High Molecular Weight Glucan of the Culinary Medicinal Mushroom Agaricus bisporus is an α-Glucan that Forms Complexes with Low Molecular Weight Galactan


Abstract
:1. Introduction
2. Results and Discussion
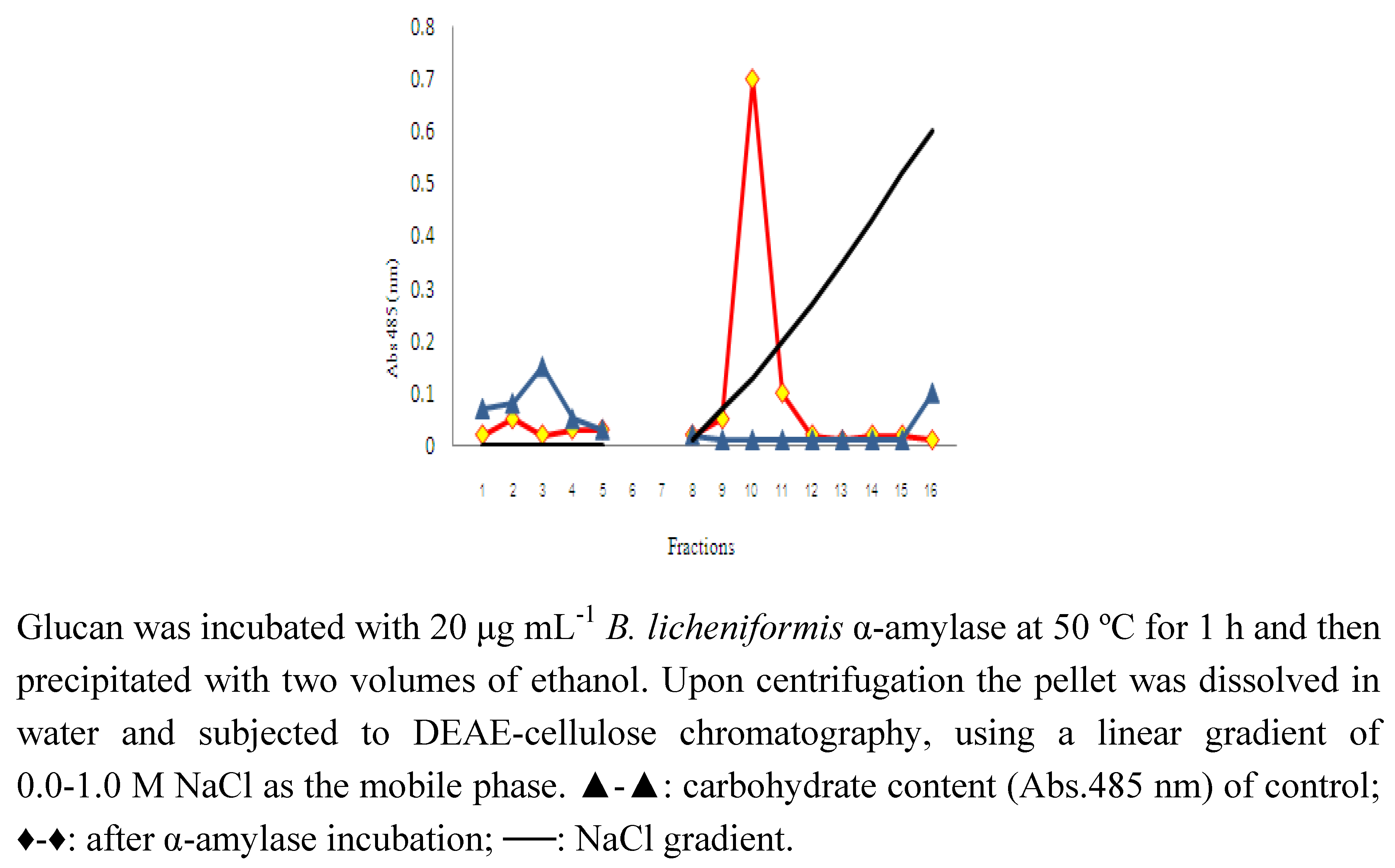

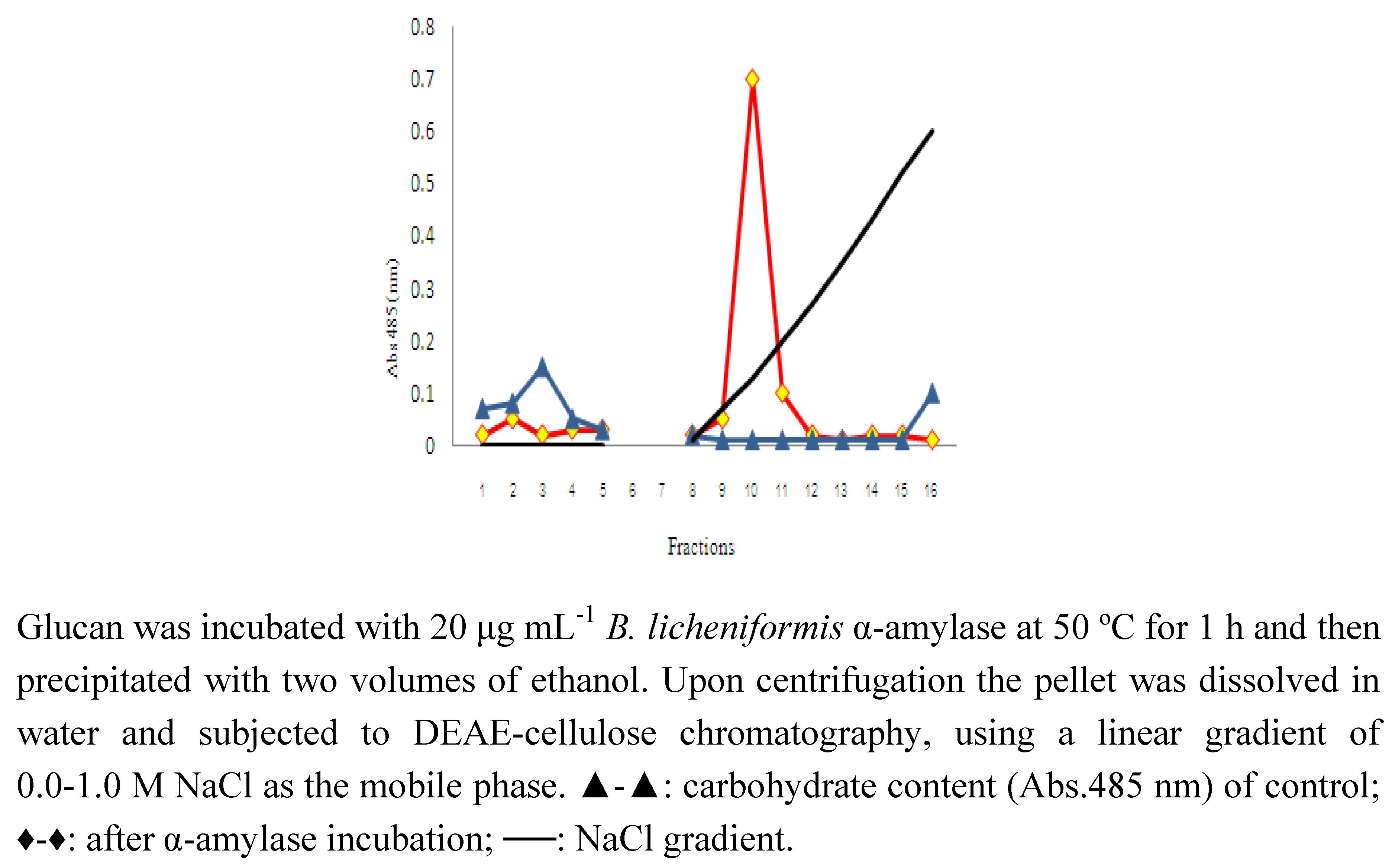{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosaccharides (%) | |||||
|---|---|---|---|---|---|
| Glucose | Mannose | Galactose | Arabinose | Ribose | Rhamnose |
| 90.1 | - | 9.8 | Tr. | Tr. | Tr. |
| Partially O-methylated alditol acetates (a) | Mol % | Linkage Type (c) |
|---|---|---|
| Sample (b) | ||
| 2,3,4,6-Me4-Glcp | 9.9 | Glcp-(1→ |
| 2,3,6-Me3-Glcp | 72.5 | 4→)-Glcp-(1→ |
| 2,3,4-Me3-Galp | 9.3 | 6→)-Galp-(1→ |
| 2,3-Me2-Glcp | 8.3 | 4,6→)-Glcp-(1→ |
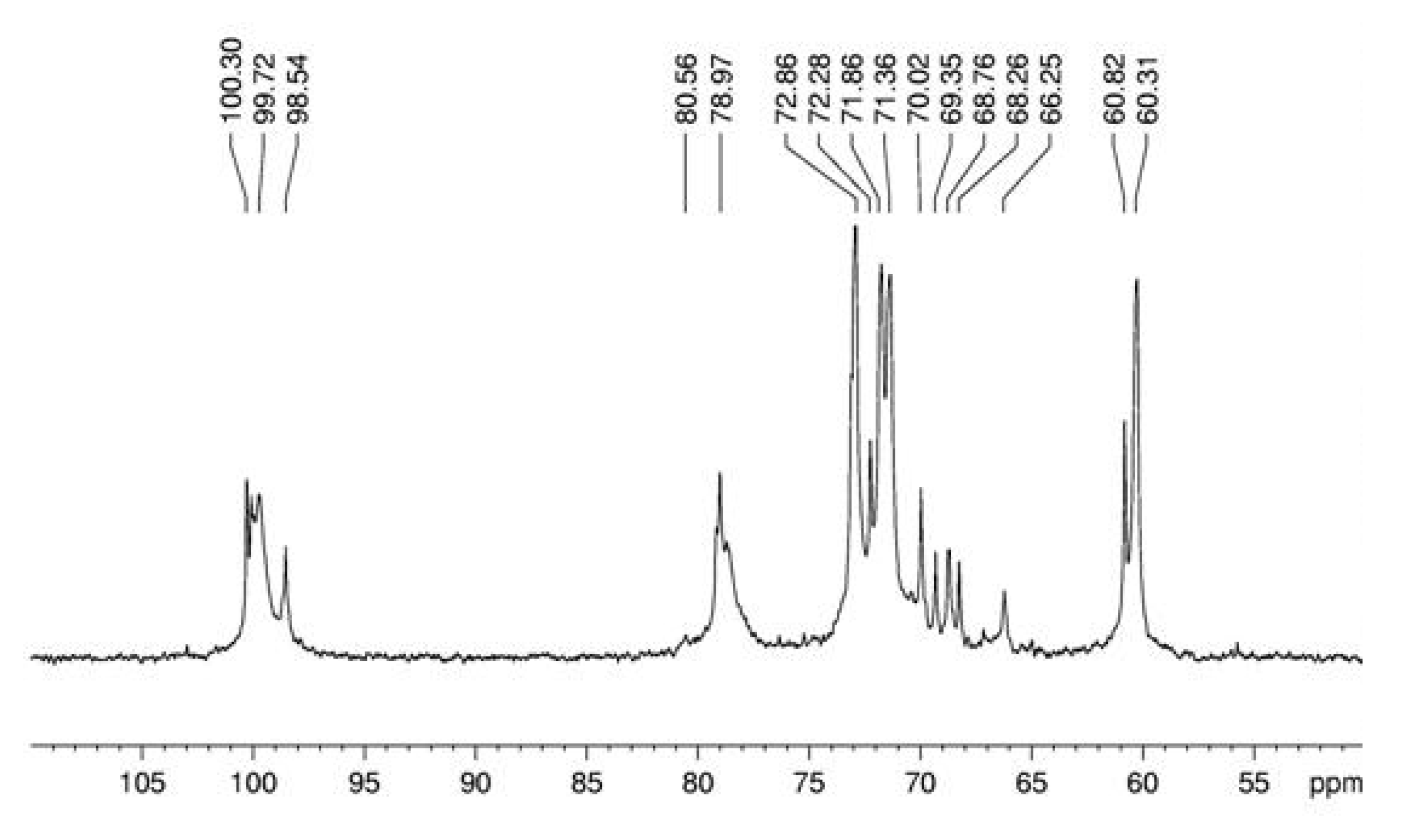
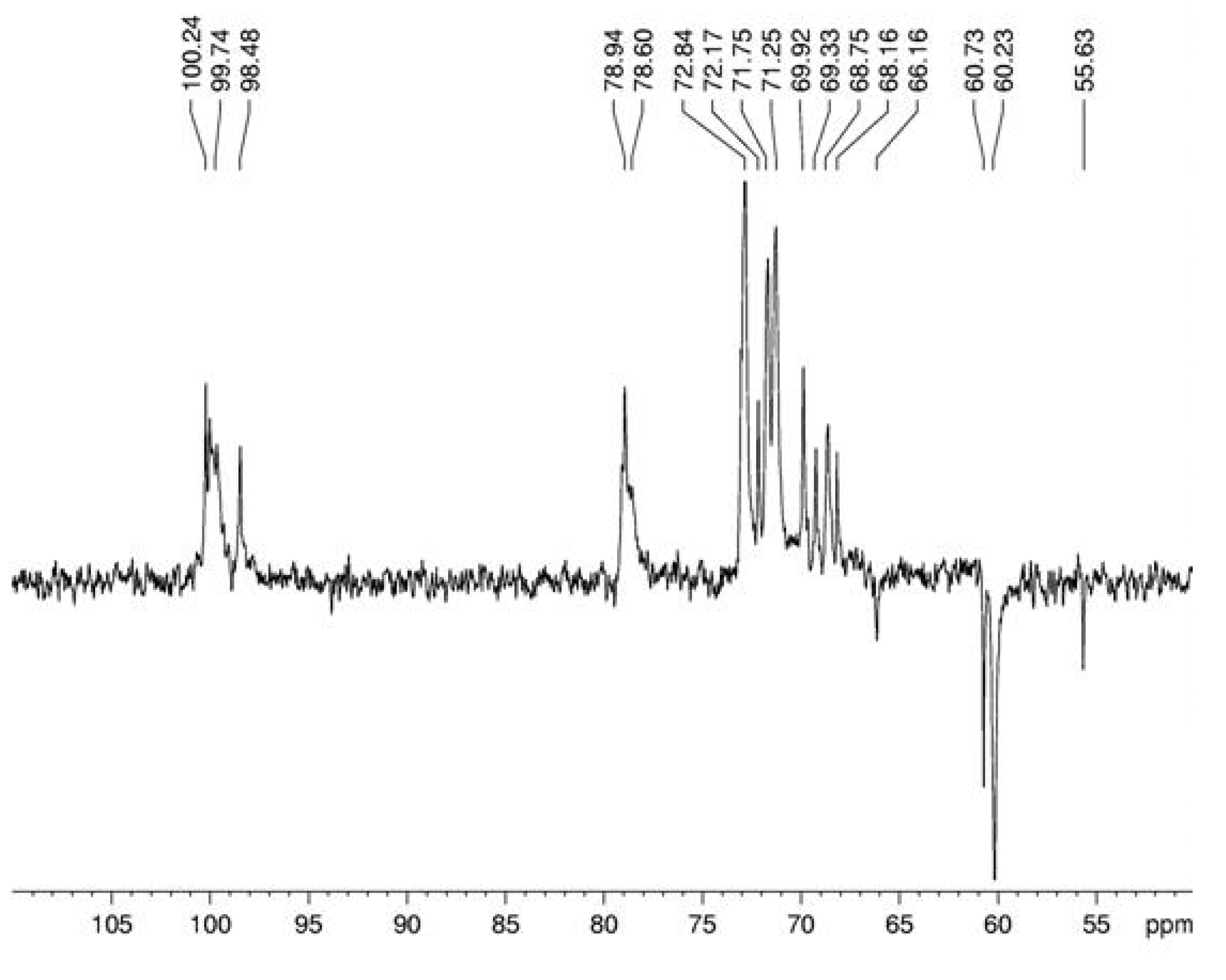
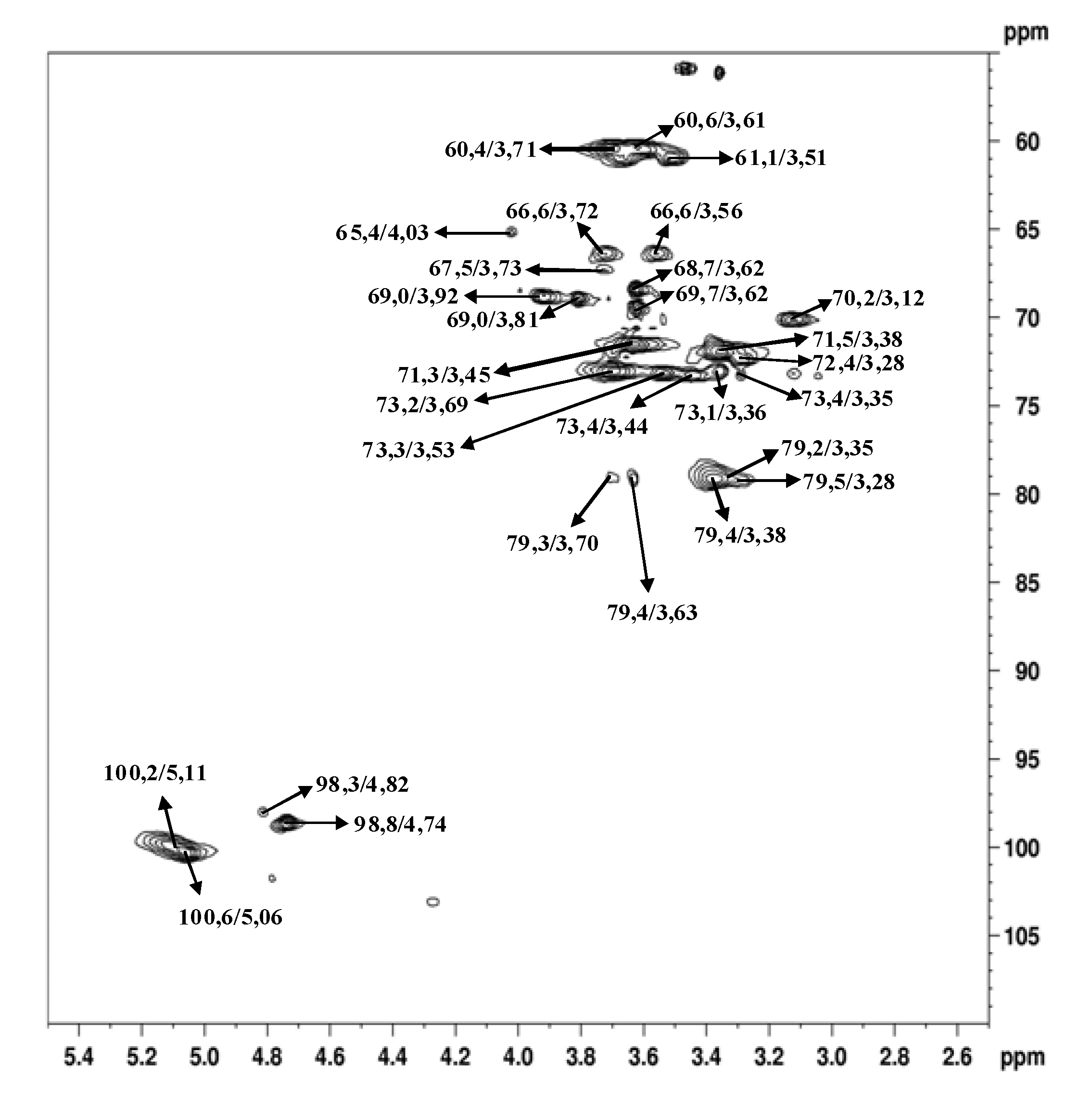
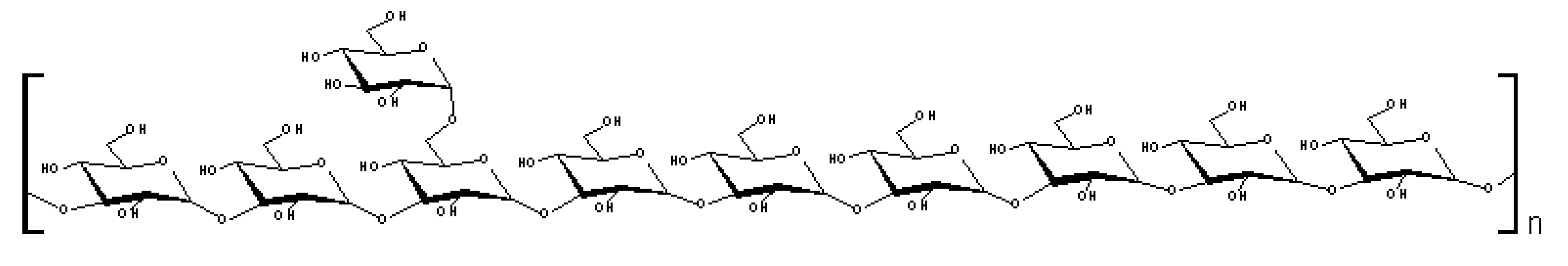
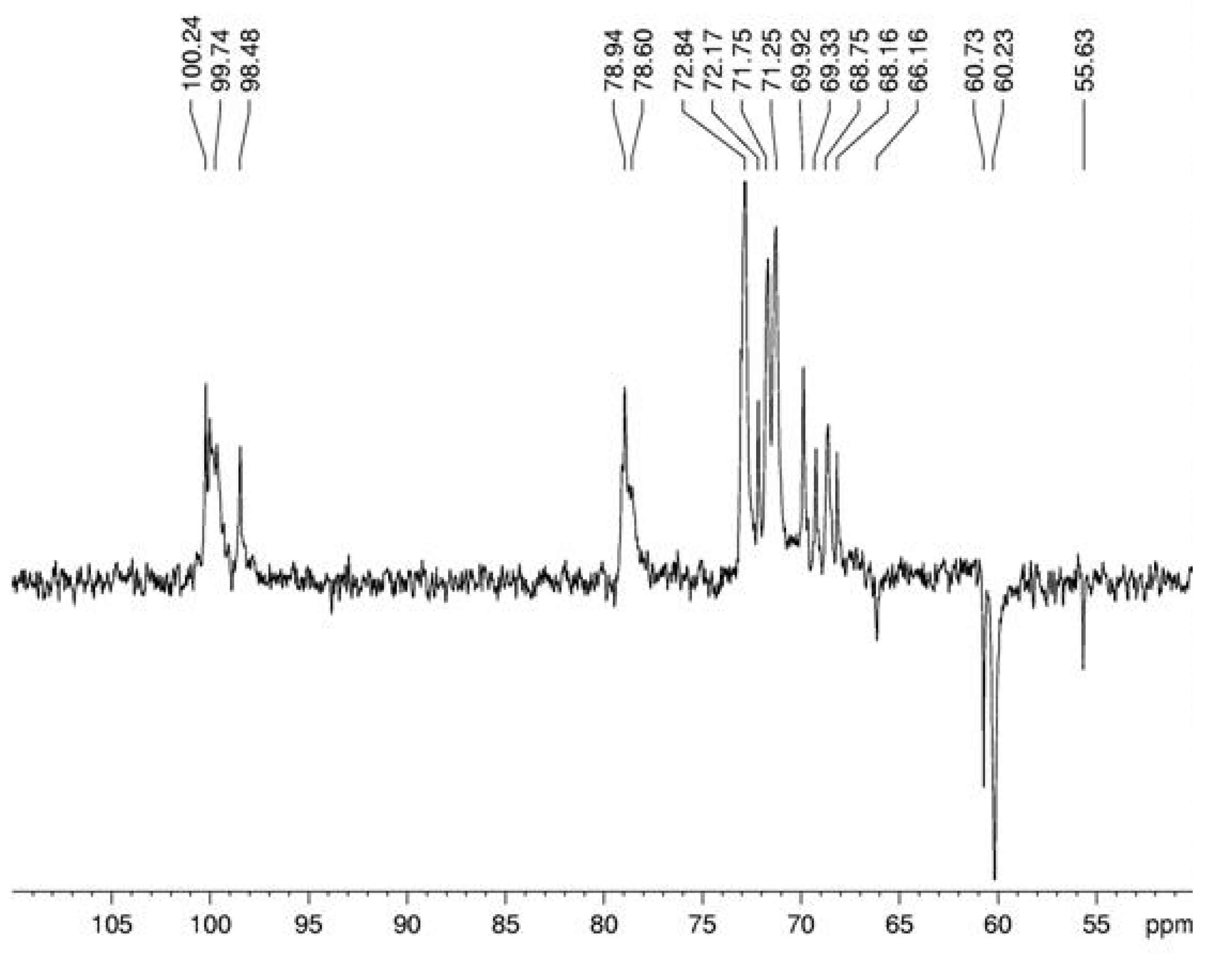
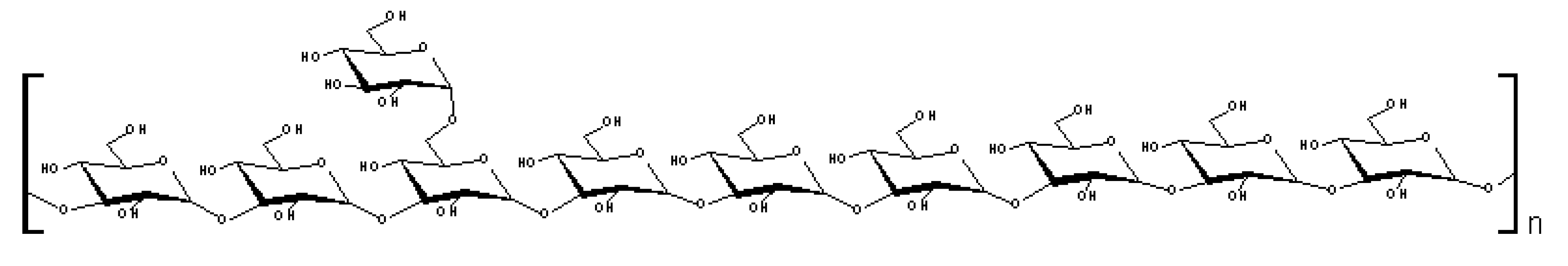
3. Experimental
3.1. Polysaccharides
3.2. General Experimental Procedures
3.3. Analyses and Determinations
3.3.1. Carbohydrate Determination
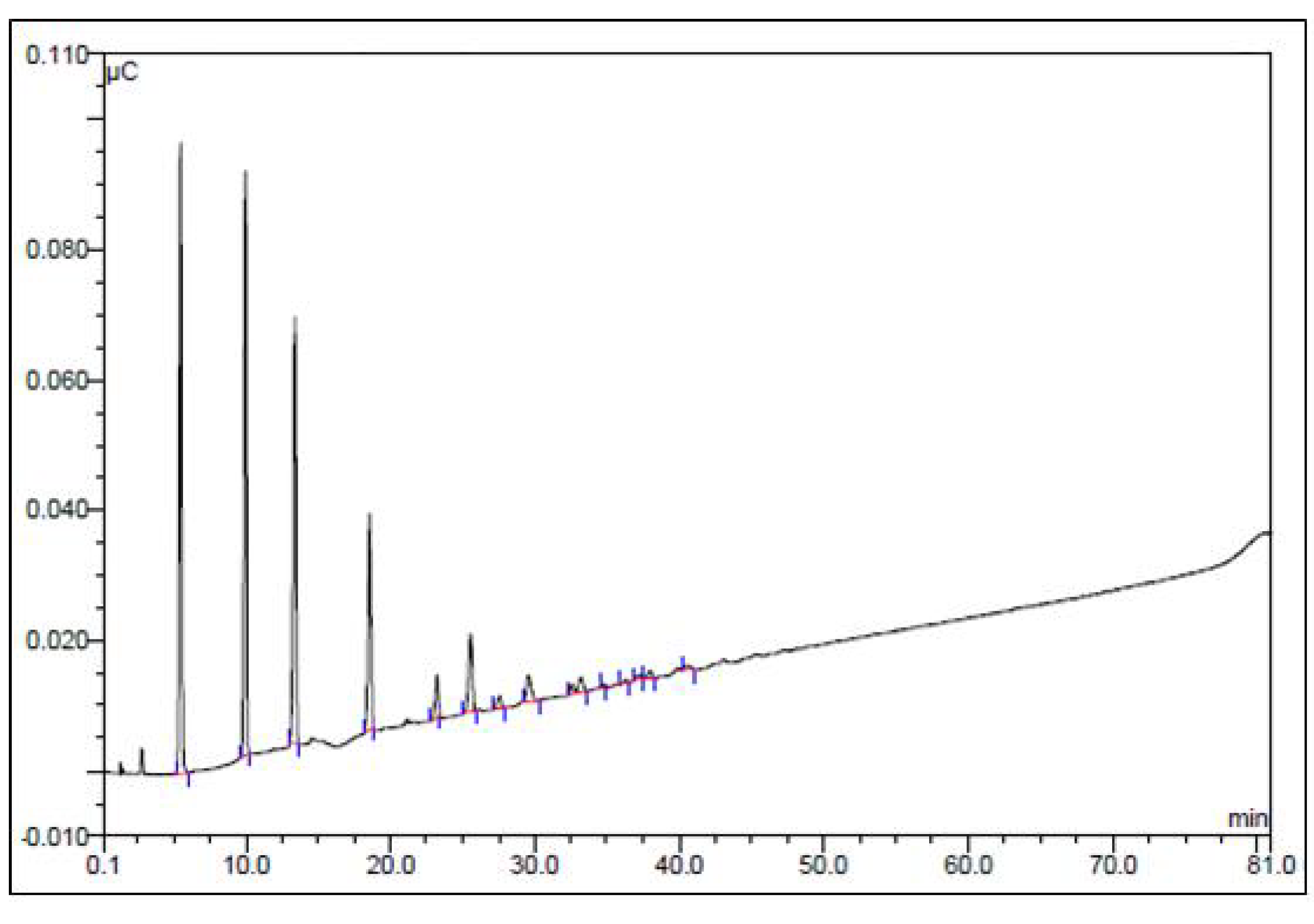
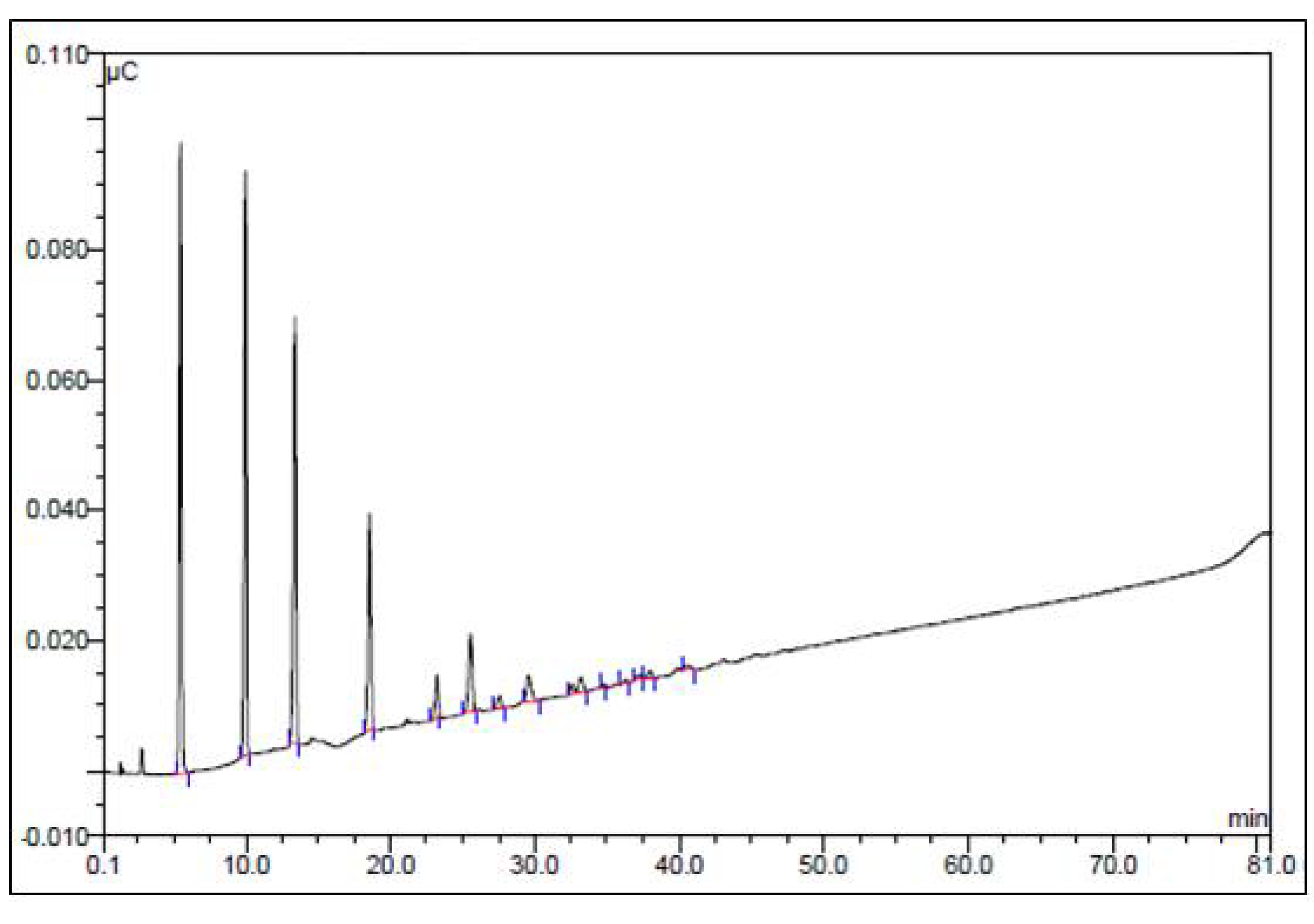
3.3.2. Analysis of Monosaccharide Composition by GC-MS
3.3.3. Analysis of Monosaccharide Composition by HPLC
3.3.4. Analysis of Sugar Oligomers
3.3.5. Spectroscopy Analyses
4. Conclusions
Acknowledgments
- Samples Availability: Samples of A. bisporus α-glucan are available from the corresponding author.
References
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef]
- Borchers, A.T.; Keen, C.L.; Gershwin, M.E. Mushrooms, tumors, and immunity: an update. Exp. Biol. Med. (Maywood). 2004, 229, 393–406. [Google Scholar]
- Lull, C.H.J.; Wichers, H.J.; Savelkoul, H.F. Antiinflammatory and immunomodulating properties of fungal metabolites. Mediat. Inflamm. 2005, 63–80. [Google Scholar]
- Zaidman, B.Z.; Yassin, M.; Mahajna, J.; Wasser, S.P. Medicinal mushroom modulators of molecular targets as cancer therapeutics. Appl. Microbiol. Biotechnol. 2005, 67, 453–468. [Google Scholar] [CrossRef]
- Akramiene, D.; Kondrotas, A.; Didziapetriene, J.; Kevelaitis, E. Effects of beta-glucans on the immune system. Medicina (Kaunas) 2007, 43, 597–606. [Google Scholar]
- Rop, O.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef]
- Wichers, H. Immunomodulation by Food: Promising concept for mitigating allergic disease? Anal. Bioanal. Chem. 2009, 395, 37–45. [Google Scholar] [CrossRef]
- Soltanian, S.; Stuyven, E.; Cox, E. . Beta-glucans as immunostimulant invertebrates and invertebrate. Crit. Rev. Microbiol. 2009, 35, 109–138. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Xu, X.; Zeng, F. Correlation between antitumor activity, molecular weight, and conformation of lentinan. Carbohydr. Res. 2005, 340, 1515–1521. [Google Scholar]
- Kojima, H.; Akaki, J.; Nakajima, S.; Kamei, K.; Tamesada, M. Structural analysis of glycogen-like polysaccharides having macrophage-activating activity in extracts. J. Nat. Med. 2010, 64, 16–23. [Google Scholar] [CrossRef]
- Kakutani, R.; Adachi, Y.; Kajiura, H.; Takata, H.; Kuriki, T.; Ohno, N. 2007. Relationship between structure and immunostimulating activity of enzymatically synthesized glycogen. Carbohydr. Res. 2007, 342, 2371–2379. [Google Scholar] [CrossRef]
- Wu, D.; Pae, M.; Ren, Z.; Guo, Z.; Smith, D.; Meydani, S.N. Dietary supplementation with white button mushroom enhances natural killer cell activity in C57BL/6 mice. J. Nutr. 2007, 137, 1472–1477. [Google Scholar]
- Jeong, S.C.; Jeong, Y.T.; Yang, B.K.; Islam, R.; Koyyalamudi, S.R.; Pang, G.; Cho, K.Y.; Song, C.H. White button mushroom (Agaricus bisporus) lowers blood glucose and cholesterol levels in diabetics and hypercholesterolemic rats. Nutr. Res. 2010, 30, 49–56. [Google Scholar] [CrossRef]
- Volman, J.J.; Mensink, R.P.; Van Griensven, L.J.L.D.; Plat, J. Effects of α-glucans from Agaricus bisporus on ex vivo cytokine production by LPS and PHA-stimulated PBMCs; a placebo-controlled study in slightly hypercholesterolemic subjects. Eur. J. Clin. Nutr. 2010, 1–7. [Google Scholar]
- Volman, J.J.; Helsper, J.P.F.G.; Wei, S.; Baars, J.J.P.; Van Griensven, L.J.L.D.; Sonnenberg, A.S.M.; Mensink, R.P.; Plat, J. Effects of mushroom derived beta-glucan-rich polysaccharide extracts on NO production by bone marrow-derived macrophages and NF-κB transactivation in Caco-2 reporter cells: can effects be explained by structure? Mol. Nutr. Food Res. 2010, 54, 268–276. [Google Scholar]
- Wei, S.; Van Griensven, L.J.L.D. Pro- and antioxidative properties of medicinal mushroom extracts. Int. J. Med. Mushrooms 2008, 10, 315–324. [Google Scholar] [CrossRef]
- Jagadish, L.K.; Venkatakrishnan, V.; Shenbhagaraman, R.; Kaviyarasan, V. Comparative study on the antioxidant, anticancer and antimicrobial property of Agaricus bisporus (J.E. Lange) Imbach before and after boiling. Afr. J. Biotechnol. 2009, 8, 654–661. [Google Scholar]
- Shnyreva, A.V.; Wei, S.; Van Griensven, L.J.L.D. Extracts of Agaricus bisporus and Phellinus linteus induce pro-apoptotic effects in the human cancer cell line K562. Int. J. Med. Mushrooms 2010, 12, 167–175. [Google Scholar] [CrossRef]
- Yamac, M.; Kanbak, G.; Zeytinoglu, M.; Senturk, H.; Bayramoglu, G.; Dokumacioglu, A.; Van Griensven, L.J.L.D. Pancreas protective effect of antioxidative Agaricus bisporus extract on rats with streptozotocin induced diabetes. Int. J. Med. Mushrooms 2010, 12. in press. [Google Scholar]
- Wei, S.; Helsper, J.P.F.G.; Van Griensven, L.J.L.D. Phenolic Compounds Present in Medicinal Mushroom Extracts Generate Reactive Oxygen Species in Human Cells In Vitro. Int. J. Med. Mushrooms 2008, 10, 1–13. [Google Scholar] [CrossRef]
- Matsui, M.; Kakut, M.; Misaki, A. Fine structural features of oyster glycogen: mode of multiple branching. Carbohydr. Polym. 1996, 31, 227–235. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, S.W.; Cheung, P.C.K.; Wang, Q. Antitumor polysaccharides from mushrooms: a review in their isolation process, structural characteristics and antitumor activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Pang, X.; Yao, W.; Yang, X.; Xie, C.; Liu, D.; Zhang, J.; Gao, X. Purification, characterization and biological activity on hepatocytes of a polysaccharide from Flammulina velutipes mycelium. Carbohydr. Polym. 2007, 70, 291–297. [Google Scholar] [CrossRef]
- Gorin, P.A.J. Carbon-13 nuclear magnetic resonance spectroscopy of polysaccharides. Adv. Carbohydr. Chem. Biochem. 1981, 38, 13–104. [Google Scholar] [CrossRef]
- Perlin, A.S.; Casu, B. Carbon-13 and proton magnetic resonance spectra of D-glucose-13C. Tetrahedron Lett. 1969, 34, 2919–2924. [Google Scholar]
- Stanek, M.; Falk, H.; Huber, A. Investigation of the branchingcharacteristic of glycogen by means of two-dimensional 1H and 13C NMR spectroscopy. Monatsh. Chem. 1998, 129, 355–364. [Google Scholar]
- Wilson, W.A.; Hughes, W.E.; Tomamichel, W.; Roach, P.J. Increased glycogen storage in yeast results in less branched glycogen. Biochem. Biophys. Res. Commun. 2004, 320, 416–423. [Google Scholar]
- Kalac, P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. Food Chem. 2009, 113, 9–16. [Google Scholar] [CrossRef]
- Rosado, F.R.; Carbonero, E.R.; Claudino, R.F.; Tischer, C.A.; Kemmelmeier, C.; Iacomini, M. The presence of partially 3-O-methylated mannogalactan from the fruit bodies of edible basidiomycetes Pleurotus ostreatus ‘florida’ Berk. and Pleurotus ostreatoroseus Sing. FEMS Microbiol. Lett. 2003, 221, 119–124. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Carbonero, E.R.; Mellinger, C.G.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Structural characterization of a polysaccharide and a b-glucan isolated from the edible mushroom Flammulina velutipes. Phytochemistry 2006, 67, 2189–2196. [Google Scholar] [CrossRef]
- Carbonero, E.R.; Gracher, A.H.P.; Komura, D.L.; Marcon, R.; Freitas, C.S.; Baggio, C.H.; Santos, A.R.S.; Torri, G.; Gorin, P.A.J.; Iacomini, M. Lentinus edodes heterogalactan: Antinociceptive and anti-inflammatory effects. Food Chem. 2008, 111, 531–537. [Google Scholar]
- Roy, S.K.; Maiti, D.; Mondal, S.; Das, D.; Islam, S.S. Structural analysis of a polysaccharide isolated from the aqueous extract of an edible mushroom, Pleurotus sajor-caju, cultivar Black Japan. Carbohydr. Res. 2008, 343, 1108–1113. [Google Scholar] [CrossRef]
- Lu, M.K.; Cheng, J.J.; Lin, C.Y.; Chang, C.C. Purification, structural elucidation, and anti-inflammatory effect of a water-soluble 1,6-branched 1,3-α-D-galactan from cultured mycelia of Poria coco. Food Chem. 2010, 118, 349–356. [Google Scholar] [CrossRef]
- Rosado, F.R.; Carbonero, E.R.; Kemmelmeier, C.; Tischer, C.A.; Gorin, P.A.J.; Iacomini, M. A partially 3-O-methylated (1→4)-linked α-D-galactan and α-D-mannan from Pleurotus ostreatoroseus Sing. FEMS Microbiol. Lett. 2002, 212, 261–265. [Google Scholar]
- Carbonero, E.R.; Gracher, A.H.P.; Rosa, M.C.C.; Torri, G.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Unusual partially 3-O-methylated α-galactan from mushrooms of the genus Pleurotus. Phytochemistry 2008, 69, 252–257. [Google Scholar]
- Umar, M.H.; Van Griensven, L.J.L.D. Hyphal regeneration and histogenesis in Agaricus bisporus. Mycol. Res. 1997, 101, 1025–1032. [Google Scholar] [CrossRef]
- Sato, T.; Norisuye, T.; Fujita, H. Triple helix of Schizophyllum commune polysaccharide in dilute solution. 5. Light scattering and refractometry in mixtures of water and dimethyl sulfoxide. Macromolecules 1983, 16, 185–189. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Moradali, M.F.; Mostafavi, H.; Ghods, S.; Hedjaroude, G.A. Immunomodulating and anticancer agents in the realm of macromycetes fungi (macrofungi). Int. Immunopharmacol. 2007, 7, 701–724. [Google Scholar] [CrossRef]
- Dubois, M.; Giles, K.A.; Hamilton, G.K.; Rebers, P.A.; Smith, F. Colorimetric method for the determination of sugars and related substances. Anal. Chem. 1956, 28, 350–56. [Google Scholar]
- Sassaki, G.L.; Souza, L.M.; Serrato, R.V.; Cipriani, T.R.; Gorin, P.A.J.; Iacomini, M. 2008. Application of acetates derivatives for gas chromatography-mass spectrometry: Novel approaches on carbohydrates, lipids and amino acids analysis. J. Chromatogr. A. 2008, 1208, 215–222. [Google Scholar] [CrossRef]
- Sassaki, G.L.; Gorin, P.A.J.; Souza, L.M.; Czelusniak, P.A.; Iacomini, M. Rapid synthesis of partially O-methylated alditol acetate standards for GC-MS: some relative activities of hydroxyl groups of methyl glycopyranosides on Purdie methylation. Carbohydr. Res. 2005, 340, 731–739. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Smiderle, F.R.; Sassaki, G.L.; Arkel, J.v.; Iacomini, M.; Wichers, H.J.; Griensven, L.J.L.D.V. High Molecular Weight Glucan of the Culinary Medicinal Mushroom Agaricus bisporus is an α-Glucan that Forms Complexes with Low Molecular Weight Galactan. Molecules 2010, 15, 5818-5830. https://doi.org/10.3390/molecules15085818
Smiderle FR, Sassaki GL, Arkel Jv, Iacomini M, Wichers HJ, Griensven LJLDV. High Molecular Weight Glucan of the Culinary Medicinal Mushroom Agaricus bisporus is an α-Glucan that Forms Complexes with Low Molecular Weight Galactan. Molecules. 2010; 15(8):5818-5830. https://doi.org/10.3390/molecules15085818
Chicago/Turabian StyleSmiderle, Fhernanda R., Guilherme L. Sassaki, Jeroen van Arkel, Marcello Iacomini, Harry J. Wichers, and Leo J.L.D. Van Griensven. 2010. "High Molecular Weight Glucan of the Culinary Medicinal Mushroom Agaricus bisporus is an α-Glucan that Forms Complexes with Low Molecular Weight Galactan" Molecules 15, no. 8: 5818-5830. https://doi.org/10.3390/molecules15085818
APA StyleSmiderle, F. R., Sassaki, G. L., Arkel, J. v., Iacomini, M., Wichers, H. J., & Griensven, L. J. L. D. V. (2010). High Molecular Weight Glucan of the Culinary Medicinal Mushroom Agaricus bisporus is an α-Glucan that Forms Complexes with Low Molecular Weight Galactan. Molecules, 15(8), 5818-5830. https://doi.org/10.3390/molecules15085818