White Spot Syndrome Virus Orf514 Encodes a Bona Fide DNA Polymerase




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
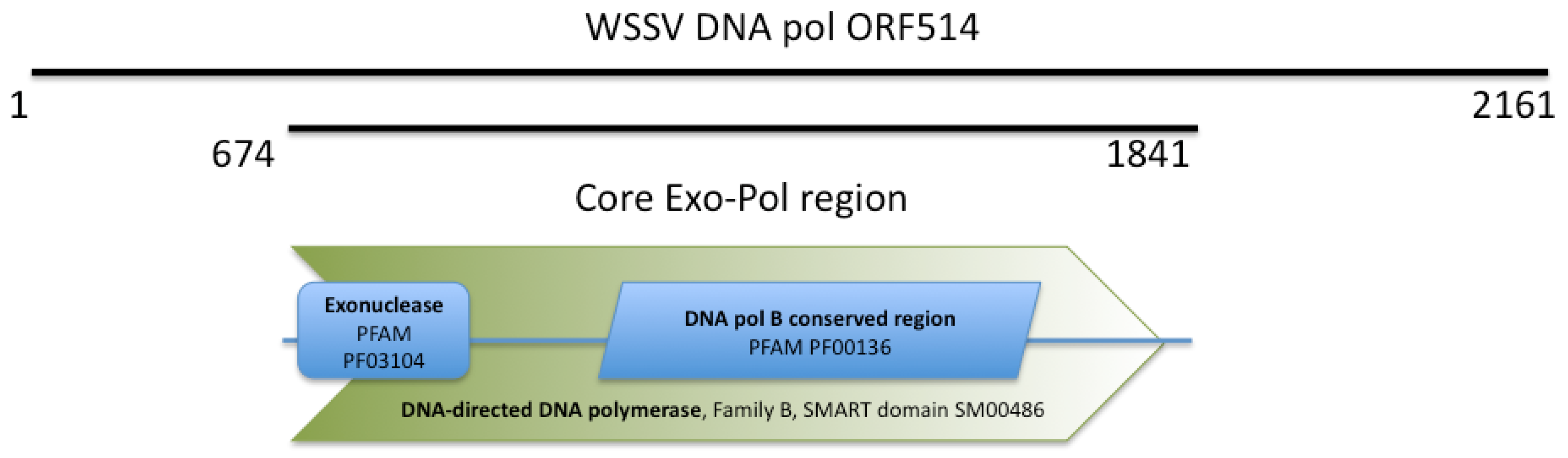
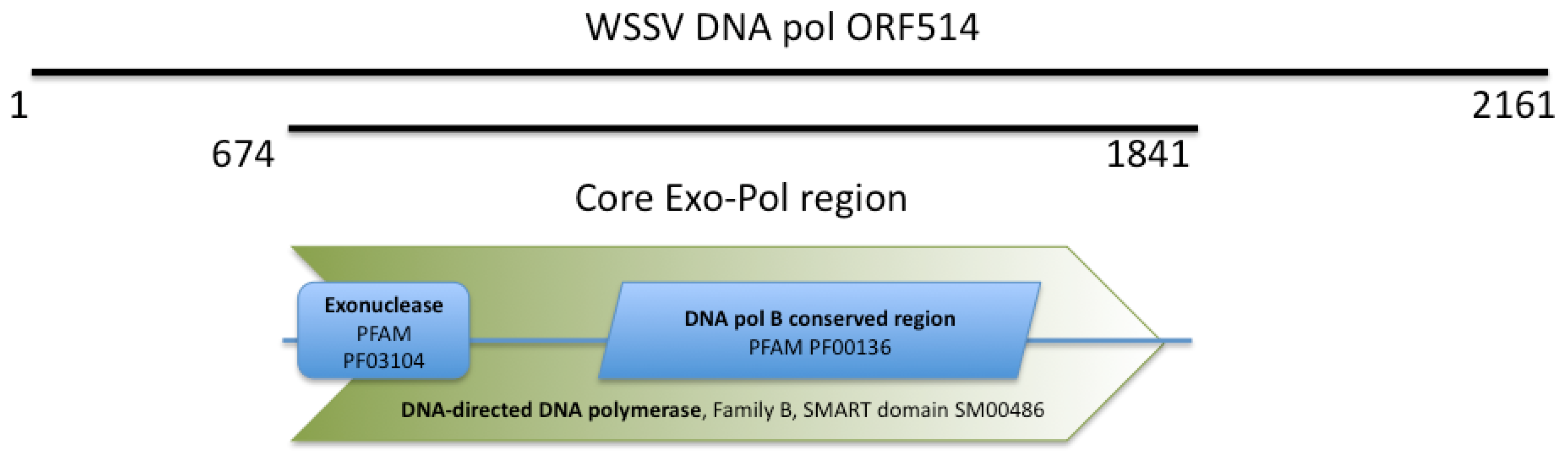
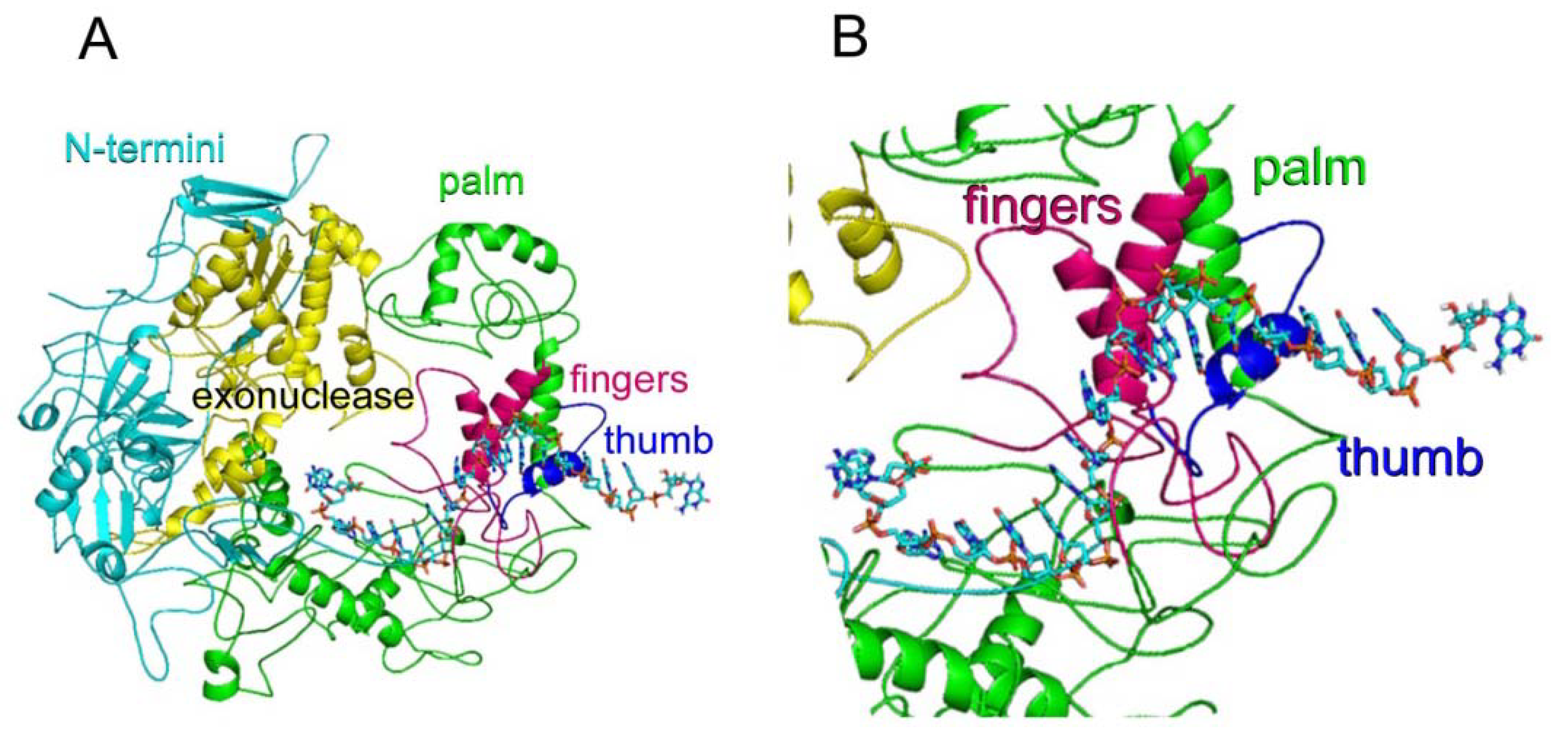
2.1. WSSV ORF514 Contains the Functional Domains of a Family B DNA Polymerase
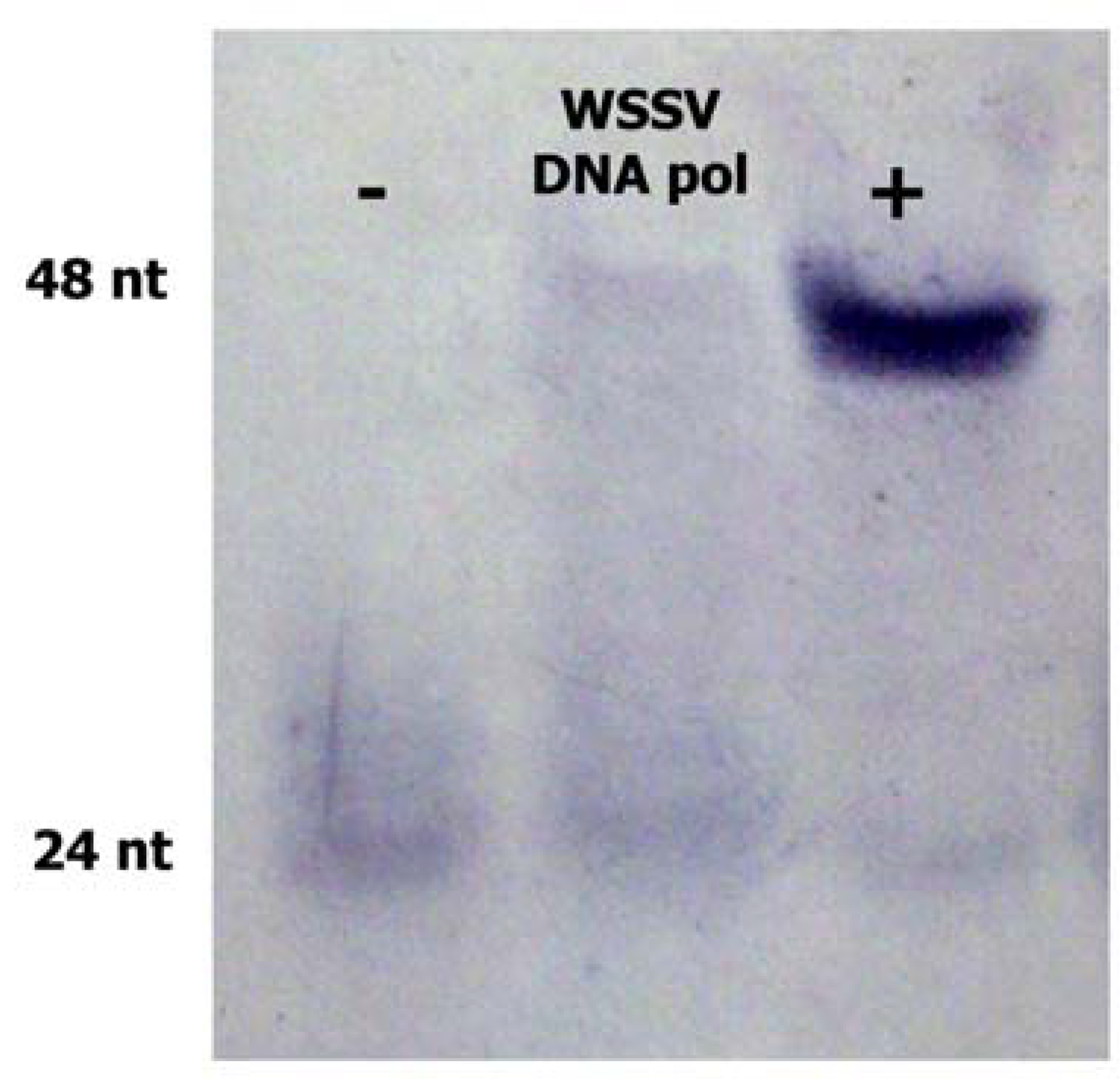
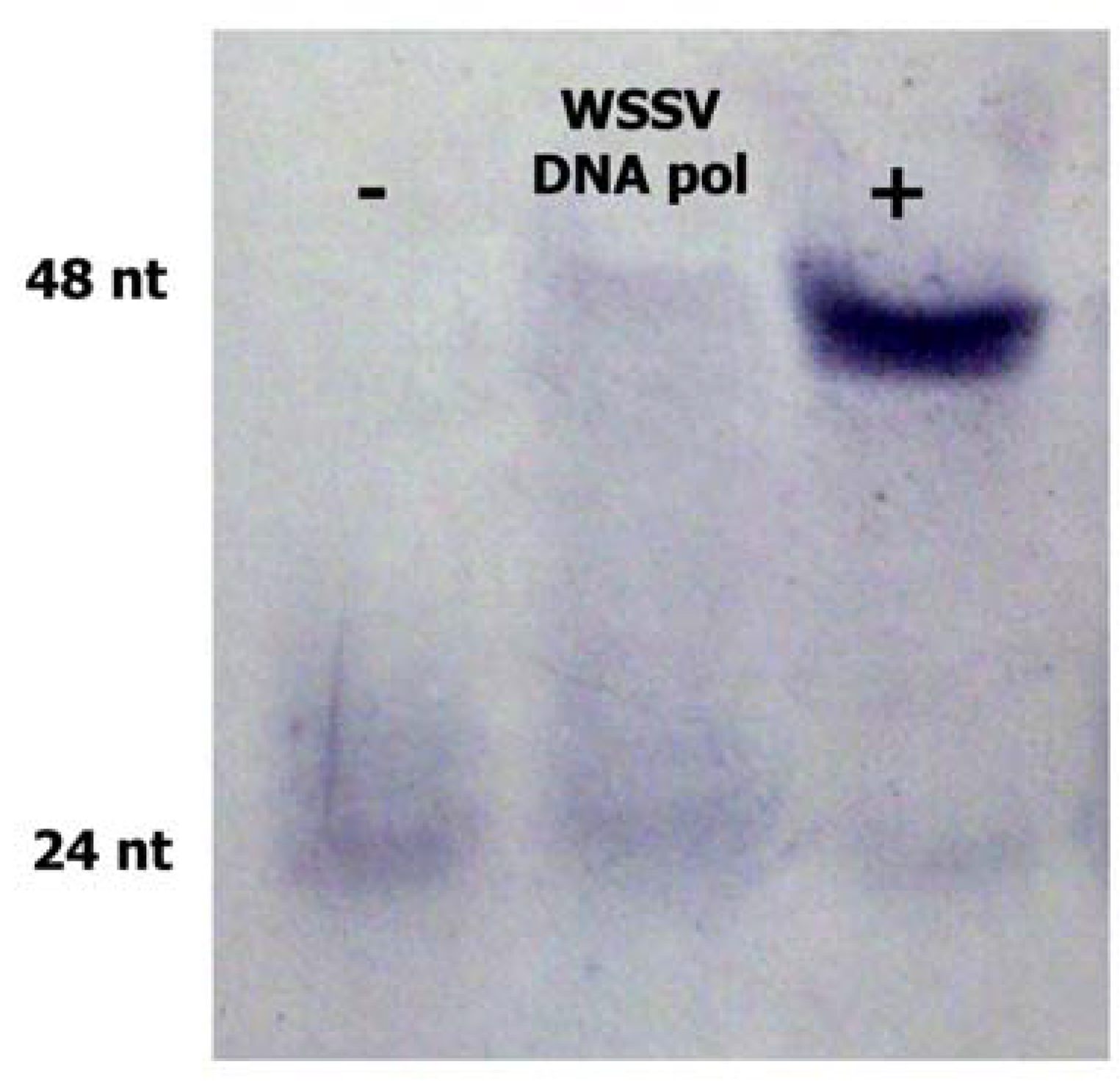
2.2. WSSV ORF514 has Polymerase Activity
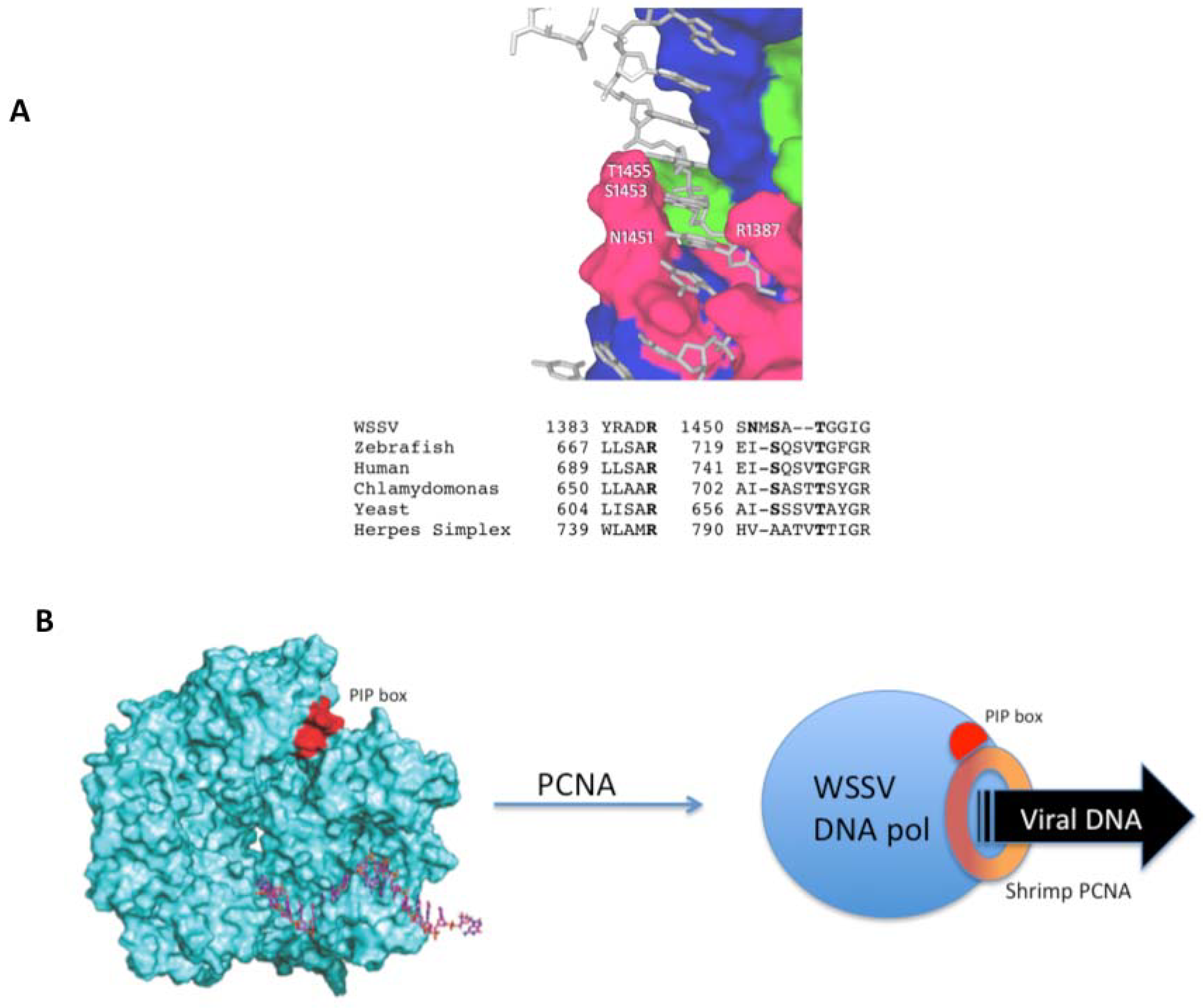
2.3. Interaction with Host Replication Factors
3. Experimental
3.1. Materials
3.2. WSSV ORF514 Expression Construct
3.3. Recombinant Expression of WSSV ORF514
3.4. WSSV ORF514 Protein Purification
3.5. DNA Polymerase Activity Assay
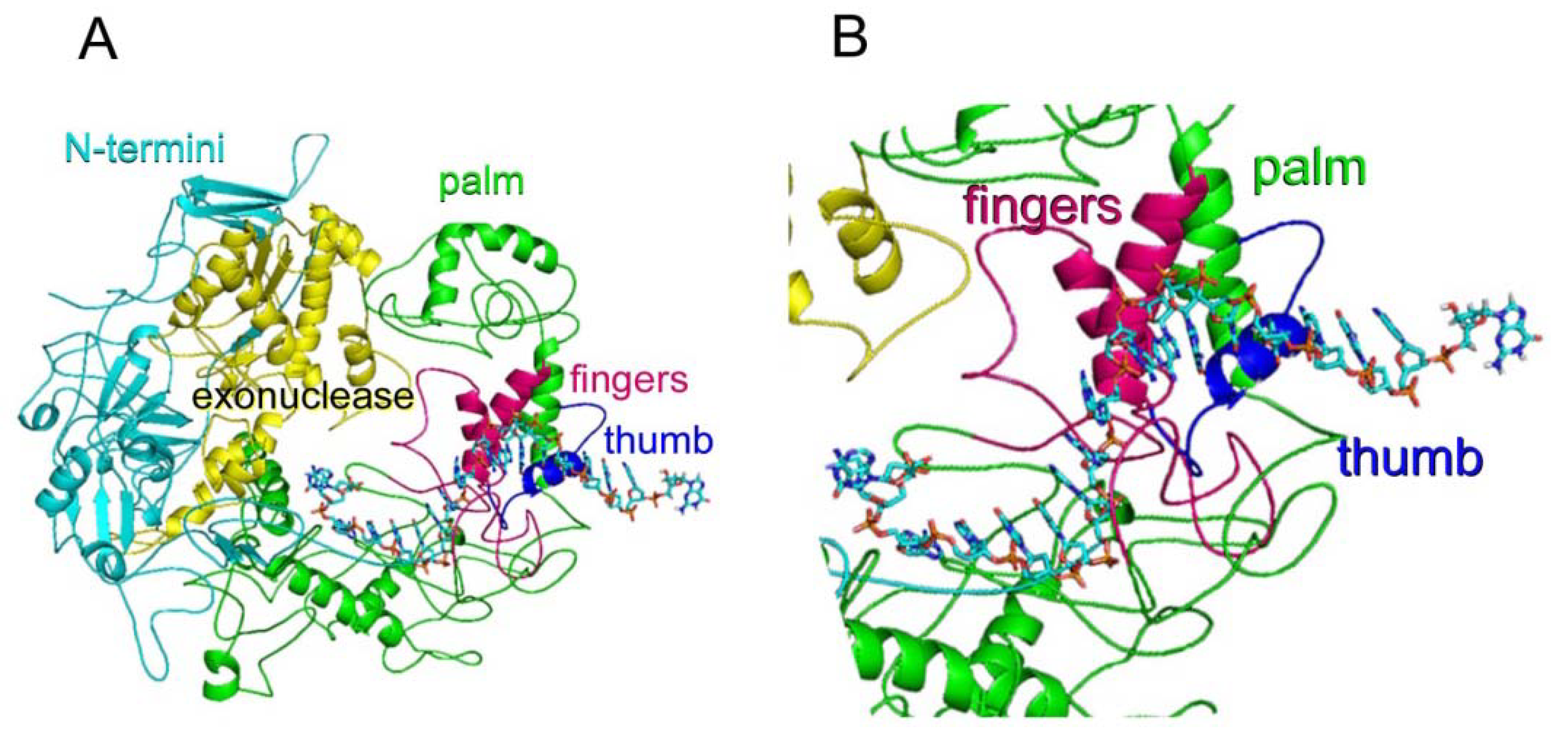
3.6. WSSV DNA ORF514 Encoded DNA Polymerase Molecular Modeling
4. Conclusions
Acknowledgements
Abbreviations
References and Notes
- Liu, H.; Soderhall, K.; Jiravanichpaisal, P. Antiviral immunity in crustaceans. Fish Shellfish Immunol. 2009, 27, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Paz, A. White spot syndrome virus: An overview on an emergent concern. Vet. Res. 2010, 41, 43. [Google Scholar] [CrossRef] [PubMed]
- van Hulten, M.C.; Witteveldt, J.; Peters, S.; Kloosterboer, N.; Tarchini, R.; Fiers, M.; Sandbrink, H.; Lankhorst, R.K.; Vlak, J.M. The white spot syndrome virus DNA genome sequence. Virology 2001, 286, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.M.; Wang, H.C.; Leu, J.H.; Wang, A.H.; Zhuang, Y.; Walker, P.J.; Kou, G.H.; Lo, C.F. Identification of the nucleocapsid, tegument, and envelope proteins of the shrimp white spot syndrome virus virion. J. Virol. 2006, 80, 3021–3029. [Google Scholar] [CrossRef] [PubMed]
- Arvizu-Flores, A.A.; Aispuro-Hernandez, E.; Garcia-Orozco, K.D.; Varela-Romero, A.; Valenzuela-Soto, E.; Velazquez-Contreras, E.F.; Rojo-Domínguez, A.; Yepiz-Plascencia, G.; Maley, F.; Sotelo-Mundo, R.R. Functional identity of the active sites of crustacean and viral thymidylate synthases. Comp. Biochem. Physiol. Pt. C 2009, 150C, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Wang, H.C.; Huang, C.J.; Peng, S.E.; Chen, Y.G.; Lin, S.J.; Chen, W.Y.; Dai, C.F.; Yu, H.T.; Wang, C.H. Transcriptional analysis of the DNA polymerase gene of shrimp white spot syndrome virus. Virology 2002, 301, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Joyce, C.M.; Steitz, T.A. Polymerase structures and function: variations on a theme? J. Bacteriol. 1995, 177, 6321–6329. [Google Scholar] [CrossRef] [PubMed]
- Steitz, T.A. DNA polymerases: structural diversity and common mechanisms. J. Biol.Chem. 1999, 274, 17395–17398. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, M.; Parris, D.S. Evidence against a simple tethering model for enhancement of herpes simplex virus DNA polymerase processivity by accessory protein UL42. J. Virol. 2002, 76, 10270–10281. [Google Scholar] [CrossRef] [PubMed]
- Digard, P.; Bebrin, W.R.; Weisshart, K.; Coen, D.M. The extreme C terminus of herpes simplex virus DNA polymerase is crucial for functional interaction with processivity factor UL42 and for viral replication. J. Virol. 1993, 67, 398–406. [Google Scholar] [PubMed]
- Weisshart, K.; Chow, C.S.; Coen, D.M. Herpes simplex virus processivity factor UL42 imparts increased DNA-binding specificity to the viral DNA polymerase and decreased dissociation from primer-template without reducing the elongation rate. J. Virol. 1999, 73, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Seybert, A.; Scott, D.J.; Scaife, S.; Singleton, M.R.; Wigley, D.B. Biochemical characterisation of the clamp/clamp loader proteins from the euryarchaeon Archaeoglobus fulgidus. Nucl. Acid. Res. 2002, 30, 4329–4338. [Google Scholar] [CrossRef]
- Warbrick, E. PCNA binding through a conserved motif. BioEssays 1998, 20, 195–199. [Google Scholar] [CrossRef]
- Tori, K.; Kimizu, M.; Ishino, S.; Ishino, Y. DNA polymerases BI and D from the hyperthermophilic archaeon Pyrococcus furiosus both bind to proliferating cell nuclear antigen with their C-terminal PIP-box motifs. J. Bacteriol. 2007, 189, 5652–5657. [Google Scholar] [CrossRef] [PubMed]
- Huber, H.E.; Tabor, S.; Richardson, C.C. Escherichia coli thioredoxin stabilizes complexes of bacteriophage T7 DNA polymerase and primed templates. J. Biol.Chem. 1987, 262, 16224–16232. [Google Scholar] [PubMed]
- Xie, Y.; Wang, B.; Li, F.; Jiang, H.; Xiang, J. Molecular cloning and characterization of proliferating cell nuclear antigen (PCNA) from Chinese shrimp Fenneropenaeus chinensis. Comp. Biochem. Physiol. Pt. B 2008, 151B, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Welch, M.; Govindarajan, S.; Ness, J.E.; Villalobos, A.; Gurney, A.; Minshull, J.; Gustafsson, C. Design parameters to control synthetic gene expression in Escherichia coli. PLoS One 2009, 4, e7002. [Google Scholar] [CrossRef] [PubMed]
- Brieba, L.G.; Eichman, B.F.; Kokoska, R.J.; Doublie, S.; Kunkel, T.A.; Ellenberger, T. Structural basis for the dual coding potential of 8-oxoguanosine by a high-fidelity DNA polymerase. EMBO J. 2004, 23, 3452–3461. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Knafels, J.D.; Chang, J.S.; Waszak, G.A.; Baldwin, E.T.; Deibel, M.R.; Thomsen, D.R.; Homa, F.L.; Wells, P.A.; Tory, M.C. Crystal structure of the herpes simplex virus 1 DNA polymerase. J. Biol. Chem. 2006, 281, 18193–18200. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.C.; Wang, J.; Steitz, T.A. Structure of the replicating complex of a pol [alpha] family DNA polymerase. Cell 2001, 105, 657–667. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Wu, Y.; L, L.; Yang, L.S.; Weng, S.P.; Chan, S.M.; He, J.G. Inhibition of white spot syndrome virus in Litopenaeus vannamei shrimp by sequence-specific siRNA. Aquaculture 2007, 271, 21–30. [Google Scholar] [CrossRef]
- Tzeng, H.F.; Chang, Z.F.; Peng, S.E.; Wang, C.H.; Lin, J.Y.; Kou, G.H.; Lo, C.F. Chimeric polypeptide of thymidine kinase and thymidylate kinase of shrimp white spot syndrome virus: thymidine kinase activity of the recombinant protein expressed in a baculovirus/insect cell system. Virology 2002, 299, 248–255. [Google Scholar] [CrossRef] [PubMed]

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De-la-Re-Vega, E.; Garcia-Orozco, K.D.; Arvizu-Flores, A.A.; Yepiz-Plascencia, G.; Muhlia-Almazan, A.; Hernández, J.; Brieba, L.G.; Sotelo-Mundo, R.R. White Spot Syndrome Virus Orf514 Encodes a Bona Fide DNA Polymerase. Molecules 2011, 16, 532-542. https://doi.org/10.3390/molecules16010532
De-la-Re-Vega E, Garcia-Orozco KD, Arvizu-Flores AA, Yepiz-Plascencia G, Muhlia-Almazan A, Hernández J, Brieba LG, Sotelo-Mundo RR. White Spot Syndrome Virus Orf514 Encodes a Bona Fide DNA Polymerase. Molecules. 2011; 16(1):532-542. https://doi.org/10.3390/molecules16010532
Chicago/Turabian StyleDe-la-Re-Vega, Enrique, Karina D. Garcia-Orozco, Aldo A. Arvizu-Flores, Gloria Yepiz-Plascencia, Adriana Muhlia-Almazan, Jesús Hernández, Luis G. Brieba, and Rogerio R. Sotelo-Mundo. 2011. "White Spot Syndrome Virus Orf514 Encodes a Bona Fide DNA Polymerase" Molecules 16, no. 1: 532-542. https://doi.org/10.3390/molecules16010532
APA StyleDe-la-Re-Vega, E., Garcia-Orozco, K. D., Arvizu-Flores, A. A., Yepiz-Plascencia, G., Muhlia-Almazan, A., Hernández, J., Brieba, L. G., & Sotelo-Mundo, R. R. (2011). White Spot Syndrome Virus Orf514 Encodes a Bona Fide DNA Polymerase. Molecules, 16(1), 532-542. https://doi.org/10.3390/molecules16010532