Anticancer and Antifungal Compounds from Aspergillus, Penicillium and Other Filamentous Fungi

 ,
,
Abstract
:1. Introduction
2. Polyketide-Derived Anticancer Compounds
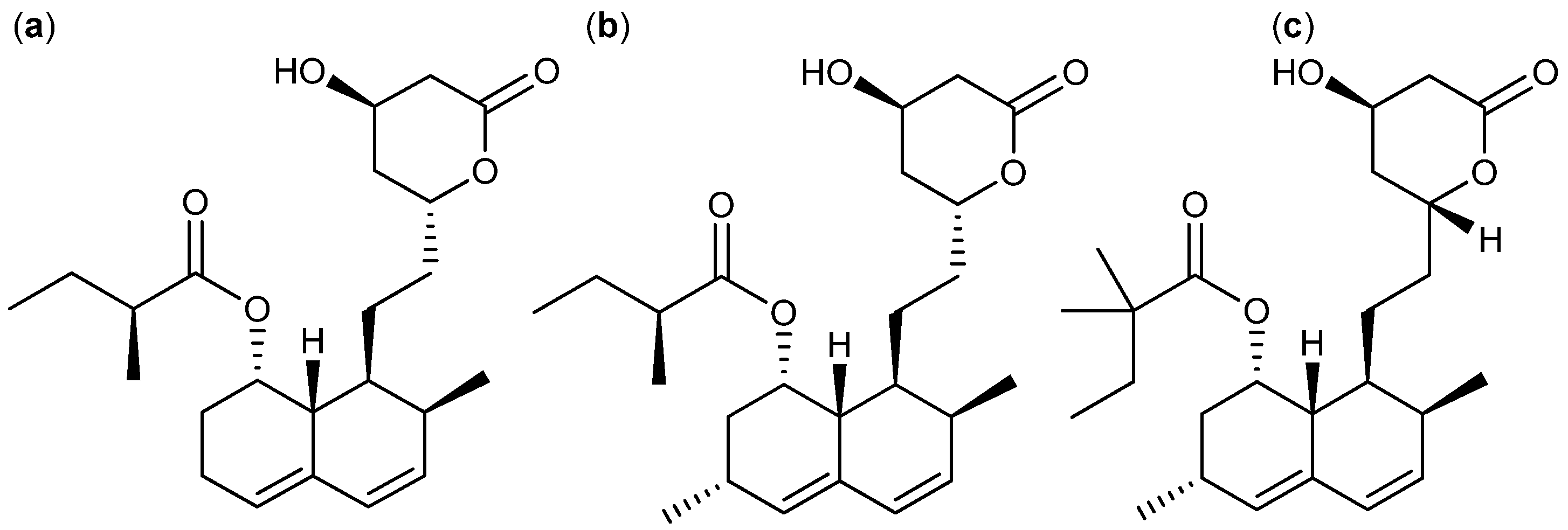
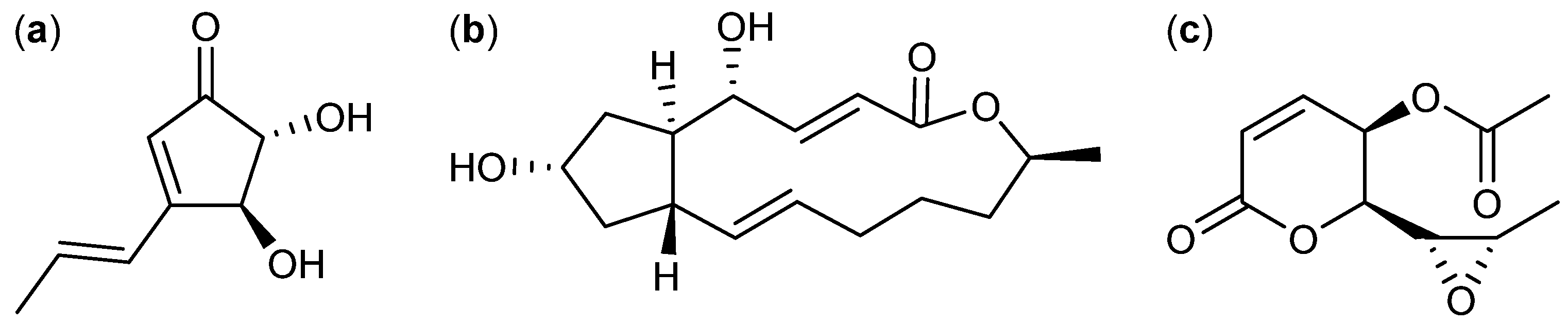
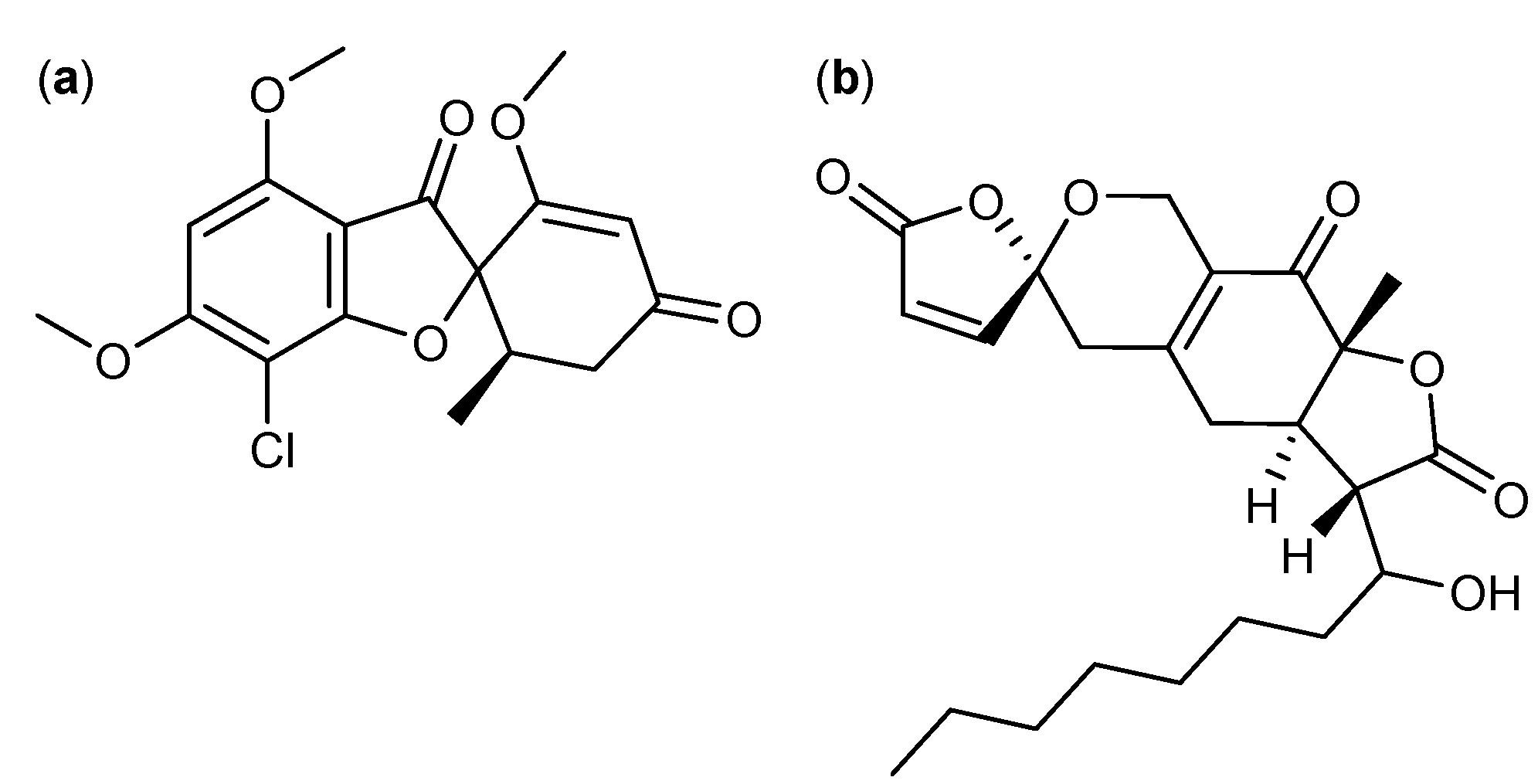
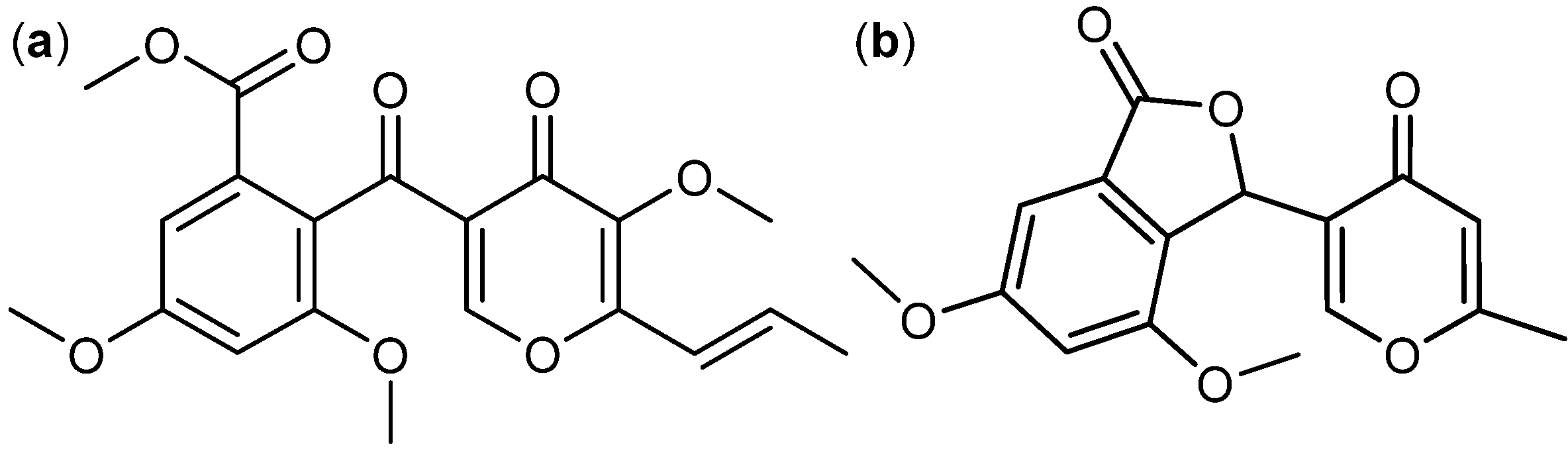
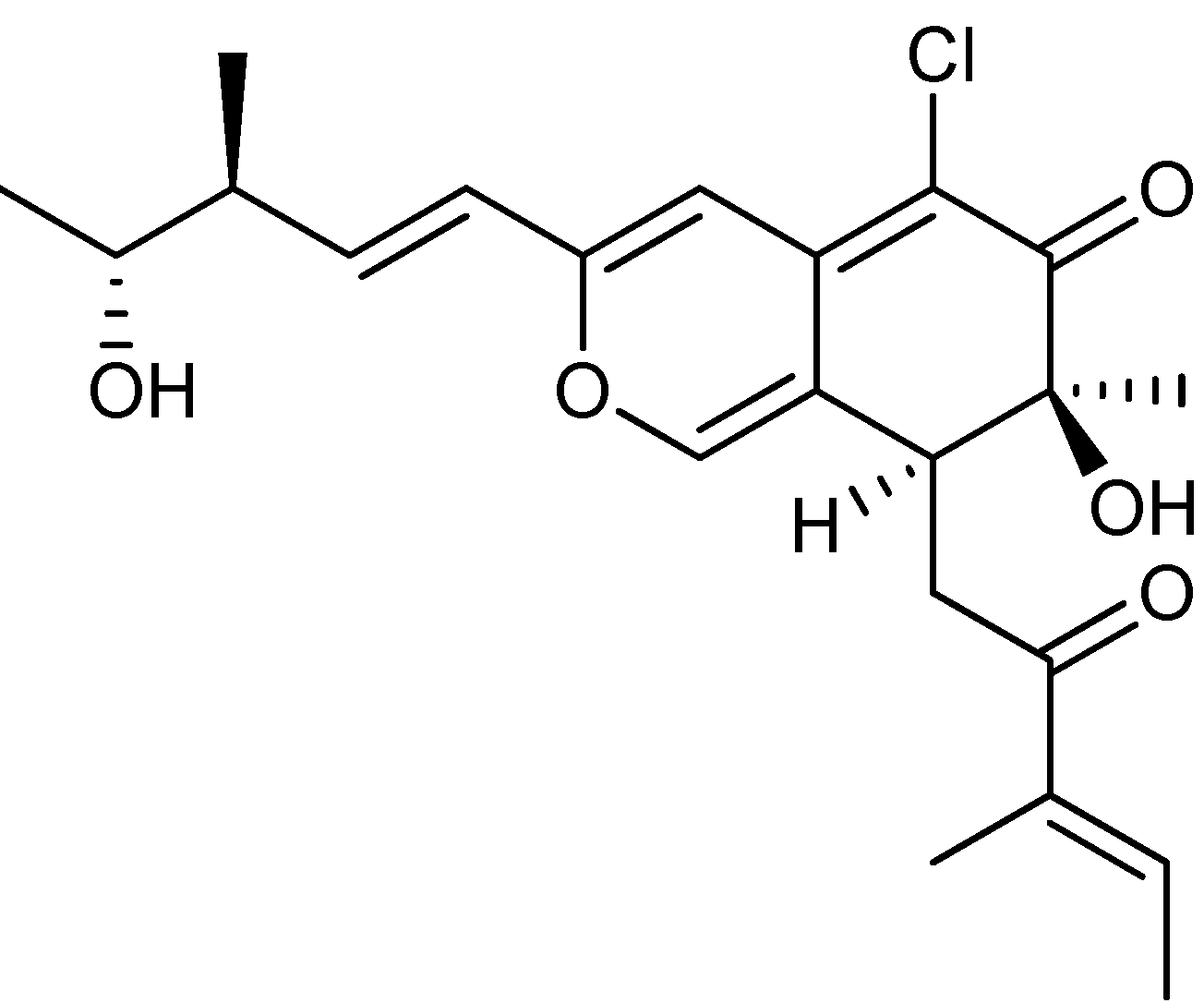
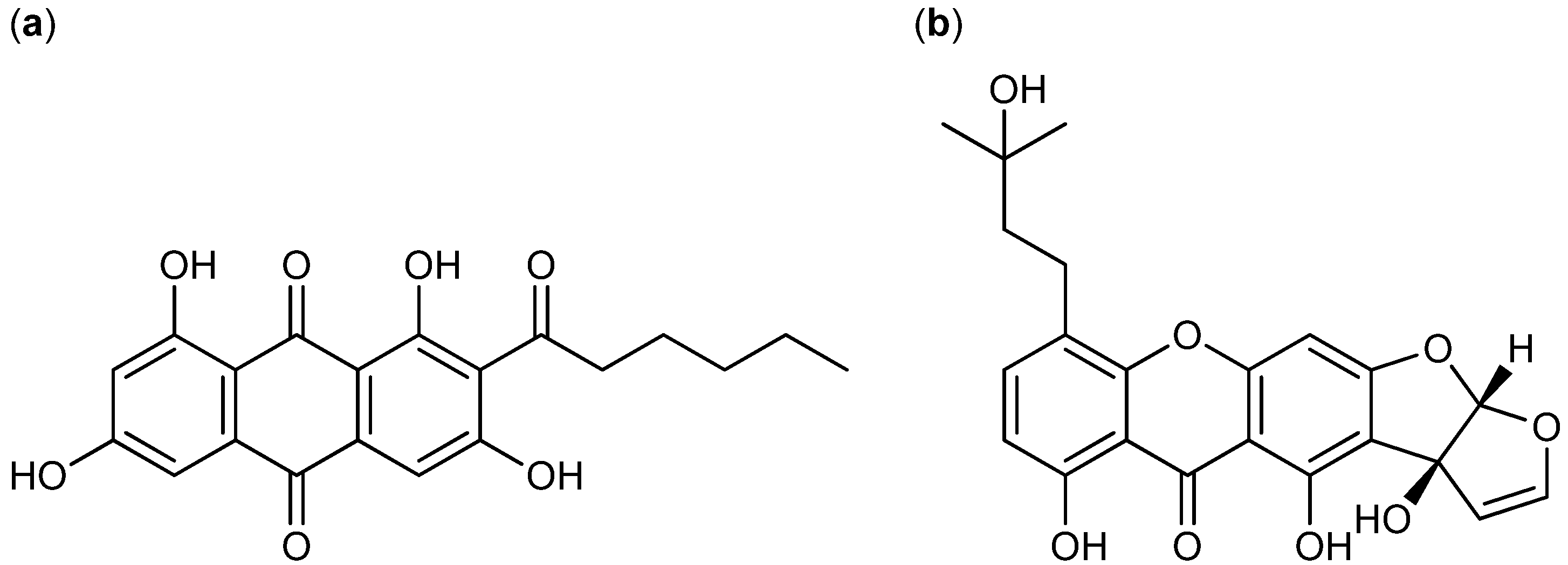
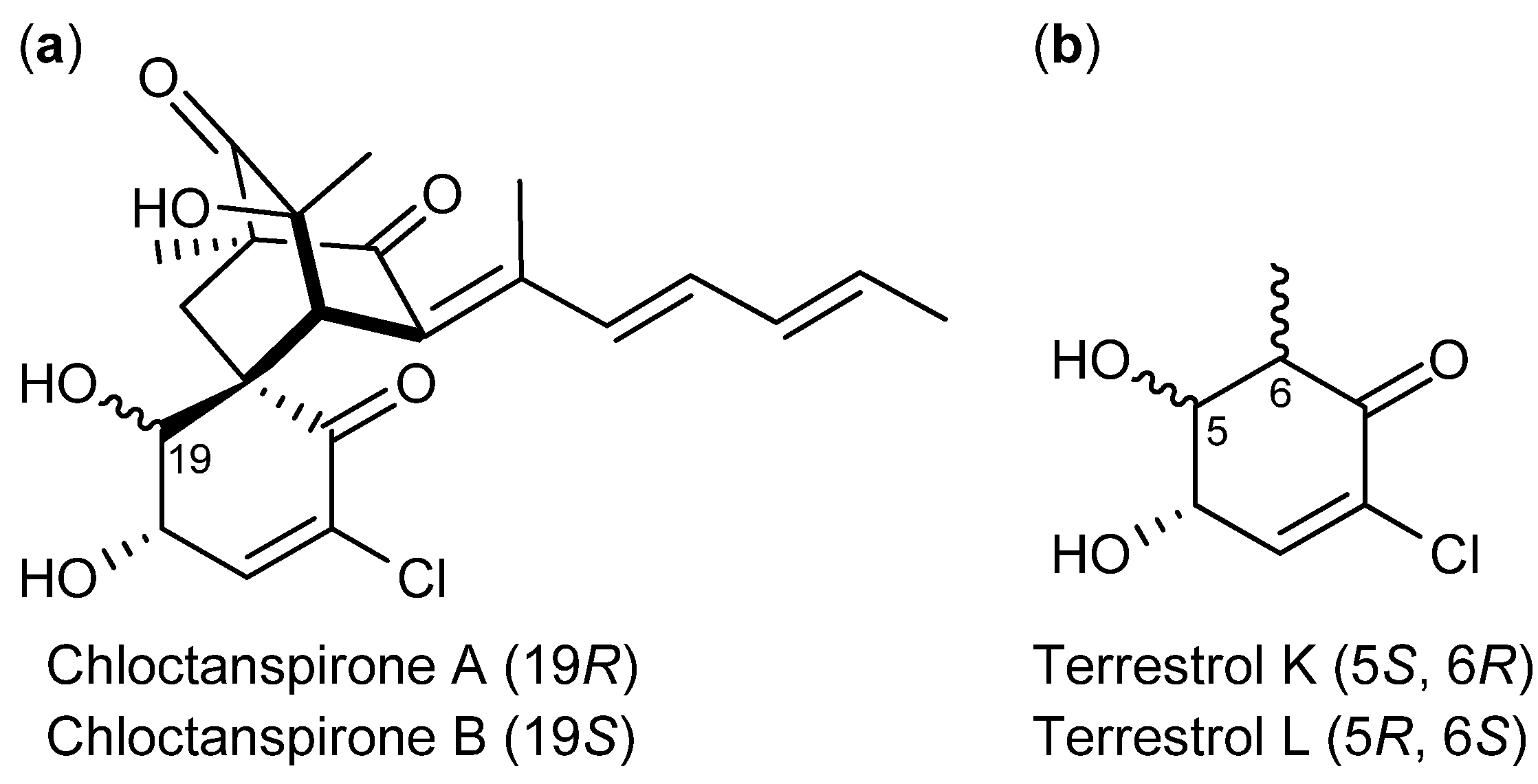
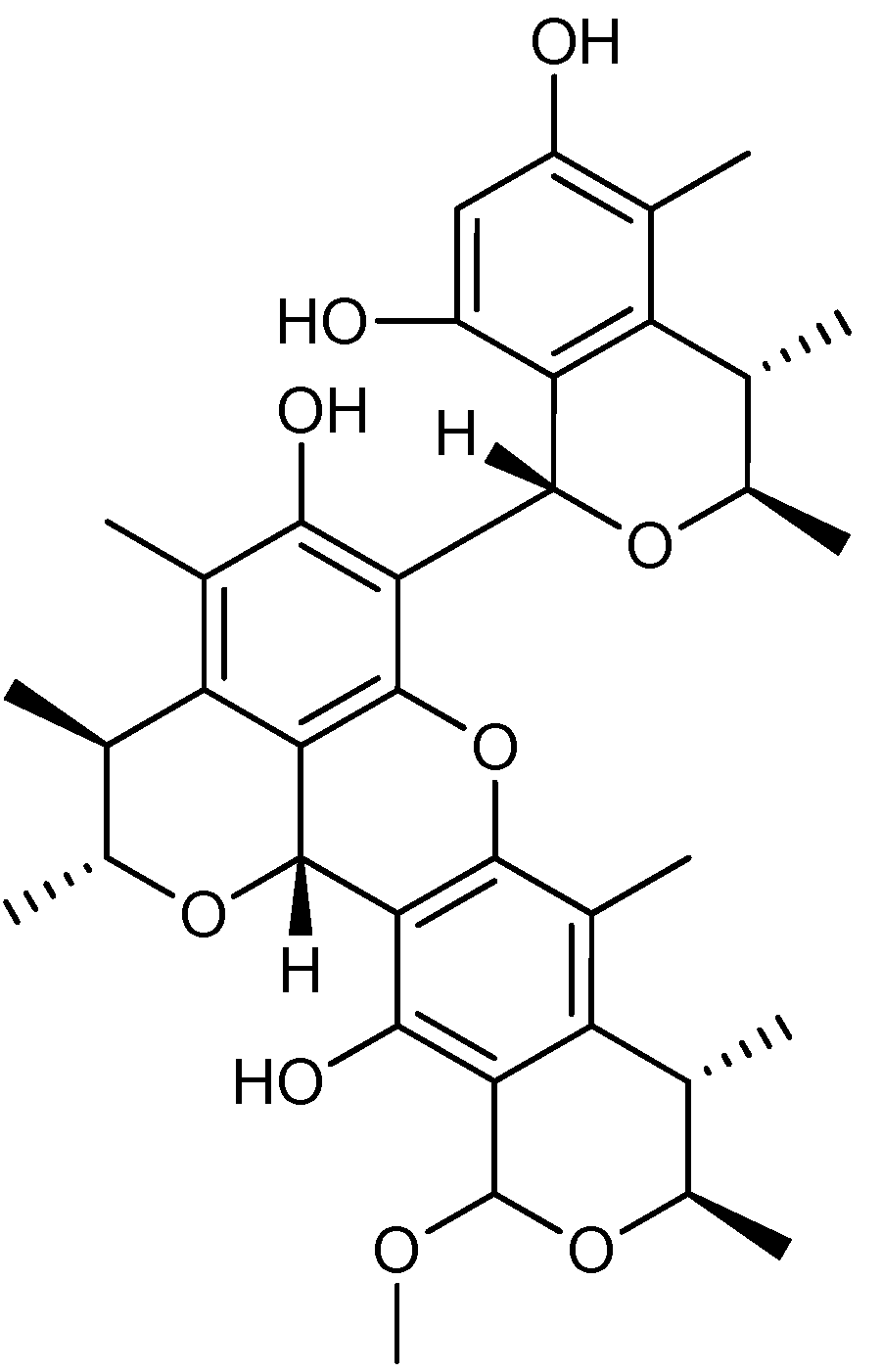
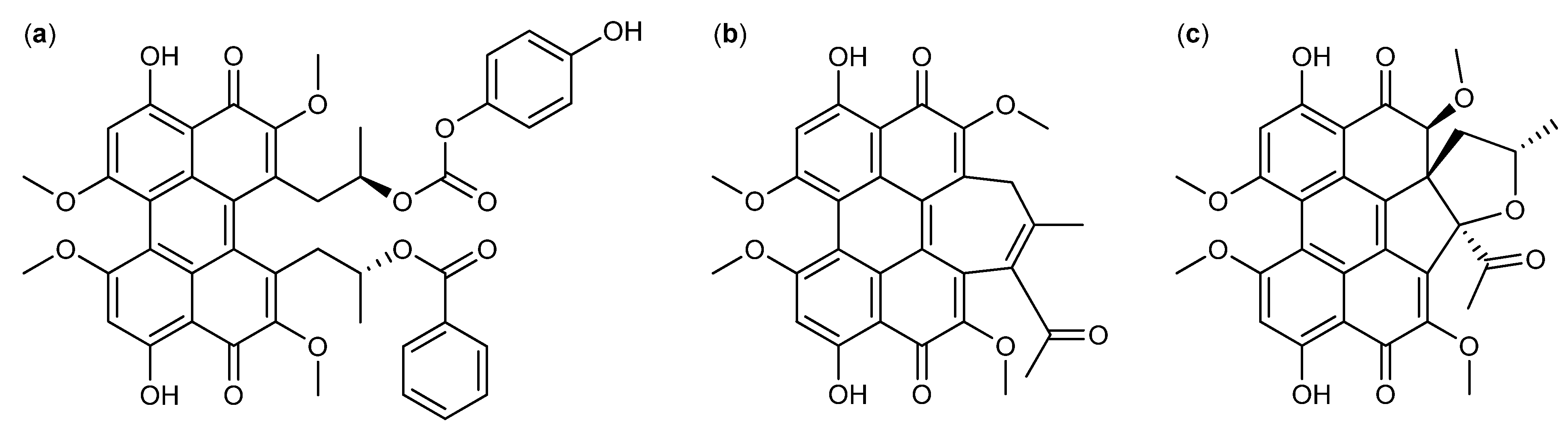
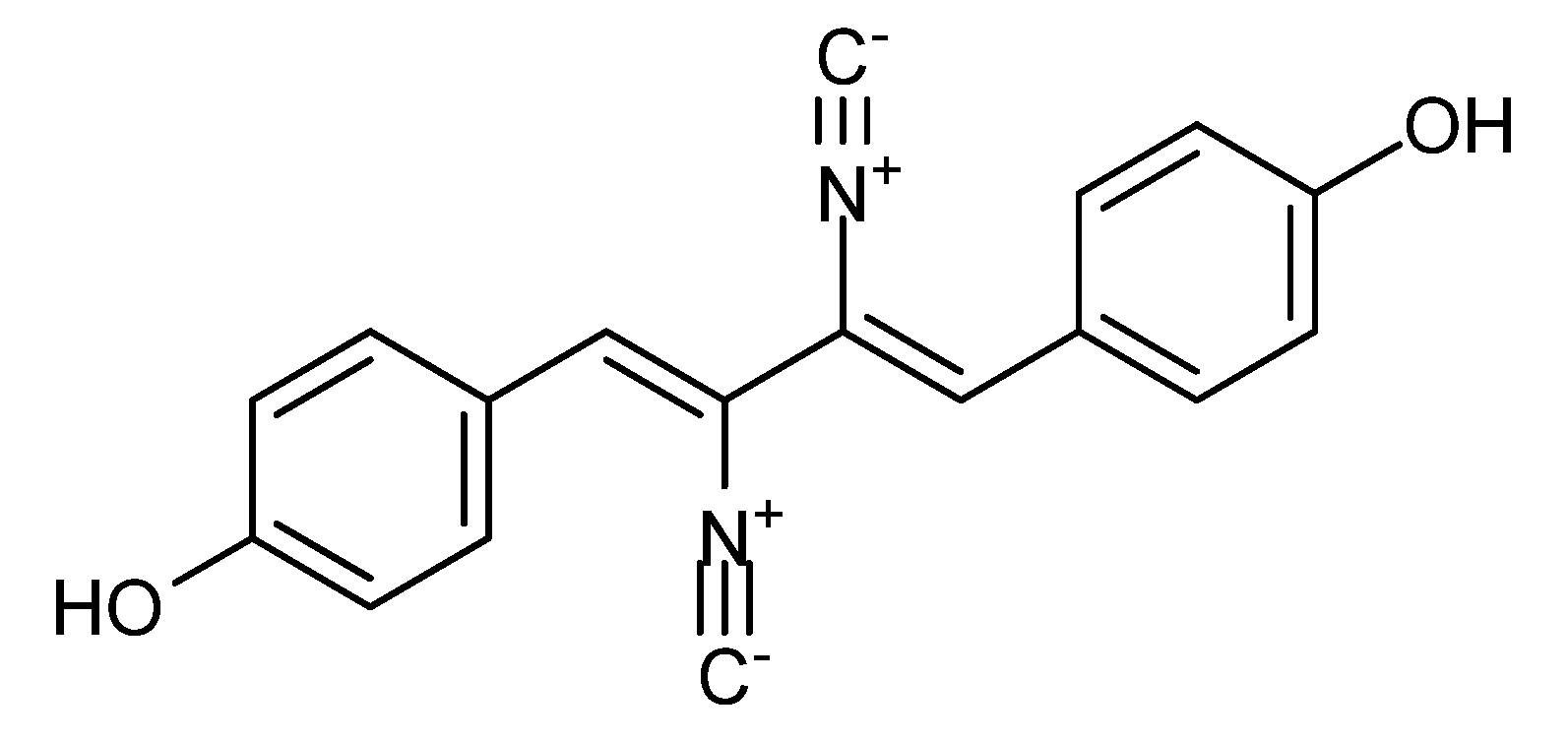
3. Nitrogen-Containing Anticancer Compounds
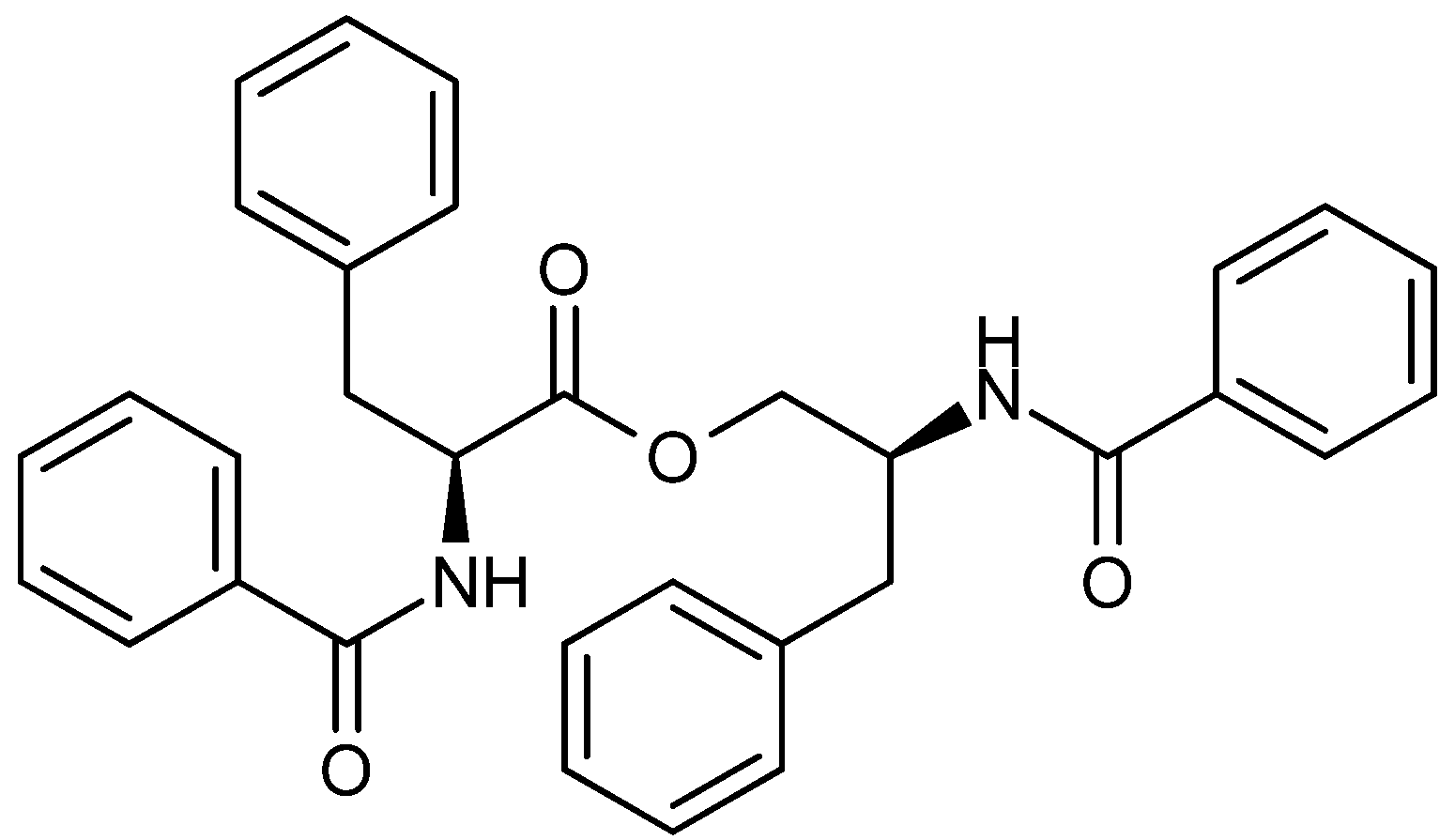
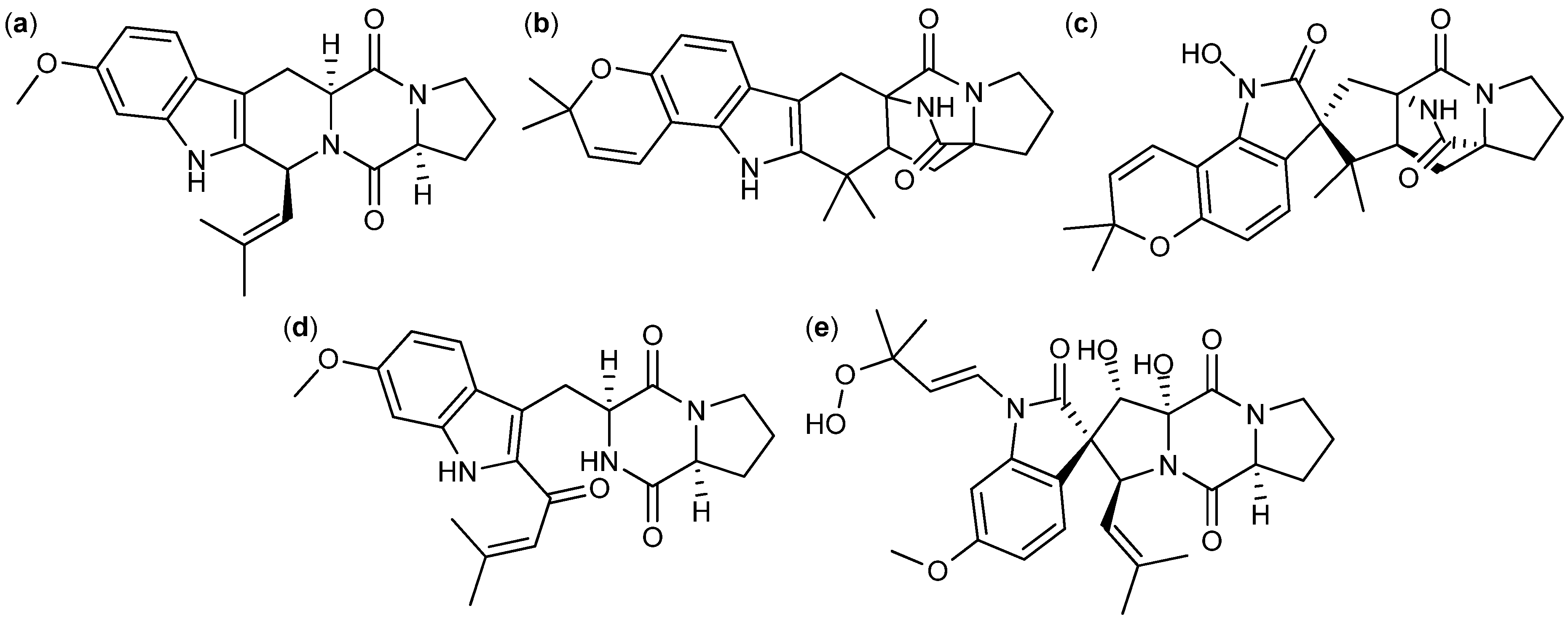
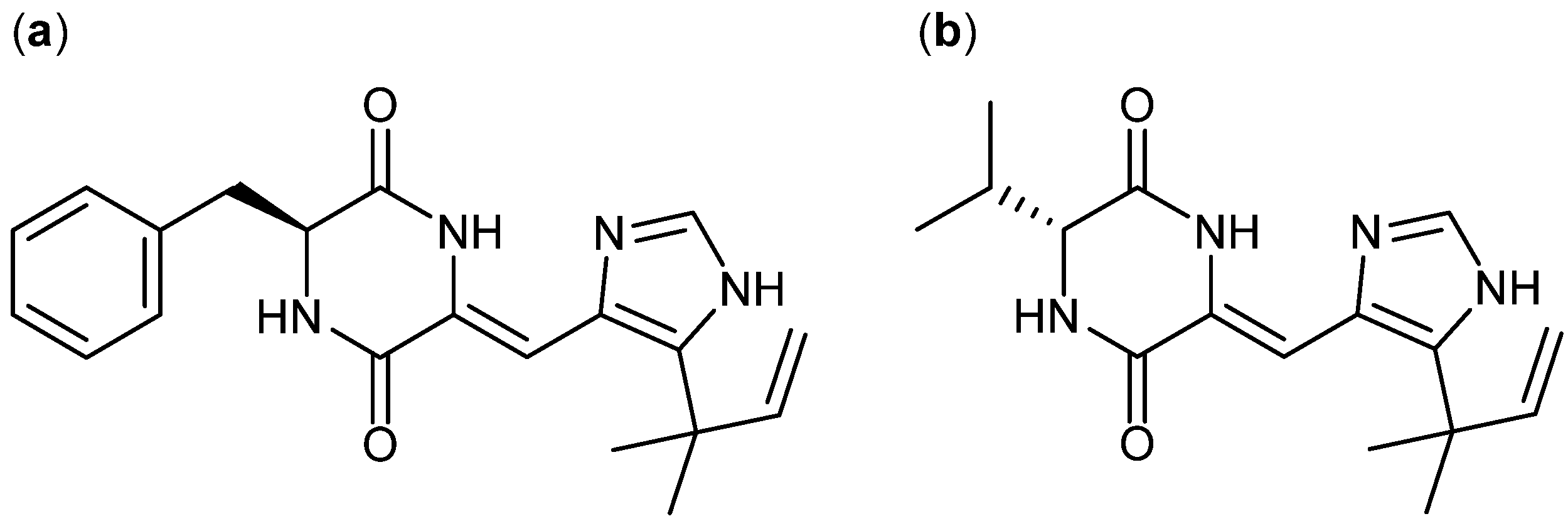
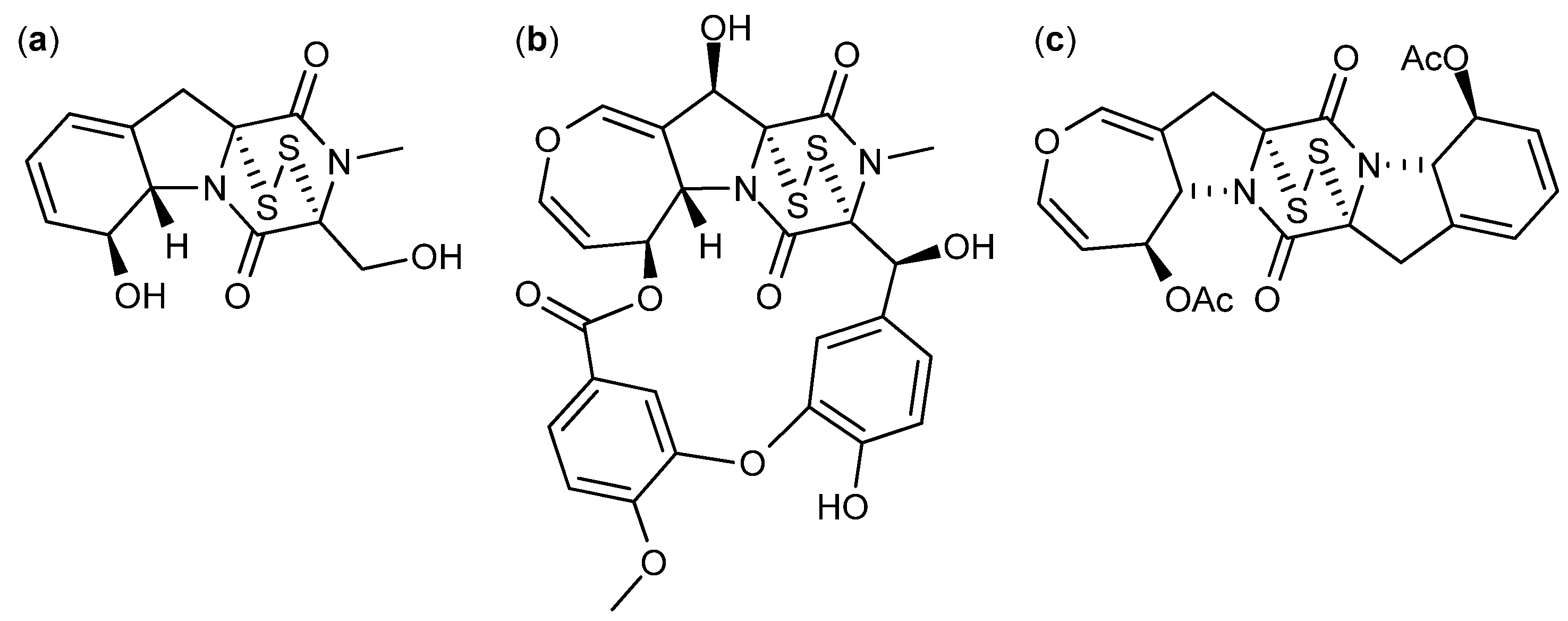
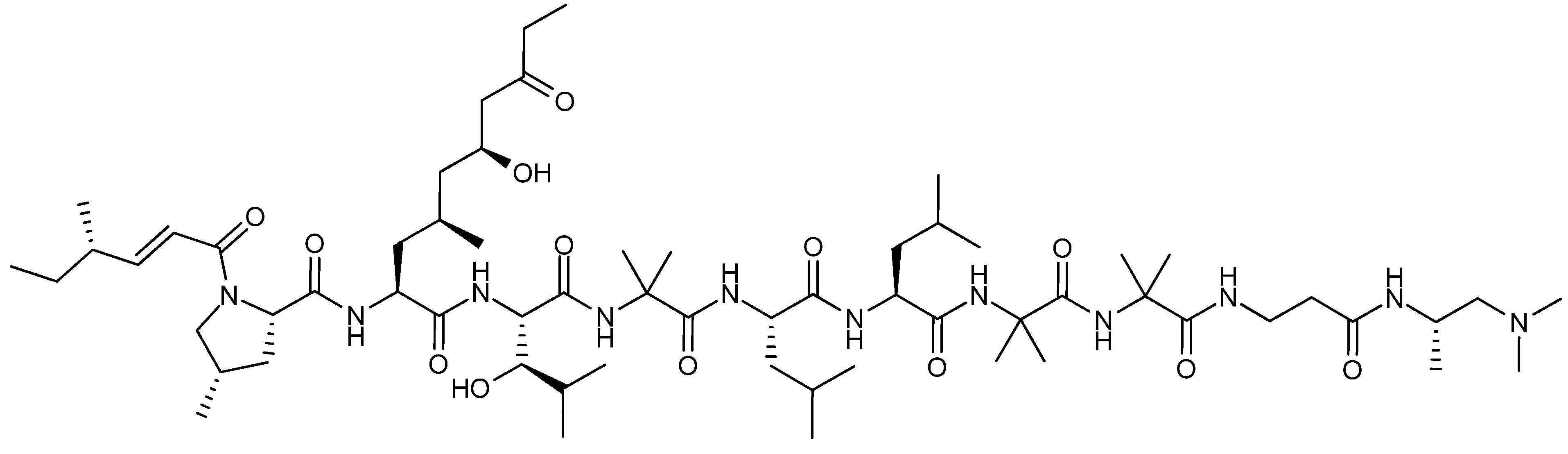
4. Terpenoid-Derived Anticancer Compounds
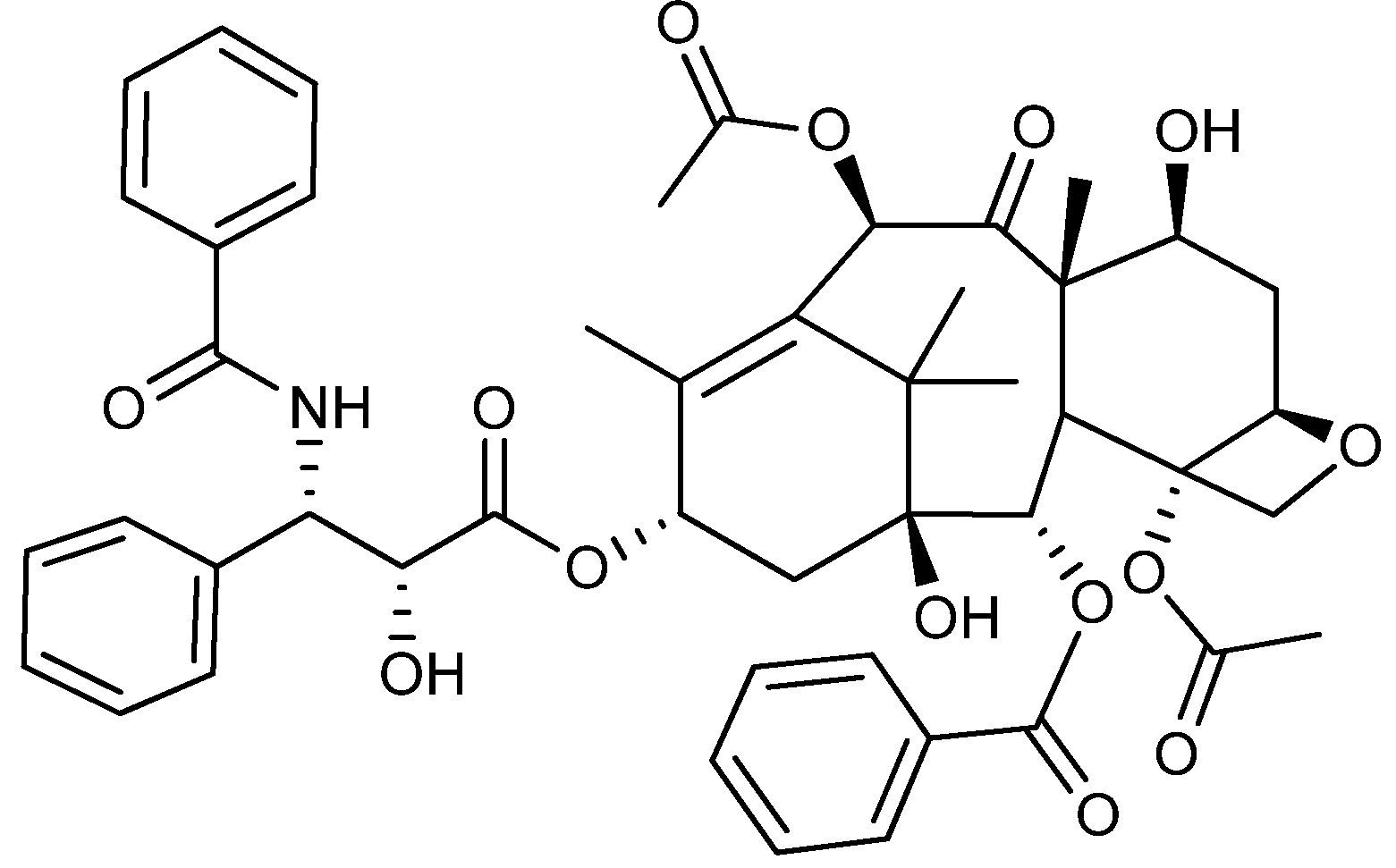

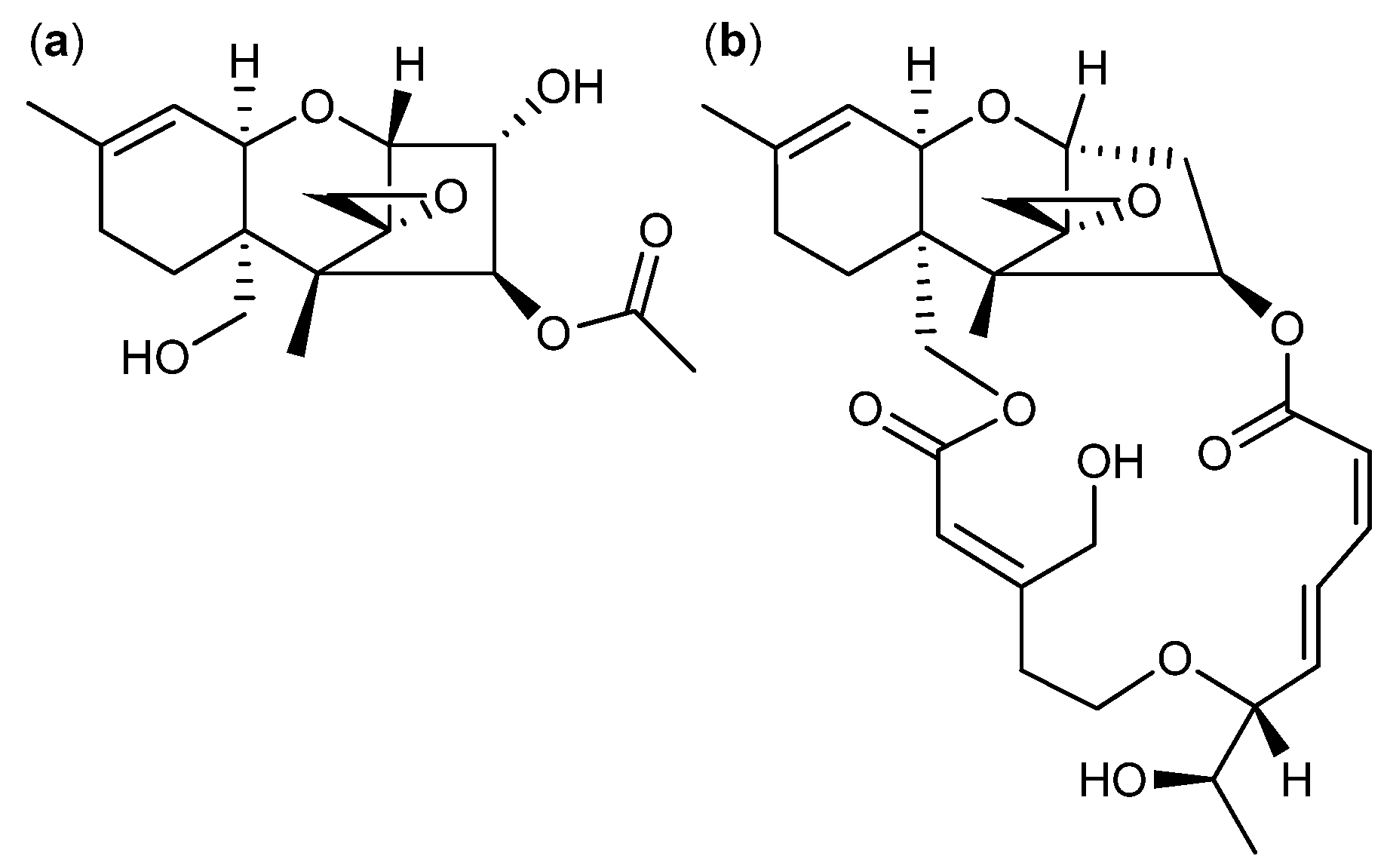

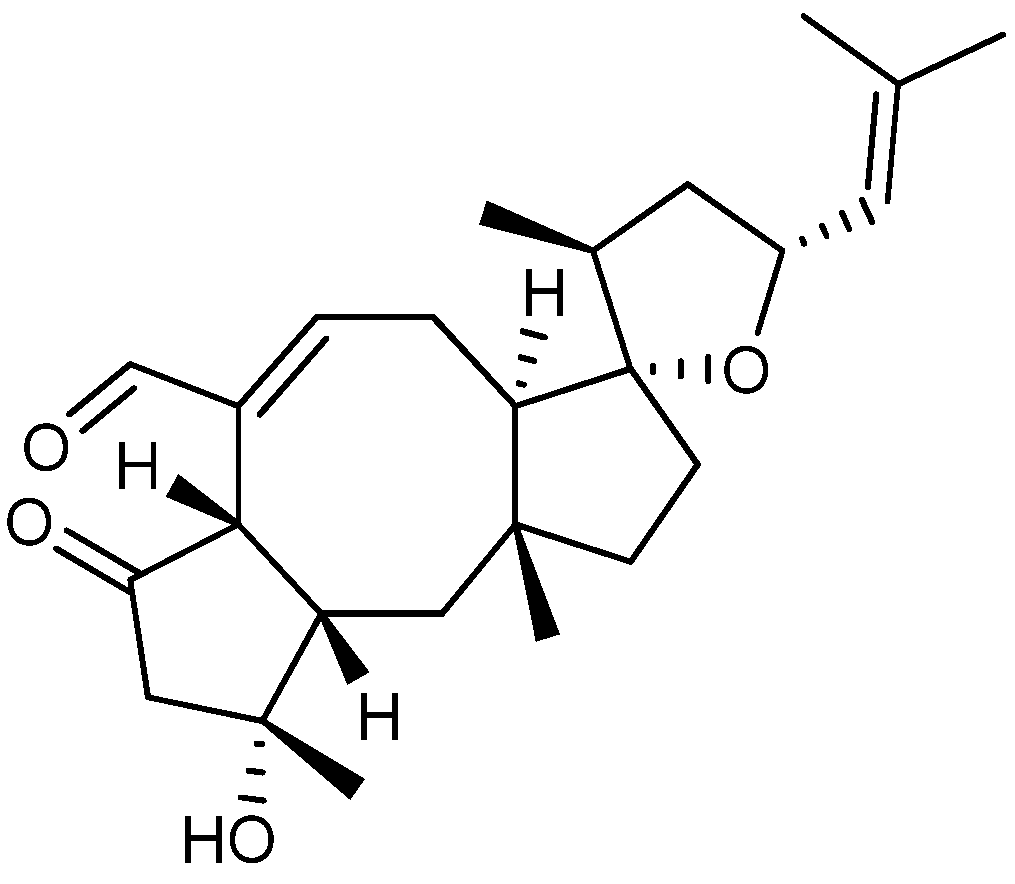
5. Anticancer Natural Products of Mixed or Unresolved Biosynthetic Origin

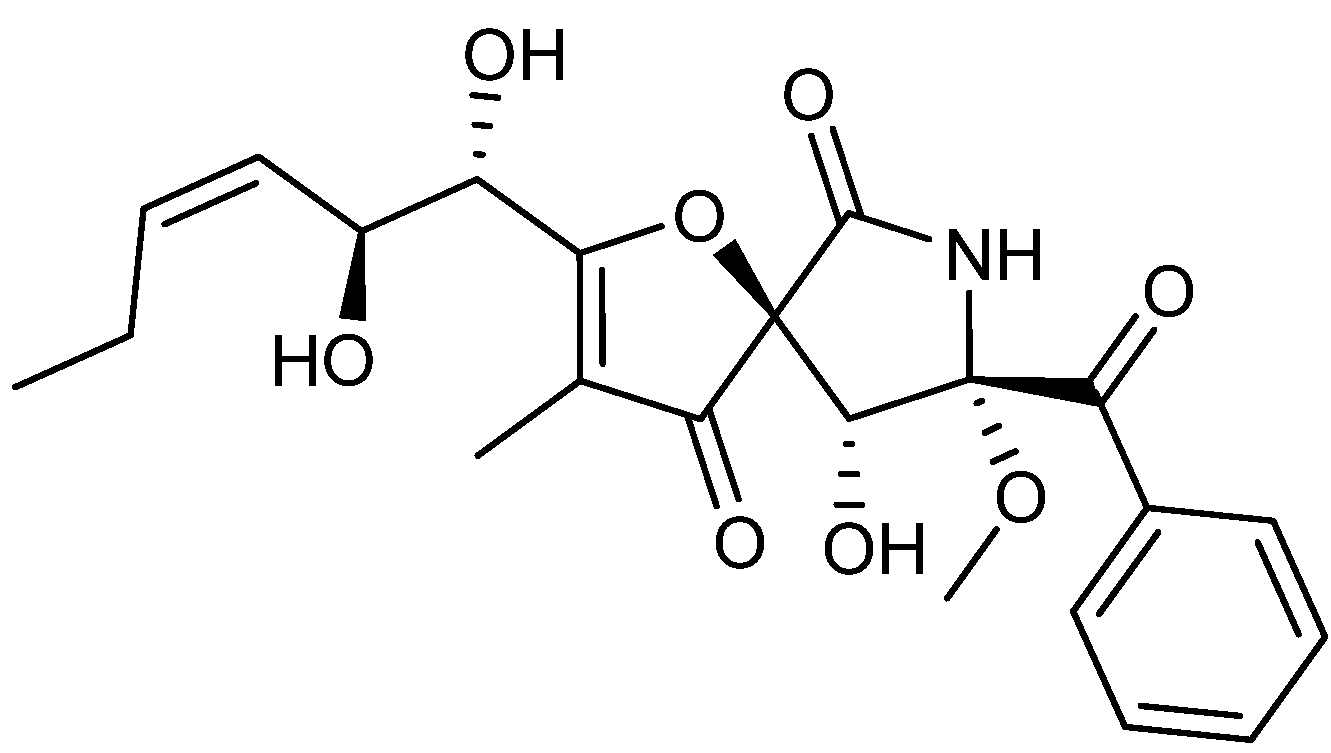
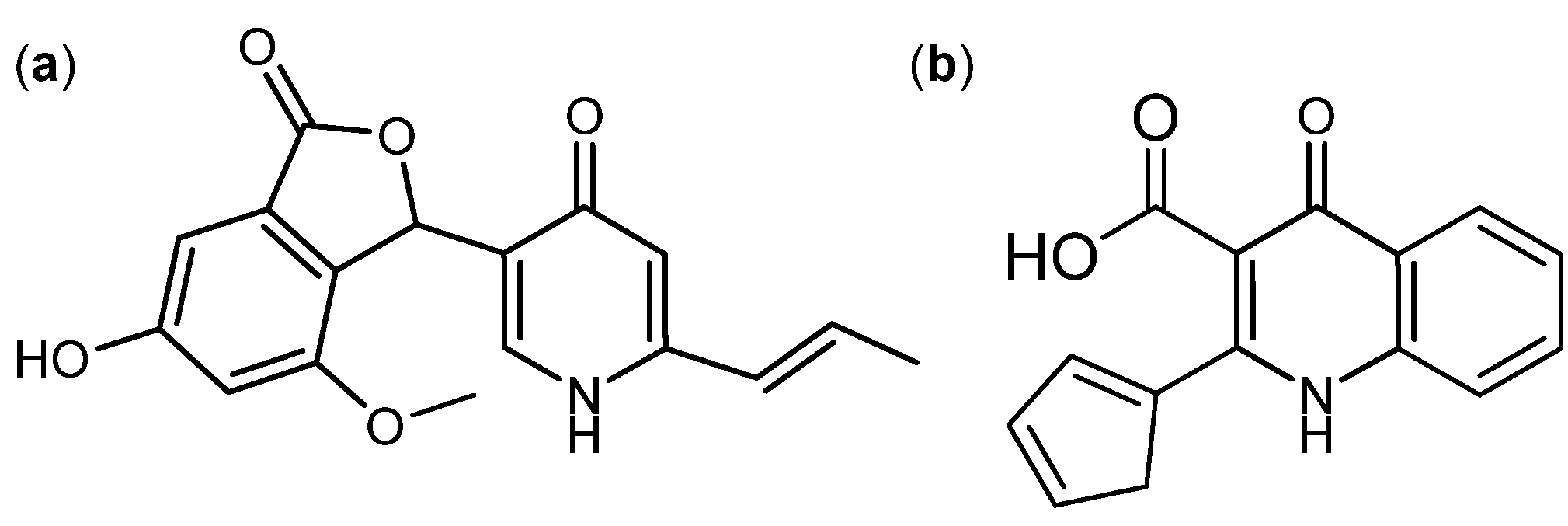
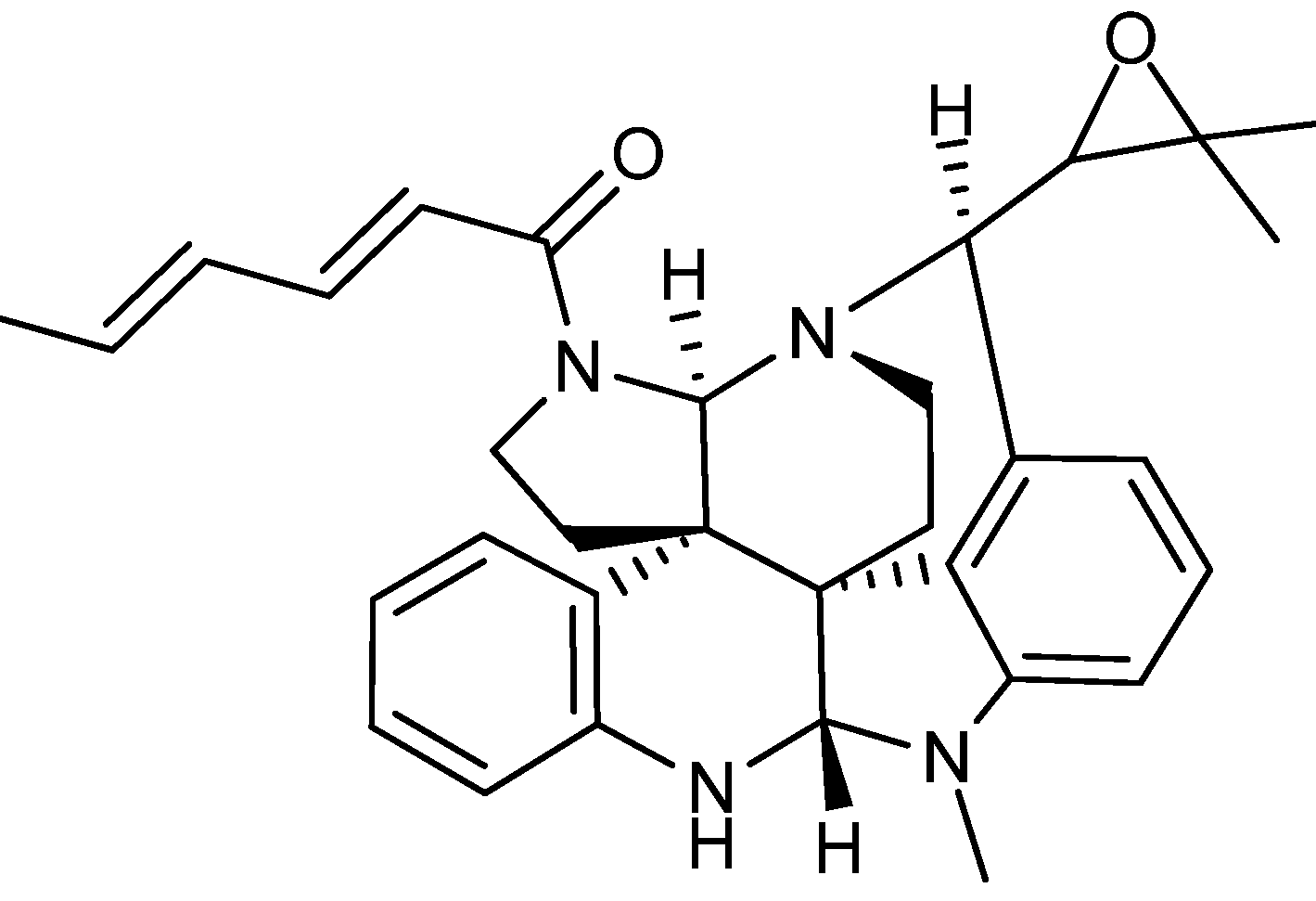
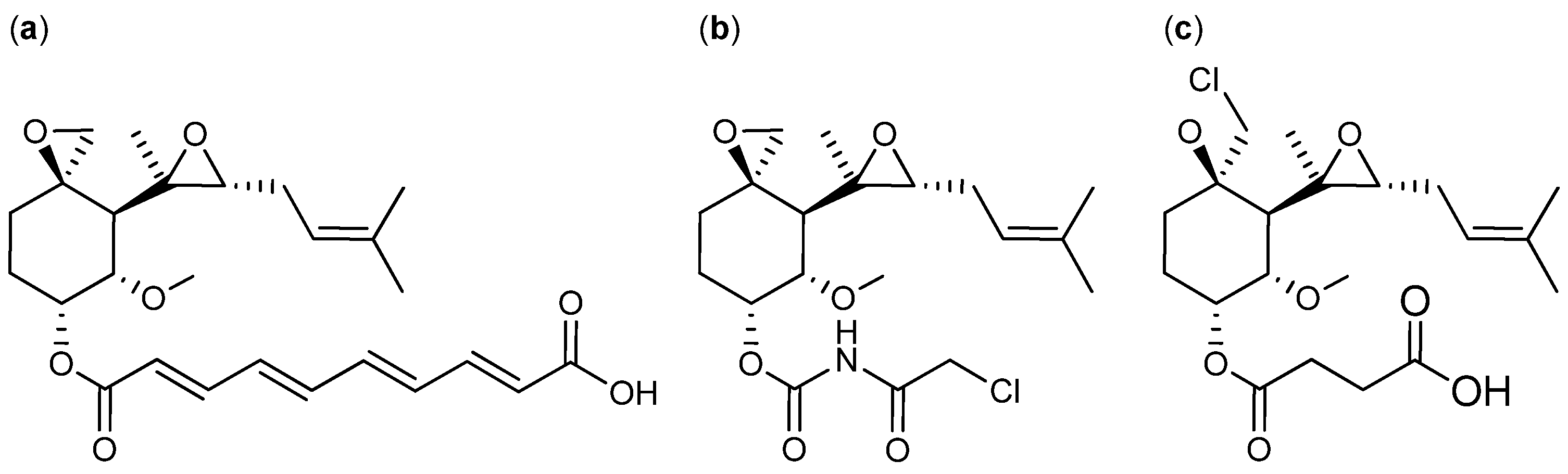
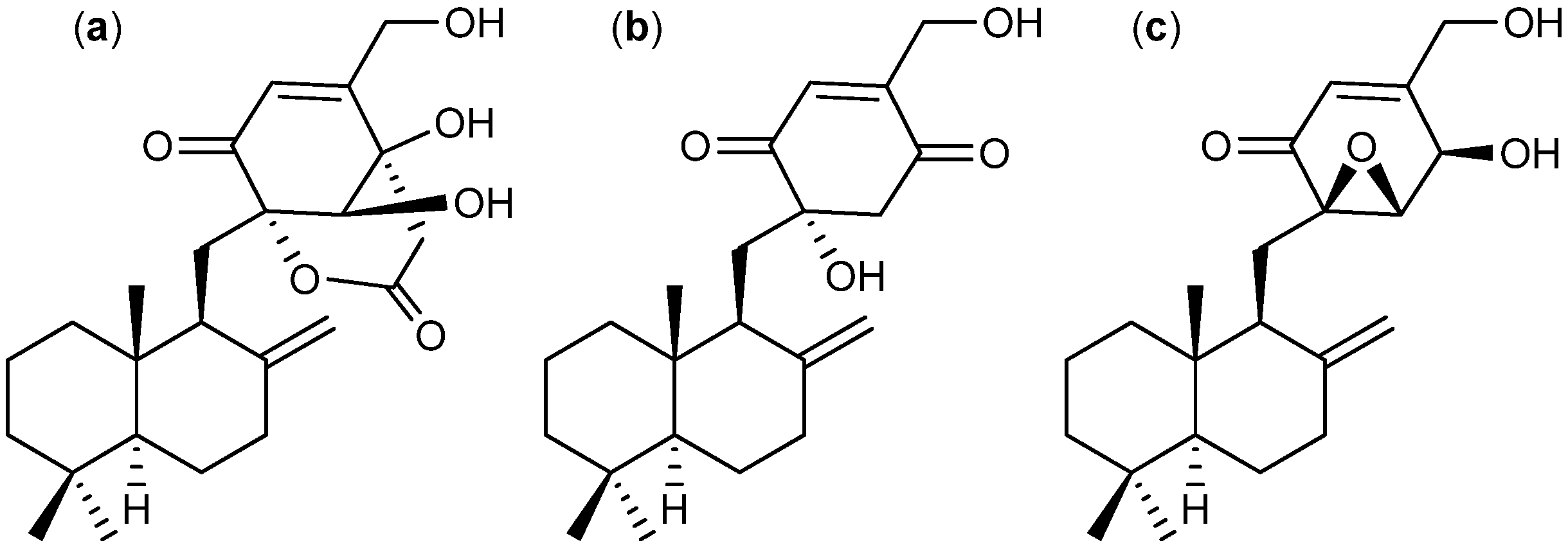
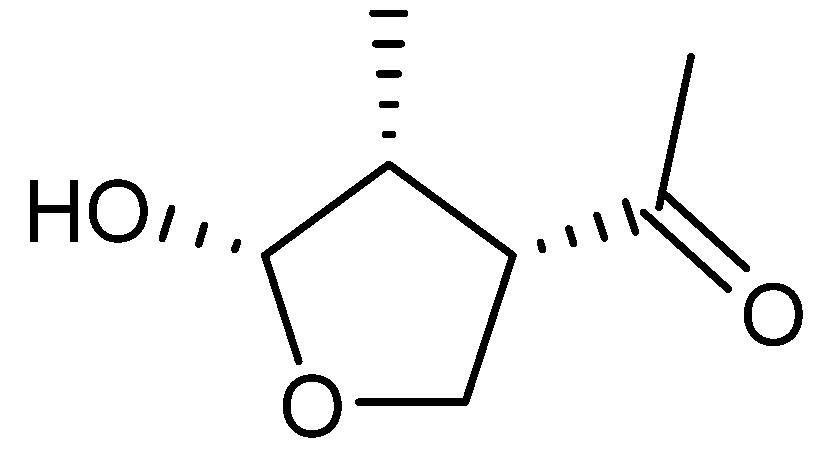
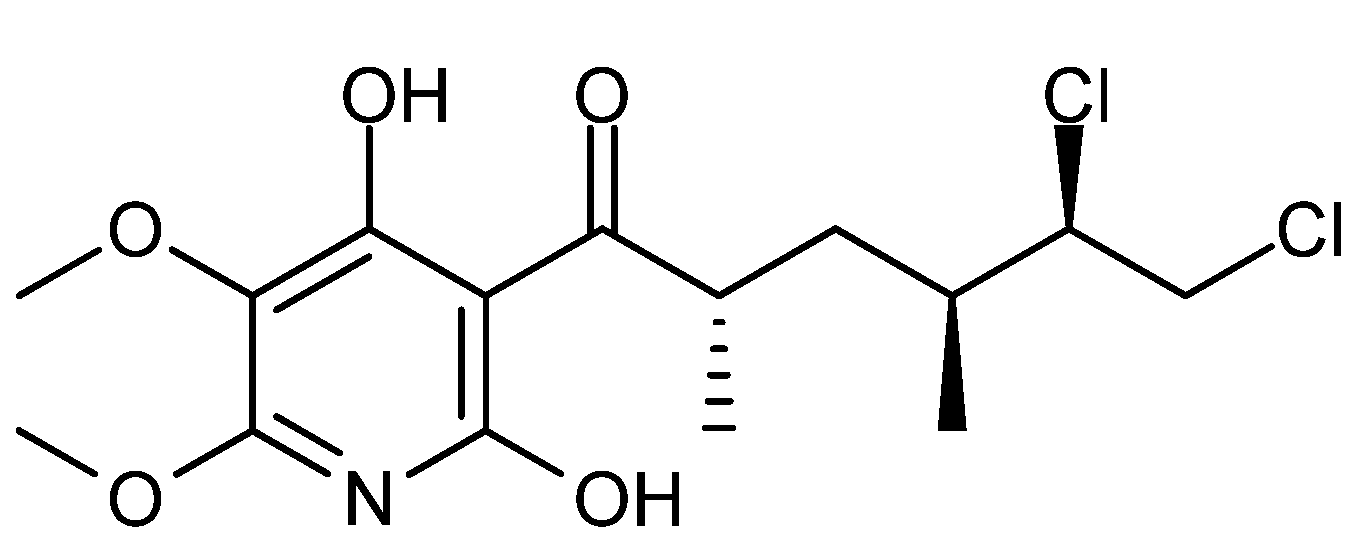

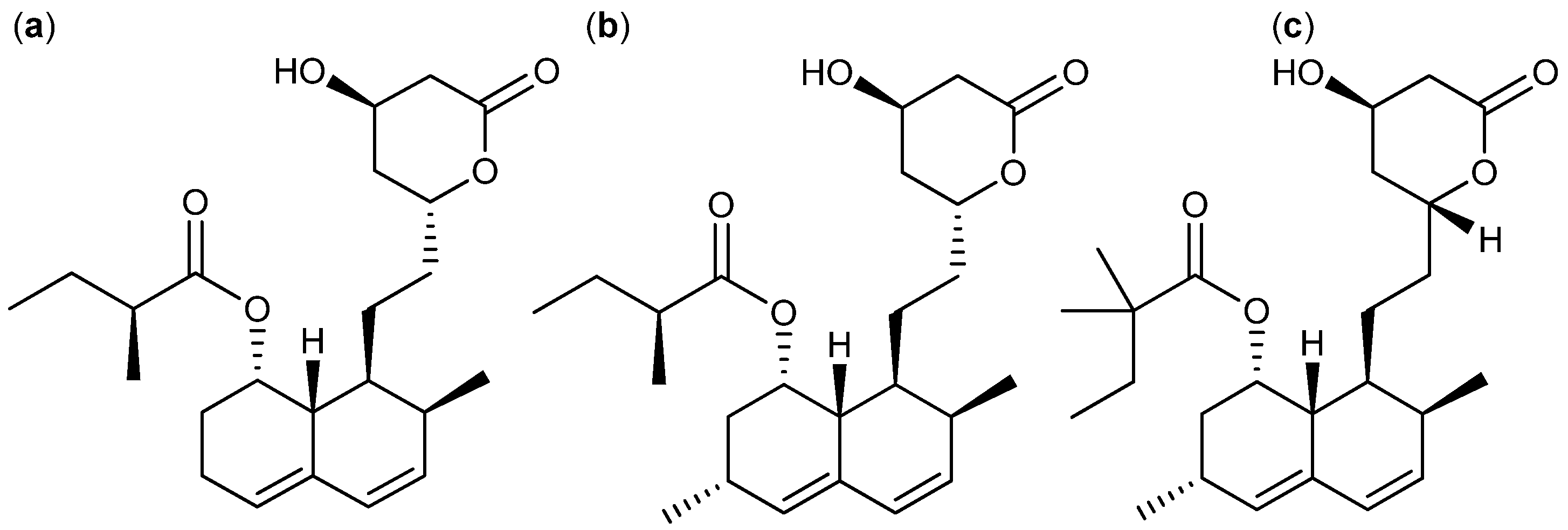{kind=link}
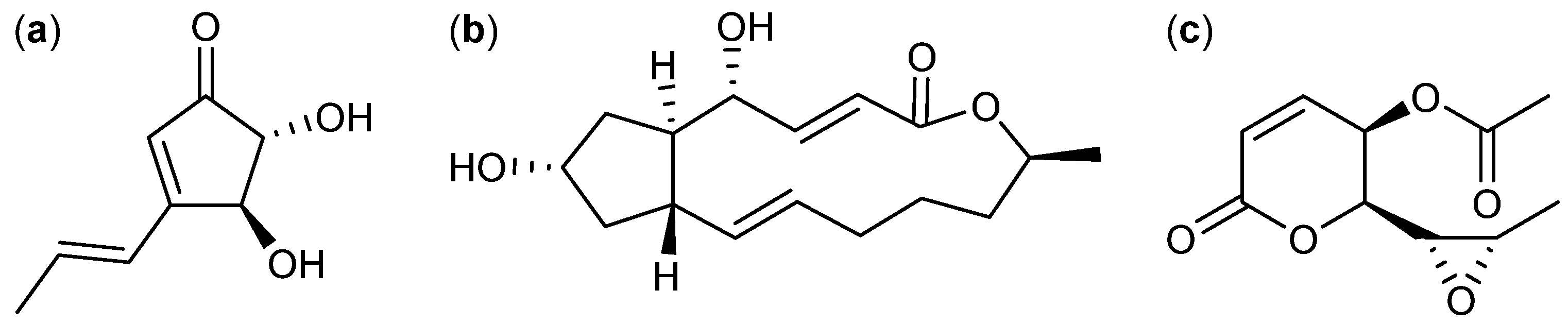{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
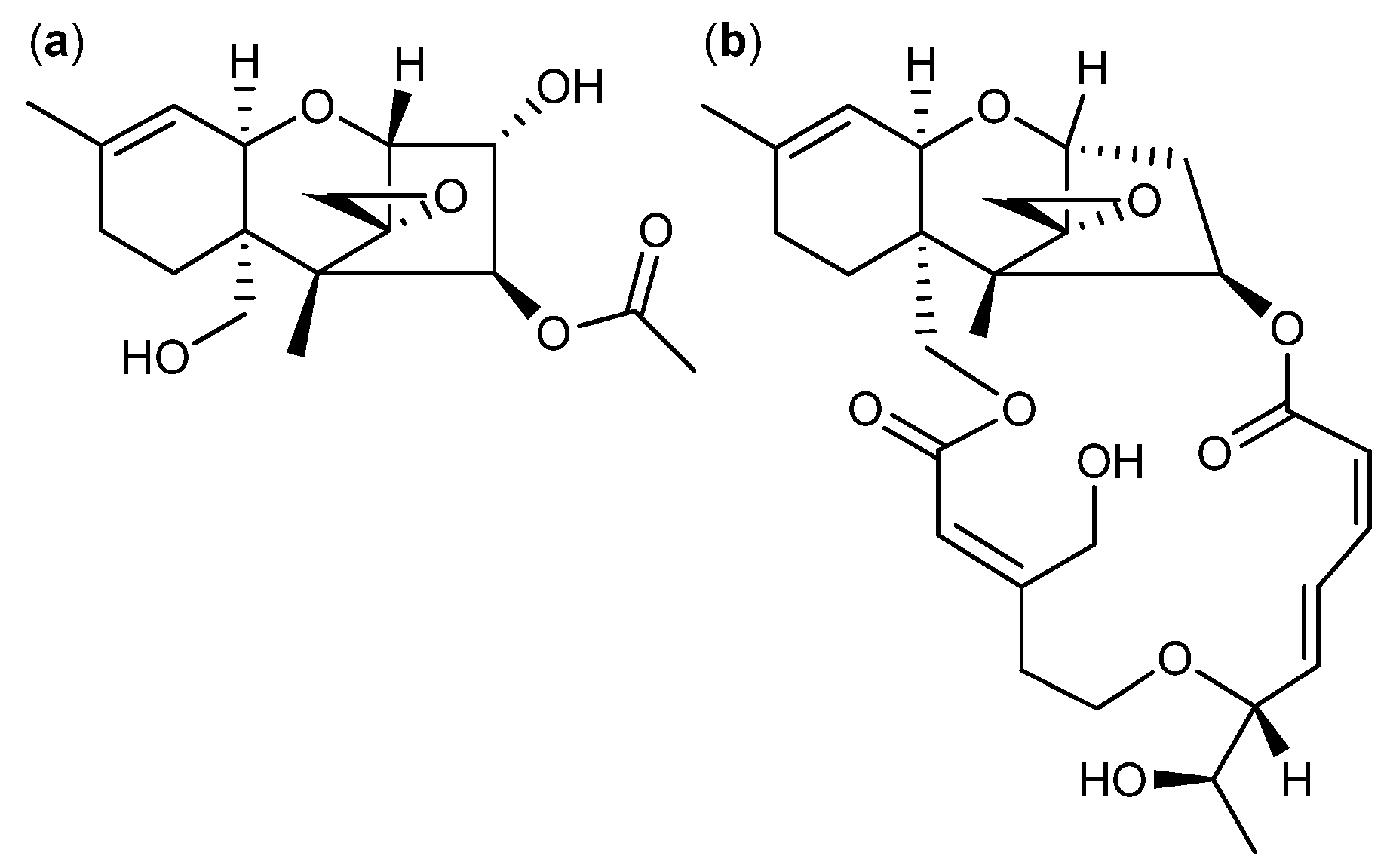{kind=link}
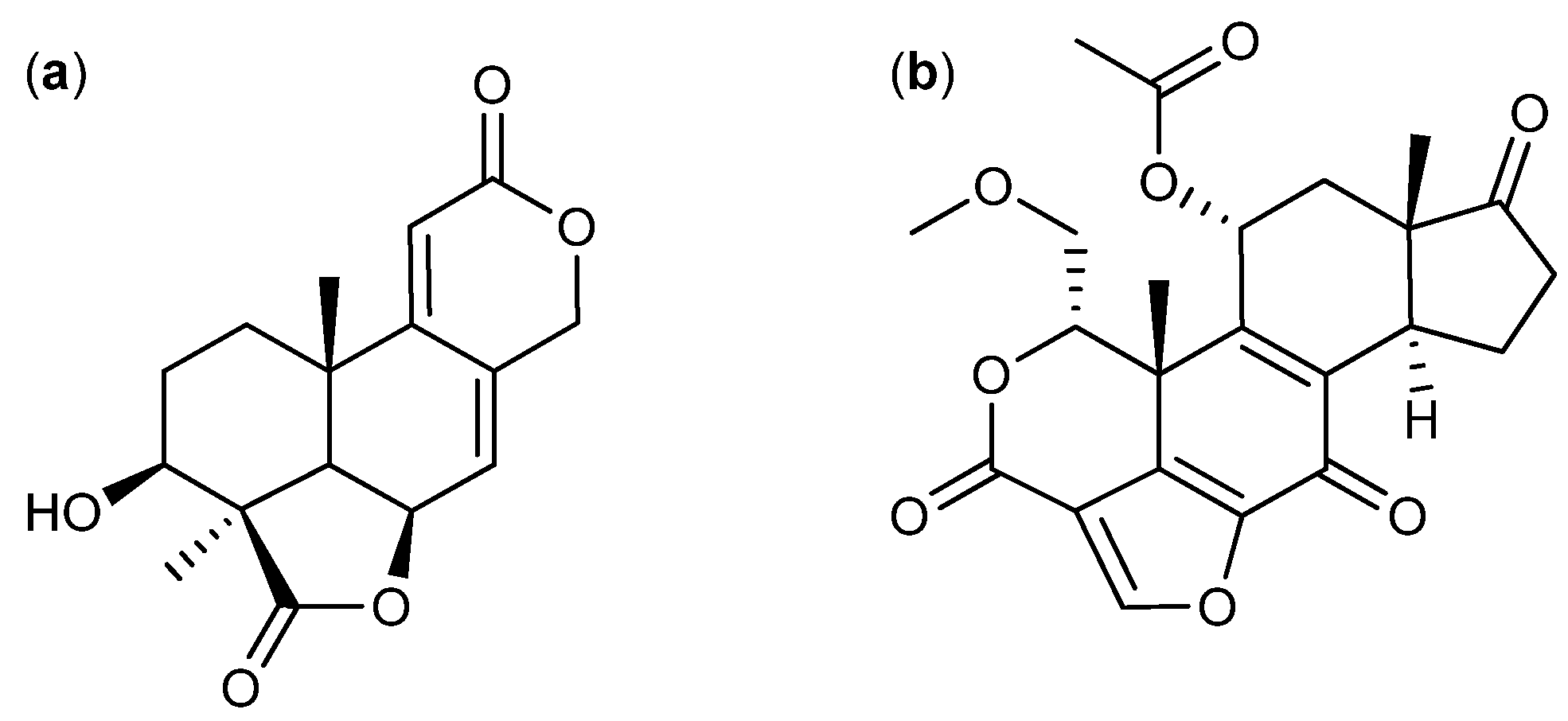{kind=link}
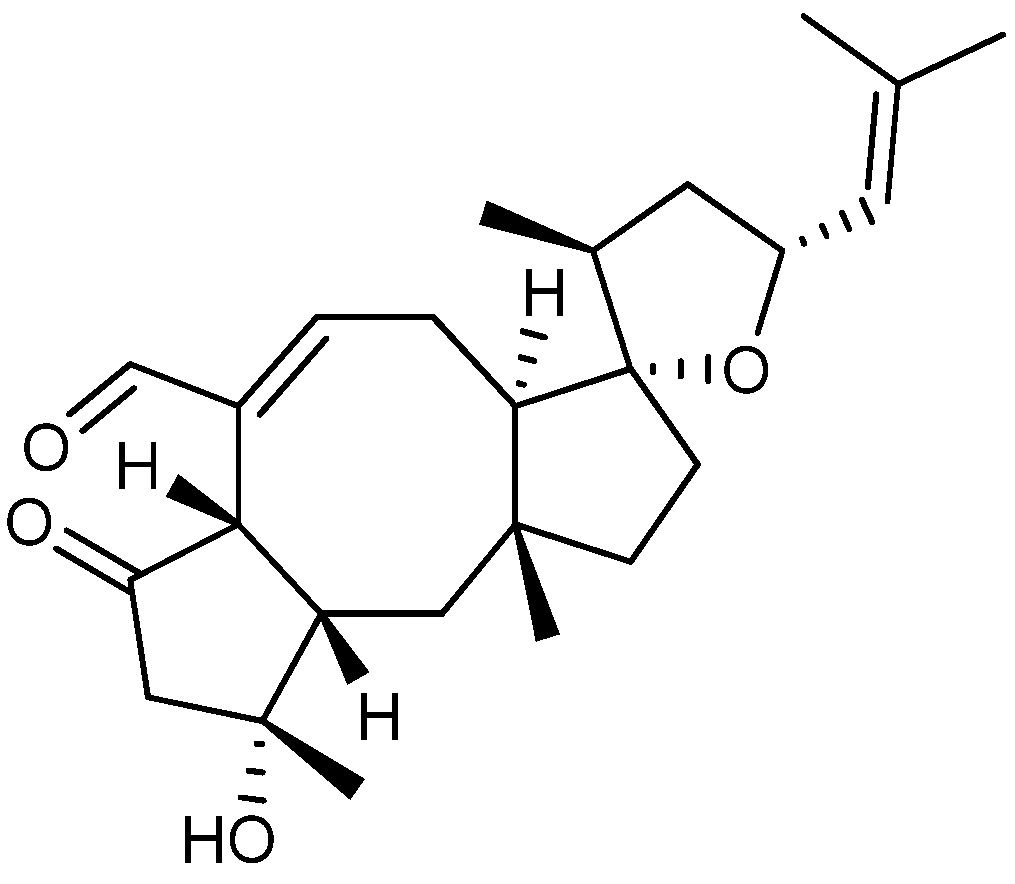{kind=link}
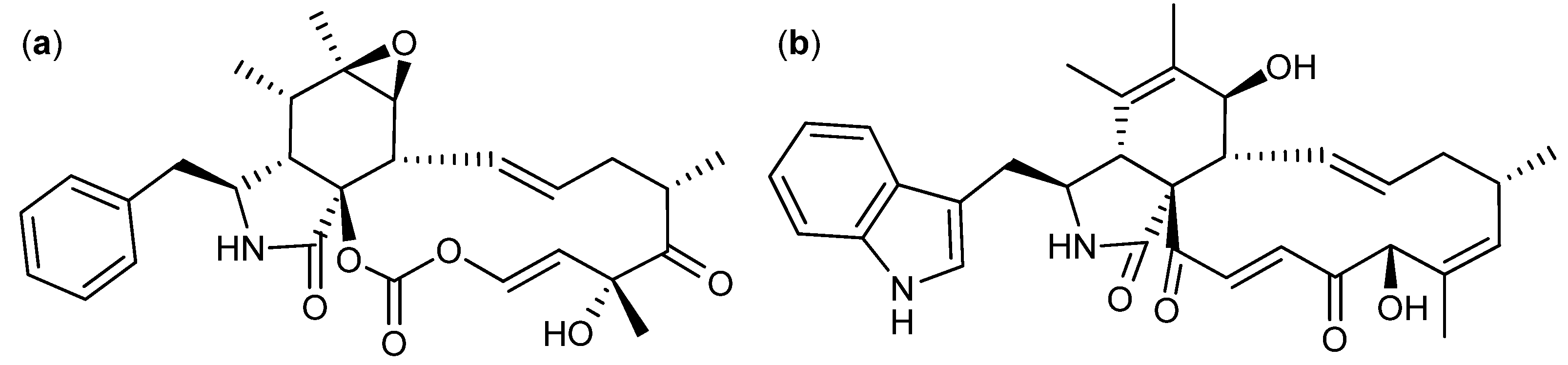{kind=link}
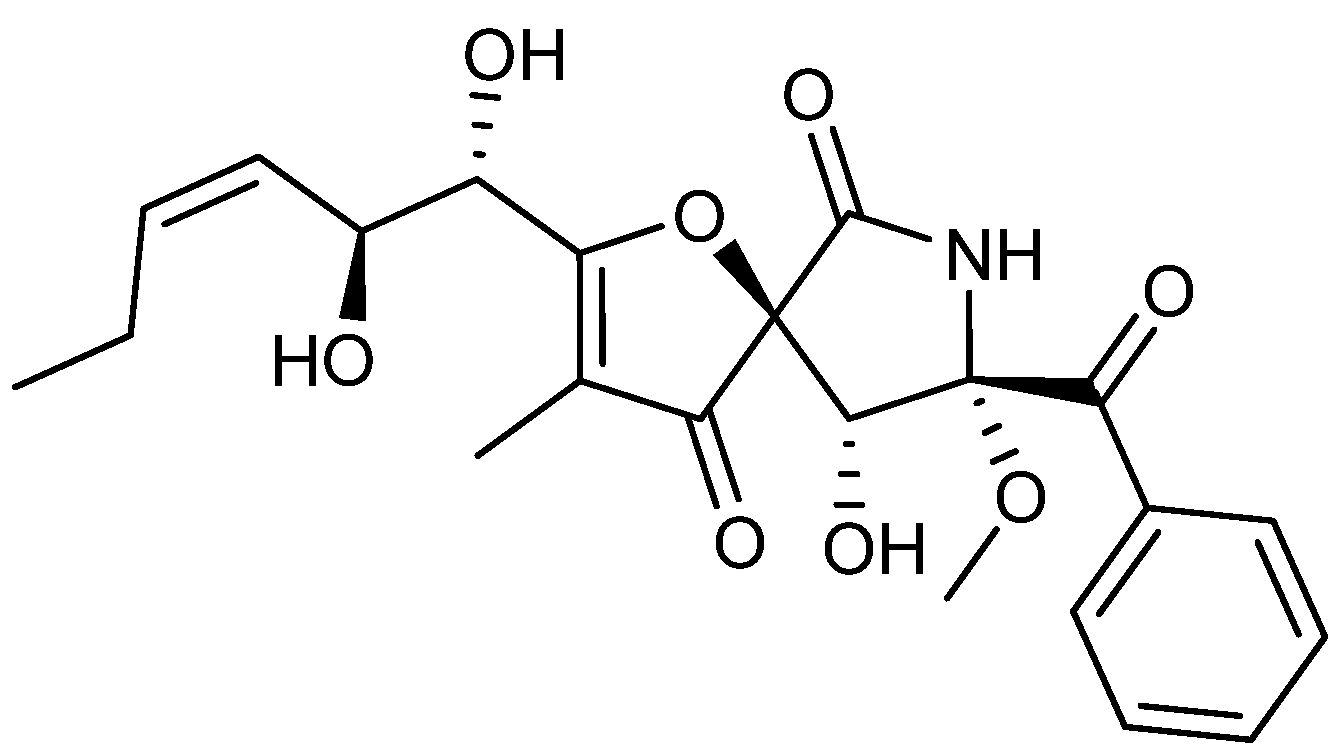{kind=link}
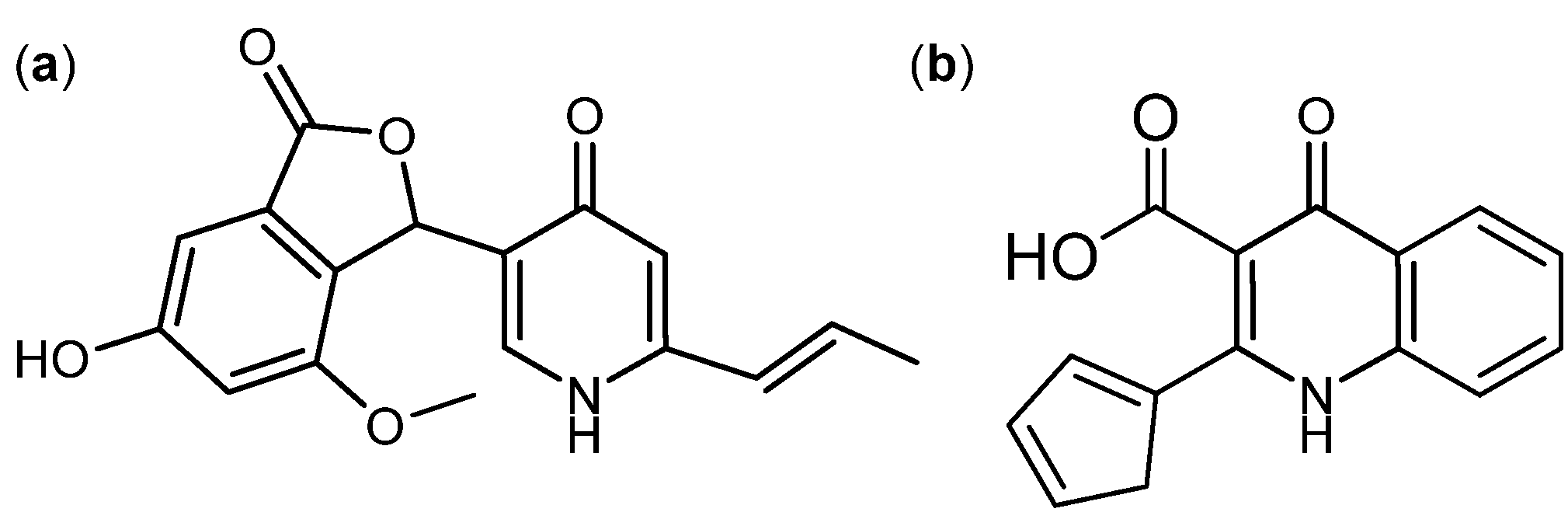{kind=link}
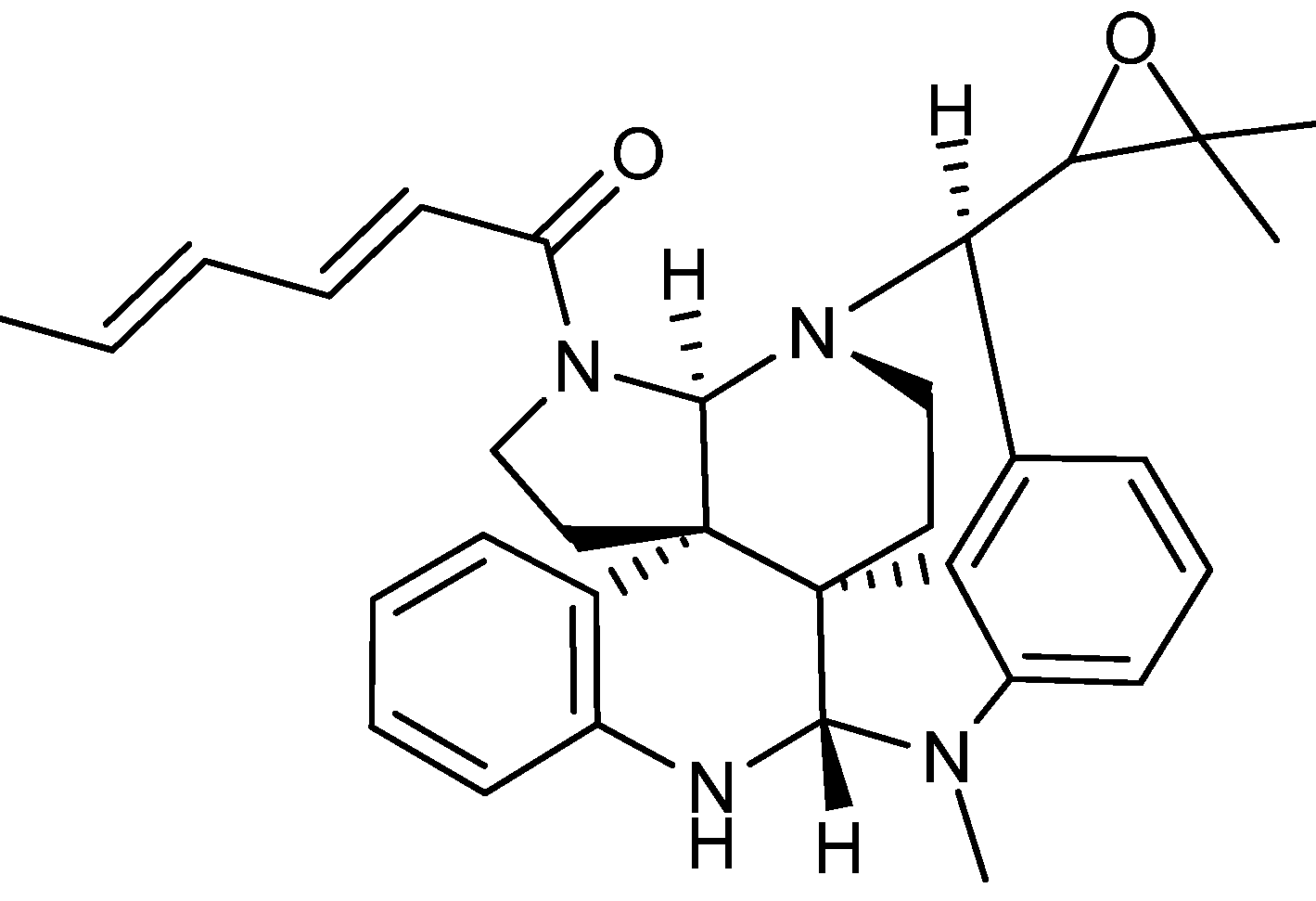{kind=link}
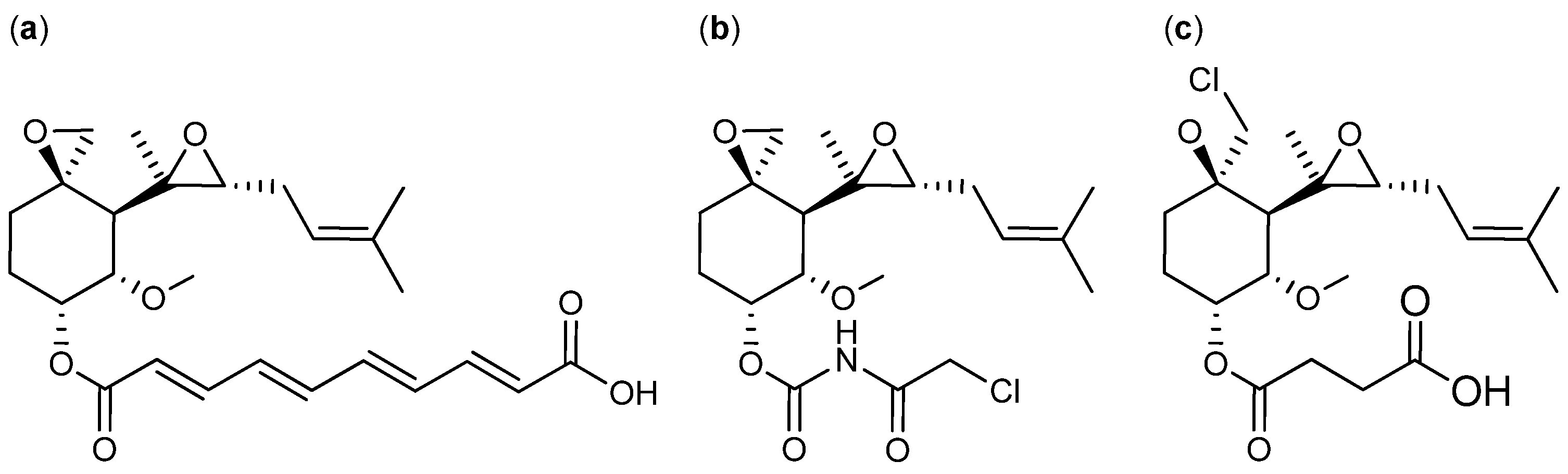{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (original published) | Compound | Type | Target cell | IC50 | Anti-fungal (+/−) |
|---|---|---|---|---|---|
| T. pinophilus [1] (P. pinophilum [48]) | 3-O-methylfunicone [50,51,52,53] | Polyketide | HeLa | - | + [48,49] |
| MCF-7 | - | ||||
| A-375P | - | ||||
| A-375M | - | ||||
| A. nidulans [46] | Asperlin [47] | Polyketide | HeLa | - | − |
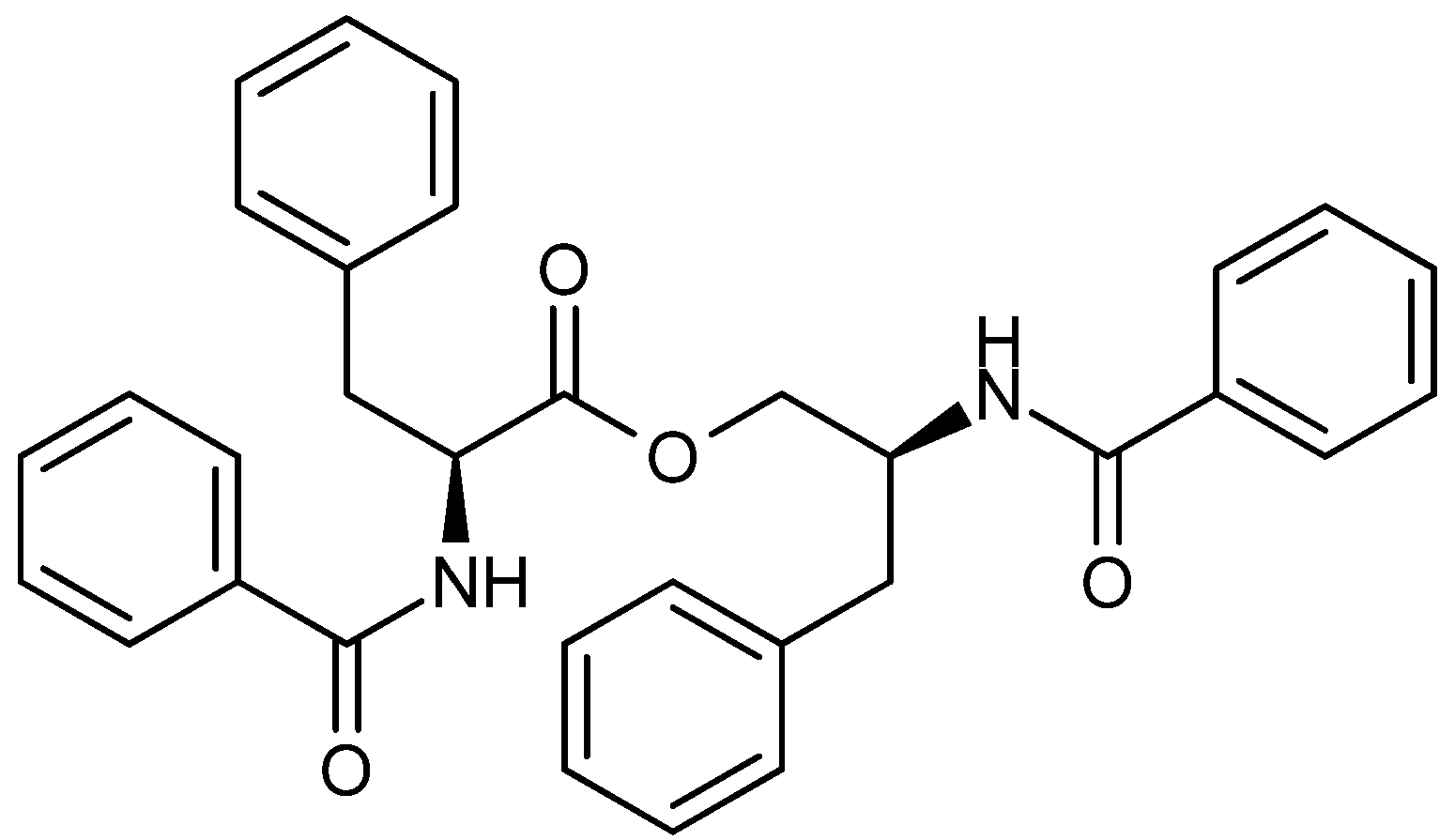
| A. flavus [95] Penicillium spp. [96] | Asperphenamate | NPR | - | - | − |
| A. flavus [95] Penicillium spp. [96] | BBP [99] | NPR | MCF-7 | 3.0 µM | |
| T47D | 4.7 µM | ||||
| MDA-MB231 | 5.2 µM | ||||
| BEL-4702 | 12.7 µM | ||||
| A-549 | 15.1 µM | ||||
| HeLa | 17.0 µM | ||||
| HL-60 | 18.3 µM | ||||
| Aspergillus sp. [131] | Acetylapoaranotin [131] | NPR | HCT-116 | 13.8 µM | − |
| AGS | 12 µM | ||||
| A-549 | 2 µM | ||||
| MCF-7 | 10 µM | ||||
| P. atramentosum [222] | Atpenin A5 [222] | Unresolved biosynthetic origin | DU-145 (co-culture) | 0.02 µg/mL | + [221] |
| A. puniceus [69] A. turkensis [69] A. pseudoustus [69] A. ustus [70] | Austocystin D [71,72] | Polyketide | SR | 16 nM (GI50) | − |
| U-87 | 4946 nM (GI50) | ||||
| MCF-7 | <1 nM (GI50) | ||||
| MDA-MB-231 | 549 nM (GI50) | ||||
| PC-3 | 3 nM (GI50) | ||||
| SW-620 | 27 nM (GI50) | ||||
| HCT-15 | 42 nM (GI50) | ||||
| MX-2 | 3358 nM (GI50) | ||||
| P. brevicompactum [20] P. paneum [223] T. stipitatus [1] (P. stipitatum [220]) | Botryodiplodin [219,220] | Unresolved biosynthetic origin | EAC | 0.6 µg/mL (ED50) | + [218] |
| L-5178 | 0.8 µg/mL (ED50) | ||||
| Sarcoma 37 | 1.5 µg/mL (ED50) | ||||
| HeLa | 2.0 µg/mL (ED50) | ||||
| P. brefeldianum [39] | Brefeldin A [42,43,44,45] | Polyketide | HL-60 | 35.7 nM | + [39,40] |
| KB | 32 nM | ||||
| HeLa | 6.4 nM | ||||
| MCF-7 | 7.1 nM | ||||
| SPC-A-1 | 3.6 nM | ||||
| BC-1 | 40 nM | ||||
| NCI-H187 | 110 nM | ||||
| Cladosporium cladosporioides [82] | Calphostin C [82,83] | Polyketide | HeLa S3 | 0.2–8.5 µM | − |
| MCF-7 | 0.2–2.7 µM | ||||
| ALL | - | ||||
| Chaetomium globosum [184] P. discolor [1] P. expansum [1] | Chaetoglobosin B [193] | Mixed biosynthetic origin | BC | 3.0 µM | + [56,186] |
| Chaetomium globosum [184] P. discolor [1] P. expansum [1] | Chaetoglobosin D [193] | Mixed biosynthetic origin | KKU-100 | 3.4 µM | |
| KKU-OCA17 | 12.2 µM | ||||
| Chaetomium globosum [184] P. discolor [1] P. expansum [1] | Chaetoglobosin U [194] | Mixed biosynthetic origin | KB | 16.0 µM | |
| Chaetomium globosum [184] P. discolor [1] P. expansum [1] | Chaetoglobosin X [195] | Mixed biosynthetic origin | H-22 | 7.5 µM | |
| Chaetomium globosum [57,58,59,60,61,63,64] T. pinophilus [1] | 11-epichaetomugilin I [62] | Polyketide | P-388 | 0.7 pM | + [56] |
| HL-60 | 1.0 pM | ||||
| L-1210 | 1.6 pM | ||||
| KB | 1.2 pM | ||||
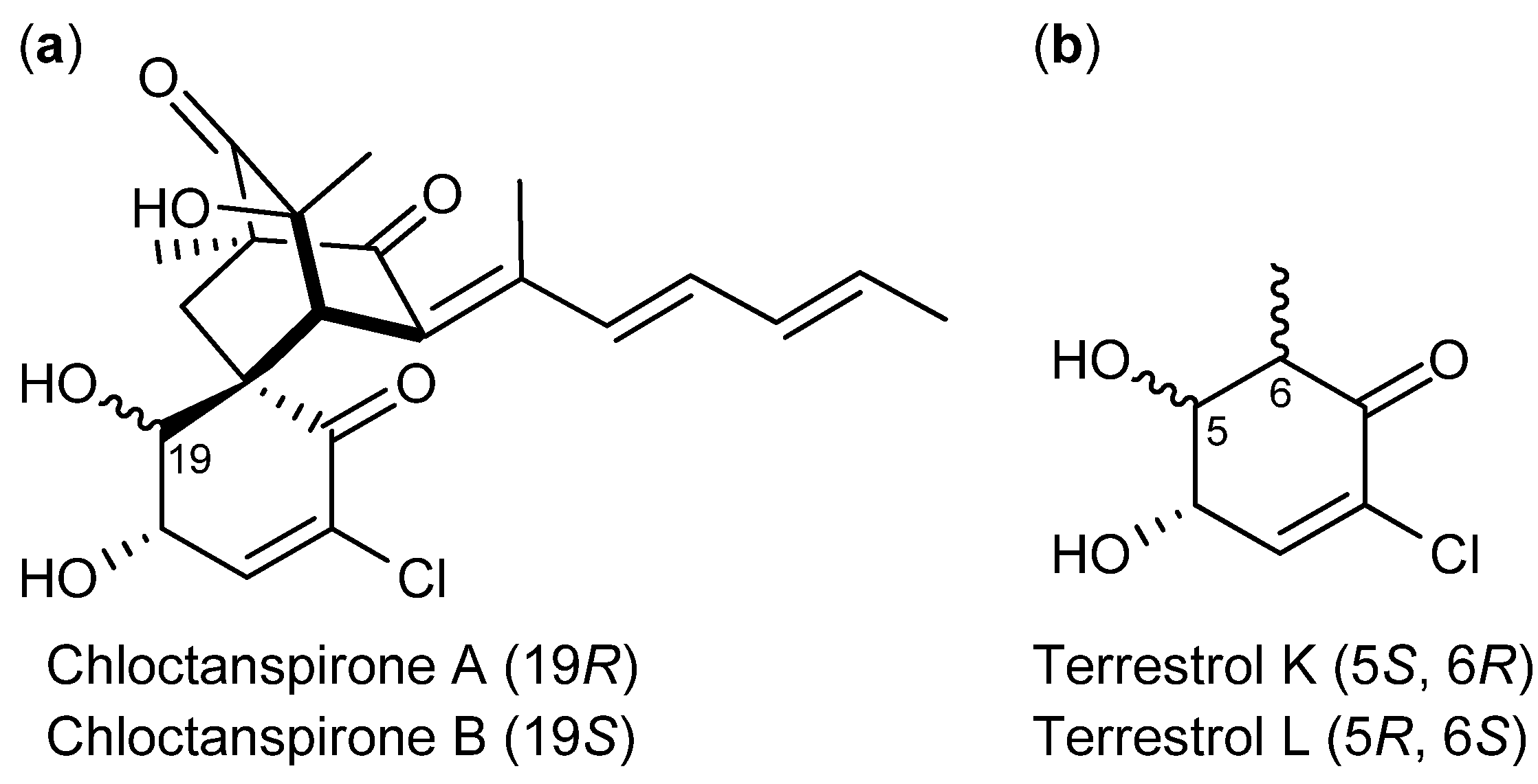
| P. chrysogenum [74] P. rubens [73] (P. terrestre [76]) | Chloctanspirone A [76] | Polyketide | HL-60 | 9.2 µM | − |
| A-549 | 39.7 µM | ||||
| P. buchwaldii [96] P. marinum [1] P. expansum [204] P. rivulorum [205] | Communesin B [206,207] | Mixed biosynthetic origin | P-388 | 0.5 µg/mL (ED50) | − |
| U-937 | 10.4 µg/mL (ED50) | ||||
| THP-1 | 11.4 µg/mL (ED50) | ||||
| NAMALWA | 9.9 µg/mL (ED50) | ||||
| MOLT-3 | 8.1 µg/mL (ED50) | ||||
| SUP-B15 | 7.2 µg/mL (ED50) | ||||
| Mariannaea elegans (Spicaria elegans [192]) A. clavatus [190] A. rhizopodus [90] Hypoxylon terricola [187] | Cytochalasin E [191,192] | Mixed biosynthetic origin | A-2780 | 0.02 µg/mL (IC100) | + [185] |
| HCT-116 | 1.0 µg/mL (IC100) | ||||
| SW-620 | 0.2 µg/mL (IC100) | ||||
| A-549 | 0.006 µM | ||||
| P-388 | 0.09 µM | ||||
| A. striatus (Emericella striata [128]) Cladorrhinum sp. | Emestrin A [129] | NPR | HL-60 | 83.5 nM | + [127] |
| A. striatus (Emericella striata [128]) Cladorrhinum sp. | Emestrin C [130] | NPR | DU-145 | 9.3 nM | |
| Penicillium sp. BCC 17468 [147] | Fudecadione A [147] | Terpene | MCF-7 | 12.6 µg/mL | − |
| KB | 22.3 µg/mL | ||||
| NCI-H187 | 24.9 µg/mL | ||||
| A. fumigatus [208] P. scabrosum [209] | Fumagillin [210] | Mixed biosynthetic origin | Angiogenesis inhibitor | - | − |
| TNP-470 [210,211,212,213,-214] | Synthetic | PC-3 | - | ||
| MDA-MB-231 | - | ||||
| Penicillium sp. [215] | Linerin [215] | Mixed biosynthetic origin | POS1 | 117 nM | |
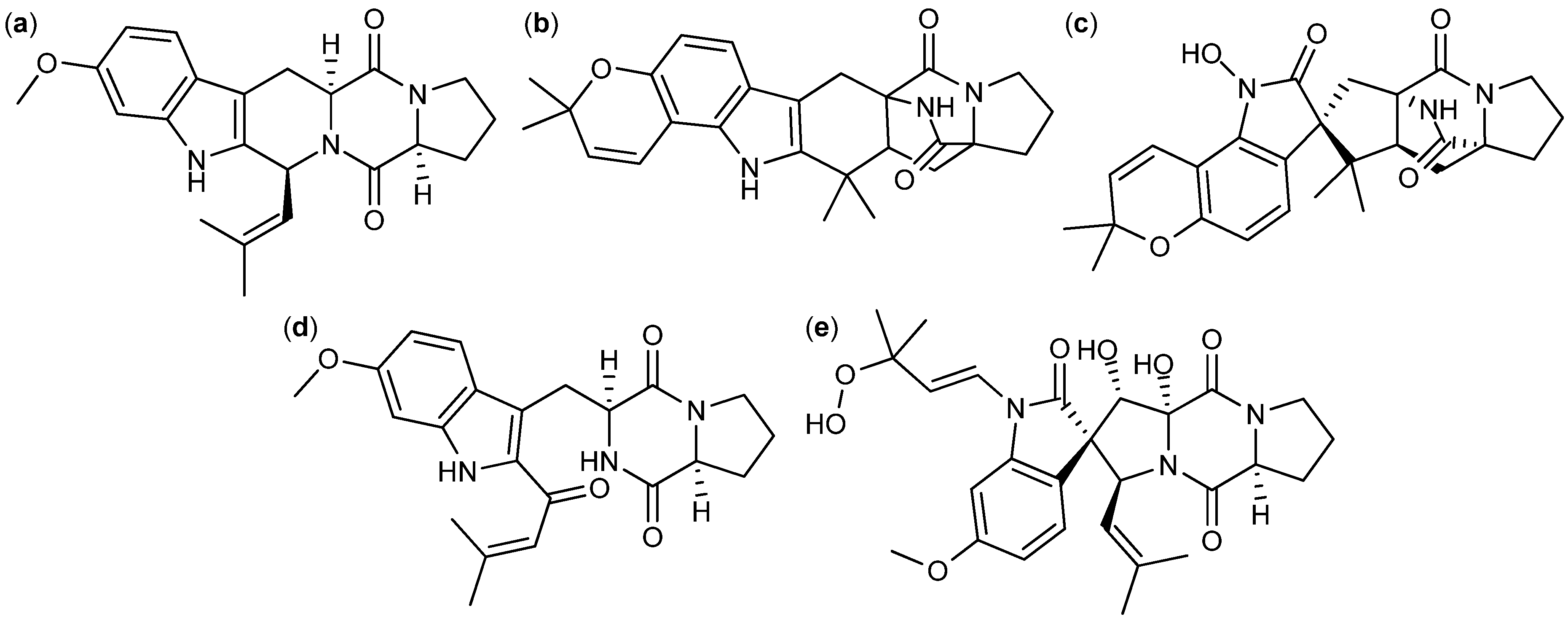
| A. fumigatus [101,102] A. fischeri [103] | Fumitremorgin C [111,113,114] | NPR | P-388 | 3.9 µg/mL (ED50) | + [224] |
| A. fumigatus [101,102] A. fischeri [103] | 12,13-dehydroxyfumitremorgin C [112] | NPR | U-937 | 1.8 µM | |
| PC-3 | 6.6 µM | ||||
| A. fumigatus [122] D. cejpii [123] | Gliotoxin [112,125,126] | NPR | MCF-7 | 985 nM | + [124] |
| T47D | 365 nM | ||||
| BT-474 | 102 nM | ||||
| ZR75-1 | 158 nM | ||||
| MDA MB231 | 38 nM | ||||
| MDA MB435 | 87 nM | ||||
| U-937 | 0.2 µM | ||||
| PC-3 | 0.4 µM | ||||
| P. griseofulvum [225] | Griseofulvin [2,3,4,226,227] | Polyketide | HeLa | 20 µM | + [226,228] |
| SCC-114 | 35 µM | ||||
| GF-15 [5] | Synthetic | SCC-114 | 0.9 µM | ||
| Hypocrella bambusae(Shiraia bambusicola [84,85]) | Hypocrellin D [85] | Polyketide | Bel-7721 | 1.8 µg/mL | + [229] |
| A-549 | 8.8 µg/mL | ||||
| Anip-973 | 38.4 µg/mL | ||||
| Synthetic analog [86] | Synthetic | MCF-7 (PDT) | - | ||
| Purpureocillium lilacinum [133] (P. lilacinum [132]) | Leucinostatin A [132,135,136,137,138] | NPR | Ehrlich | 1.6 mg/kg (LD50) | + [132,134] |
| L-1210 | 410.3 nM (IC100) | ||||
| BT-20 | 2.3 nM | ||||
| MCF-7 | 4 nM | ||||
| G-361 | 7 nM | ||||
| HT-144 | 6 nM | ||||
| A-549 | 16 nM | ||||
| A-427 | 59 nM | ||||
| SK-MES-1 | 12 µM | ||||
| Caov-3 | 17 nM | ||||
| Caov-4 | 53 nM | ||||
| SKOV-3 | 1,236 nM | ||||
| HT-29 | 119 nM | ||||
| LoVo | 114 nM | ||||
| HEp-2 | 40 nM | ||||
| HL-60 | 12 nM | ||||
| HSB-2 | 5 nM | ||||
| K-562 | 6 nM | ||||
| KG-1 | 46 nM | ||||
| RPMI-1788 | 158 nM | ||||
| SKW-6.4 | 6 nM | ||||
| U-937 | 319 nM | ||||
| Raji | 11 nM | ||||
| A. parasiticus [65,66] A. nidulans [67,68] | Norsolorinic acid [67,68] | Polyketide | T-24 | 10.5 µM | − |
| MCF-7 | 12.7 µM | ||||
| A. amoenus [115] | Notoamides [116,230] | NPR | HeLa/L-1210 | 22–52 µg/mL | − |
| Bipolaris sp. [169,170] A. calidoustus [69] A. insuetus [69] A. keveii [69] (A. ustus [171]) Sarocladium oryzae [173] (Cephalosporium caerulens [172] (D. maydis [174] D. sorghicola [174] | Ophiobolin A [175,176] | Terpene | A-549 | 0.1 µM | + [180,181] |
| HT-29 | 0.1 µM | ||||
| Mel-20 | 0.1 µM | ||||
| P-388 | 0.06 µM | ||||
| OVCAR-3 | 0.3 µM | ||||
| Bipolaris oryzae [179] | 3-anhydro-6-hydroxyophiobolin A [179] | Terpene | HepG2 | 6.5 µM | |
| K-562 | 4.1 µM | ||||
| Aspergillus section Usti [69] (A. ustus [177]) | Ophiobolin O [177,178] | Terpene | MFC-7 | 17.9 µM | |
| P-388 | 4.7 µM | ||||
| Talaromyces sp. (Penicillium sp.) [199] | Penicidone B [199] | Mixed biosynthetic origin | SW1116 | 54.2 µM | − |
| K-562 | 21.1 µM | ||||
| KB | 29.6 µM | ||||
| HeLa | 35.2 µM | ||||
| Penicillium sp. [200] Auxarthron reticulatum [201] | Penicinoline [202] | Mixed biosynthetic origin | HepG2 | 13.2 µM | − |
| Penicilliumsp. [200] Auxarthron reticulatum [201] | Methyl-penicinoline [202] | Mixed biosynthetic origin | HepG2 | 11.3 µM | |
| T. pinophilus [1] (P. simplicissimum [54]) | Penisimplicissin [55] | Polyketide | HL-60 | −6.7 (log10 GI50) | − |
| CCRF-CEM | −5.8 (log10 GI50) | ||||
| Phaeosphaeria sp. [87] | Phaeosphaerin B [87] | Polyketide | PC-3 | 2.4 µM | − |
| DU-145 | 9.5 µM | ||||
| LNCaP | 2.7 µM | ||||
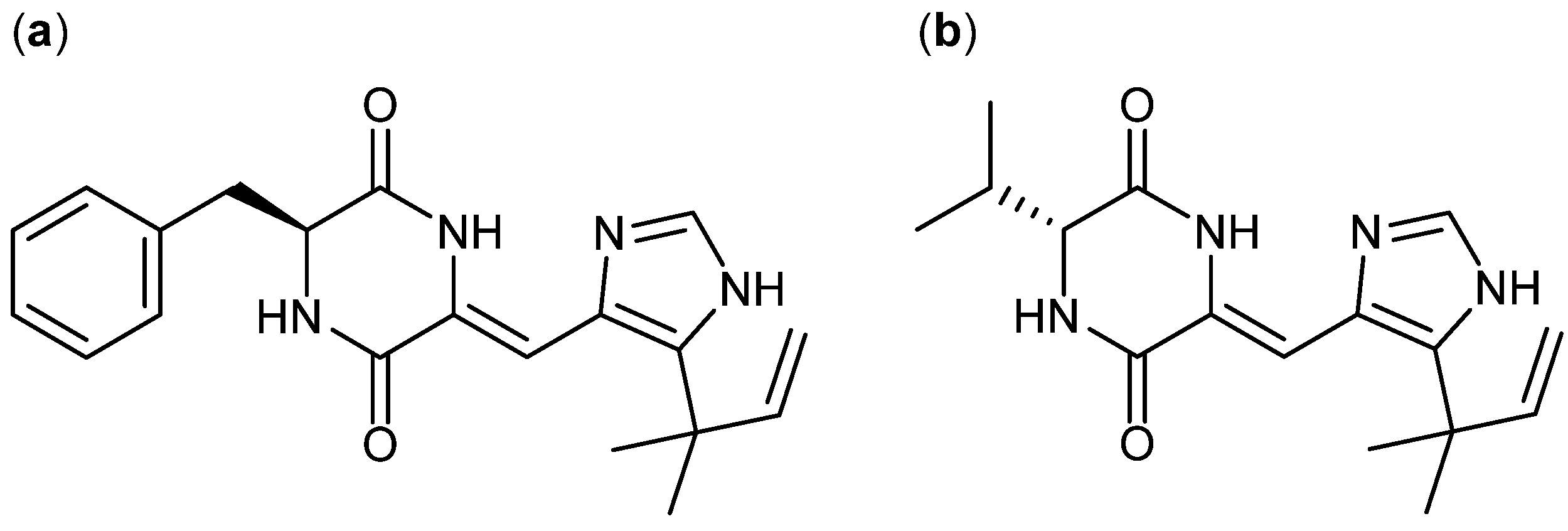
| A. ustus [117] | Phenylahistin [118] | NPR | A-431 | 0.22 µM | − |
| A-549 | 0.30 µM | ||||
| HeLa | 0.20 µM | ||||
| K-562 | 0.19 µM | ||||
| P-388 | 0.33 µM | ||||
| MCF-7 | 3.7 µM | ||||
| TE-671 | 0.18 µM | ||||
| A. taichungensis [79] | Prenylterphenyllin A [79] | Polyketide | A-549 | 8.3 µM | − |
| HL-60 | 1.5 µM | ||||
| A. candidus [77,78] | 4”-deoxyisoterprenin [78] | Polyketide | KB3-1 | 6.2 µM | |
| A. taichungensis [79] | Prenylcandidusin B [79] | Polyketide | P-388 | 1.6 µM | |
| Pseudeurotium ovalis [196] A. fumigatus [197] | Pseurotin D [198] | Mixed biosynthetic origin | MFC-7 | 15.6 µM | − |
| T. purpurogenus mutant BD-1-6 [1] (P. purpurogenum [216]) | Purpurogemutantidin [216] | Mixed biosynthetic origin | K-562 | 0.9 µM | + [217] |
| HeLa | 16.6 µM | ||||
| BGC-823 | 31.0 µM | ||||
| MCF-7 | 26.3 µM | ||||
| T. purpurogenus mutant BD-1-6 [1] (P. purpurogenum [216]) | Macrophorin A [216] | Mixed biosynthetic origin | HL-60 | 0.9 µM | |
| A. parasiticus [231] | Sequoiamonascin A [231] | Polyketide | MCF-7 | 1% | − |
| NCI-H460 | 1% | ||||
| SF-268 | 2% | ||||
| Percent cell growth compared to untreated cells at 10 µM | |||||
| P. solitum [20,25] (P. brevicompactum [18]) P. hirsutum [20] (P. citrinum [19]) | Compactin [28] | Polyketide | AML | 2.6 µM (IC100) | + [18] |
| A. terreus [26] Monascus sp. [27] | Lovastatin [29,30,31,32] | Polyketide | OVHS-1 | 39 µM | + [22,23,24] |
| Calu-1 | 3 µM | ||||
| H-460 | 3 µM | ||||
| A-549 | 10 µM | ||||
| H-441 | 30 µM | ||||
| Ovca-432 | 2 µM | ||||
| A-2780 | 3 µM | ||||
| Hey | 3 µM | ||||
| Ovca-429 | 5 µM | ||||
| HOC-7 | 10 µM | ||||
| DOV-13 | 11 µM | ||||
| Skov-3 | 21 µM | ||||
| A-2780-ADR | 21 µM | ||||
| A-2780-CIS | 22 µM | ||||
| MCF-7 | 1.7 µM | ||||
| HepG2 | 2.7 µM | ||||
| HeLa | 1.5 µM | ||||
| AML | - | ||||
| Simvastatin [1,20,29,33,34] | Synthetic | DLRP | 0.9 µM | + [23,24] | |
| H-1299 | 1.3 µM | ||||
| HT-144 | 1.0 µM | ||||
| MI-14 | 0.8 µM | ||||
| SK-MEL-28 | 0.8 µM | ||||
| BT-474A | 4.2 µM | ||||
| SKBR-3 | 2.2 µM | ||||
| MDA-MB-453 | 5.4 µM | ||||
| BT-20 | 1.7 µM | ||||
| AML | - | ||||
| A. ochraceus [106] A. westerdijkiae [107] | Stephacidin B [106] | NPR | PC-3 | 0.4 µM | − |
| LNCaP | 0.06 µM | ||||
| A-2780 | 0.3 µM | ||||
| A-2780/DDP | 0.4 µM | ||||
| A-2780/Tax | 0.3 µM | ||||
| HCT-116 | 0.5 µM | ||||
| HCT-116/mdr+ | 0.5 µM | ||||
| HCT-116/topo | 0.4 µM | ||||
| MCF-7 | 0.3 µM | ||||
| SKBR-3 | 0.3 µM | ||||
| LX-1 | 0.4 µM | ||||
| Taxomyces andreanea [140] P. raistrickii [141] | Taxol [139,142,143,144,145] | Terpene | Clinical use: Ovarian tumors Breast tumors Lung tumors Kaposi’s sarcoma | - | + [146] |
| A. terreus [148] | Terrecyclic acid A [148,149,232] | Terpene | NCI-H460 | 10.6 µM | + [148] |
| MCF-7 | 24.1 µM | ||||
| SF-268 | 14.7 µM | ||||
| P-388 | 63-125 mg/kg (LD50) | ||||
| A. terreus [37] | Terrein [38] | Polyketide | MCF-7 | 1.1 nM | + [36] |
| PANC-1 | 9.8 µM | ||||
| HepG2 | 66.8 µM | ||||
| Fusarium spp. [151] Isaria japonica [157] | 4-Acetyl-12,13-epoxy-9-trichothecene- 3,15-diol (AETD) [157] | Terpene | HL-60 | 10 nM | + [154] |
| U-937 | 22 nM | ||||
| HeLa | 45 nM | ||||
| MCF-7 | 53 nM | ||||
| Hep-G2 | 170 nM | ||||
| Fusarium spp. [151] Myrothecium roridum [158] | 12’hydroxyroridin E [158] | Terpene | L-1210 | 0.2 µM | |
| BC-1 | 0.9 µM | ||||
| NCI-H187 | 1.5 µM | ||||
| P. citrinum [80] | Tricitrinol B [80] | Polyketide | HL-60 | 3.2 µM | − |
| HCT-116 | 4.8 µM | ||||
| KB | 3.9 µM | ||||
| A. fumigatus [104,105,108,109] | Ds2-tryprostatin B [108] | NPR | NCI-H-522 | 15.8 µM (GI50) | − |
| MCF-7 | 15.9 µM (GI50) | ||||
| PC-3 | 11.9 µM (GI50) | ||||
| A. fumigatus [104,105,108,109] | 18-oxotryprostatin A [110] | NPR | A-549 | 1.3 µM | |
| A. fumigatus [104,105,108,109] | Spirotryprostatin E [109] | NPR | MOLT-4 | 3.1 µM | |
| HL-60 | 2.3 µM | ||||
| A-549 | 3.1 µM | ||||
| Penicillium sp. [150] | Unnamed chloro-trinoreremophilane sesquiterpene [150] | Terpene | HL-60 | 11.8 µM | − |
| A-549 | 12.2 µM | ||||
| A. wentii [160] | Wentilactones [161] | Terpene | SMMC-7721 | 19 µM | + [233] |
| T. wortmannii [1] (P. wortmannii [162]) Or similar species. | Wortmannin [163,164,165,166,167,168] | Terpene | HL-60 | 30 nM | + [162] |
| K-562 | 25 nM | ||||
| KNS-62 | 100–200 nM | ||||
| Colo-699 | 100–200 nM | ||||
| P. chrysogenum [1] D. cejpii [90] (D. albus [90]) | Xanthocillin X [89,91,92] | NPR | Ehrlich ascites carcinoma-mouse strain | 40 mg/kg (LD50) | + [89] |
| K-562 | 0.4 µg/mL | ||||
| HeLa | 1.2 µg/mL | ||||
| MCF-7 | 12 µg/mL | ||||
| HepG2 | 7 µg/mL | ||||
| NCI-H460 | 10 µg/mL | ||||
| Du-145 | 8 µg/mL | ||||
| MDA-MB-231 | 8 µg/mL | ||||
| Aspergillus sp. [93] | BU-4704 [93] | NPR | HCT-116 | 0.6 µg/mL | |
| B16-F10 | 4.3 µg/mL | ||||
| P. chrysogenum [1] | Xanthocillin X di-methoxy [94] | NPR | HepG2 | 0.2 µg/mL | |
| MCF-7 | 0.4 µg/mL | ||||
| KB | 0.4 µg/mL |
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Panda, D.; Rathinasamy, K.; Santra, M.K.; Wilson, L. Kinetic suppression of microtubule dynamic instability by griseofulvin: implications for its possible use in the treatment of cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 9878–9883. [Google Scholar] [CrossRef]
- Rebacz, B.; Larsen, T.O.; Clausen, M.H.; Rønnest, M.H.; Löffler, H.; Ho, A.D.; Krämer, A. Identification of griseofulvin as an inhibitor of centrosomal clustering in a phenotype-based screen. Cancer Res. 2007, 67, 6342–6350. [Google Scholar] [CrossRef]
- Ho, Y.-S.; Duh, J.-S.; Jeng, J.-H.; Wang, Y.-J.; Liang, Y.-C.; Lin, C.-H.; Tseng, C.-J.; Yu, C.-F.; Chen, R.-J.; Lin, J.-K. Griseofulvin potentiates antitumorigenesis effects of nocodazole through induction of apoptosis and G2/M cell cycle arrest in human colorectal cancer cells. Int. J. Cancer 2001, 91, 393–401. [Google Scholar]
- Rønnest, M.H.; Rebacz, B.; Markworth, L.; Terp, A.H.; Larsen, T.O.; Krämer, A.; Clausen, M.H. Synthesis and structure-activity relationship of griseofulvin analogues as inhibitors of centrosomal clustering in cancer cells. J. Med. Chem. 2009, 52, 3342–3347. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Global species numbers of fungi: are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers. Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Klejnstrup, M.L.; Frandsen, R.J.N.; Holm, D.K.; Nielsen, M.T.; Mortensen, U.H.; Larsen, T.O.; Nielsen, J.B. Genetics of polyketide metabolism in Aspergillus nidulans. Metabolites 2012, 2, 100–133. [Google Scholar] [CrossRef]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar] [CrossRef]
- Henrikson, J.C.; Hoover, A.R.; Joyner, P.M.; Cichewicz, R.H. A chemical epigenetics approach for engineering the in situ biosynthesis of a cryptic natural product from Aspergillus niger. Org. Biomol. Chem. 2009, 7, 435–438. [Google Scholar] [CrossRef]
- Laatsch, H. AntiBase 2012. Available online: http://eu.wiley.com/WileyCDA/WileyTitle/productCd-3527334068.html/ (accessed on 5 September 2013).
- Nielsen, K.F.; Månsson, M.; Rank, C.; Frisvad, J.C.; Larsen, T.O. Dereplication of microbial natural products by LC-DAD-TOFMS. J. Nat. Prod. 2011, 74, 2338–2348. [Google Scholar] [CrossRef]
- Hansen, M.E.; Smedsgaard, J.; Larsen, T.O. X-hitting: an algorithm for novelty detection and dereplication by UV spectra of complex mixtures of natural products. Anal. Chem. 2005, 77, 6805–6817. [Google Scholar] [CrossRef]
- Larsen, T.O.; Petersen, B.O.; Duus, J.Ø.; Sørensen, D.; Frisvad, J.C.; Hansen, M.E. Discovery of new natural products by application of X-hitting, a novel algorithm for automated comparison of full UV spectra, combined with structural determination by NMR spectroscopy. J. Nat. Prod. 2005, 68, 871–874. [Google Scholar] [CrossRef]
- Wang, L.-W.; Zhang, Y.-L.; Lin, F.-C.; Hu, Y.-Z.; Zhang, C.-L. Natural products with antitumor activity from endophytic fungi. Mini-Rev. Med. Chem. 2011, 11, 1056–1074. [Google Scholar] [CrossRef]
- Kharwar, R.N.; Mishra, A.; Gond, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Taylor, J.W.; Wingfield, M.J.; Abaci, O.; Aime, C.; et al. The Amsterdam declaration on fungal nomenclature. IMA Fungus 2011, 2, 105–112. [Google Scholar]
- Brown, A.G.; Smale, T.C.; King, T.J.; Hasenkamp, R.; Thompson, R.H. Crystal and molecular structure of compactin, a new antifungal metabolite from Penicillium brevicompactum. J. Chem. Soc. Perk. T. 1 1976, 1976, 1165–1173. [Google Scholar]
- Endo, A.; Kuroda, M.; Tsujita, Y. ML-236B, and ML-236C, new inhibitors of cholesterogenesis produced by Penicillium citrinum. J. Antibiot. 1976, 29, 1346–1348. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Filtenborg, O. Terverticillate penicillia: Chemotaxonomy and mycotoxin production. Mycologia 1989, 81, 837–861. [Google Scholar] [CrossRef]
- Wong, W.-L.; Dimitroulakos, J.; Minden, M.; Penn, L. HMG-CoA reductase inhibitors and the malignant cell: the statin family of drugs as triggers of tumor-specific apoptosis. Leukemia 2002, 16, 508–519. [Google Scholar] [CrossRef]
- Qiao, J.; Kontoyiannis, D.P.; Wan, Z.; Li, R.; Liu, W. Antifungal activity of statins against Aspergillus species. Med. Mycol. 2007, 45, 589–593. [Google Scholar] [CrossRef]
- Chamilos, G.; Lewis, R.E.; Dimitrios, P.; Kontoyiannis, D.P. Lovastatin has significant activity against zygomycetes and interacts synergistically with voriconazole. Antimicrob. Agents Chemother. 2006, 50, 96–103. [Google Scholar] [CrossRef]
- Macreadie, I.G.; Johnson, G.; Schlosser, T.; Macreadie, P.I. Growth inhibition of Candida species and Aspergillus fumigatus by statins. FEMS Microbiol. Lett. 2006, 262, 9–13. [Google Scholar] [CrossRef]
- Larsen, T.O.; Lange, L.; Schnorr, K.; Stender, S.; Frisvad, J.C. Solistatinol, a novel phenolic compactin analogue from Penicillium solitum. Tetrahedron Lett. 2007, 48, 1261–1264. [Google Scholar] [CrossRef]
- Alberts, A.W.; Chen, J.; Kuron, G.; Hunt, V.; Huff, J.; Hoffman, C.; Rothrock, J.; Lopez, M.; Joshua, H.; Harris, E.; et al. Mevinolin: A highly potent competitive inhibitor of hydroxymethylglutaryl-coenzyme A reductase and a cholesterol-lowering agent. Proc. Natl. Acad. Sci. USA 1980, 77, 3957–61. [Google Scholar] [CrossRef]
- Endo, A.; Monacolin, K. A new hypocholesterolemic agent produced by a Monascus speicies. J. Antibiot. 1979, 32, 852–854. [Google Scholar] [CrossRef]
- Sharma, S.; Hulcher, F.; Dodge, W. Effects of cholesterol synthesis inhibitor on the proliferation of avian granulocytic and monocytic progenitor cells and primary acute monocytic leukemia cells. J. Nutr. 1984, 2, 65–69. [Google Scholar]
- Newman, A.; Clutterbuck, R.D.; Powles, R.L.; Catovsky, D.; Millar, J.L. A comparison of the effect of the 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase inhibitors simvastatin, lovastatin and pravastatin on leukaemic and normal bone marrow progenitors. Leukemia Lymphoma 1997, 24, 533–537. [Google Scholar] [CrossRef]
- Maksimova, E.; Yie, T.-A.; Rom, W.N. In vitro mechanisms of lovastatin on lung cancer cell lines as a potential chemopreventive agent. Lung 2008, 186, 45–54. [Google Scholar] [CrossRef]
- Martirosyan, A.; Clendening, J.W.; Goard, C.A.; Penn, L.Z. Lovastatin induces apoptosis of ovarian cancer cells and synergizes with doxorubicin: potential therapeutic relevance. BMC Cancer 2010, 10, 103. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Al-Abd, A.M.; Lightfoot, D.A.; El-Shemy, H.A. Anti-cancer characteristics of mevinolin against three different solid tumor cell lines was not solely p53-dependent. J. Enzym. Inhib. Med. Ch. 2012, 27, 673–679. [Google Scholar] [CrossRef]
- Relja, B.; Meder, F.; Wilhelm, K.; Henrich, D.; Marzi, I.; Lehnert, M. Simvastatin inhibits cell growth and induces apoptosis and G0/G1 cell cycle arrest in hepatic cancer cells. Int. J. Mol. Med. 2010, 26, 735–741. [Google Scholar]
- Glynn, S.A.; O’Sullivan, D.; Eustace, A.J.; Clynes, M.; O’Donovan, N. The 3-hydroxy-3-methylglutaryl-coenzyme A reductase inhibitors, simvastatin, lovastatin and mevastatin inhibit proliferation and invasion of melanoma cells. BMC Cancer 2008, 8, 9. [Google Scholar] [CrossRef]
- Osmak, M. Statins and cancer: Current and future prospects. Cancer Lett. 2012, 324, 1–12. [Google Scholar] [CrossRef]
- Ghisalberti, E.L.; Narbey, M.J.; Rowland, C.Y. Metabolites of Aspergillus terreus antagonistic towards the take-all fungus. J. Nat. Prod. 1990, 53, 520–522. [Google Scholar] [CrossRef]
- Raistrick, H.; Smith, G. LXXI. Studies in the biochemistry of micro-organisms. XLII. The metabolic products of Aspergillus terreus Thom. A new mould metabolic product-terrein. Biochem. J. 1935, 29, 606–611. [Google Scholar]
- Liao, W.-Y.; Shen, C.-N.; Lin, L.-H.; Yang, Y.-L.; Han, H.-Y.; Chen, J.-W.; Kuo, S.-C.; Wu, S.-H.; Liaw, C.-C. Asperjinone, a nor-neolignan, and terrein, a suppressor of ABCG2-expressing breast cancer cells, from thermophilic Aspergillus terreus. J. Nat. Prod. 2012, 75, 630–635. [Google Scholar] [CrossRef]
- Härri, E.; Loeffler, W.; Sigg, H.P.; Stähelin, H.; Tamm, C. Über die isolierung neuer stoffwechselprodukte aus Penicillium brefeldianum. Helv. Chim. Acta 1963, 46, 1235–1246. [Google Scholar] [CrossRef]
- Betina, V.; Nemec, P.; Dobias, J.; Barath, Z. Cyanein, a new antibiotic from Penicillium cyaneum. Folia Biol. 1962, 7, 353–357. [Google Scholar]
- Singleton, V.; Bohonos, N.; Ullstrup, A. Decumbin, a new compound from a species of Penicillium. Nature 1958, 181, 1072–1073. [Google Scholar] [CrossRef]
- Shao, R.G.; Shimizu, T.; Pommier, Y. Brefeldin A is a potent inducer of apoptosis in human cancer cells independently of p53. Exp. Cell Res. 1996, 227, 190–196. [Google Scholar] [CrossRef]
- Chapman, J.R.; Tazaki, H.; Mallouh, C.; Konno, S. Brefeldin A-induced apoptosis in prostatic cancer DU-145 cells: A possible p53-independent death pathway. BJU Int. 1999, 83, 703–708. [Google Scholar]
- Wang, J.; Huang, Y.; Fang, M.; Zhang, Y.; Zheng, Z.; Zhao, Y.; Su, W. Brefeldin A, a cytotoxin produced by Paecilomyces sp. and Aspergillus clavatus isolated from Taxus mairei and Torreya grandis. FEMS Imunol. Med. Mic. 2002, 34, 51–57. [Google Scholar] [CrossRef]
- Chinworrungsee, M.; Wiyakrutta, S.; Sriubolmas, N.; Chuailua, P.; Suksamrarn, A. Cytotoxic activities of trichothecenes isolated from an endophytic fungus belonging to order Hypocreales. Archi. Pharmacal Res. 2008, 31, 611–616. [Google Scholar] [CrossRef]
- Argoudelia, A.D.; Zieserl, J.F. The structure of U-13,933, a new antibiotic. Tetrahedron Lett. 1966, 7, 1969–1973. [Google Scholar] [CrossRef]
- He, L.; Nan, M.-H.; Oh, H.C.; Kim, Y.H.; Jang, J.H.; Erikson, R.L.; Ahn, J.S.; Kim, B.Y. Asperlin induces G2/M arrest through ROS generation and ATM pathway in human cervical carcinoma cells. Biochem. Bioph. Res. Co. 2011, 409, 489–493. [Google Scholar] [CrossRef]
- Brian, P.W. Studies on the biological activity of griseofulvin. Ann. Bot. 1949, 13, 59–77. [Google Scholar]
- Hector, R.F. An overview of antifungal drugs and their use for treatment of deep and superficial mycoses in animals. Clin. Tech. Small Anim. Pract. 2005, 20, 240–249. [Google Scholar] [CrossRef]
- Oxford, A.E.; Raistrick, H.; Simonart, P. XXIX. Studies in the biochemistry of microorganisms. LX. Griseofulvin, C17H17O6Cl, a metabolic product of Penicillium griseofulvum Dierckx. Bioch. J. 1939, 33, 240–248. [Google Scholar]
- Grisham, L.M.; Wilson, L.; Bensch, K.G. Antimitotic action of griseofulvin does not involve disruption of microtubules. Nature 1973, 224, 294–296. [Google Scholar]
- Stierle, D.B.; Stierle, A.A.; Bugni, T. Sequoiamonascins A–D: Novel anticancer metabolites isolated from a redwood endophyte. J. Org. Chem. 2003, 68, 4966–4969. [Google Scholar] [CrossRef]
- De Stefano, S.; Nicoletti, R.; Milone, A.; Zambardino, S. 3-O-Methylfunicone, a fungitoxic metabolite produced by the fungus Penicillium pinophilum. Phytochemistry 1999, 52, 1399–1401. [Google Scholar] [CrossRef]
- Stammati, A.; Nicoletti, R.; De Stefano, S.; Zampaglioni, F.; Zucco, F. Cytostatic properties of a novel compound derived from Penicillium pinophilum: An in vitro study. Altern. Lab. Anim. 2002, 30, 69–75. [Google Scholar]
- Buommino, E.; Nicoletti, R.; Gaeta, G.M.; Orlando, M.; Ciavatta, M.L.; Baroni, A.; Tufano, M.A. 3-O-methylfunicone, a secondary metabolite produced by Penicillium pinophilum, induces growth arrest and apoptosis in HeLa cells. Cell Proliferat. 2004, 37, 413–426. [Google Scholar] [CrossRef]
- Buommino, E.; Boccellino, M.; de Filippis, A.; Petrazzuolo, M.; Cozza, V.; Nicoletti, R.; Ciavatta, M.L.; Quagliuolo, L.; Tufano, M.A. 3-O-methylfunicone produced by Penicillium pinophilum affects cell motility of breast cancer cells, downregulating avb5 integrin and inhibiting metalloproteinase-9 secretion. Mol. Carcinogen. 2007, 46, 930–940. [Google Scholar] [CrossRef]
- Baroni, A.; de Luca, A.; de Filippis, A.; Petrazzuolo, M.; Manente, L.; Nicoletti, R.; Tufano, M.A.; Buommino, E. 3-O-methylfunicone, a metabolite of Penicillium pinophilum, inhibits proliferation of human melanoma cells by causing G(2)+M arrest and inducing apoptosis. Cell Proliferat. 2009, 42, 541–553. [Google Scholar] [CrossRef]
- Buommino, E.; Tirino, V.; de Filippis, A.; Silvestri, F.; Nicoletti, R.; Ciavatta, M.L.; Pirozzi, G.; Tufano, M. A 3-O-methylfunicone, from Penicillium pinophilum, is a selective inhibitor of breast cancer stem cells. Cell Proliferat. 2011, 44, 401–409. [Google Scholar] [CrossRef]
- Komai, S.-I.; Hosoe, T.; Itabashi, T.; Nozawa, K.; Yaguchi, T.; Fukushima, K.; Kawai, K.-I. New vermistatin derivatives isolated from Penicillium simplicissimum. Heterocycles 2005, 65, 2771–2776. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B.; Girtsman, T. Caspase-1 inhibitors from an extremophilic fungus that target specific leukemia cell lines. J. Nat. Prod. 2012, 75, 344–350. [Google Scholar] [CrossRef]
- Qin, J.-C.; Zhang, Y.-M.; Gao, J.-M.; Bai, M.-S.; Yang, S.-X.; Laatsch, H.; Zhang, A.-L. Bioactive metabolites produced by Chaetomium globosum, an endophytic fungus isolated from Ginkgo biloba. Bioorgan. Med. Chem. Lett. 2009, 19, 1572–1574. [Google Scholar] [CrossRef]
- Yamada, T.; Doi, M.; Shigeta, H.; Muroga, Y.; Hosoe, S.; Numata, A.; Tanaka, R. Absolute stereostructures of cytotoxic metabolites, chaetomugilins A–C, produced by a Chaetomium species separated from a marine fish. Tetrahedron Lett. 2008, 49, 4192–4195. [Google Scholar] [CrossRef]
- Yasuhide, M.; Yamada, T.; Numata, A.; Tanaka, R. Chaetomugilins, new selectively cytotoxic metabolites, produced by a marine fish-derived Chaetomium species. J. Antibiot. 2008, 61, 615–622. [Google Scholar] [CrossRef]
- Yamada, T.; Yasuhide, M.; Shigeta, H.; Numata, A.; Tanaka, R. Absolute stereostructures of chaetomugilins G and H produced by a marine-fish-derived Chaetomium species. J. Antibiot. 2009, 62, 353–357. [Google Scholar] [CrossRef]
- Muroga, Y.; Yamada, T.; Numata, A.; Tanaka, R. Chaetomugilins I–O, new potent cytotoxic metabolites from a marine-fish-derived Chaetomium species. Stereochemistry and biological activities. Tetrahedron 2009, 65, 7580–7586. [Google Scholar] [CrossRef]
- Muroga, Y.; Yamada, T.; Numata, A.; Tanaka, R. 11- and 4’-epimers of chaetomugilin A, novel cytostatic metabolites from marine fish-derived fungus Chaetomium globosum. Helv. Chim. Acta 2010, 93, 542–549. [Google Scholar] [CrossRef]
- Yamada, T.; Muroga, Y.; Jinno, M.; Kajimoto, T.; Usami, Y.; Numata, A.; Tanaka, R. New class azaphilone produced by a marine fish-derived Chaetomium globosum. The stereochemistry and biological activities. Bioorgan. Med. Chem. 2011, 19, 4106–4113. [Google Scholar] [CrossRef]
- Yamada, T.; Jinno, M.; Kikuchi, T.; Kajimoto, T.; Numata, A.; Tanaka, R. Three new azaphilones produced by a marine fish-derived Chaetomium globosum. J. Antibiot. 2012, 65, 413–417. [Google Scholar] [CrossRef]
- Chen, G.-D.; Li, Y.-J.; Gao, H.; Chen, Y.; Li, X.-X.; Li, J.; Guo, L.-D.; Cen, Y.-Z.; Yao, X.-S. New azaphilones and chlorinated phenolic glycosides from Chaetomium elatum with caspase-3 inhibitory activity. Planta Med. 2012, 78, 1683–1689. [Google Scholar] [CrossRef]
- Lee, L.; Bennett, J.W.; Goldblatt, L.A.; Lundin, R.E. Norsolorinic acid from a mutant strain of Aspergillus parasiticus. J. Am. Oil Chemi. Soc. 1971, 48, 93–94. [Google Scholar] [CrossRef]
- Bennett, J.W.; Lee, L.S.; Vinnett, C. The correlation of aflatoxin and norsolorinic acid production. J. Am. Oil Chem. Soc. 1971, 48, 368–370. [Google Scholar] [CrossRef]
- Wang, C.C.C.; Chiang, Y.-M.; Kuo, P.-L.; Chang, J.-K.; Hsu, Y.-L. Norsolorinic acid inhibits proliferation of T24 human bladder cancer cells by arresting the cell cycle at the G0/G1 phase and inducing a Fas/membrane-bound Fas ligand-mediated apoptotic pathway. Clini. Exp. Pharmacol. P. 2008, 35, 1301–1308. [Google Scholar] [CrossRef]
- Wang, C.C.C.; Chiang, Y.-M.; Kuo, P.-L.; Chang, J.-K.; Hsu, Y.-L. Norsolorinic acid from Aspergillus nidulans inhibits the proliferation of human breast adenocarcinoma MCF-7 cells via Fas-mediated pathway. Basic Clin. Pharmacol. 2008, 102, 491–497. [Google Scholar] [CrossRef]
- Samson, R.A.; Varga, J.; Meijer, M.; Frisvad, J.C. New taxa in Aspergillus section. Usti. Stud. Mycol. 2011, 69, 81–97. [Google Scholar] [CrossRef]
- Steyn, P.S.; Vleggaar, R. Austocystins. Six novel dihydrofuro (3',2':4,5)furo(3,2-b)xanthenones from Aspergillus ustus. J. Chem. Soc. Perk. T. 1 1974, 1974, 2250–2256. [Google Scholar]
- Ireland, C.; Aalbersberg, W.; Andersen, R.; Ayral-Kaloustian, S.; Berlinck, R.; Bernan, V.; Carter, G.; Churchill, A.; Clardy, J.; Concepcion, G.; et al. Anticancer agents from unique natural products sources. Pharm. Biol. 2003, 41, 15–38. [Google Scholar] [CrossRef]
- Marks, K.M.; Park, E.S.; Arefolov, A.; Russo, K.; Ishihara, K.; Ring, J.E.; Clardy, J.; Clarke, A.S.; Pelish, H.E. The selectivity of austocystin D arises from cell-line-specific drug activation by cytochrome P450 enzymes. J. Nat. Prod. 2011, 74, 567–573. [Google Scholar] [CrossRef]
- Houbraken, J.; Frisvad, J.C.; Samson, R. A. Fleming’s penicillin producing strain is not Penicillium chrysogenum but P. rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef]
- Houbraken, J.; Frisvad, J.C.; Seifert, K.A.; Overy, D.P.; Tuthill, D.M.; Valdez, J.G.; Samson, R.A. New penicillin-producing Penicillium species and an overview of section Chrysogena. Persoonia 2012, 29, 78–100. [Google Scholar] [CrossRef]
- Liu, W.; Gu, Q.; Zhu, W.; Cui, C.; Fan, G.; Zhu, T.; Liu, H.; Fang, Y. Chloctanspirones A and B, novel chlorinated polyketides with an unprecedented skeleton, from marine sediment derived fungus Penicillium terrestre. Tetrahedron Lett. 2005, 46, 4993–4996. [Google Scholar]
- Li, D.; Chen, L.; Zhu, T.; Kurtán, T.; Mándi, A.; Zhao, Z.; Li, J.; Gu, Q. Chloctanspirones A and B, novel chlorinated polyketides with an unprecedented skeleton, from marine sediment derived fungus Penicillium terrestre. Tetrahedron 2011, 67, 7913–7918. [Google Scholar] [CrossRef]
- Takahashi, C.; Yoshihira, K.; Natori, S.; Umeda, M. The structures of toxic metabolites of Aspergillus candidus. I. The compounds A and E, cytotoxic p-terphenyls. Chem. Pharm. Bull. 1976, 24, 613–620. [Google Scholar]
- Wei, H.; Inada, H.; Hayashi, A.; Higashimoto, K.; Pruksakorn, P.; Kamada, S.; Arai, M.; Ishida, S. Prenylterphenyllin and its dehydroxyl analogs, new cytotoxic substances from a marine-derived fungus Aspergillus candidus IF10. J. Antibiot. 2007, 60, 586–590. [Google Scholar] [CrossRef]
- Cai, S.; Sun, S.; Zhou, H.; Kong, X.; Zhu, T.; Li, D.; Gu, Q. Prenylated polyhydroxy-p-terphenyls from Aspergillus taichungensis ZHN-7-07. J. Nat. Prod. 2011, 75, 1106–1110. [Google Scholar]
- Du, L.; Liu, H.-C.; Fu, W.; Li, D.-H.; Pan, Q.-M.; Zhu, T.-J.; Geng, M.-Y.; Gu, Q.-Q. Unprecedented citrinin trimer tricitinol B functions as a novel topoisomerase IIα inhibitor. J. Med. Chem. 2011, 54, 5796–5810. [Google Scholar] [CrossRef]
- Mulrooney, C.A.; O’Brien, E.M.; Morgan, B.J.; Kozlowski, M.C. Perylenequinones: Isolation, synthesis, and biological activity. Eur. J. Org. Chem. 2012, 2012, 3887–3904. [Google Scholar] [CrossRef]
- Kobayashi, E.; Ando, K.; Nakano, H.; Iida, T.; Ohno, H.; Morimoto, M.; Tamaoki, T. Calphostins (UCN-1028), novel and specific inhibitors of protein kinase C. I. Fermentation, isolation, physico-chemical properties and biological activities. J. Antibiot. 1989, 42, 1470–1474. [Google Scholar]
- Zhu, D.M.; Narla, R.K.; Fang, W.H.; Chia, N.C.; Uckun, F.M. Calphostin C triggers calcium-dependent apoptosis in human acute lymphoblastic leukemia cells. Clini. Cancer Res. 1998, 4, 2967–2976. [Google Scholar]
- Wu, H.; Lao, X.-F.; Wang, Q.-W.; Lu, R.-R. The shiraiachromes: novel fungal perylenequinone pigments from Shiraia bambusicola. J. Nat. Prod. 1989, 52, 948–951. [Google Scholar] [CrossRef]
- Fang, L.; Qing, C.; Shao, H.; Yang, Y.; Dong, Z.; Wang, F.; Zhao, W.; Yang, W.; Liu, J. Hypocrellin D, a cytotoxic fungal pigment from fruiting bodies of the ascomycete Shiraia bambusicola. J. Antibiot. 2006, 59, 351–354. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, L.; Xie, J.; Qiu, H.; Gu, Y.; Zhao, J. Novel surfactant-like hypocrellin derivatives to achieve simultaneous drug delivery in blood plasma and cell uptake. Photochem. Photobiol. 2010, 86, 667–672. [Google Scholar] [CrossRef]
- Li, G.; Wang, H.; Zhu, R.; Sun, L.; Wang, L.; Li, M.; Li, Y.; Liu, Y.; Zhao, Z.; Lou, H. Phaeosphaerins A−F, cytotoxic perylenequinones from an endolichenic fungus, Phaeosphaeria sp. J. Nat. Prod. 2012, 75, 142–147. [Google Scholar] [CrossRef]
- Gao, X.; Chooi, Y.-H.; Ames, B.D.; Wang, P.; Walsh, C.T.; Tang, Y. Fungal indole alkaloid biosynthesis: Genetic and biochemical investigation of the tryptoquialanine pathway in Penicillium aethiopicum. J. Am. Chem. Soc. 2011, 133, 2729–2741. [Google Scholar]
- Shang, Z.; Li, X.; Meng, L.; Li, C.; Gao, S.; Huang, C.; Wang, B. Chemical profile of the secondary metabolites produced by a deep-sea sediment-derived fungus Penicillium commune SD-118. Chin. J. Oceanol. Limn. 2012, 30, 305–314. [Google Scholar] [CrossRef]
- Varga, J.; Due, M.; Frisvad, J.C.; Samson, R.A. Taxonomic revision of Aspergillus section Clavati based on molecular, morphological and physiological data. Stud. Mycol. 2007, 59, 89–106. [Google Scholar] [CrossRef]
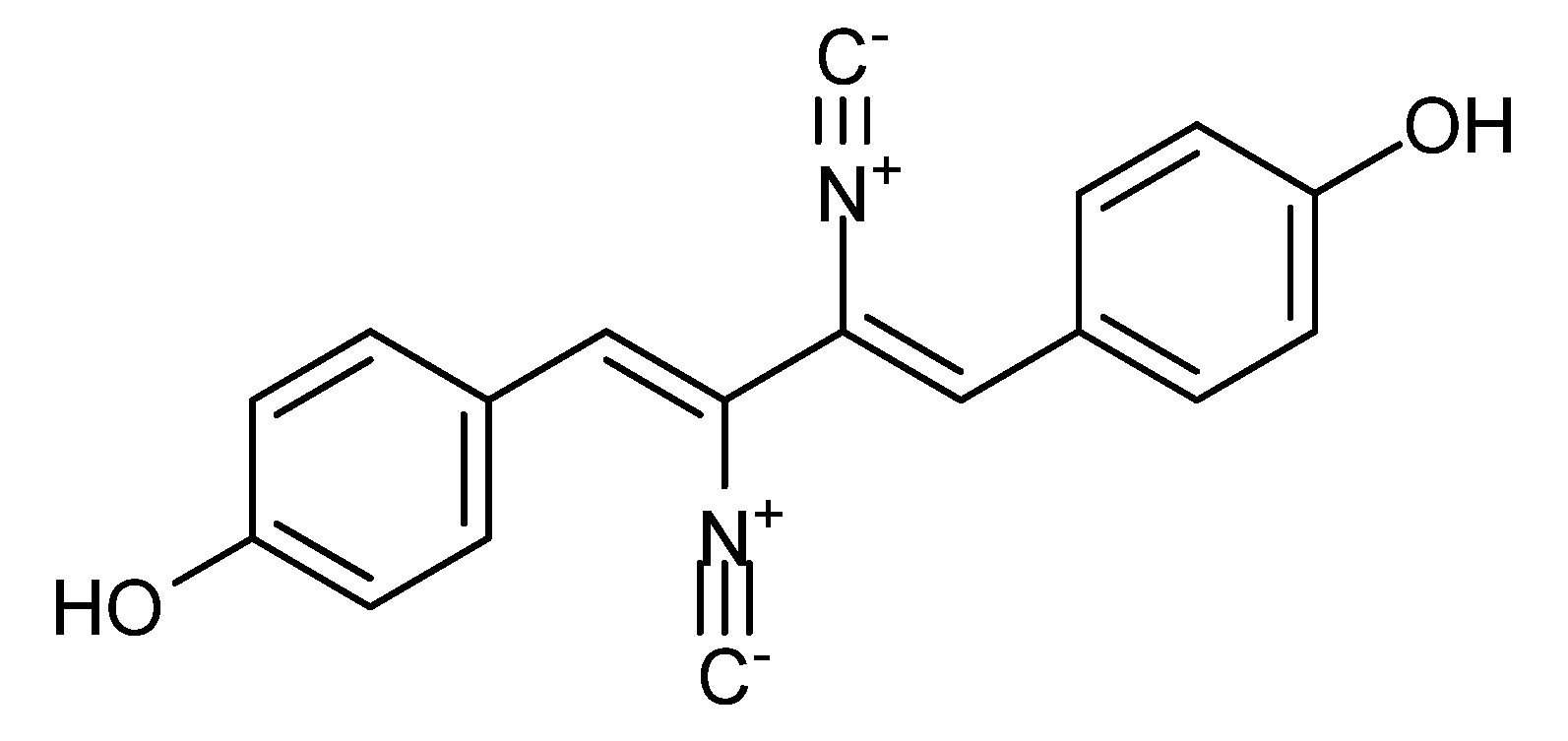
- Ando, K.; Suzuki, S.; Takatsuki, A.; Arima, K.; Tamura, G. A new antibiotic, 1-(p-hydroxyphenyl)-2,3-diisocyano-4-(p-methoxyphenyl)-buta-1,3-diene. I. Isolation and biological properties. J. Antibiot. 1968, 21, 582–586. [Google Scholar] [CrossRef]
- Kozlovskiĭ, A.G.; Zhelifonova, V.P.; Antipova, T.V; Adanin, V.M.; Novikova, N.D.; Deshevaia, E.A.; Schlegel, B.; Dahse, H.M.; Gollmick, F.A.; Grafe, U. Penicillium expansum, a resident fungal strain of the orbital complex Mir, producing xanthocillin X and questiomycin A. Prikl. Biokhim. Mikrobiol. 2004, 40, 344–349. [Google Scholar]
- Tsunkawa, M.; Ohkusa, N.; Kobaru, S.; Narita, Y.; Mirate, S.; Sawada, Y.; Oki, T. BU-4704, a new member of the xanthocillin class. J. Antibiot. 1993, 46, 687–688. [Google Scholar] [CrossRef]
- Zhan, X.; Zhao, H.; Guan, Y.; Xiao, C.; He, Q. Mechanism of inhibiting proliferation by xanthocillin X dimethyl in tumor cells. Zhongguo Xinyao Zazhi 2010, 19, 832–836. [Google Scholar]
- Clark, A.M.; Hufford, C.D.; Robertson, L.W. Two metabolites from Aspergillus flavipes. Lloydia 1977, 40, 146–151. [Google Scholar]
- Frisvad, J.C.; Houbraken, J.; Popma, S.; Samson, R.A. Two new Penicillium species Penicillium buchwaldii and Penicillium spathulatum, producing the anticancer compound asperphenamate. FEMS microbiol. Lett. 2013, 339, 77–92. [Google Scholar] [CrossRef]
- Wu, P.-L.; Lin, F.-W.; Wu, T.-S.; Kuoh, C.-S.; Lee, K.-H.; Lee, S.-J. Cytotoxic and anti-HIV principles from the rhizomes of Begonia nantoensis. Chem. Pharm. Bull. 2004, 52, 345–349. [Google Scholar]
- Yuan, L.; Wang, J.H.; Sun, T.M. Total synthesis and anticancer activity studies of the stereoisomers of asperphenamate and patriscabratine. Chin. Chem. Lett. 2010, 21, 155–158. [Google Scholar] [CrossRef]
- Li, Y.; Luo, Q.; Yuan, L.; Miao, C.; Mu, X.; Xiao, W.; Li, J.; Sun, T.; Ma, E. JNK-dependent Atg4 upregulation mediates asperphenamate derivative BBP-induced autophagy in MCF-7 cells. Toxicol. Appl. Pharm. 2012, 263, 21–31. [Google Scholar] [CrossRef]
- Borthwick, A.D. 2,5-Diketopiperazines: Synthesis, reactions, medicinal chemistry, and bioactive natural products. Chem. Rev. 2012, 112, 3641–3716. [Google Scholar] [CrossRef]
- Yamazaki, M.; Suzuki, S.; Miyaki, K. Tremorgenic toxins from Aspergillus fumigatus Fres. Chem. Pharm. Bull. 1971, 19, 1739–1740. [Google Scholar] [CrossRef]
- Abraham, W.-R.; Arfmann, H. 12,13-Dihydroxy-fumitremorgin C from Aspergillus fumigatus. Phytochemistry 1990, 29, 1025–1026. [Google Scholar] [CrossRef]
- Samson, R.A.; Hong, S.; Peterson, S.W.; Frisvad, J.C.; Varga, J. Polyphasic taxonomy of Aspergillus section Fumigati and its teleomorph Neosartorya. Stud. Mycol. 2007, 59, 147–203. [Google Scholar] [CrossRef]
- Cui, C.-B.; Kakeya, H.; Okada, G.; Onose, R.; Ubukata, M.; Takahashi, I.; Isono, K.; Osada, H. Tryprostatins A and B, novel mammalian cell cycle inhibitors produced by Aspergillus fumigatus. J. Antibiot. 1995, 48, 1382–1384. [Google Scholar] [CrossRef]
- Cui, C.; Kakeya, H.; Osada, H. Novel mammalian cell cycle inhibitors, spirotryprostatins A and B, produced by Aspergillus fumigatus, which inhibit mammalian cell cycle at G2/M phase. Tetrahedron 1996, 52, 12651–12666. [Google Scholar] [CrossRef]
- Qian-Cutrone, J.; Huang, S.; Shu, Y.-Z.; Vyas, D.; Fairchild, C.; Menendez, A.; Krampitz, K.; Dalterio, R.; Klohr, S.E.; Gao, Q. Stephacidin A and B: two structurally novel, selective inhibitors of the testosterone-dependent prostate LNCaP cells. J. Am. Chem. Soc. 2002, 124, 14556–14557. [Google Scholar] [CrossRef]
- Finefield, J.M.; Frisvad, J.C.; Sherman, D.H.; Williams, R.M. Fungal origins of the bicyclo[2.2.2]diazaoctane ring system of prenylated indole alkaloids. J. Nat. Prod. 2012, 75, 812–833. [Google Scholar]
- Zhao, S.; Smith, K.S.; Deveau, A.M.; Dieckhaus, C.M.; Johnson, M.A.; Macdonald, T.L.; Cook, J.M. Biological activity of the tryprostatins and their diastereomers on human carcinoma cell lines. J. Med. Chem. 2002, 45, 1559–1562. [Google Scholar] [CrossRef]
- Wang, F.; Fang, Y.; Zhu, T.; Zhang, M.; Lin, A.; Gu, Q.; Zhu, W. Seven new prenylated indole diketopiperazine alkaloids from holothurian-derived fungus Aspergillus fumigatus. Tetrahedron 2008, 64, 7986–7991. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.-L.; Fang, Y.-C.; Zhu, T.-J.; Gu, Q.-Q.; Zhu, W.-M. Cytotoxic alkaloids and antibiotic nordammarane triterpenoids from the marine-derived fungus Aspergillus sydowi. J. Nat. Prod. 2008, 71, 985–989. [Google Scholar] [CrossRef]
- Numata, A.; Takahashi, C.; Miyamoto, T.; Matsushita, T.; Kawai, K.; Usami, Y.; Matsumura, E.; Inoue, M.; Ohishi, H.; Shingu, T. Structures of cytotoxic substances and new quinazoline derivatives produced by a fungus from a saltwater fish. Tennen Yuki Kagobutsu Toronkai Koen Yoshishu 1991, 33, 723–730. [Google Scholar]
- Wang, Y.; Li, Z.-L.; Bai, J.; Zhang, L.-M.; Wu, X.; Zhang, L.; Pei, Y.-H.; Jing, Y.-K.; Hua, H.-M. 2,5-diketopiperazines from the marine-derived fungus Aspergillus fumigatus YK-7. Chem. Biodivers. 2012, 9, 385–393. [Google Scholar] [CrossRef]
- Rabindran, S.K.; He, H.; Singh, M.; Brown, E.; Collins, K.I.; Annable, T.; Greenberger, L.M. Reversal of a novel multidrug resistance mechanism in human colon carcinoma cells by fumitremorgin C. Cancer Res. 1998, 1998, 5850–5858. [Google Scholar]
- Rho, M.C.; Hayashi, M.; Fukami, A.; Obata, R.; Sunazuka, T.; Tomoda, H.; Komiyama, K.; Omura, S. Reversal of multidrug resistance by 7-O-benzoylpyripyropene A in multidrug-resistant tumor cells. J. Antibiot. 2000, 53, 1201–1206. [Google Scholar] [CrossRef]
- Jurjevic, Z.; Peterson, S.W.; Horn, B.W. Aspergillus section Versicolores: nine new species and multilocus DNA sequence based phylogeny. IMA Fungus 2012, 3, 59–79. [Google Scholar] [CrossRef]
- Kato, H.; Yoshida, T.; Tokue, T.; Nojiri, Y.; Hirota, H.; Ohta, T.; Williams, R.M.; Tsukamoto, S. Notoamides A–D: Prenylated indole alkaloids isolated from a marine-derived fungus, Aspergillus sp. Angew. Chem. Int. Edit. 2007, 46, 2254–2256. [Google Scholar] [CrossRef]
- Kanoh, K.; Kohno, S.; Asari, T.; Harada, T.; Katada, J.; Muramatsu, M.; Kawashima, H.; Sekiya, H.; Uno, I. (−)-phenylahistin: A new mammalian cell cycle inhibitor produced by Aspergillus ustus. Bioorg. Med. Chem. Lett. 1997, 7, 2847–2852. [Google Scholar] [CrossRef]
- Kanoh, K.; Kohno, S.; Katada, J.; Hayashi, Y.; Muramatsu, M.; Uno, I. Antitumor activity of phenylahistin in vitro and in vivo. Biosci. 1999, 63, 1130–1133. [Google Scholar]
- Yamazaki, Y.; Tanaka, K.; Nicholson, B.; Deyanat-Yazdi, G.; Potts, B.; Yoshida, T.; Oda, A.; Kitagawa, T.; Orikasa, S.; Kiso, Y.; et al. Synthesis and structure−Activity relationship study of antimicrotubule agents phenylahistin derivatives with a didehydropiperazine-2,5-dione structure. J. Med. Chem. 2012, 55, 1056–1071. [Google Scholar] [CrossRef]
- Hayashi, Y.; Orikasa, S.; Tanaka, K.; Kanoh, K.; Kiso, Y. Total synthesis of anti-microtubule diketopiperazine derivatives: phenylahistin and aurantiamine. J. Org. Chem. 2000, 65, 8402–8405. [Google Scholar] [CrossRef]
- Larsen, T.O.; Frisvad, J.C.; Jensen, S.R. Aurantiamine, a diketopiperazine from two varieties of Penicillium aurantiogriseum. Phytochemistry 1992, 31, 1613–1615. [Google Scholar] [CrossRef]
- Menzel, A.O.; Wintersteiner, O.; Hoogerheide, J.O. The isolation of gliotoxin and fumigacin from culture filtrates of Aspergillus fumigatus. J. Biol. Chem. 1944, 152, 419–429. [Google Scholar]
- Kaouadji, M. Gliotoxin: Uncommon 1H couplings and revised 1H- and 13C-NMR assignments. J. Nat. Prod. 1990, 53, 717–719. [Google Scholar] [CrossRef]
- Carberry, S.; Molloy, E.; Hammel, S.; O’Keeffe, G.; Jones, G.W.; Kavanagh, K.; Doyle, S. Gliotoxin effects on fungal growth: Mechanisms and exploitation. Fungal Genet. Biol. 2012, 49, 302–312. [Google Scholar] [CrossRef]
- Kidd, J.G. Effects of an antibiotic from Aspergillus fumigatus Fresenius on tumor cells in vitro, and its possible identity with gliotoxin. Science 1947, 105, 511–513. [Google Scholar]
- Vigushin, D.M.; Mirsaidi, N.; Brooke, G.; Sun, C.; Pace, P.; Inman, L.; Moody, C.J.; Coombes, R.C. Gliotoxin is a dual inhibitor of farnesyltransferase and geranylgeranyltransferase I with antitumor activity against breast cancer in vivo. Med. Mycol. 2004, 21, 21–30. [Google Scholar]
- Seya, H.; Nozawa, K.; Nakajima, S.; Kawai, K.-I.; Udagawa, S.-I. Studies on fungal products. Part 8. Isolation and structure of emestrin, a novel antifungal macrocyclic epidithiodioxopiperazine from Emericeella striata. X-Ray molecular structure of emestrin. J. Chem. Soc. Perk. T. 1 1986, 1986, 109–116. [Google Scholar]
- Seya, H.; Nakajima, S.; Kawai, K.-I.; Udagawa, S.-I. Structure and absolute configuration of emestrin, a new macrocyclic epidithiodioxopiperazine from Emericella striata. J. Chem. Soc. Chem. Comm. 1985, 739, 657–658. [Google Scholar]
- Ueno, Y.; Umemori, K.; Nilmi, E.; Tanuma, S.; Nagata, S.; Sugamata, M.; Ihara, T.; Sekljlma,, M.; Kawai, K.-I.; Ueno, I.; Takhiro, F. Induction of apoptosis by T-2 toxin and other natural toxins in HL-60 human promyelotic leukemia cells. Nat. Toxins 1995, 3, 129–137. [Google Scholar]
- Onodera, H.; Hasegawa, A.; Tsumagari, N.; Nakai, R.; Ogawa, T.; Kanda, Y. MPC1001 and its analogues: New antitumor agents from the fungus Cladorrhinum species. Org. Lett. 2004, 6, 4101–4104. [Google Scholar] [CrossRef]
- Choi, E.J.; Park, J.-S.; Kim, Y.-J.; Jung, J.-H.; Lee, J.K.; Kwon, H.C.; Yang, H.O. Apoptosis-inducing effect of diketopiperazine disulfides produced by Aspergillus sp. KMD 901 isolated from marine sediment on HCT116 colon cancer cell lines. J. Appl. Microbiol. 2011, 110, 304–313. [Google Scholar] [CrossRef]
- Arai, T.; Mikami, Y.; Fukushima, K.; Utsumi, T.; Yazawa, K. A new antibiotic, leucinostatin, derived from Penicillium lilacinum. J. Antibiot. 1973, 26, 157–161. [Google Scholar] [CrossRef]
- Luangsa-Ard, J.; Houbraken, J.; van Doorn, T.; Hong, S.-B.; Borman, A.M.; Hywel-Jones, N.L.; Samson, R.A. Purpureocillium, a new genus for the medically important Paecilomyces lilacinus. FEMS microbiol. Lett. 2011, 321, 141–149. [Google Scholar] [CrossRef]
- Fukushima, K.; Arai, T.; Mori, Y.; Tsuboi, M.; Suzuki, M. Studies on peptide antibiotics, leucinostatins. I. Separation, physico- chemical properties and biological activities of leucinostatins A and B. J. Antibiot. 1983, 36, 1606–1612. [Google Scholar]
- Strobel, G.A.; Torczynski, R.; Bollon, A. Acremonium sp.—A leucinostatin A producing endophyte of European yew (Taxus baccata). Plant Sci. 1997, 128, 97–108. [Google Scholar] [CrossRef]
- Ishiguro, K.; Arai, T. Action of the peptide antibiotic leucinostatin. Antimicrobi. Agents Chemither. 1976, 9, 893–898. [Google Scholar] [CrossRef]
- Kawada, M.; Inoue, H.; Ohba, S.-I.; Masuda, T.; Momose, I.; Ikeda, D. Leucinostatin A inhibits prostate cancer growth through reduction of insulin-like growth factor-I expression in prostate stromal cells. Int. J. Cancer 2010, 126, 810–818. [Google Scholar]
- Strobel, G.A.; Hess, W.M. Glucosylation of the peptide leucinostatin A, produced by an endophytic fungus of European yew, may protect the host from leucinostatin toxicity. Chem. Biol. 1997, 4, 529–536. [Google Scholar] [CrossRef]
- Wani, M.C.; Taylor, L.H.; Wall, M.E.; Coggon, P.; McPhail, A.T. Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar]
- Niedens, B.R.; Parker, S.R.; Stierle, D.B.; Stierle, A.A. First fungal aromatic L-amino acid decarboxylase from a paclitaxel-producing Penicillium raistrickii. Mycologia 2013, 91, 619–626. [Google Scholar]
- Kohler, D.R.; Goldspiel, B.R. Paclitaxel (taxol). Pharmacotherapy 1994, 14, 3–34. [Google Scholar]
- Visalakchi, S.; Muthumary, J. Taxol (anticancer drug) producing endophytic fungi: An overview. Int. J. Pharm. Bio Sci. 2010, 1, 1–9. [Google Scholar]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of microtubule assembly in vitro by taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef]
- Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef]
- Young, D.H.; Michelotti, E.L.; Swindell, C.S.; Krauss, N.E. Antifungal properties of taxol and various analogues. Experientia 1992, 48, 882–885. [Google Scholar] [CrossRef]
- Pittayakhajonwut, P.; Dramae, A.; Intaraudom, C.; Boonyuen, N.; Nithithanasilp, S.; Rachtawee, P.; Laksanacharoen, P. Two new drimane sesquiterpenes, fudecadiones A and B, from the soil fungus Penicillium sp. BCC 17468. Planta Med. 2011, 77, 74–76. [Google Scholar] [CrossRef]
- Hirota, A.; Nakagawa, M.; Hirota, H.; Takahashi, T. Terrecylic acid A, a new antibiotic from Aspergillus terreus IV. Absolute stereochemistry of terrecyclic acid A. J. Antibiot. 1986, 39, 1–4. [Google Scholar] [CrossRef]
- Wijeratne, E.M.K.; Turbyville, T.J.; Zhang, Z.; Bigelow, D.; Pierson, L.S.; VanEtten, H.D.; Whitesell, L.; Canfield, L.M.; Gunatilaka, A.A.L. Cytotoxic constituents of Aspergillus terreus from the rhizosphere of Opuntia versicolor of the Sonoran desert. J. Nat. Prod. 2003, 66, 1567–1573. [Google Scholar] [CrossRef]
- Wu, G.; Lin, A.; Gu, Q.; Zhu, T.; Li, D. Four new chloro-eremophilane sesquiterpenes from an antarctic deep-sea derived fungus, Penicillium sp. PR19N-1. Mar. Drugs 2013, 11, 1399–1408. [Google Scholar]
- Woloshuk, C.P.; Shim, W.-B. Aflatoxins, fumonisins, and trichothecenes: a convergence of knowledge. FEMS Microbiol. Rev. 2013, 37, 94–109. [Google Scholar] [CrossRef]
- Sudakin, D. Trichothecenes in the environment: relevance to human health. Toxicol. Lett. 2003, 143, 97–107. [Google Scholar] [CrossRef]
- Shifrin, V.I.; Anderson, P. Trichothecene mycotoxins trigger a ribotoxic stress response that activates c-Jun N-terminal kinase and p38 mitogen-activated protein kinase and induces apoptosis. J. Biol. Chem. 1999, 274, 13985–13992. [Google Scholar] [CrossRef]
- Campos, F.F.; Johann, S.; Cota, B.B.; Alves, T.M.A.; Rosa, L.H.; Caligiorne, R.B.; Cisalpino, P.S.; Rosa, C.A.; Zani, C.L. Antifungal activity of trichothecenes from Fusarium sp. against clinical isolates of Paracoccidioides brasiliensis. Mycoses 2011, 54, E122–E129. [Google Scholar] [CrossRef]
- Sun, T.; Zheng, W.; Peng, H.; Zhang, A.; Chen, Y.; Tan, R.; Shen, P. A small molecule IFB07188 inhibits proliferation of human cancer cells by inducing G2/M cell cycle arrest and apoptosis. Biomedi. Pharmacother. 2012, 66, 512–518. [Google Scholar] [CrossRef]
- Pae, H.O.; Oh, G.S.; Choi, B.M.; Seo, E.A.; Oh, H.; Shin, M.K.; Kim, T.H.; Kwon, T.O.; Chung, H.T. Induction of apoptosis by 4-acetyl-12,13-epoxyl-9-trichothecene-3,15-diol from Isaria japonica Yasuda through intracellular reactive oxygen species formation and caspase-3 activation in human leukemia HL-60 cells. Toxicol. in Vitro 2003, 17, 49–57. [Google Scholar] [CrossRef]
- OH, G.-S.; Hong, K.-H.; OH, H.; Pae, H.-O.; Kim, I.-K.; Kwon, T.-O.; Shin, M.-K.; Chung, H.-T. 4-Acetyl-12,13-epoxyl-9-trichothecene-3 ,15-diol isolated from the fruiting bodies of Iaria japonica YASUDA induces apoptosis of human leukemia cells (HL-60). Biol. Pharm. Bull. 2001, 24, 785–789. [Google Scholar] [CrossRef]
- Xu, J.; Takasaki, A.; Kobayashi, H.; Oda, T.; Yamada, J.; Mangindaan, R.E.P.; Ukai, K.; Nagai, H.; Namikoshi, M. Four new macrocyclic trichothecenes from two strains of marine-derived fungi of the genus Myrothecium. J. Antibiot. 2006, 59, 451–455. [Google Scholar] [CrossRef]
- Goodwin, W.; Haas, C.D.; Fabian, C.; Heller-Bettinger, I.; Hoogstraten, B. Phase I clinical evaluation of anguidine (diacetoxyscirpenol, NSC-141537). Cancer 1977, 42, 23–26. [Google Scholar]
- Dornera, J.W.; Colea, R.J.; Springerb, J.P.; Coxc, R.H.; Cutlerd, H.; Wicklow, D.T. Isolation and identification of two new biologically active norditerpene dilactones from Aspergillus wentii. Phytochemistry 1980, 19, 1157–1161. [Google Scholar] [CrossRef]
- Zhang, Z.; Miao, L.; Sun, W.; Jiao, B.; Wang, B.; Yao, L.; Huang, C. Wentilactone B from Aspergillus wentii induces apoptosis and inhibits proliferation and migration of human hepatoma SMMC-7721 cells. Bio. Pharm. Bull. 2012, 35, 1964–1971. [Google Scholar] [CrossRef]
- Brian, P.W.; Curtis, P.J.; Hemming, H.G.; Norris, G.L.F. Wortmannin, an antibiotic produced by Penicillium wortmanni. Trans. Br. Mycol. Soc. 1957, 40, 365–368. [Google Scholar] [CrossRef]
- Nakamura, M.; Nakashima, S.; Katagiri, Y.; Ajozawa, Y. Effect of wortmannin and 2-(4-Morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002) on N-formyl-methionyl-leucyl-phenylalanine-induced phospholipase D activation in differentiated HL60 Cells. Biochem. Pharmacol. 1997, 53, 1929–1936. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Q.; Zhang, L.; Wu, Y.; Shu, Y. Wortmannin induced apoptosis of leukemia cells by reducing PI3K/Akt. Chin. German J. Clin. Oncol. 2010, 9, 734–738. [Google Scholar] [CrossRef]
- Yun, J.; Lv, Y.G.; Yao, Q.; Wang, L.; Li, Y.P.; Yi, J. Wortmannin inhibits proliferation and induces apoptosis of MCF-7 breast cancer cells. Eur. J. Gynaecol. Oncol. 2012, 33, 367–369. [Google Scholar]
- Ng, S.S.W.; Tsao, M.; Chow, S.; Hedley, D.W. Inhibition of phosphatidylinositide 3-kinase enhances gemcitabine-induced apoptosis in human pancreatic cancer cells. Cancer Res. 2000, 60, 5451–5455. [Google Scholar]
- Ng, S.S.W.; Tsao, M.; Nicklee, T.; Hedley, D.W. Wortmannin inhibits PKB/Akt phosphorylation and promotes gemcitabine antitumor activity in orthotopic human pancreatic cancer xenografts in immunodeficient mice. Clini. Cancer Res. 2001, 7, 3269–3275. [Google Scholar]
- Boehle, A.S.; Kurdow, R.; Boenicke, L.; Schniewind, B.; Faendrich, F.; Dohrmann, P.; Kalthoff, H. Wortmannin inhibits growth of human non-small-cell lung cancer in vitro and in vivo. Langenbeck Arch. Surg. 2002, 387, 234–239. [Google Scholar] [CrossRef]
- Au, T.K.; Chick, W.S.; Leung, P.C. The biology of ophiobolins. Life Sci. 2000, 67, 733–742. [Google Scholar] [CrossRef]
- Ahn, J.-W.; Lee, M.-K.; Choi, S.-U.; Lee, C.-O.; Kim, B.-S. Cytotoxic ophiobolins produced by Bipolaris sp. J. Microbiol. Biotechn. 1998, 8, 406–408. [Google Scholar]
- Singh, S.B.; Smith, J.L.; Sabnis, G.S.; Dombrowski, A.W.; Schaeffer, J.M.; Goetz, M.A.; Bills, G.F. Structure and conformation of ophiobolin K and 6- epiophiobolin K from Aspergillus ustus as a nematocidal agent. Tetrahedron 1991, 47, 6931–6938. [Google Scholar] [CrossRef]
- Nozoe, S.; Itai, A.; Tsuda, K.; Okuda, S. The chemical transformation of cephalonic acid. Tetrahedron Lett. 1967, 42, 4113–4117. [Google Scholar] [CrossRef]
- Bills, G.F.; Platas, G.; Gams, W. Conspecificity of the cerulenin and helvolic acid producing “Cephalosporium caerulens”, and the hypocrealean fungus Sarocladium oryzae. Mycolo. Res. 2004, 108, 1291–1300. [Google Scholar] [CrossRef]
- Sugawara, F.; Strobel, G.; Strange, R.N.; Siedow, J.N.; Van Duyne, G.D.; Clardy, J. Phytotoxins from the pathogenic fungi Drechslera maydis and Drechslera sorghicola. Proc. Natl. Acad. Sci. USA 1987, 84, 3081–3085. [Google Scholar]
- De Vries-van Leeuwen, I.J.; Kortekaas-Thijssen, C.; Mandouckou, J.A.N.; Kas, S.; Evidente, A.; de Boer, A.H. Fusicoccin—A selectively induces apoptosis in tumor cells after interferon-alpha priming. Cancer Lett. 2010, 293, 198–206. [Google Scholar] [CrossRef]
- Shen, X.; Krasnoff, S.B.; Lu, S.W.; Dunbar, C.D.; O’Neal, J.; Turgeon, B.G.; Yoder, O.C.; Gibson, D.M.; Hamann, M.T. Characterization of 6-epi-3-anhydroophiobolin B from Cochliobolus heterostrophus. J. Nat. Prod. 1999, 62, 895–897. [Google Scholar] [CrossRef]
- Yang, T.; Lu, Z.; Meng, L.; Wei, S.; Hong, K.; Zhu, W.; Huang, C. The novel agent ophiobolin O induces apoptosis and cell cycle arrest of MCF-7 cells through activation of MAPK signaling pathways. Bioorgan. Med. Chem. 2012, 22, 579–585. [Google Scholar] [CrossRef]
- Zhang, D.; Fukuzawa, S.; Satake, M.; Li, X.; Kuranaga, T.; Niitsu, A.; Yoshizawa, K.; Tachibana, K. Ophiobolin O and 6-epi-ophiobolin O, two new cytotoxic sesterterpenes from the marine derived fungus Aspergillus sp. Nat. Prod. Commun. 2012, 7, 1411–1414. [Google Scholar]
- Wang, Q.-X.; Yang, J.-L.; Qi, Q.-Y.; Bao, L.; Yang, X.-L.; Liu, M.-M.; Huang, P.; Zhang, L.-X.; Chen, J.-L.; Cai, L.; et al. 3-Anhydro-6-hydroxy-ophiobolin A, a new sesterterpene inhibiting the growth of methicillin-resistant Staphylococcus aureus and inducing the cell death by apoptosis on K562, from the phytopathogenic fungus Bipolaris oryzae. Bioorgan. Med. Chem. Lett. 2013, 23, 3547–3550. [Google Scholar] [CrossRef]
- Li, E.; Clark, A.M.; Rotella, D.P.; Hufford, C.D. Microbial metabolites of ophiobolin A and antimicrobial evaluation of ophiobolins. J. Nat. Prod. 1995, 58, 74–81. [Google Scholar] [CrossRef]
- Krizsán, K.; Bencsik, O.; Nyilasi, I.; Galgóczy, L.; Vágvölgyi, C.; Papp, T. Effect of the sesterterpene-type metabolites, ophiobolins A and B, on zygomycetes fungi. FEMS Microbiol. Lett. 2010, 313, 135–140. [Google Scholar] [CrossRef]
- Geris, R.; Simpson, T.J. Meroterpenoids produced by fungi. Nat. Prod. Rep. 2009, 26, 1063–1094. [Google Scholar] [CrossRef]
- Schümann, J.; Hertweck, C. Molecular basis of cytochalasan biosynthesis in fungi: gene cluster analysis and evidence for the involvement of a PKS-NRPS hybrid synthase by RNA silencing. J. Am. Chem. Soc. 2007, 129, 9564–9565. [Google Scholar] [CrossRef]
- Sekita, S.; Yoshihira, K.; Natori, S.; Kuwano, H. Structures of chaetoglobosin A and B, cytotoxic metabolites of Chaetomium globosum. Tetrahedron Lett. 1973, 14, 2109–2112. [Google Scholar] [CrossRef]
- Fu, J.; Zhou, Y.; Li, H.; Ye, Y.; Guo, J. Antifungal metabolites from Phomopsis sp By254, an endophytic fungus in Gossypium hirsutum. Afr. J. Microbiol. Res. 2011, 5, 1231–1236. [Google Scholar]
- Wicklow, D.T.; Rogers, K.D.; Dowd, P.F.; Gloer, J.B. Bioactive metabolites from Stenocarpella maydis, a stalk and ear rot pathogen of maize. Fungal Biol. 2011, 115, 133–142. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Berestetskiy, A.; Evidente, A. Production of phytotoxins by Phoma exigua var. exigua, a potential mycoherbicide against perennial thistles. J. Agr. Food Chem. 2008, 56, 6304–6309. [Google Scholar]
- Xu, H.; Fang, W.-S.; Chen, X.-G.; He, W.-Y.; Cheng, K.-D. Cytochalasin D from Hypocrella Bambusae. J. Asian Nat. Prod. Res. 2001, 3, 151–155. [Google Scholar] [CrossRef]
- Demain, A.L.; Hunt, N.A.; Malik, V.; Kobbe, B.; Hawkins, H.; Matsuo, K.; Wogan, G.N. Improved procedure for production of cytochalasin E and tremorgenic mycotoxins by Aspergillus clavatus. Appl. Environ. Microbiol. 1976, 31, 138–140. [Google Scholar]
- Wagenaar, M.M.; Corwin, J.; Strobel, G.; Clardy, J. Three new cytochalasins produced by an endophytic fungus in the genus Rhinocladiella. J. Nat. Prod. 2000, 63, 1692–1695. [Google Scholar] [CrossRef]
- Liu, R.; Gu, Q.; Zhu, W.; Cui, C.; Fan, G.; Fang, Y.; Zhu, T.; Liu, H. 10-Phenyl-[12]-cytochalasins Z7, Z8, and Z9 from the marine-derived fungus Spicaria elegans. J. Nat. Prod. 2006, 69, 871–875. [Google Scholar] [CrossRef]
- Thohinung, S.; Kanokmedhakul, S.; Kanokmedhakul, K.; Kukongviriyapan, V.; Tusskorn, O.; Soytong, K. Cytotoxic 10-(indol-3-yl)-[13]cytochalasans from the fungus Chaetomium elatum ChE01. Arch. Pharm. Res. 2010, 33, 1135–1141. [Google Scholar] [CrossRef]
- Ding, G.; Song, Y.C.; Chen, J.R.; Xu, C.; Ge, H.M.; Wang, X.T.; Tan, R.X. Chaetoglobosin U, a cytochalasan alkaloid from endophytic Chaetomium globosum IFB-E019. J. Nat. Prod. 2006, 69, 302–304. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Ren, W.; Zhao, D.; Zhu, Y.; Wu, X. Bioactive metabolites from Chaetomium globosum L18, an endophytic fungus in the medicinal plant Curcuma wenyujin. Phytomedicine 2012, 19, 364–368. [Google Scholar] [CrossRef]
- Bloch, P.; Tamm, C.; Bollinger, P.; Petcher, T.J.; Weber, H.P. 13. Pseurotin, a new metabolite of Pseudeurotium ovalis Stolk having an unusual hetero-spirocyclic system. Helv. Chim. Acta 1976, 59, 133–137. [Google Scholar] [CrossRef]
- Wenke, J.; Anke, H.; Sterner, O. Pseurotin A and 8-O-demethylp-seurotin A from Aspergillus fumigatus and their inhibitory activities on chitin synthase. Biosci. Biotechnol. Biochem. 1993, 57, 961–964. [Google Scholar] [CrossRef]
- Martinez-Luis, S.; Cherigo, L.; Arnold, E.; Spadafore, C.; Gerwick, W.H.; Cubilla-Rios, L. Antiparasitic and anticancer constituentd of the endophytic fungus Aspergillus sp. strain F1544. Nat. Prod. Commun. 2012, 7, 165–168. [Google Scholar]
- Ge, H.M.; Shen, Y.; Zhu, C.H.; Tan, S.H.; Ding, H.; Song, Y.C.; Tan, R.X. Penicidones A–C, three cytotoxic alkaloidal metabolites of an endophytic Penicillium sp. Phytochemistry 2008, 69, 571–576. [Google Scholar] [CrossRef]
- Shao, C.-L.; Wang, C.-Y.; Gu, Y.-C.; Wei, M.-Y.; Pan, J.-H.; Deng, D.-S.; She, Z.-G.; Lin, Y.-C. Penicinoline, a new pyrrolyl 4-quinolinone alkaloid with an unprecedented ring system from an endophytic fungus Penicillium sp. Bioorg. Med. Chem. Lett. 2010, 20, 3284–3286. [Google Scholar] [CrossRef]
- Elsebai, M.F.; Rempel, V.; Schnakenburg, G.; Kehraus, S.; Christa, E.M.; Gabriele, M.K. Identification of a potent and selective cannabinoid CB1 receptor antagonist from Auxarthron reticulatum. ACS Med. Chem. Lett. 2011, 2, 866–869. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, L.; Zhu, T.; Gu, Q.; Li, D. Unusual pyrrolyl 4-quinolinone alkaloids from the marine-derived fungus Penicillium sp. ghq208. Chem. Pharm. Bull. 2012, 60, 1458–1460. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–173. [Google Scholar]
- Larsen, T.O.; Frisvad, J.C.; Ravn, G.; Skaaning, T. Mycotoxin production by Penicillium expansum on blackcurrant and cherry juice. Food Addit. Contam. 1998, 15, 671–675. [Google Scholar] [CrossRef]
- Dalsgaard, P.W.; Blunt, J.W.; Munro, M.H.G.; Frisvad, J.C.; Christophersen, C. Communesins G and H, new alkaloids from the psychrotolerant fungus Penicillium rivulum. J. Nat. Prod. 2005, 68, 258–261. [Google Scholar] [CrossRef]
- Numata, A.; Takahashi, C.; Ito, Y.; Takada, T.; Kawai, K.; Usami, Y.; Matsumura, E.; Imachia, M.; Ito, T.; Hasegawa, T. Communesins, cytotoxic metabolites of a fungus isolated from a marine alga. Tetrahedron 1993, 34, 2355–2358. [Google Scholar] [CrossRef]
- Jadulco, R.; Edrada, R.A.; Ebel, R.; Berg, A.; Schaumann, K.; Wray, V.; Steube, K.; Proksch, P. New communesin derivatives from the fungus Penicillium sp. derived from the Mediterranean sponge Axinella verrucosa. J. Nat. Prod. 2004, 67, 78–81. [Google Scholar] [CrossRef]
- Eble, T.E.; Hanson, F.R. Fumagillin, an antibiotic from Aspergillus fumigatus H-3. Antibiot. Chemother. 1951, 1, 54–58. [Google Scholar]
- Frisvad, J.C.; Samson, R.A.; Stolk, A.C. A new species of Penicillium, P. scabrosum. Persoonia 1990, 14, 177–182. [Google Scholar]
- Ingber, D.; Fujita, T.; Kishimoto, S.; Sudo, K.; Kanamaru, T.; Brem, H.; Folkman, J. Synthetic analogues of fumagillin that inhibit angiogenesis and suppress tumour growth. Nature 1990, 348, 555–557. [Google Scholar] [CrossRef]
- Kusaka, M.; Sudo, K.; Fujita, T.; Marui, S.; Itoh, F.; Ingber, D.; Folkman, J. Potent anti-angiogenic action of AGM-1470: comparison to the fumagillin parent. Biochem. Bioph. Res. Co. 1991, 174, 1070–1076. [Google Scholar] [CrossRef]
- Yamaoka, M.; Yamamoto, T.; Ikeyama, S.; Sudo, K.; Fujita, T. Angiogenesis inhibitor TNP-470 (AGM-1470) potently inhibits the tumor growth of hormone-independent human breast and prostate carcinoma cell lines. Cancer Res. 1993, 51, 5233–5236. [Google Scholar]
- Kruger, E.A.; Figg, W.D. TNP-470: An angiogenesis inhibitor in clinical development for cancer. Expert Opin. Inv. Drug 2000, 9, 1383–1396. [Google Scholar] [CrossRef]
- Bhargava, P.; Marshall, J.L.; Rizvi, N.; Dahut, W.; Yoe, J.; Figuera, M.; Phipps, K.; Ong, V.S.; Kato, A.; Hawkins, M.J. A Phase I and pharmacokinetic study of TNP-470 administered weekly to patients with advanced cancer. Clin. Cancer Res. 1999, 5, 1989–1995. [Google Scholar]
- Vansteelandt, M.; Blanchet, E.; Egorov, M.; Petit, F.; Toupet, L.; Bondon, A.; Monteau, F.; le Bizec, B.; Thomas, O.P.; Pouchus, Y.F.; et al. Ligerin, an antiproliferative chlorinated sesquiterpenoid from a marine-derived Penicillium strain. J. Nat. Prod. 2013, 76, 297–301. [Google Scholar] [CrossRef]
- Fang, S.-M.; Cui, C.-B.; Li, C.-W.; Wu, C.-J.; Zhang, Z.-J.; Li, L.; Huang, X.-J.; Ye, W.-C. Purpurogemutantin and purpurogemutantidin, new drimenyl cyclohexenone derivatives produced by a mutant obtained by diethyl sulfate mutagenesis of a marine-derived Penicillium purpurogenum G59. Mar. Drugs 2012, 10, 1266–1287. [Google Scholar] [CrossRef]
- Sassa, T.; Ishizaki, A.; Nukina, M.; Ikeda, M.; Sugiyama, T. Isolation and identification of new antifungal macrophorins E, F and G as malonyl meroterpenes from Botryosphaeria berengeriana. Biosci. Biotechnol. Biochem. 1998, 62, 2260–2262. [Google Scholar] [CrossRef]
- Cabedo, N.; López-Gresa, M.P.; Primo, J.; Ciavatta, M.L.; González-Mas, M.C. Isolation and structural elucidation of eight new related analogues of the mycotoxin (−)-botryodiplodin from Penicillium coalescens. J. Agr. Food Chem. 2007, 55, 6977–6983. [Google Scholar]
- Fuska, J.; Proksa, B.; Uhrín, D. The antibiotic PSX-1 produced by Penicillium stipitatum is identical with botryodiplodin. Folia Microbiol. 1988, 33, 238–240. [Google Scholar] [CrossRef]
- Fuska, J.; Kuhr, I.; Nemec, P.; Fuskova, A. Antitumor antibiotics produced by Penicillium stipitatum THOM. J. Antibiot. 1973, 27, 123–127. [Google Scholar]
- Omura, S.; Tomoda, H.; Kimura, K.; Zhen, D.-Z.; Kumagai, H.; Igarashi, K.; Imamura, N.; Takahashi, Y.; Tanaka, Y.; Iwai, Y. Atpenins, new antifungal antibiotics produced by Penicillium sp. J. Antibiot. 1988, 41, 1769–1773. [Google Scholar] [CrossRef]
- Kawada, M.; Momose, I.; Someno, T.; Tsujiuchi, G.; Ikeda, D. New atpenins, NBRI23477 A and B, inhibit the growth of human prostate cancer cells. J. Antibiot. 2009, 62, 243–246. [Google Scholar] [CrossRef]
- Boysen, M.; Skouboe, P.; Frisvad, J.; Rossenl, L. Reclassification of the Penicillium roqueforti group into three species on the basis of molecular genetic and biochemical profiles. Microbiology 1996, 142, 541–549. [Google Scholar] [CrossRef]
- Li, X.-J.; Zhang, Q.; Zhang, A.-L.; Gao, J.-M. Metabolites from Aspergillus fumigatus, an endophytic fungus associated with Melia azedarach, and their antifungal, antifeedant, and toxic activities. J. Agr. Food Chem. 2012, 60, 3424–3431. [Google Scholar] [CrossRef]
- Ma, G.; Khan, S.I.; Jacob, M.R.; Babu, L.; Li, Z.; Pasco, D.S.; Walker, L.A.; Khan, I.A.; Tekwani, B.L. Antimicrobial and antileishmanial activities of hypocrellins A and B. Antimicrob. Agents Ch. 2004, 48, 4450–4452. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Kato, H.; Samizo, M.; Nojiri, Y.; Onuki, H.; Hirota, H.; Ohta, T. Notoamides F-K, prenylated indole alkaloids isolated from a marine-derived Aspergillus sp. J. Nat. Prod. 2008, 71, 2064–2067. [Google Scholar] [CrossRef]
- Nakagawa, M.; Hirota, A.; Sakai, H. Terrecyclic acid A, a new antibiotic from Aspergillus terreus. I. Taxonomy, production, and chemical and biological properties. J. Antibiot. 1982, 35, 778–782. [Google Scholar]
- Sun, H.-F.; Li, X.-M.; Meng, L.; Cui, C.-M.; Gao, S.-S.; Li, C.-S.; Huang, C.-G.; Wang, B.-G. Asperolides A–C, tetranorlabdane diterpenoids from the marine alga-derived endophytic fungus Aspergillus wentii EN-48. J. Nat. Prod. 2012, 75, 148–152. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bladt, T.T.; Frisvad, J.C.; Knudsen, P.B.; Larsen, T.O. Anticancer and Antifungal Compounds from Aspergillus, Penicillium and Other Filamentous Fungi. Molecules 2013, 18, 11338-11376. https://doi.org/10.3390/molecules180911338
Bladt TT, Frisvad JC, Knudsen PB, Larsen TO. Anticancer and Antifungal Compounds from Aspergillus, Penicillium and Other Filamentous Fungi. Molecules. 2013; 18(9):11338-11376. https://doi.org/10.3390/molecules180911338
Chicago/Turabian StyleBladt, Tanja Thorskov, Jens Christian Frisvad, Peter Boldsen Knudsen, and Thomas Ostenfeld Larsen. 2013. "Anticancer and Antifungal Compounds from Aspergillus, Penicillium and Other Filamentous Fungi" Molecules 18, no. 9: 11338-11376. https://doi.org/10.3390/molecules180911338
APA StyleBladt, T. T., Frisvad, J. C., Knudsen, P. B., & Larsen, T. O. (2013). Anticancer and Antifungal Compounds from Aspergillus, Penicillium and Other Filamentous Fungi. Molecules, 18(9), 11338-11376. https://doi.org/10.3390/molecules180911338