Enhancement of Diosgenin Production in Dioscorea zingiberensis Cell Cultures by Oligosaccharides from Its Endophytic Fungus Fusarium oxysporum Dzf17


Abstract
:Abbreviations
| DP | degree of polymerization |
| DP2-5 | fraction composed of oligosaccharides with DPs from 2 to 5 |
| DP5-8 | fraction composed of oligosaccharides with DPs from 5 to 8 |
| DP8-12 | fraction composed of oligosaccharides with DPs from 8 to 12 |
| DP4 | oligosaccharide with DP as four |
| DP7 | oligosaccharide with DP as seven |
| DP10 | oligosaccharide with DP as ten; dw: dry weight |
| HPLC | high performance liquid chromatography |
| TFA | trifluoroacetic acid |
| TLC | thin-layer chromatography |
| WPS | water-extracted mycelial polysaccharide |
1. Introduction
2. Results
2.1. Effects of Oligosaccharide Fractions DP2-5, DP5-8 and DP8-12
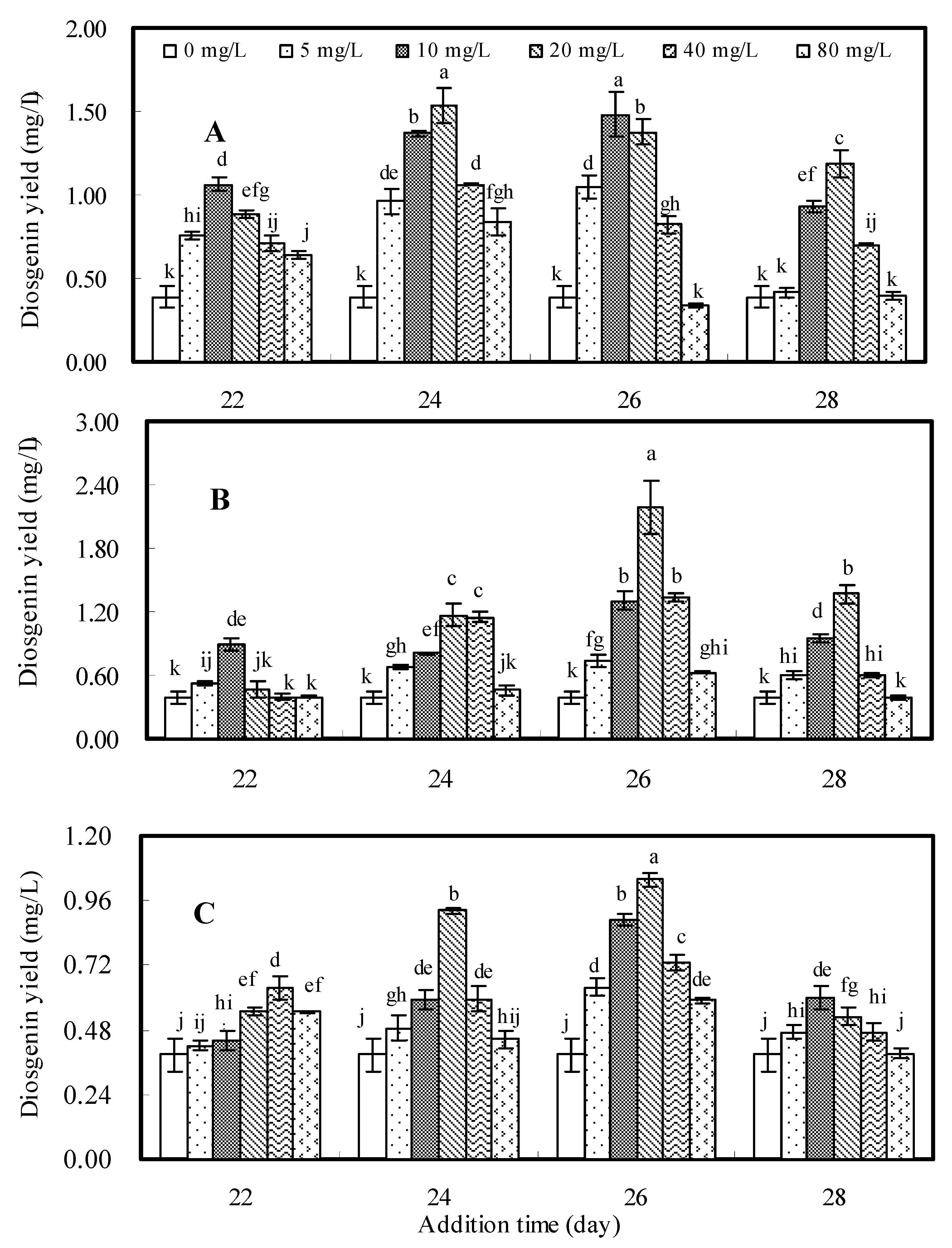
2.2. Effects of Oligosaccharides DP4, DP7 and DP10
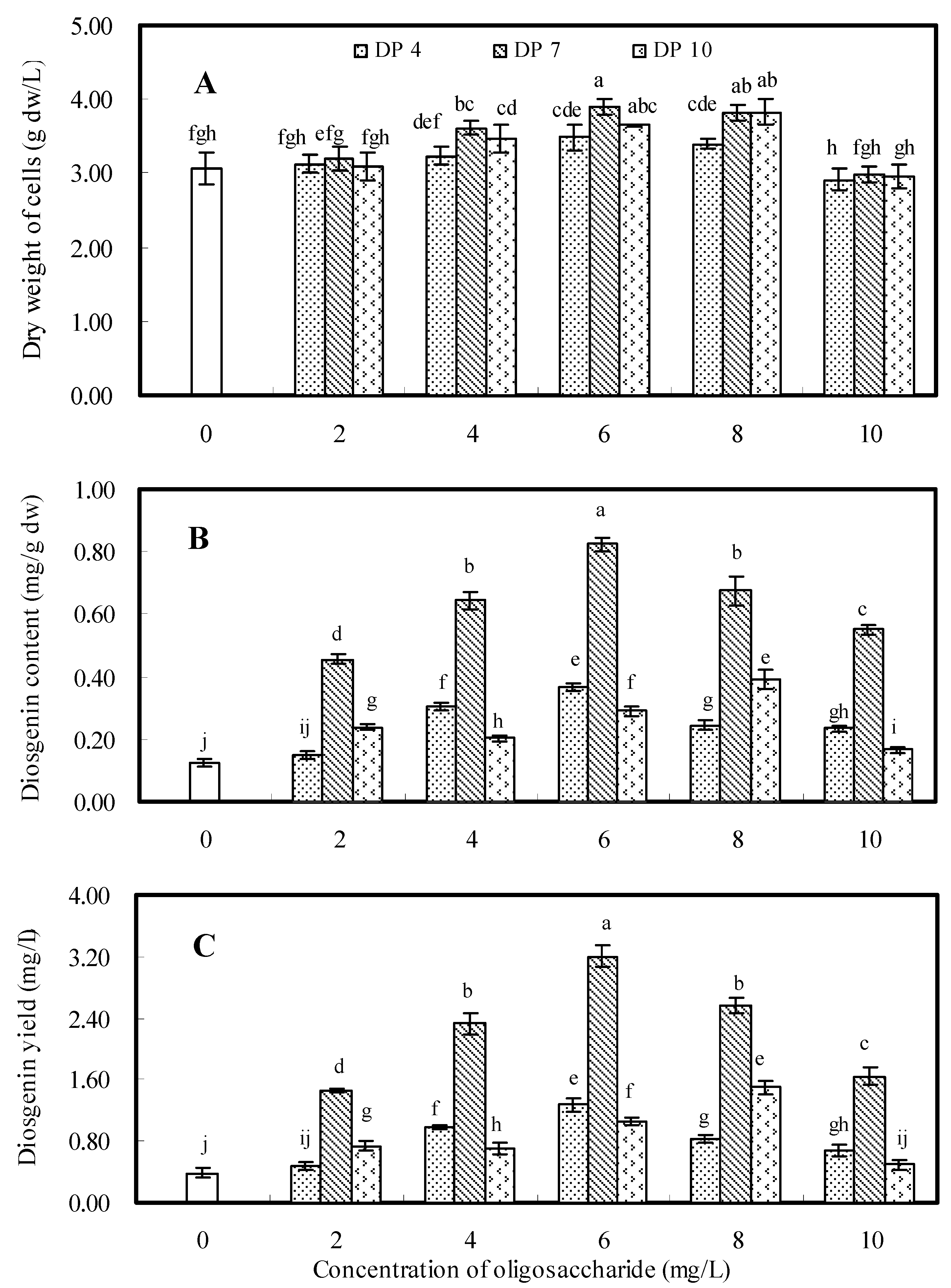
2.3. Dynamics of Cell Growth and Diosgenin Accumulation Elicited once by DP7
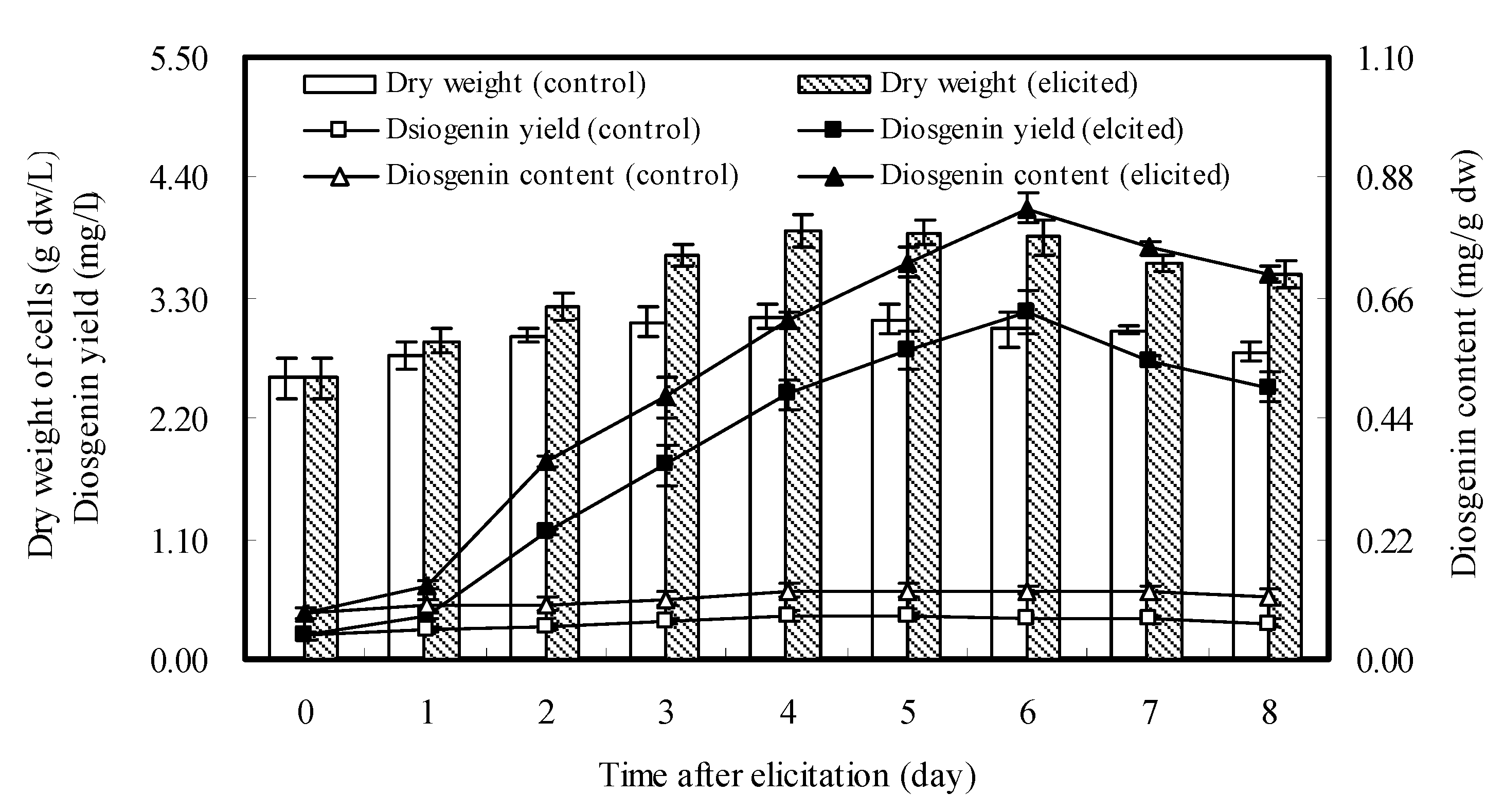
2.4. Effects of Repeated Elicitation of DP7 on Cell Growth and Diosgenin Accumulation

{kind=link}
{kind=link}
{kind=link}
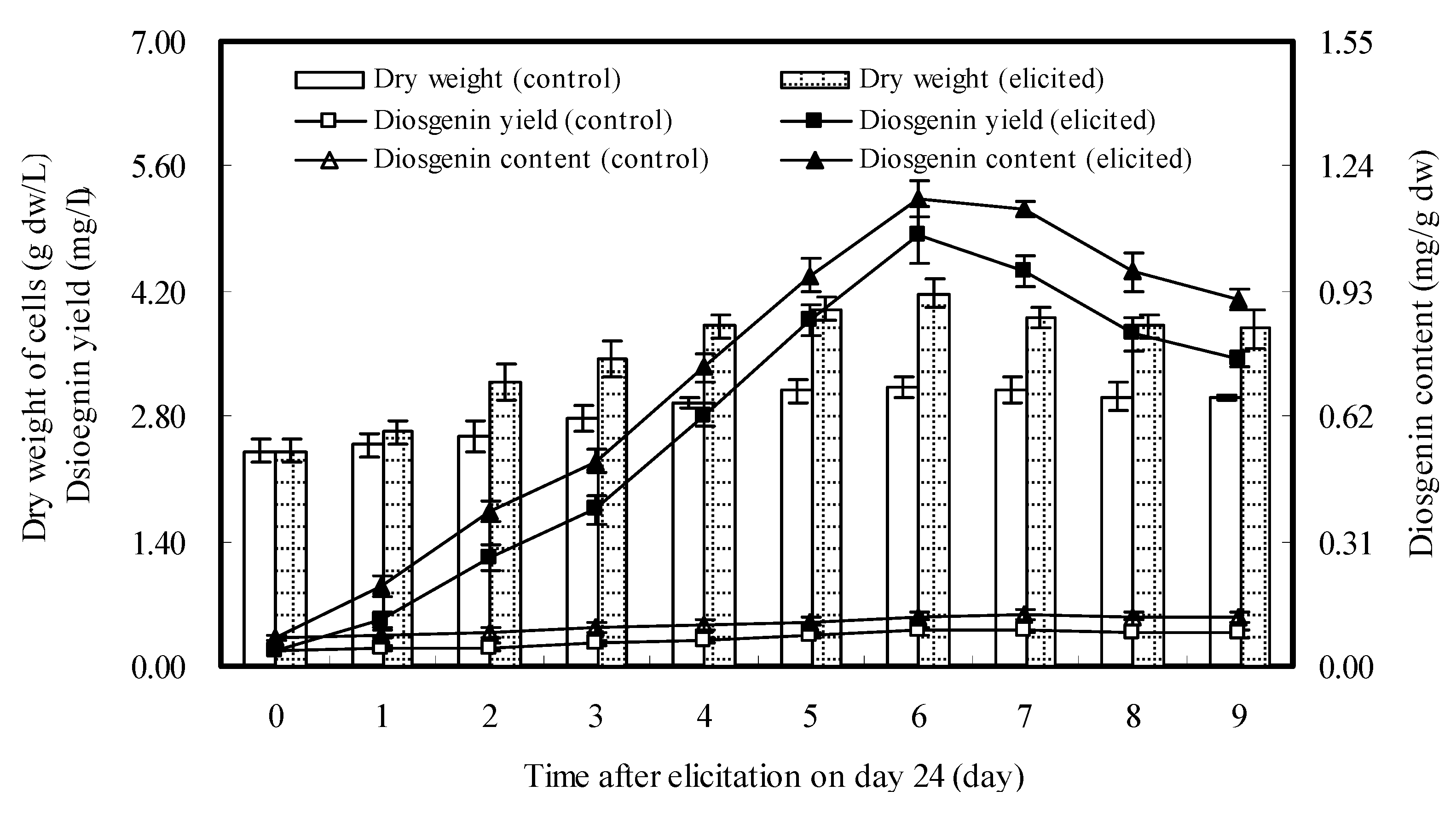{kind=link}
| Addition time | Dry weight | Diosgenin content | Diosgenin yield |
|---|---|---|---|
| (day) | (g dw/L) | (mg/g dw) | (g/L) |
| Control (without elicitation) | 3.070 ± 0.217 c | 0.126 ± 0.012 d | 0.387 ± 0.061 d |
| Days 24 and 26 | 3.809 ± 0.164 a | 0.931 ± 0.039 a | 3.548 ± 0.280 a |
| Day 26 | 3.898 ± 0.143 a | 0.821 ± 0.029 b | 3.203 ± 0.222 b |
| Days 26 and 28 | 3.332 ± 0.057 b | 0.733 ± 0.021 c | 2.440 ± 0.030 c |
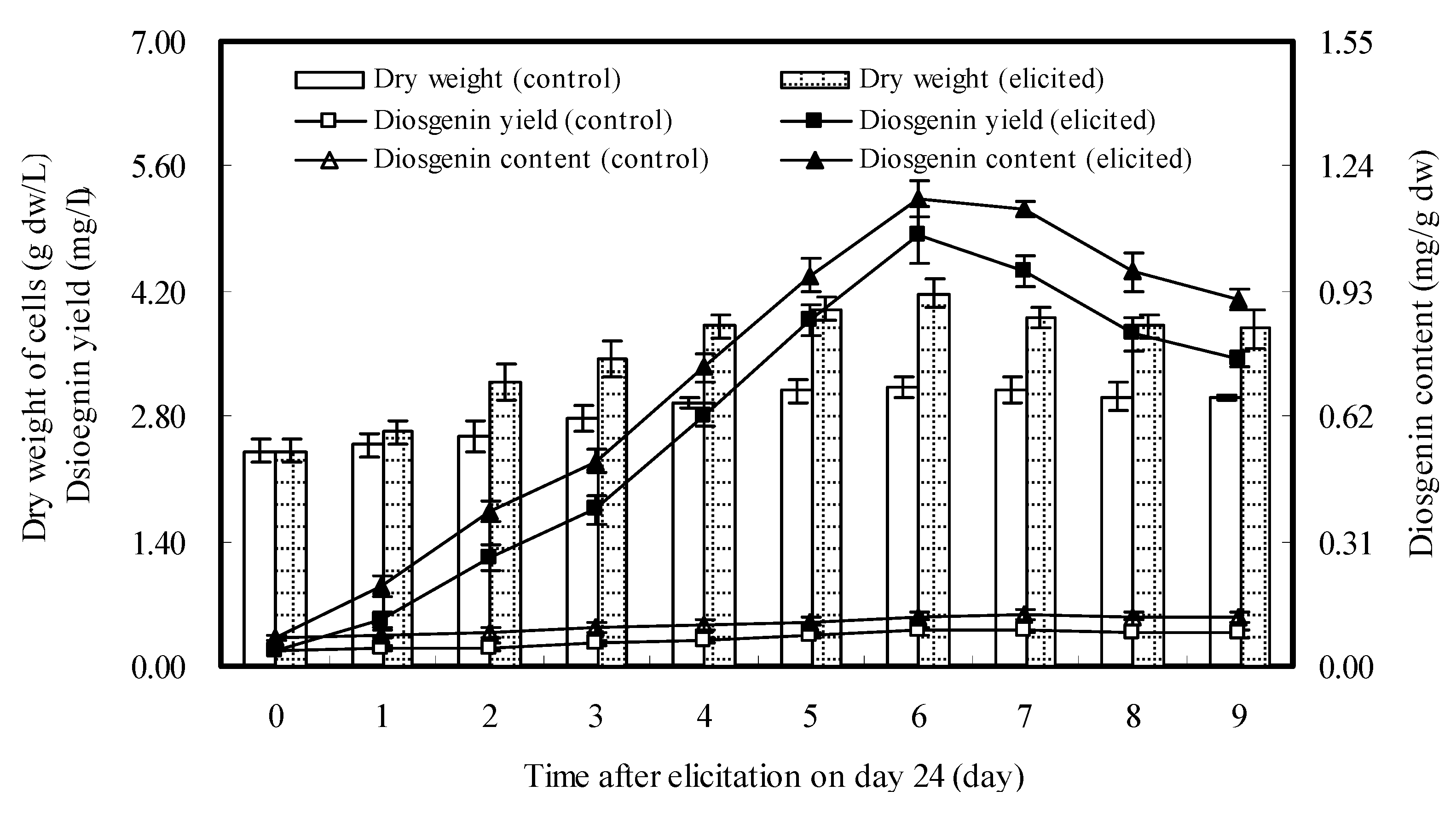
3. Discussion
4. Experimental
4.1. Cell Suspension Cultures
4.2. Preparation of Oligosaccharides
4.3. Elicitation Treatment of the Suspension Cells
4.4. Determination of Cell Dry Weight
4.5. Diosgenin Extraction and Quantification
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgements
References and Notes
- Sautour, M.; Mitaine-Offer, A.-C.; Lacaille-Dubois, M.-A. The Dioscorea genus: A review of bioactive steroid saponins. J. Nat. Med. 2007, 61, 91–101. [Google Scholar] [CrossRef]
- Roman, I.D.; Thewles, A.; Coleman, R. Fractionation of livers following diosgenin treatment to elevate binary cholesterol. Biochim. Biophys. Acta 1995, 1255, 77–81. [Google Scholar] [CrossRef]
- Gong, G.; Qin, Y.; Huang, W.; Zhou, S.; Wu, X.; Yang, X.; Zhao, Y.; Li, D. Protective effects of diosgenin in the hyperlipidemic rat model and in human vascular endothelial cells against hydrogen peroxide-induced apoptosis. Chem. Biol. Interact. 2010, 184, 366–375. [Google Scholar] [CrossRef]
- Chen, P.-S.; Shih, Y.-W.; Huang, H.-C.; Cheng, H.-W. Diosgenin, a steroidal saponin, inhibits migration and invasion of human prostate cancer PC-3 cells by reducing matrix metalloproteinases expression. PLoS One 2011, 6, e20164. [Google Scholar]
- Aradhana; Rao, A.R.; Kale, R.K. Diosgenin—A growth stimulator of mammary gland of ovariectomized mouse. Indian J. Exp. Biol. 1992, 30, 367–370. [Google Scholar]
- Wang, Y.; Pan, K.; Hsieh, T.; Chang, T.; Lin, W.; Hsu, J.T. Diosgenin, a plant-derived sapogenin, exhibits antiviral activity in vitro against hepatitis C virus. J. Nat. Prod. 2011, 74, 580–584. [Google Scholar] [CrossRef]
- Raju, J.; Mehta, R. Cancer chemopreventive and therapeutic effects of diosgenin, a food saponin. Nutr. Cancer 2009, 61, 27–35. [Google Scholar] [CrossRef]
- Tada, Y.; Kanda, N.; Haratake, A.; Tobiishi, M.; Uchiwa, H.; Watanabe, S. Novel effects of diosgenin on skin aging. Steroids 2009, 74, 504–511. [Google Scholar] [CrossRef]
- Chapagain, B.P.; Saharan, V.; Pelah, D.; Yadav, R.C.; Wiesman, Z. Bioproduction of diosgenin in callus cultures of Balanites aegyptiaca: Effect of growth regulators, explants and somatic embryogenesis. Nat. Prod. Commun. 2006, 1, 215–221. [Google Scholar]
- Lambert, N.; Baccou, J.C.; Sauvaire, Y. Screening for diosgenin in rhizomes from 3 Costus species (C. deistellii, C. igneous, C. lucanusianus). Planta Med. 1988, 54, 366–367. [Google Scholar]
- Huang, W.; Zhang, H.; Ni, J.; Zuo, H.; Qiu, L.; Li, H.; Li, H. The best utilization of Dioscorea zingiberensis C. H. Wright by an eco-friendly process. Bioresource Technol. 2008, 99, 7407–7411. [Google Scholar]
- Zhou, L.; Wu, J. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep. 2006, 23, 789–810. [Google Scholar] [CrossRef]
- Dornenburg, H.; Knorr, D. Strategies for the improvement of secondary metabolite production in plant cell cultures. Enzyme Microb. Technol. 1995, 17, 674–684. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, C.; Li, J.; Wang, S.; Wu, J. Heptasaccharide and octasaccharide isolated from Paris polyphylla var. yunnanensis and their plant growth-regulatory activity. Plant Sci. 2003, 165, 571–575. [Google Scholar] [CrossRef]
- Zhou, L.; Cao, X.; Zhang, R.; Peng, Y.; Zhao, S.; Wu, J. Stimulation of saponin production in Panax ginseng hairy roots by two oligosaccharides from Paris polyphylla var yunnanensis. Biotechnol. Lett. 2007, 29, 631–634. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, L.; Wu, J. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures. Appl. Microbiol. Biotechnol. 2010, 87, 137–144. [Google Scholar] [CrossRef]
- Sanchez-Sampedro, M.A.; Femandez-Tarrago, J.; Corchete, P. Yeast extract and methyl jasmonate-induced silymarin production in cell cultures of Silybum marianum (L.) Gaertn. J. Biotechnol. 2005, 119, 60–69. [Google Scholar]
- Marero, L.M.; Jin, J.H.; Shin, J.H.; Lee, H.J.; Chung, I.S.; Lee, H.J. Effect of fungal elicitation on indirubin production from a suspension culture of Polygonum tinctorium. Enzyme Microb. Technol. 1997, 21, 97–101. [Google Scholar] [CrossRef]
- Chen, H.; Chen, F. Effects of yeast elicitor on the growth and secondary metabolism of a high-tanshinone-producing line of the Ti transformed Salvia miltiorrhiza cells in suspension culture. Process Biochem. 2000, 35, 837–840. [Google Scholar] [CrossRef]
- Rojas, R.; Alba, J.; Magana-Plaza, I.; Cruz, F.; Ramos-Valdivia, A.C. Stimulated production of diosgenin in Dioscorea galeottiana cell suspension cultures by abiotic and biotic factors. Biotechnol. Lett. 1999, 21, 907–911. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C.; Draeger, S.; Rommert, A.-K.; Krohn, K. Endophytic fungi: a source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Kharwar, R.N.; Mishra, A.; Gong, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z.; Tan, R. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Zhang, R.; Li, P.; Xu, L.; Chen, Y.; Sui, P.; Zhou, L.; Li, J. Enhancement of diosgenin production in Dioscorea zingiberensis cell culture by oligosaccharide elicitor from its endophytic fungus Fusarium oxysporum. Nat. Prod. Commun. 2009, 4, 1459–1462. [Google Scholar]
- Field, R.A. Oligosaccharide signaling molecules. In Plant-Derived Natural Products; Osbourn, A.E., Lanzotti, V., Eds.; Springer Science + Business Media: New York, NY, USA, 2009; pp. 349–359. [Google Scholar]
- Suryanarayanan, T.S.; Thirunavukkarasu, N.; Govindarajulu, M.B.; Sasse, F.; Jansen, R.; Murali, T.S. Fungal endophytes and bioprospectin. Fungal Biol. Rev. 2009, 23, 9–19. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef]
- Linden, J.C.; Phisalaphong, M. Oligosaccharides potentiate methyl jasmonate -induced production of paclitaxel in Taxus canadensis. Plant Sci. 2000, 158, 41–51. [Google Scholar] [CrossRef]
- Wang, Z.; Zhong, J. Repeated elcitation enhances taxane production in suspension cultures of Taxus chinensis in bioreactors. Biotechnol. Lett. 2002, 24, 445–448. [Google Scholar] [CrossRef]
- Cheng, X.; Guo, B.; Zhou, H.; Ni, W.; Liu, C. Repeated elicitation enhances phenylethanoid glycosides accumulation in cell suspension cultures of Cistanche deserticola. Biochem. Eng. J. 2005, 24, 203–207. [Google Scholar] [CrossRef]
- Cheng, X.; Zhou, H.; Cui, X.; Ni, W.; Liu, C. Improvement of phenylethanoid glycosides biosynthesis in Cistanche deserticola cell suspension cultures by chitosan elicitor. J. Biotechnol. 2006, 121, 253–260. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Li, P.; Mou, Y.; Shan, T.; Xu, J.; Li, Yan.; Lu, S.; Zhou, L. Effects of polysaccharide elicitors from endophytic Fusarium oxysporium Dzf17 on growth and diosgenin production in cell suspension culture of Dioscorea zingiberensis. Molecules 2011, 16, 9003–9016. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, W.; Ni, J. A promising clean process for production of diosgenin from Dioscorea zingiberensis C. H. Wright. J. Clean. Prod. 2010, 18, 242–247. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the oligosaccharides are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, P.; Mao, Z.; Lou, J.; Li, Y.; Mou, Y.; Lu, S.; Peng, Y.; Zhou, L. Enhancement of Diosgenin Production in Dioscorea zingiberensis Cell Cultures by Oligosaccharides from Its Endophytic Fungus Fusarium oxysporum Dzf17. Molecules 2011, 16, 10631-10644. https://doi.org/10.3390/molecules161210631
Li P, Mao Z, Lou J, Li Y, Mou Y, Lu S, Peng Y, Zhou L. Enhancement of Diosgenin Production in Dioscorea zingiberensis Cell Cultures by Oligosaccharides from Its Endophytic Fungus Fusarium oxysporum Dzf17. Molecules. 2011; 16(12):10631-10644. https://doi.org/10.3390/molecules161210631
Chicago/Turabian StyleLi, Peiqin, Ziling Mao, Jingfeng Lou, Yan Li, Yan Mou, Shiqiong Lu, Youliang Peng, and Ligang Zhou. 2011. "Enhancement of Diosgenin Production in Dioscorea zingiberensis Cell Cultures by Oligosaccharides from Its Endophytic Fungus Fusarium oxysporum Dzf17" Molecules 16, no. 12: 10631-10644. https://doi.org/10.3390/molecules161210631
APA StyleLi, P., Mao, Z., Lou, J., Li, Y., Mou, Y., Lu, S., Peng, Y., & Zhou, L. (2011). Enhancement of Diosgenin Production in Dioscorea zingiberensis Cell Cultures by Oligosaccharides from Its Endophytic Fungus Fusarium oxysporum Dzf17. Molecules, 16(12), 10631-10644. https://doi.org/10.3390/molecules161210631