I.M. Comparison of methods for assessing synergic antibiotic interactions. Int. J. Antimicrob. Agents 2000, 15, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Nisha, S.A.; Sakthivel, R.; Pandian, S.K. Eugenol (an essential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane. J. Ethnopharmacol. 2010, 130, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zengin, H.; Baysal, A.H. Antibacterial and Antioxidant Activity of Essential Oil Terpenes against Pathogenic and Spoilage-Forming Bacteria and Cell Structure-Activity Relationships Evaluated by SEM Microscopy. Molecules 2014, 19, 17773-17798. https://doi.org/10.3390/molecules191117773
Zengin H, Baysal AH. Antibacterial and Antioxidant Activity of Essential Oil Terpenes against Pathogenic and Spoilage-Forming Bacteria and Cell Structure-Activity Relationships Evaluated by SEM Microscopy. Molecules. 2014; 19(11):17773-17798. https://doi.org/10.3390/molecules191117773
Chicago/Turabian StyleZengin, Hatice, and Ayse H. Baysal. 2014. "Antibacterial and Antioxidant Activity of Essential Oil Terpenes against Pathogenic and Spoilage-Forming Bacteria and Cell Structure-Activity Relationships Evaluated by SEM Microscopy" Molecules 19, no. 11: 17773-17798. https://doi.org/10.3390/molecules191117773
APA StyleZengin, H., & Baysal, A. H. (2014). Antibacterial and Antioxidant Activity of Essential Oil Terpenes against Pathogenic and Spoilage-Forming Bacteria and Cell Structure-Activity Relationships Evaluated by SEM Microscopy. Molecules, 19(11), 17773-17798. https://doi.org/10.3390/molecules191117773