Changes in Nutritional Metabolites of Young Ginger (Zingiber officinale Roscoe) in Response to Elevated Carbon Dioxide


Abstract
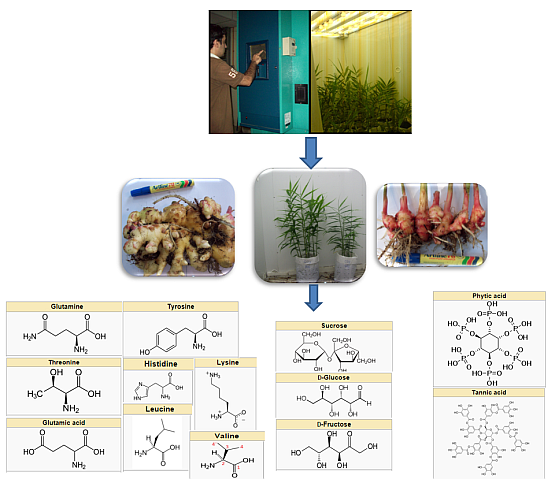
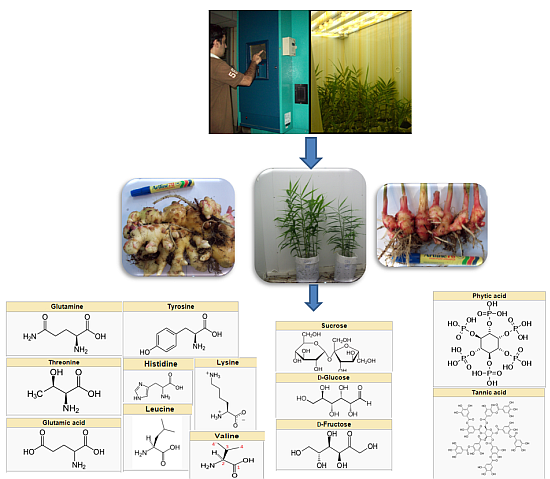
:
1. Introduction
2. Results and Discussion
2.1. Effect of Ambient and Elevated CO2 on Total Carbohydrate and Protein Content

{kind=link}
| Variety | Part | Total Carbohydrate | Total Protein | ||
|---|---|---|---|---|---|
| 400 | 800 | 400 | 800 | ||
| H. Bentong | Leaf | 142.80 ± 12.44 e | 201.50 ± 11.62 c | 16.30 ± 1.51 d | 11.80 ± 1.46 e |
| Rhizome | 176.50 ± 11.85 d | 288.40 ± 13.13 a | 21.70 ± 1.56 b | 16.70 ± 1.75 d | |
| H. Bara | Leaf | 140.50 ± 11.63 e | 189.60 ± 11.92 c | 16.80 ± 1.33 d | 12.40 ± 1.92 e |
| Rhizome | 168.20 ± 11.41 d | 244.70 ± 11.81 b | 27.50 ± 1.87 a | 18.80 ± 1.38 c | |
2.2. Effect of Ambient and Elevated CO2 on Sugars (Sucrose, Glucose and Fructose) Content
| Variety | Part | Sucrose | Glucose | Fructose | |||
|---|---|---|---|---|---|---|---|
| 400 | 800 | 400 | 800 | 400 | 800 | ||
| H. Bentong | Leaf | 25.00 ± 1.84 d | 38.20 ± 1.42 b | 10.60 ± 1.77 f | 27.20 ± 2.44 c | 7.40 ± 0.94 e | 15.60 ± 1.62 c |
| Rhizome | 33.50 ± 1.19 c | 44.60 ± 2.31 a | 17.40 ± 1.38 e | 31.50 ± 1.82 b | 8.60 ± 1.33 d | 21.30 ± 2.71 a | |
| H. Bara | Leaf | 20.60 ± 1.65 e | 37.50 ± 1.78 b | 6.30 ± 0.95 g | 22.60 ± 1.69 d | 5.90 ± 1.74 f | 19.40 ± 1.18 b |
| Rhizome | 27.60 ± 1.22 d | 41.80 ± 1.49 a | 18.60 ± 1.29 e | 36.20 ± 1.28 a | 9.50 ± 2.52 d | 22.10 ± 2.66 a | |
2.3. Effect of Ambient and Elevated CO2 on Amino Acids Content in Ginger
2.4. Effect of Ambient and Elevated CO2 on Antinutrient Content
| H. Bentong | H. Bara | |||||||
|---|---|---|---|---|---|---|---|---|
| Leaf | Rhizome | Leaf | Rhizome | |||||
| 400 | 800 | 400 | 800 | 400 | 800 | 400 | 800 | |
| Glutamine | 41.30 ± 1.17 a | 36.10 ± 2.14 ab | 20.10 ± 1.53 c | 37.00 ± 1.27 ab | 22.40 ± 2.18 c | 33.20 ± 1.18 b | 14.80 ± 0.88 d | 31.70 ± 2.06 b |
| Histidine | 20.10 ± 1.44 b | 27.50 ± 1.48 a | ND | 12.50 ± 1.14 d | 12.50 ± 1.38 d | 15.30 ± 1.22 c | ND | 6.30 ± 0.36 e |
| Glutamic acid | 28.10 ± 1.73 d | 37.20 ± 1.73 c | 28.80 ± 1.03 d | 53.10 ± 2.16 a | 16.40 ± 0.86 f | 27.50 ± 1.32 d | 22.50 ± 1.16 e | 43.70 ± 1.44 b |
| Threonine | 16.40 ± 1.93 f | 18.20 ± 1.66 f | 22.40 ± 1.55 e | 43.50 ± 1.28 b | 21.80 ± 1.22 e | 28.40 ± 1.25 d | 34.20 ± 2.18 c | 64.80 ± 2.83 a |
| Leucine | 37.50 ± 2.3 d | 28.40 ± 1.15 e | 22.10 ± 1.83 f | 40.70 ± 1.66 c | 42.70 ± 1.72 c | 54.70 ± 2.17 b | 41.50 ± 2.16 c | 88.90 ± 3.29 a |
| Lysine | ND | 4.20 ± 0.87 d | 2.80 ± 0.38 d | 5.50 ± 0.79 d | 13.60 ± 0.69 c | 20.50 ± 1.18 b | 14.40 ± 1.06 c | 29.40 ± 2.04 a |
| Valine | 29.40 ± 1.29 c | 41.10 ± 2.31 b | 17.20 ± 1.55 d | 31.70 ± 1.27 c | 32.50 ± 1.62 c | 51.60 ± 2.77 a | 19.40 ± 1.16 d | 41.60 ± 1.26 b |
| Tyrosine | 12.40 ± 1.17 d | 19.40 ± 1.66 c | 10.20 ± 1.73 d | 21.90 ± 1.06 c | 18.30 ± 1.17 c | 33.70 ± 2.19 b | 18.20 ± 1.29 c | 40.80 ± 1.18 a |
| Total | 185.20 ± 1.88 d | 212.10 ± 3.52 c | 123.60 ± 3.52 e | 245.90 ± 1.44 b | 209.90 ± 2.88 c | 235.20 ± 3.40 b | 165.00 ± 2.77 d | 347.10 ± 3.28 a |
| H. Bentong | H. Bara | |||||||
|---|---|---|---|---|---|---|---|---|
| Leaf | Rhizome | Leaf | Rhizome | |||||
| 400 | 800 | 400 | 800 | 400 | 800 | 400 | 800 | |
| Cyanide | 117.5 ± 2.6 e | 223.1 ± 4.7 b | 87.4 ± 1.4 f | 102.7 ± 1.5 e | 188.2 ± 3.9 c | 336.1 ± 2.7 a | 92.4 ± 3.7 f | 129.3 ± 2.4 d |
| Tannin | ND | ND | ND | 22.5 ± 2.2 b | ND | 19.2 ± 1.1 c | ND | 27.5 ± 1.9 a |
| Phytic acid | 21.4 ± 1.3 d | 17.1 ± 3.9 d | 44.2 ± 2.6 b | 46.8 ± 2.8 b | 36.2 ± 1.7 c | 33.4 ± 1.2 c | 52.9 ± 3.2 a | 54.1 ± 2.3 a |
3. Experimental Section
3.1. Plant Materials
3.2. Growth Chamber Microclimate
3.3. Total Carbohydrate Content
3.4. Total Protein Content
3.5. High Performance Liquid Chromatography (HPLC) of Sugars
3.5.1. Extraction
3.5.2. HPLC Method
3.6. Ultra High Performance Liquid Chromatography Analysis of Amino Acids
3.6.1. Extraction
3.6.2. UHPLC Method
3.7. Antinutrient Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Onoda, Y.; Hirose, T.; Hikosaka, K. Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three natural CO2 springs in japan. Ecol. Res. 2007, 22, 475–484. [Google Scholar]
- Wang, S.Y.; Bunce, J.A.; Maas, J. Elevated carbon dioxide increases contents of antioxidant compounds in field-grown strawberries. J. Agric. Food Chem. 2003, 51, 4315–4320. [Google Scholar]
- Iason, G.R.; Dicke, M.; Hartley, S.E. The Ecology of Plant Secondary Metabolites: From Genes to Global Processes; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Luo, Z.-B.; Calfapietra, C.; Scarascia-Mugnozza, G.; Liberloo, M.; Polle, A. Carbon-based secondary metabolites and internal nitrogen pools in populus nigra under free air CO2 enrichment (face) and nitrogen fertilisation. Plant Soil 2008, 304, 45–57. [Google Scholar]
- Mattson, W.J.; Julkunen Tiitto, R.; Herms, D.A. CO2 enrichment and carbon partitioning to phenolics: Do plant responses accord better with the protein competition or the growth differentiation balance models? Oikos 2005, 111, 337–347. [Google Scholar]
- Poorter, H.; Navas, M.L. Plant growth and competition at elevated CO2: On winners, losers and functional groups. New Phytol. 2003, 157, 175–198. [Google Scholar]
- Moore, B.; Cheng, S.H.; Sims, D.; Seemann, J. The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric co2. Plant Cell Environ. 1999, 22, 567–582. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z. Impact of elevated carbon dioxide on primary, secondary metabolites and antioxidant responses of eleais guineensis jacq.(oil palm) seedlings. Molecules 2012, 17, 5195–5211. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z. Enhancement of leaf gas exchange and primary metabolites under carbon dioxide enrichment up-regulates the production of secondary metabolites in labisia pumila seedlings. Molecules 2011, 16, 3761–3777. [Google Scholar]
- Jackson, R.; Luo, Y.; Cardon, Z.; Sala, O.; Field, C.; Mooney, H. Photosynthesis, growth and density for the dominant species in a CO2-enriched grassland. J. Biogeogr. 1995, 221–225. [Google Scholar]
- Cheng, W.; Sims, D.A.; Luo, Y.; Coleman, J.S.; Johnson, D.W. Photosynthesis, respiration, and net primary production of sunflower stands in ambient and elevated atmospheric CO2 concentrations: An invariant npp: Gpp ratio? Glo. Chang. Biol. 2000, 6, 931–941. [Google Scholar]
- Sims, D.A.; Cheng, W.; Luo, Y.; Seemann, J.R. Photosynthetic acclimation to elevated CO2 in a sunflower canopy. J. Exp. Bot. 1999, 50, 645–653. [Google Scholar]
- Pritchard, S.; Rogers, H.; Prior, S.A.; Peterson, C. Elevated CO2 and plant structure: A review. Glob. Chang. Biol. 1999, 5, 807–837. [Google Scholar]
- Ghasemzadeh, A.; Jaafar, H.Z. Antioxidant potential and anticancer activity of young ginger (zingiber officinale roscoe) grown under different co2 concentration. J. Med. Plants Res. 2011, 5, 3247–3255. [Google Scholar]
- Kerepesi, I.; Galiba, G. Osmotic and salt stress-induced alteration in soluble carbohydrate content in wheat seedlings. Crop Sci. 2000, 40, 482–487. [Google Scholar]
- Bowes, G. Growth at elevated CO2: Photosynthetic responses mediated through rubisco. Plant Cell Environ. 1991, 14, 795–806. [Google Scholar]
- Salvucci, M.E.; Crafts Brandner, S.J. Inhibition of photosynthesis by heat stress: The activation state of rubisco as a limiting factor in photosynthesis. Physiol. Plant. 2004, 120, 179–186. [Google Scholar]
- Ke, D.; Mateos, M.; Siriphanich, J.; Li, C.; Kader, A.A. Carbon dioxide action on metabolism of organic and amino acids in crisphead lettuce. Postharvest Biol. Technol. 1993, 3, 235–247. [Google Scholar]
- Sicher, R.C. Effects of CO2 enrichment on soluble amino acids and organic acids in barley primary leaves as a function of age, photoperiod and chlorosis. Plant Sci. 2008, 174, 576–582. [Google Scholar]
- Ghasemzadeh, A.; Jaafar, H.Z.; Karimi, E.; Ibrahim, M.H. Combined effect of CO2 enrichment and foliar application of salicylic acid on the production and antioxidant activities of anthocyanin, flavonoids and isoflavonoids from ginger. BMC Complement. Altern. Med. 2012, 12, 229. [Google Scholar]
- Den-Hertog, J.; Stulen, L.; Fonseca, E.; Delea, P. Modulation of carbon and nitrogen allocation in urtica diocia and plantago major by elevated CO2: Impact of accumulation of non-structural carbohydrates and ontogenic drift. Physiol. Plant. 1996, 98, 77–88. [Google Scholar]
- Baxter, R.; Ashenden, T.W.; Farrar, J. Effects of elevated co2 and nutrient status on growth, dry matter partitioning and nutrient content of poa alpinia var. Vivpara l. J. Exp. Bot. 1997, 48, 1477–1486. [Google Scholar]
- Landolt, W.; Pfenninger, I. The effect of elevated co2 and soil type on non-structural carbohydrates in beech leaves and norway spruce needles growing in model ecosystems. Acta Oecol. 1997, 18, 351–359. [Google Scholar]
- Taub, D.R.; Miller, B.; Allen, H. Effects of elevated co2 on the protein concentration of food crops: A meta analysis. Glob. Chang. Biol. 2008, 14, 565–575. [Google Scholar]
- Hogy, P.; Fangmeier, A. Atmospheric CO2 enrichment affects potatoes: 2. Tuber quality traits. Eur. J. Agric. 2009, 30, 85–94. [Google Scholar]
- Terao, T.; Miura, S.; Yanagihara, T.; Hirose, T.; Nagata, K.; Tabuchi, H.; Kim, H.Y.; Lieffering, M.; Okada, M.; Kobayashi, K. Influence of free air CO2 enrichment (face) on the eating quality of rice. J. Sci. Food Agric. 2005, 85, 1861–1868. [Google Scholar]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar]
- Jones, C.; Hartley, S. A protein competition model of phenolic allocation. Oikos 1999, 86, 17. [Google Scholar]
- Ehleringer, J.R.; Cerling, T.E.; Dearing, M.D. Atmospheric CO2 as a global change driver influencing plant-animal interactions. Integr. Comp. Biol. 2002, 42, 424–430. [Google Scholar]
- Behboudian, M.H.; Tod, C. Postharvest attributes of virosa tomato fruit produced in an enriched carbon dioxide environment. Hort. Sci. 1995, 30, 490–491. [Google Scholar]
- Cha-um, S.; Ulziibat, B.; Kirdmanee, C. Effects of temperature and relative humidity during in vitro acclimatization, on physiological changes and growth characters of phalaenopsis adapted to in vivo. Aust. J. Crop Sci. 2011, 4, 750–756. [Google Scholar]
- De Souza, A.P.; Gaspar, M.; Da Silva, E.A.; Ulian, E.C.; Waclawovsky, A.J.; Dos santos, R.V.; Teixeira, M.M.; Souza, G.M.; Buckeridge, M.S. Elevated CO2 increases photosynthesis, biomass and productivity, and modifies gene expression in sugarcane. Plant Cell Environ. 2008, 31, 1116–1127. [Google Scholar]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400. [Google Scholar]
- Demmers-Derks, H.; Mitchell, R.; Mitchell, V.; Lawlor, D. Response of sugar beet (beta vulgaris l.) yield and biochemical composition to elevated co2 and temperature at two nitrogen applications. Plant Cell Environ. 1998, 21, 829–836. [Google Scholar]
- Norby, R.J.; O’Neill, E.G.; Luxmoore, R. Effects of atmospheric CO2 enrichment on the growth and mineral nutrition of quercus alba seedlings in nutrient-poor soil. Plant Physiol. 1986, 82, 83–89. [Google Scholar]
- Ainsworth, E.A.; Rogers, A.; Leakey, A.D.; Heady, L.E.; Gibon, Y.; Stitt, M.; Schurr, U. Does elevated atmospheric [CO2] alter diurnal c uptake and the balance of c and n metabolites in growing and fully expanded soybean leaves? J. Exp. Bot. 2007, 58, 579–591. [Google Scholar]
- Geiger, M.; Walch-Liu, P.; Engels, C.; Harnecker, J.; Schulze, E.; Ludewig, F.; Sonnewald, U.; Scheible, W.; Stitt, M. Enhanced carbon dioxide leads to a modified diurnal rhythm of nitrate reductase activity in older plants, and a large stimulation of nitrate reductase activity and higher levels of amino acids in young tobacco plants. Plant Cell Environ. 1998, 21, 253–268. [Google Scholar]
- Manderscheid, R.; Bender, J.; Jager, H.-J.; Weigel, H. Effects of season long co2 enrichment on cereals. Ii. Nutrient concentrations and grain quality. Agric. Ecosyst. Environ. 1995, 54, 175–185. [Google Scholar]
- Wu, G.; Chen, F.; Ge, F. Effects of elevated co2 on the growth and foliar chemistry of transgenic bt cotton. J. Integr. Plant Biol. 2007, 49, 1361–1369. [Google Scholar]
- Wrigley, C.; Bietz, J.; Pomeranz, Y. Proteins and amino acids. In Wheat: Chemistry and Technology; Pomeranz, Y., Ed.; American Association of Cereal Chemists: Saint Paul, Minnesota, USA, 1988; Volume 1, pp. 159–275. [Google Scholar]
- Kader, A.A. Biology and technology: An overview. Postharvest Technol. Hortic. Crops 2002, 3311, 39–48. [Google Scholar]
- Oksanen, E.; Sober, J.; Karnosky, D. Impacts of elevated CO2 and/or O3 on leaf ultrastructure of aspen (populus tremuloides) and birch (betula papyrifera) in the aspen face experiment. Environ. Pollut. 2001, 115, 437–446. [Google Scholar]
- Lindroth, R.L.; Kopper, B.J.; Parsons, W.F.; Bockheim, J.G.; Karnosky, D.F.; Hendrey, G.R.; Pregitzer, K.S.; Isebrands, J.; Sober, J. Consequences of elevated carbon dioxide and ozone for foliar chemical composition and dynamics in trembling aspen (populus tremuloides) and paper birch (betula papyrifera). Environ. Pollut. 2001, 115, 395–404. [Google Scholar]
- Vasic, M.A.; Tepic, A.N.; Mihailovic, V.M.; Mikic, A.M.; Gvozdanovic-Varga, J.M.; Sumic, Z.M.; Todorovic, V.J. Phytic acid content in different dry bean and faba bean landraces and cultivars. Seeds 2012, 29, 79–85. [Google Scholar]
- Sivaci, A. Seasonal changes of total carbohydrate contents in three varieties of apple (malus sylvestris miller) stem cuttings. Sci. Hortic. 2006, 109, 234–237. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Gomez, L.; Rubio, E.; Auge, M. A new procedure for extraction and measurement of soluble sugars in ligneous plants. J. Sci. Food. Agric. 2002, 82, 360–369. [Google Scholar]
- Wilson, A.; Work, T.; Bushway, A.; Bushway, R. Hplc determination of fructose, glucose, and sucrose in potatoes. J. Food Sci. 1981, 46, 300–301. [Google Scholar]
- Rezaul Haque, M.; Howard Bradbury, J. Total cyanide determination of plants and foods using the picrate and acid hydrolysis methods. Food Chem. 2002, 77, 107–114. [Google Scholar]
- Makkar, H.P.; Blummel, M.; Borowy, N.K.; Becker, K. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. J. Sci. Food. Agric. 1993, 61, 161–165. [Google Scholar]
- Vaintraub, I.A.; Lapteva, N.A. Colorimetric determination of phytate in unpurified extracts of seeds and the products of their processing. Anal. Biochem. 1988, 175, 227–230. [Google Scholar]
- Sample Availability: Samples of the Halia Bentong and Halia Bara are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghasemzadeh, A.; Jaafar, H.Z.E.; Karimi, E.; Ashkani, S. Changes in Nutritional Metabolites of Young Ginger (Zingiber officinale Roscoe) in Response to Elevated Carbon Dioxide. Molecules 2014, 19, 16693-16706. https://doi.org/10.3390/molecules191016693
Ghasemzadeh A, Jaafar HZE, Karimi E, Ashkani S. Changes in Nutritional Metabolites of Young Ginger (Zingiber officinale Roscoe) in Response to Elevated Carbon Dioxide. Molecules. 2014; 19(10):16693-16706. https://doi.org/10.3390/molecules191016693
Chicago/Turabian StyleGhasemzadeh, Ali, Hawa Z. E. Jaafar, Ehsan Karimi, and Sadegh Ashkani. 2014. "Changes in Nutritional Metabolites of Young Ginger (Zingiber officinale Roscoe) in Response to Elevated Carbon Dioxide" Molecules 19, no. 10: 16693-16706. https://doi.org/10.3390/molecules191016693
APA StyleGhasemzadeh, A., Jaafar, H. Z. E., Karimi, E., & Ashkani, S. (2014). Changes in Nutritional Metabolites of Young Ginger (Zingiber officinale Roscoe) in Response to Elevated Carbon Dioxide. Molecules, 19(10), 16693-16706. https://doi.org/10.3390/molecules191016693