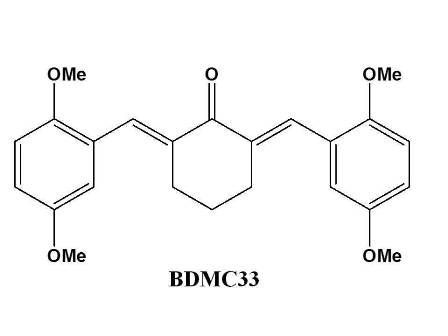
A Curcumin Derivative, 2,6-Bis(2,5-dimethoxybenzylidene)-cyclohexanone (BDMC33) Attenuates Prostaglandin E2 Synthesis via Selective Suppression of Cyclooxygenase-2 in IFN-g/LPS-Stimulated Macrophages

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abbreviations
| BDMC33 | 2,6-bis(2,5-dimethoxybenzylidene)cyclohexanone |
| COX | cyclooxygenase |
| DMSO | dimethyl sulfoxide |
| DXM | dexamethasone |
| FBS | foetal bovine serum |
| HRP | horseradish peroxidase |
| IFN-γ | interferon-gamma |
| LPS | lipopolysaccharide |
| NO | nitric oxide |
| PGE2 | prostaglandin E2 |
| NSAID | non-steroidal anti-inflammatory drug |
| RAW 264.7 | murine macrophage cells |
1. Introduction
2. Results and Discussion
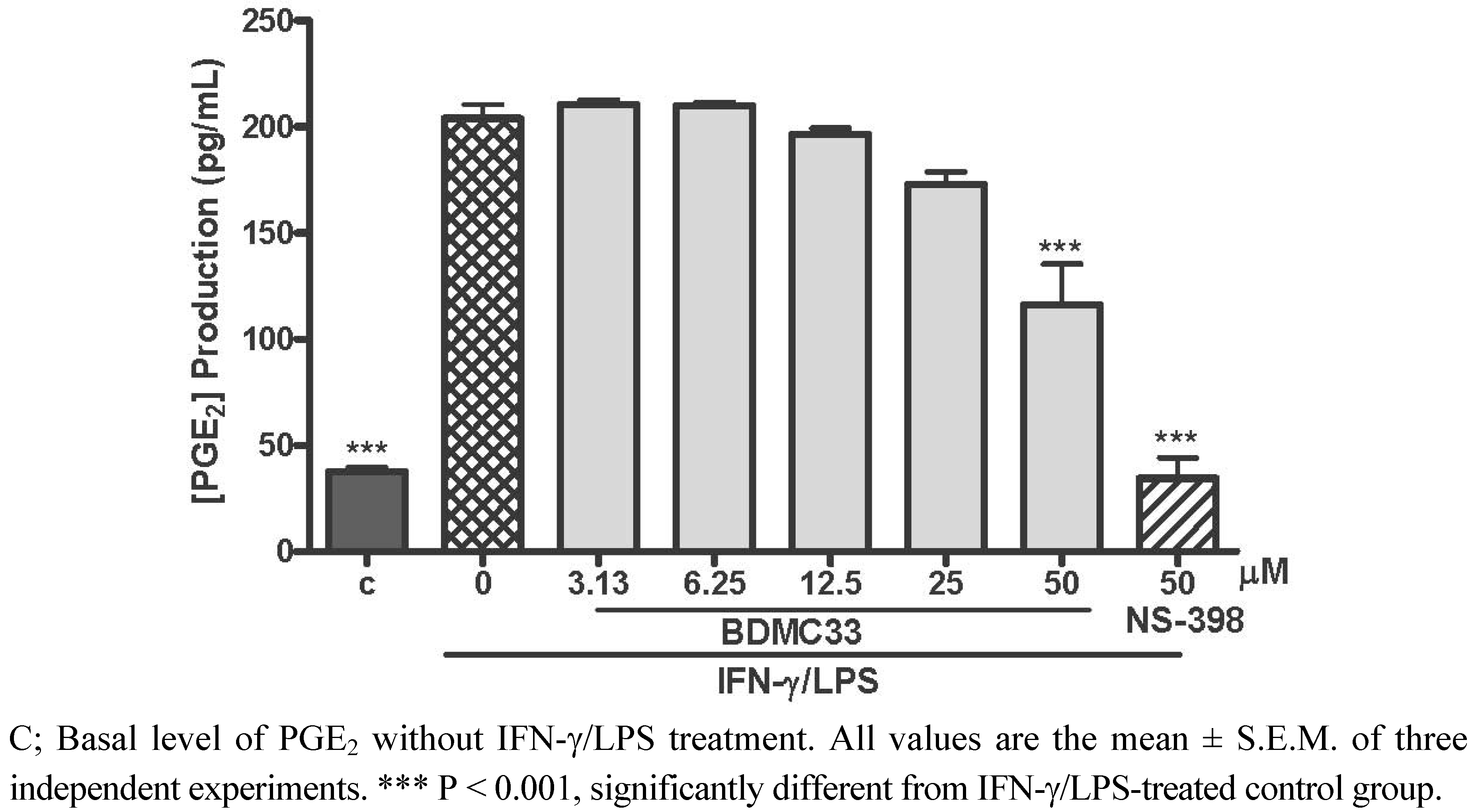
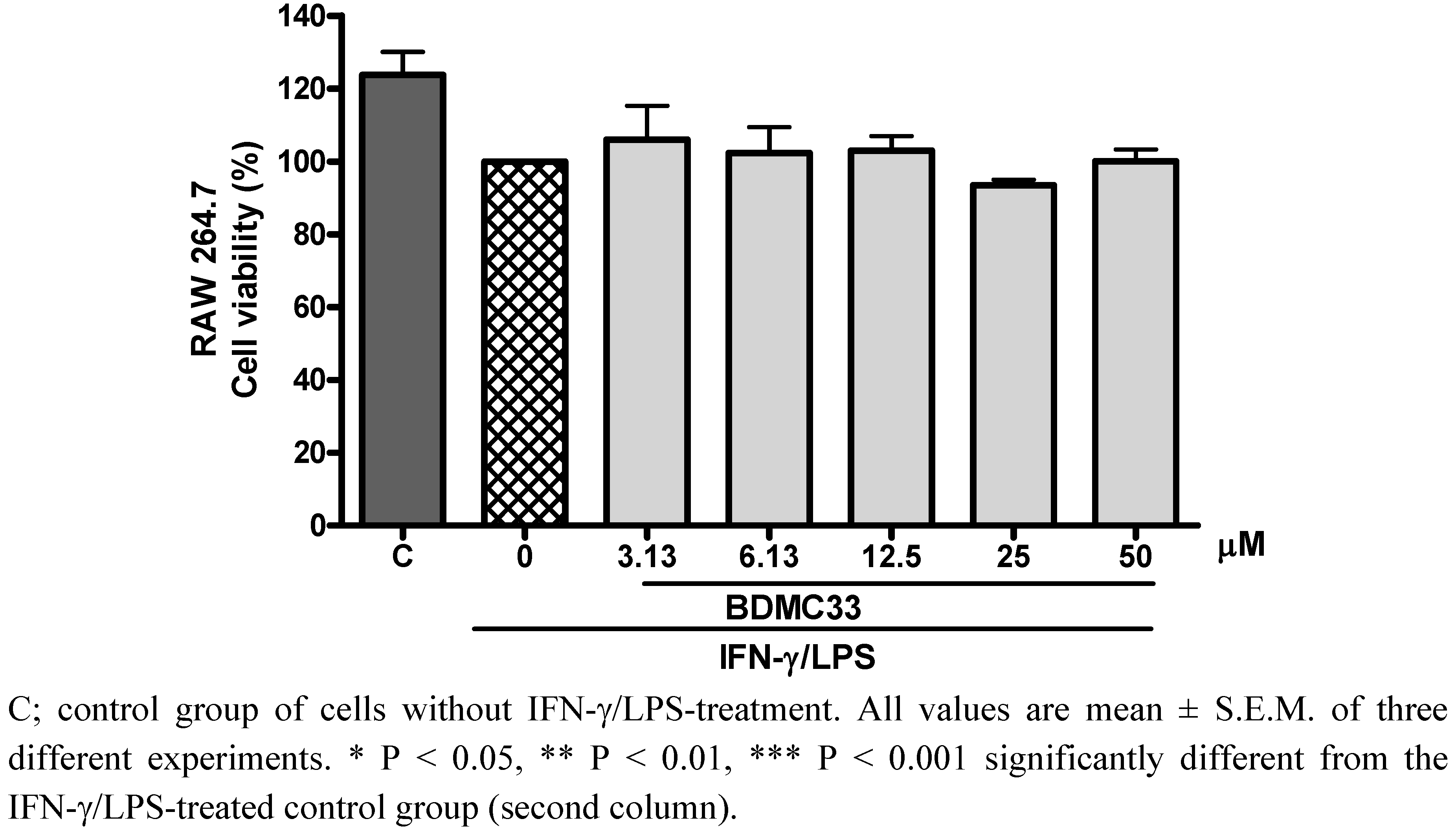
2.1. Effects of BDMC33 on PGE2 Secretion and Cell Viability
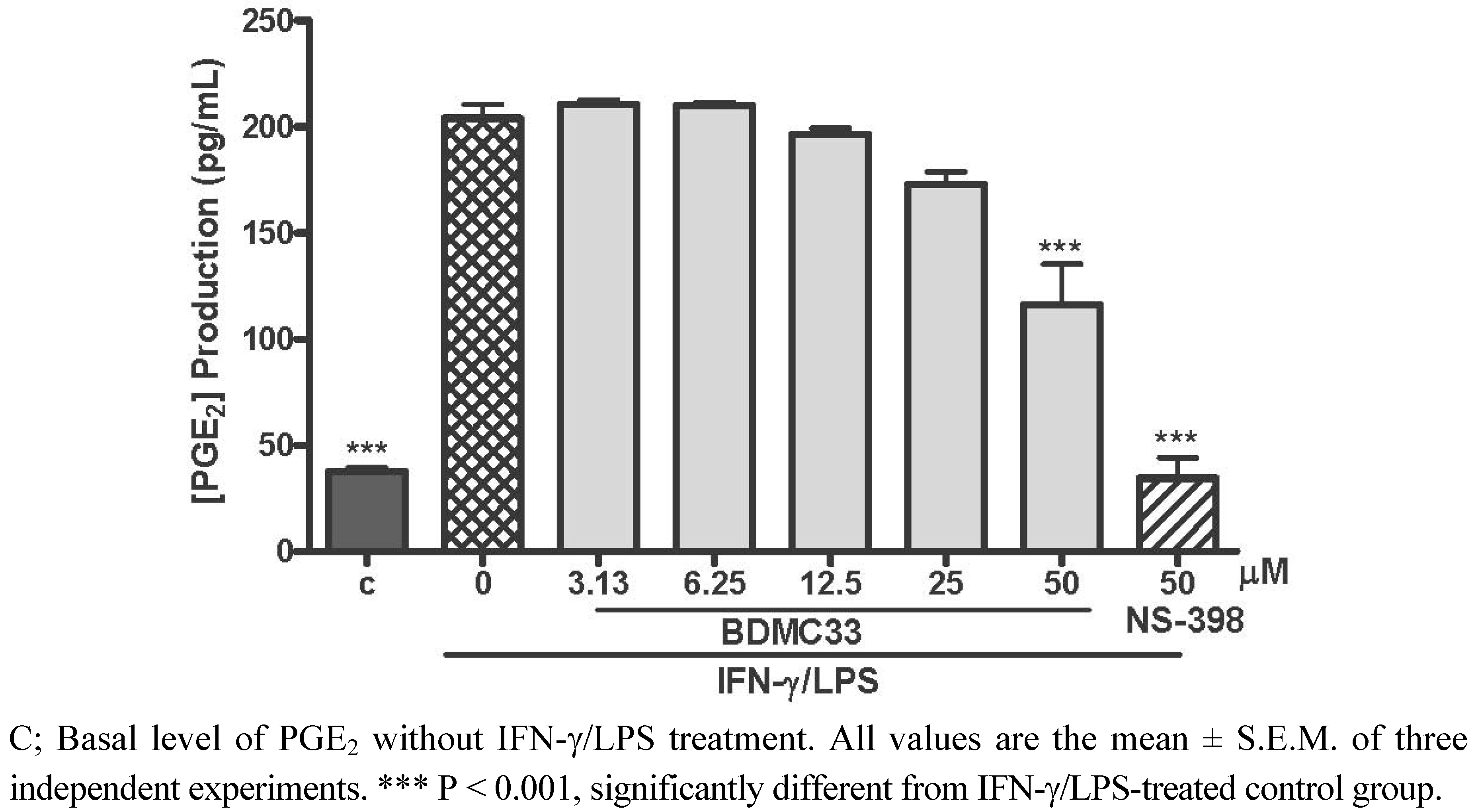
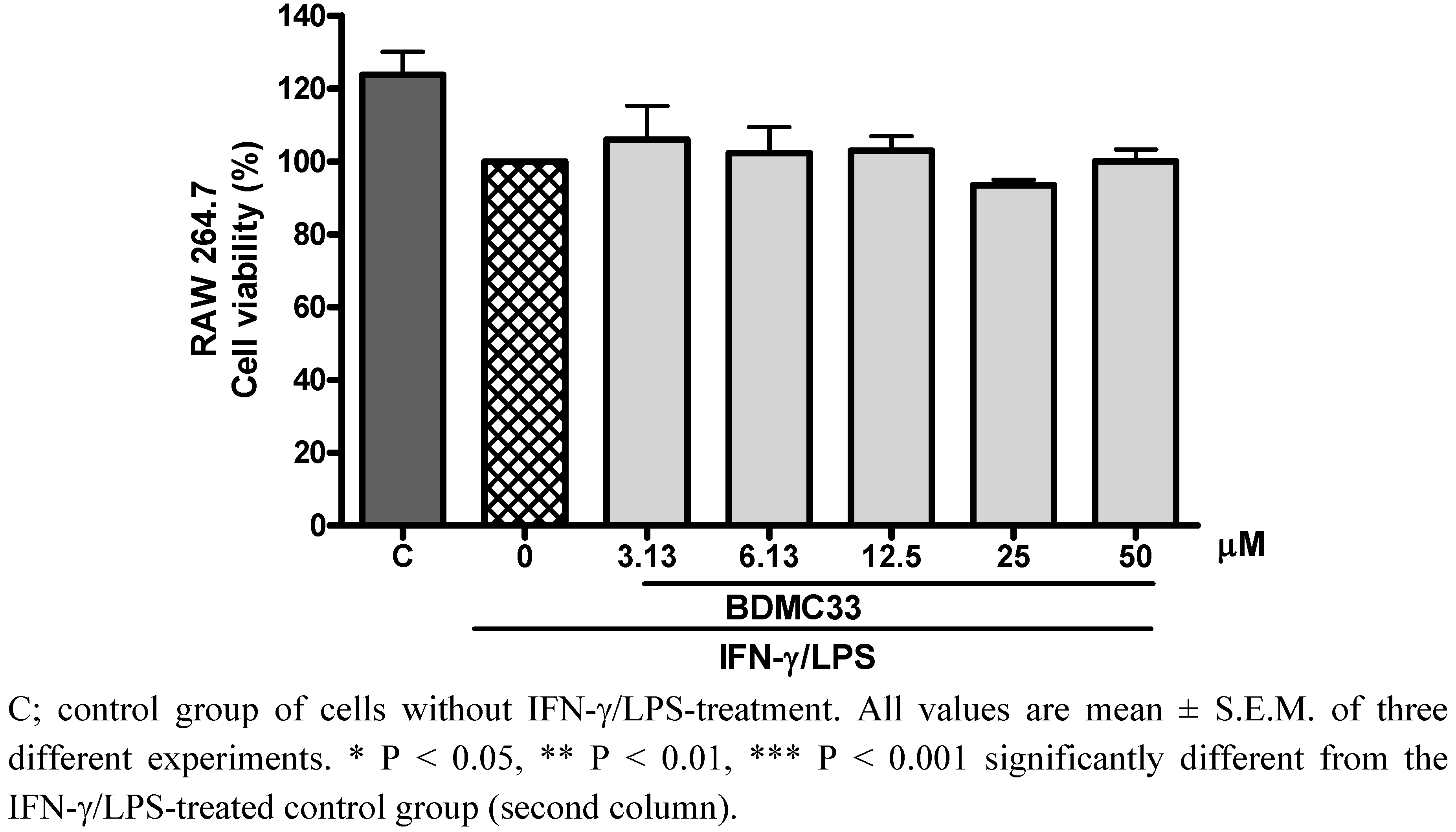
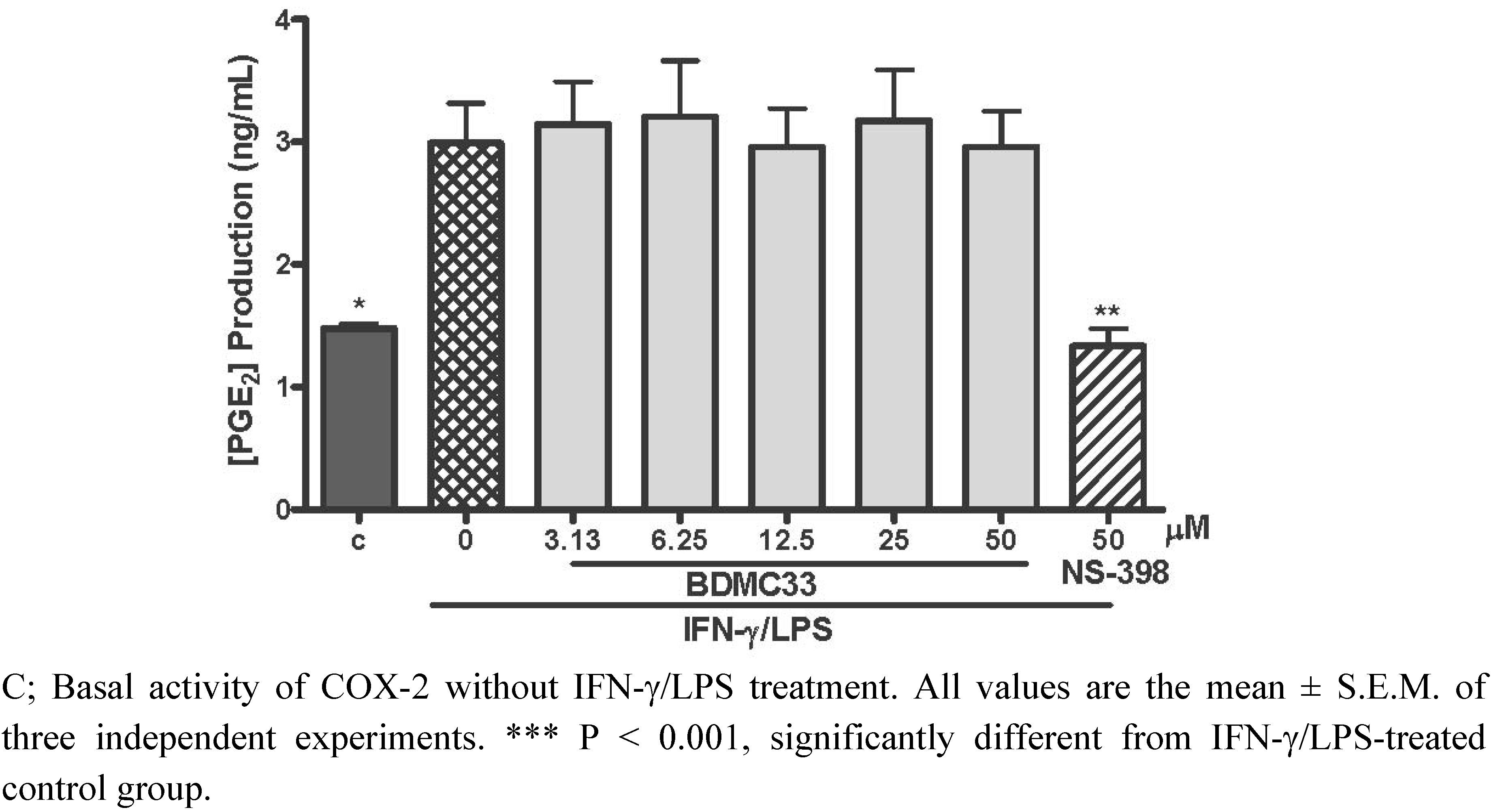
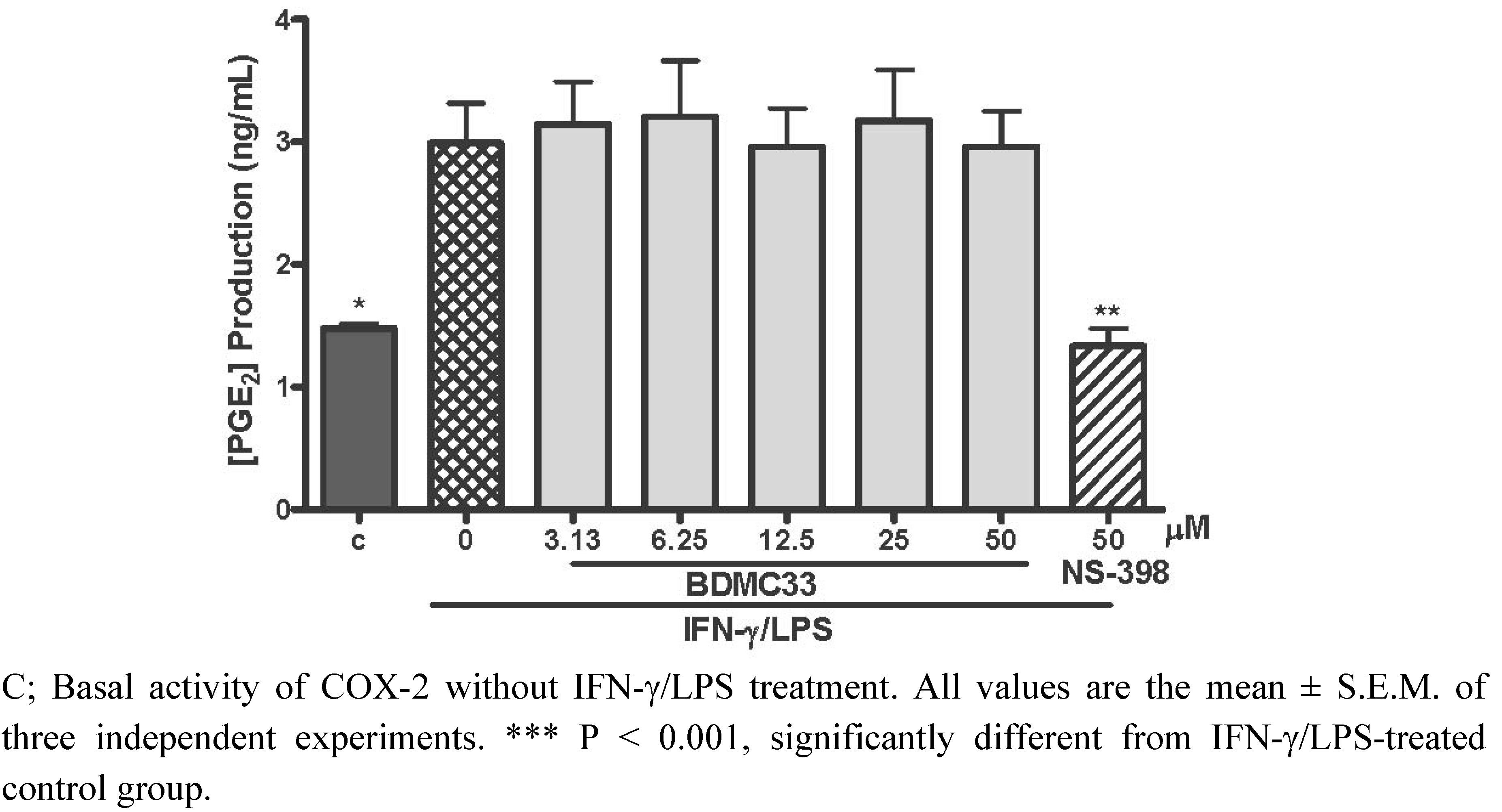
2.2. Effects of BDMC33 on COX-2 Enzymatic Activity
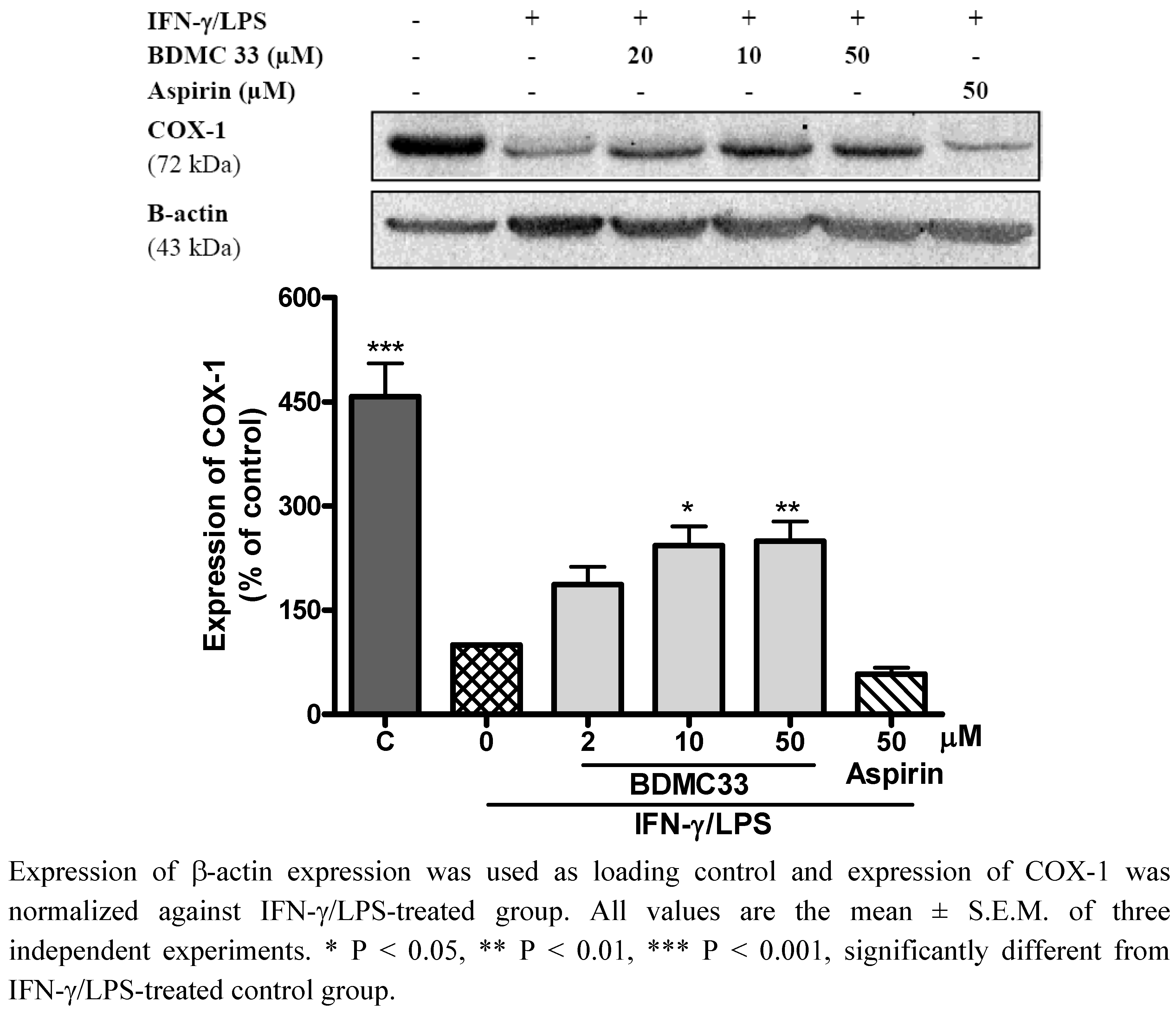
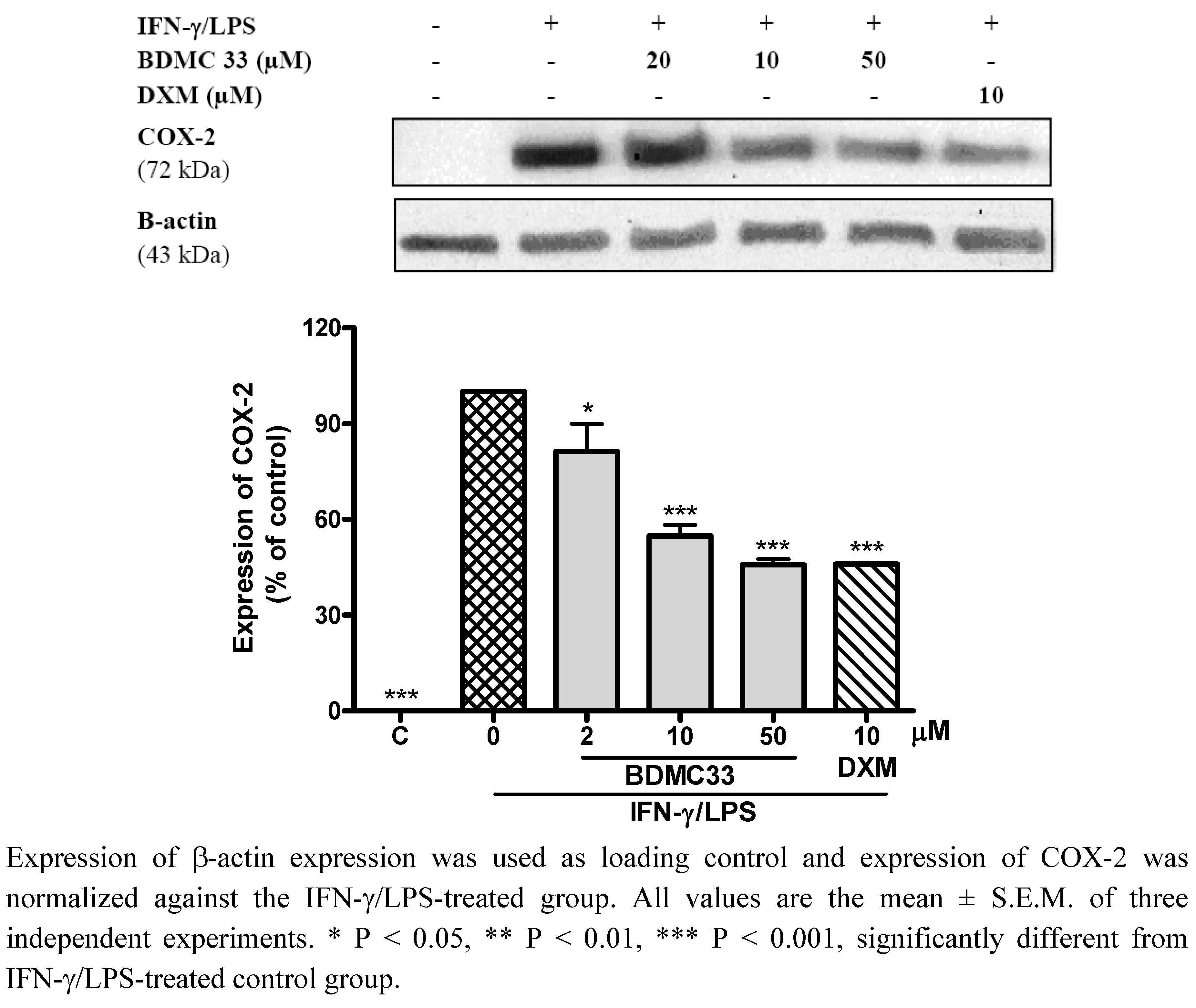
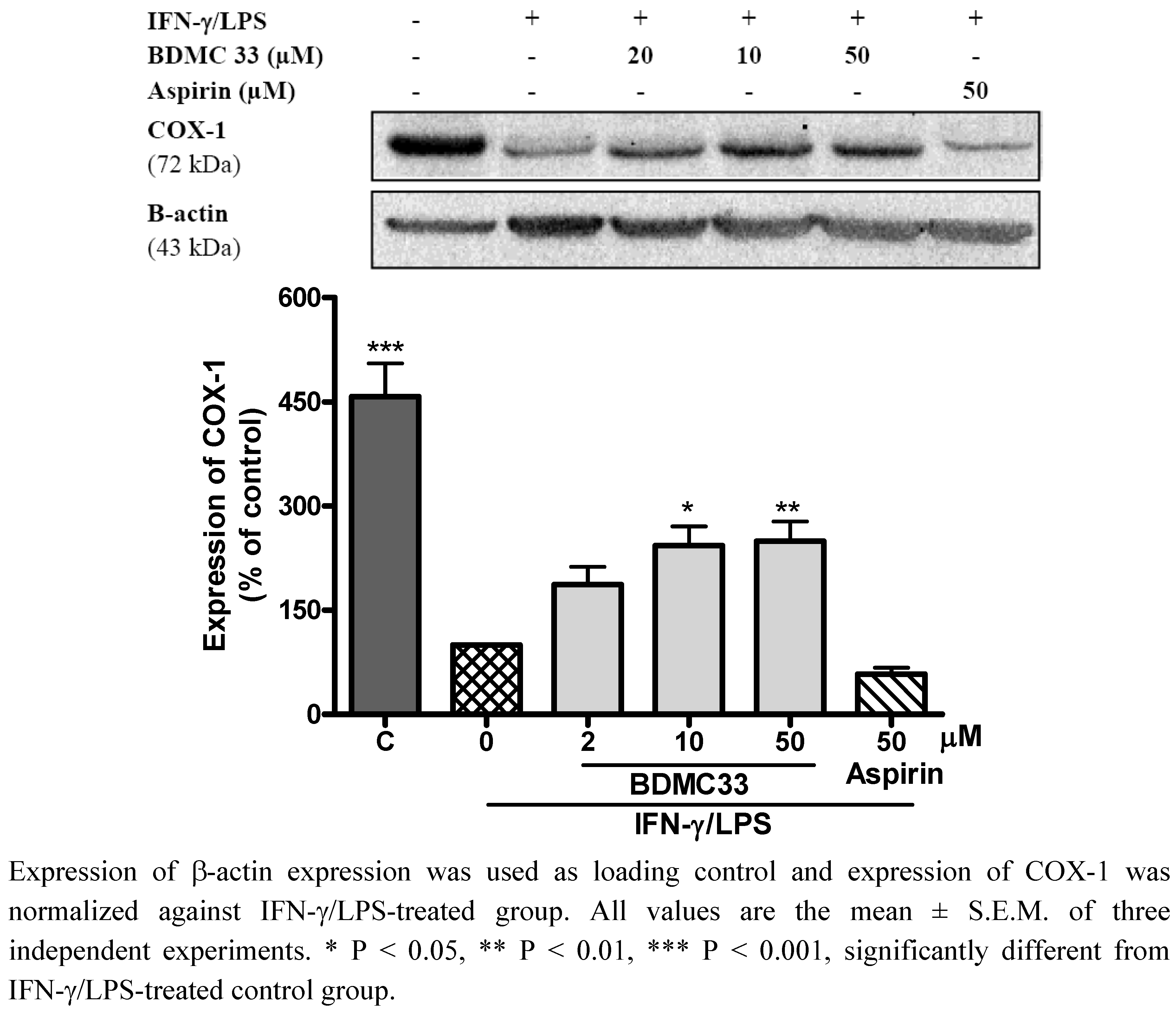
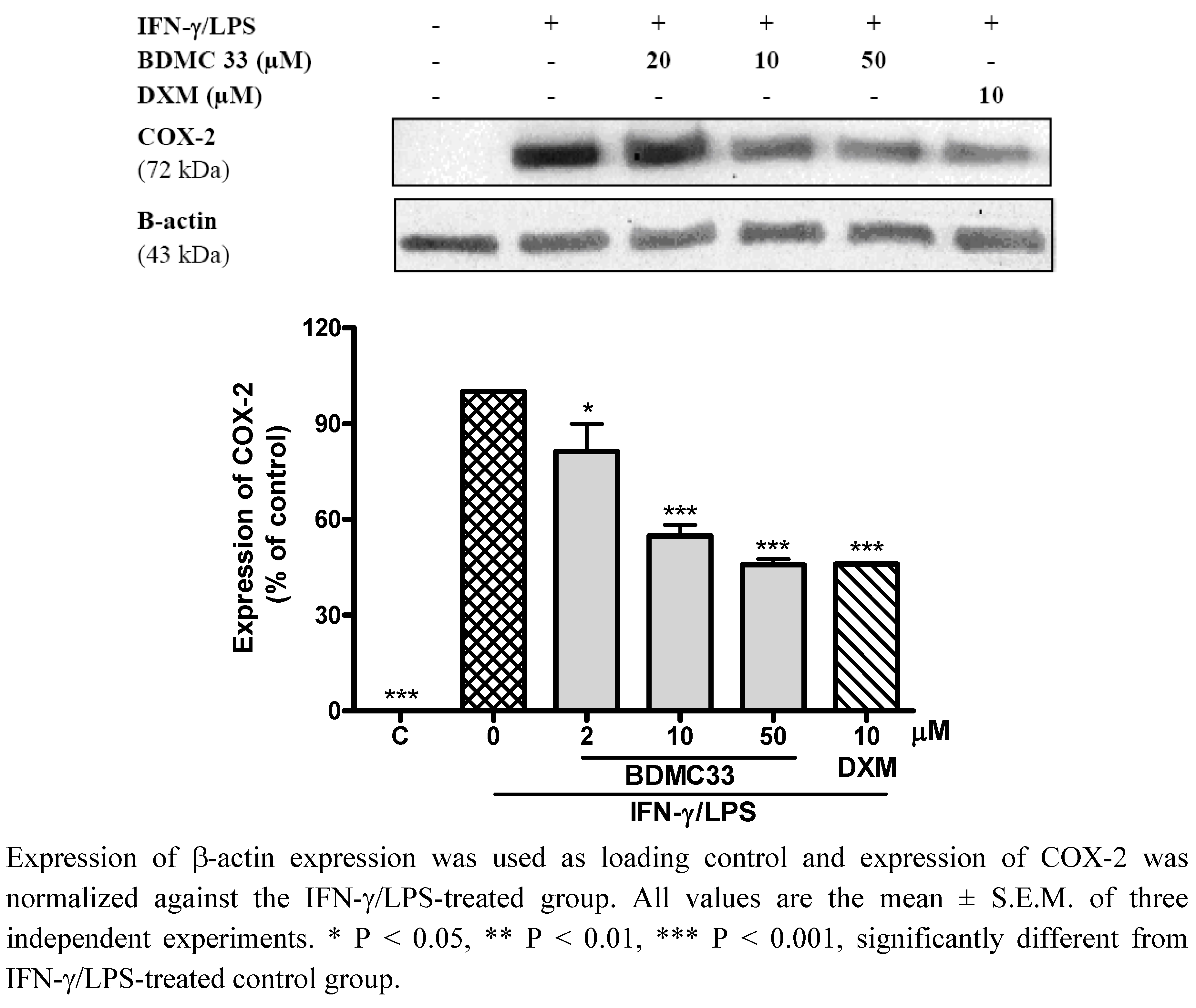
2.3. Effects of BDMC33 on COX Expression
3. Experimental
3.1. Materials
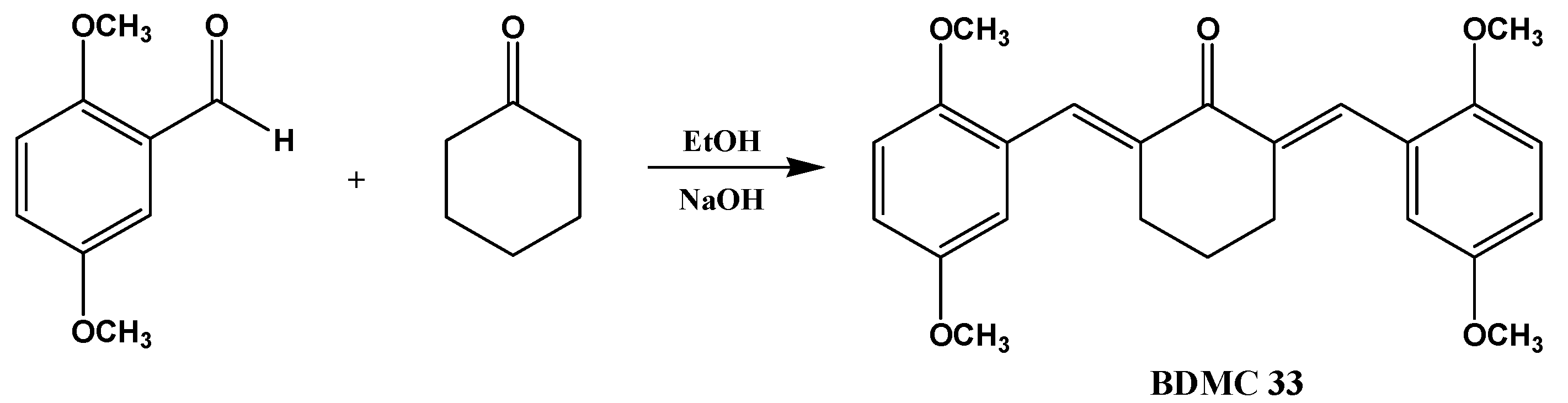
3.2. Synthesis of 2,6-Bis(2,5-dimethoxybenzylidene)cyclohexanone (BDMC33)

3.3. Cell Culture
3.4. Cell Stimulation and Treatment
3.5. Cell Viability
3.6. Determination of PGE2
3.7. Indirect Determination of Endogenous COX-2 Activity
3.8. Western Blotting Analysis of COX Expression
3.9. Statistical Analysis
4. Conclusions
Acknowledgment
References
- Serhan, C.N.; Levy, B. Success of prostaglandin E2 in structure–function is a challenge for structure-based therapeutics. Proc. Natl. Acad. Sci. USA 2003, 100, 8609–8611. [Google Scholar] [CrossRef]
- Park, G.Y.; Christman, G.W. Involvement of cyclooxygenase-2 and prostaglandins in the pathogenesis of the inflammatory lung disease. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 290, L797–L805. [Google Scholar] [CrossRef]
- Park, J.Y.; Pilinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Pelletier, J.P.; Fahmi, H. Cyclooxygenase-2 and prostaglandins in articular tissues. Semin. Arthritis Rheum. 2003, 33, 155–167. [Google Scholar] [CrossRef]
- Chandrasekharan, N.V.; Dai, H.; Roos, K.L.; Evanson, N.K.; Tomsik, J.; Elton, T.S.; Simmons, D.L. COX-3, a cyclooxygenase-1 variant inhibited by acetaminophen and other analgesic/antipyretic drugs: Cloning, structure, and expression. Proc. Natl. Acad. Sci. USA 2002, 9, 13926–13931. [Google Scholar]
- Hämäläinen, M.; Nieminen, R.; Asmawi, M.Z.; Vuorela, P.; Vapaatalo, H.; Moilanen, E. Effects of flavonoids on prostaglandin E2 production and on COX-2 and mPGES-1 expressions in activated macrophages. Planta Med. 2011, 77, 1504–1511. [Google Scholar]
- Lee, S.J.; Kavanaugh, A. Pharmacological treatment of established rheumatoid arthritis. Best Pract. Res. Clin. Rheumatol. 2003, 17, 811–829. [Google Scholar]
- Porter, S.N.; Howarth, G.S.; Butler, R.N. Non-steroidal anti-inflammatory drugs and apoptosis in the gastrointestinal tract: Potential role of the pentose phosphate pathways. Eur. J. Pharmacol. 2000, 397, 1–9. [Google Scholar] [CrossRef]
- Chandru, H.; Sharada, A.C.; Bettadaiah, B.K.; Kumar, C.S.A.; Rangappa, K.S.; Sunila; Jayashree, K. In vivo growth inhibitory and anti-angiogenic effects of synthetic novel dienone cyclopropoxy curcumin analogs on mouse Ehrlich ascites tumor. Bioorg. Med. Chem. 2007, 15, 7696–7703. [Google Scholar] [CrossRef]
- Weber, W.M.; Hunsaker, L.A.; Roybal, N.; Bobrovnikova-Marjon, E.V.; Abcouwer, S.F.; Royer, R.E.; Deck, L.M.; Vander Jagt, D.L. Activation of NFκB is inhibited by curcumin and related enones. Bioorg. Med. Chem. 2006, 14, 2450–2461. [Google Scholar] [CrossRef]
- Wei, Q.Y.; Ma, J.P.; Cai, Y.J.; Yang, L.; Liu, Z.L. Cytotoxic and apoptotic activities of iarylheptanoids and gingerol-related compounds from the rhizome of Chinese ginger. J. Ethnopharmacol. 2005, 102, 177–184. [Google Scholar] [CrossRef]
- Adam, B.K.; Ferstl, E.M.; Davis, M.C.; Herold, S.K.; Camalier, R.F.; Hollingshead, M.G.; Kaur, G.; Sausville, E.A.; Rickles, F.R.; Snyder, J.P.; et al. Synthesis and Biological evaluation of novel curcumin analogs as anti-cancer and anti-angiogensesis agents. Bioorg. Med. Chem. 2004, 12, 3871–3883. [Google Scholar] [CrossRef]
- Lee, K.H.; Aziz, F.H.A.; Abas, F.; Shaari, K.; Israf, D.A.; Lajis, N.H. Synthesis and biological evaluation of curcumin-like diarylpentanoid analogues for anti-inflammatory, antioxidant and anti-tyrosinase activities. Eur. J. Med. Chem. 2009, 44, 3195–3200. [Google Scholar] [CrossRef]
- Gilroy, D.W.; Colville-Nash, P.R. New insights into the role of COX 2 in inflammation. J. Mol. Med. 2000, 78, 121–129. [Google Scholar] [CrossRef]
- Matsuura, H.; Sakaue, M.; Subbaramaiahi, K.; Kamitani, H.; Eling, T.E.; Dannenbergi, A.J.; Tanabe, T.; Inoue, H. Arata, J.; Jetten, A.M. Regulation of Cyclooxygenase-2 by Interferon γ and Transforming Growth Factor α in Normal Human Epidermal Keratinocytes and Squamous Carcinoma Cells. J. Biol. Chem. 1999, 247, 29138–29148. [Google Scholar]
- Chen, J.X.; Berrym, L.C.; Christman, B.W.; Tanner, M.; Myers, P.R.; Meyrick, B.O. NO regulates LPS-stimulated cyclooxygenase gene expression and activity in pulmonary artery endothelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 280, L450–L457. [Google Scholar]
- Höcherl, K.; Dreher, F.; Kurtz, A.; Bucher, M. Cyclooxygenase-2 inhibition attenuates lipopolysaccharide-induced cardiovascular failure. Hypertension 2002, 40, 947–953. [Google Scholar] [CrossRef]
- Kinne, R.W.; Stuhlmüller, B.; Burmester, G.R. Cells of the synovium in rheumatoid arthritis Macrophages. Arthritis Res. Ther. 2007, 9, 224–240. [Google Scholar] [CrossRef]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase Isozymes: The Biology of Prostaglandin Synthesis and Inhibition. Pharmacol. Rev. 2004, 56, 387–437. [Google Scholar] [CrossRef]
- Ye, X; Liu, S.F. Lipopolysaccharide down-regulates promoter selective transcription factor 1 (Sp1) binding activity by promoting Sp1 protein dephosphorylation and degradation. J. Biol. Chem. 2002, 227, 31863–31870. [Google Scholar]
- Sample Availability: Samples of compound BDMC33 is available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, K.-H.; Abas, F.; Alitheen, N.B.M.; Shaari, K.; Lajis, N.H.; Ahmad, S. A Curcumin Derivative, 2,6-Bis(2,5-dimethoxybenzylidene)-cyclohexanone (BDMC33) Attenuates Prostaglandin E2 Synthesis via Selective Suppression of Cyclooxygenase-2 in IFN-g/LPS-Stimulated Macrophages. Molecules 2011, 16, 9728-9738. https://doi.org/10.3390/molecules16119728
Lee K-H, Abas F, Alitheen NBM, Shaari K, Lajis NH, Ahmad S. A Curcumin Derivative, 2,6-Bis(2,5-dimethoxybenzylidene)-cyclohexanone (BDMC33) Attenuates Prostaglandin E2 Synthesis via Selective Suppression of Cyclooxygenase-2 in IFN-g/LPS-Stimulated Macrophages. Molecules. 2011; 16(11):9728-9738. https://doi.org/10.3390/molecules16119728
Chicago/Turabian StyleLee, Ka-Heng, Faridah Abas, Noorjahan Banu Mohamed Alitheen, Khozirah Shaari, Nordin Haji Lajis, and Syahida Ahmad. 2011. "A Curcumin Derivative, 2,6-Bis(2,5-dimethoxybenzylidene)-cyclohexanone (BDMC33) Attenuates Prostaglandin E2 Synthesis via Selective Suppression of Cyclooxygenase-2 in IFN-g/LPS-Stimulated Macrophages" Molecules 16, no. 11: 9728-9738. https://doi.org/10.3390/molecules16119728
APA StyleLee, K.-H., Abas, F., Alitheen, N. B. M., Shaari, K., Lajis, N. H., & Ahmad, S. (2011). A Curcumin Derivative, 2,6-Bis(2,5-dimethoxybenzylidene)-cyclohexanone (BDMC33) Attenuates Prostaglandin E2 Synthesis via Selective Suppression of Cyclooxygenase-2 in IFN-g/LPS-Stimulated Macrophages. Molecules, 16(11), 9728-9738. https://doi.org/10.3390/molecules16119728