Plant-Derived Antimalarial Agents: New Leads and Efficient Phytomedicines. Part II. Non-Alkaloidal Natural Products

Abstract
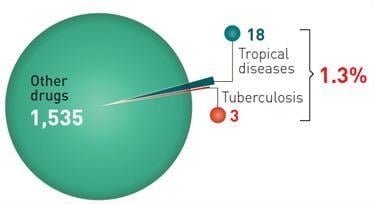
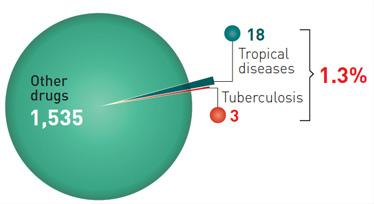
:
1. Introduction
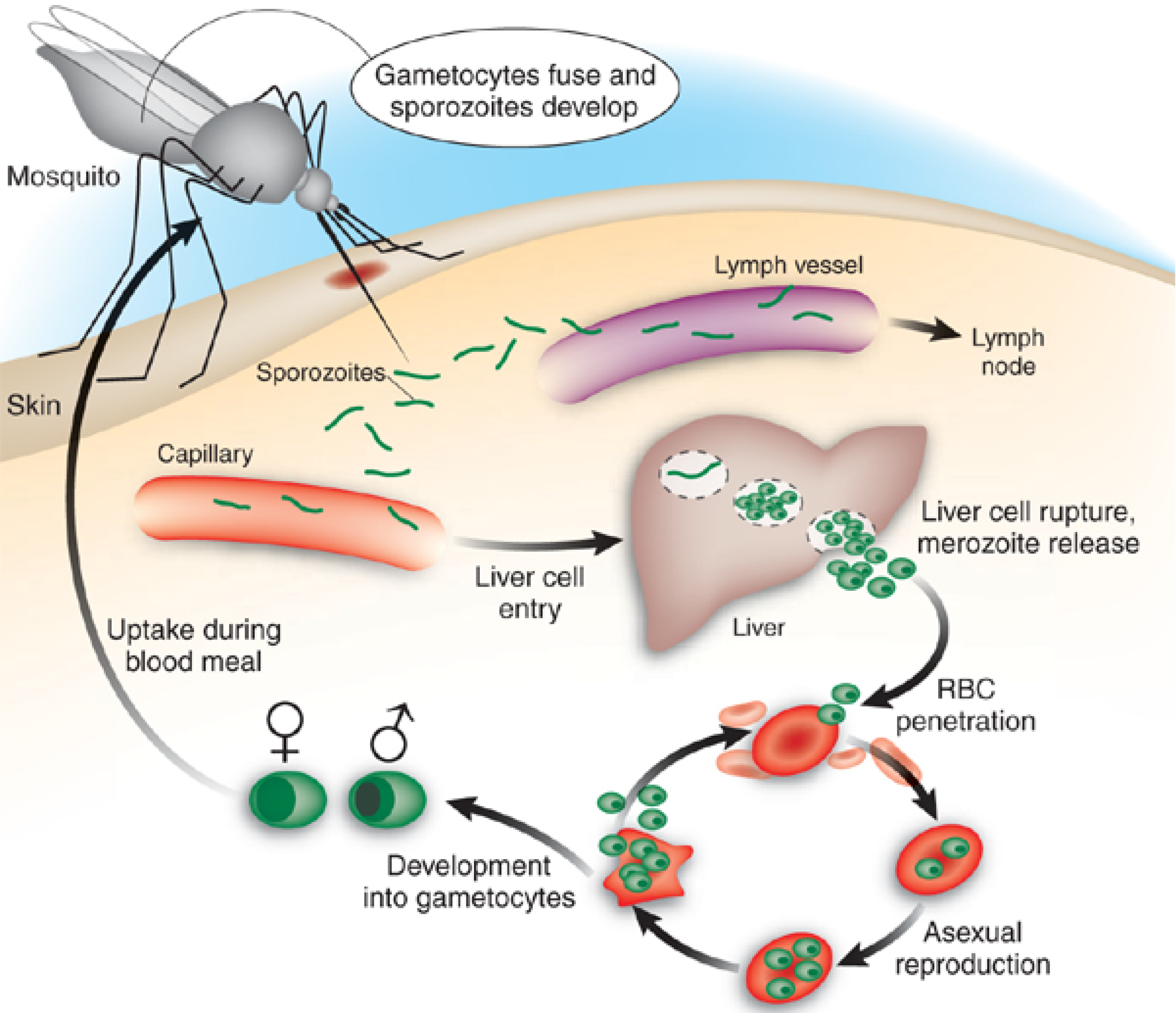

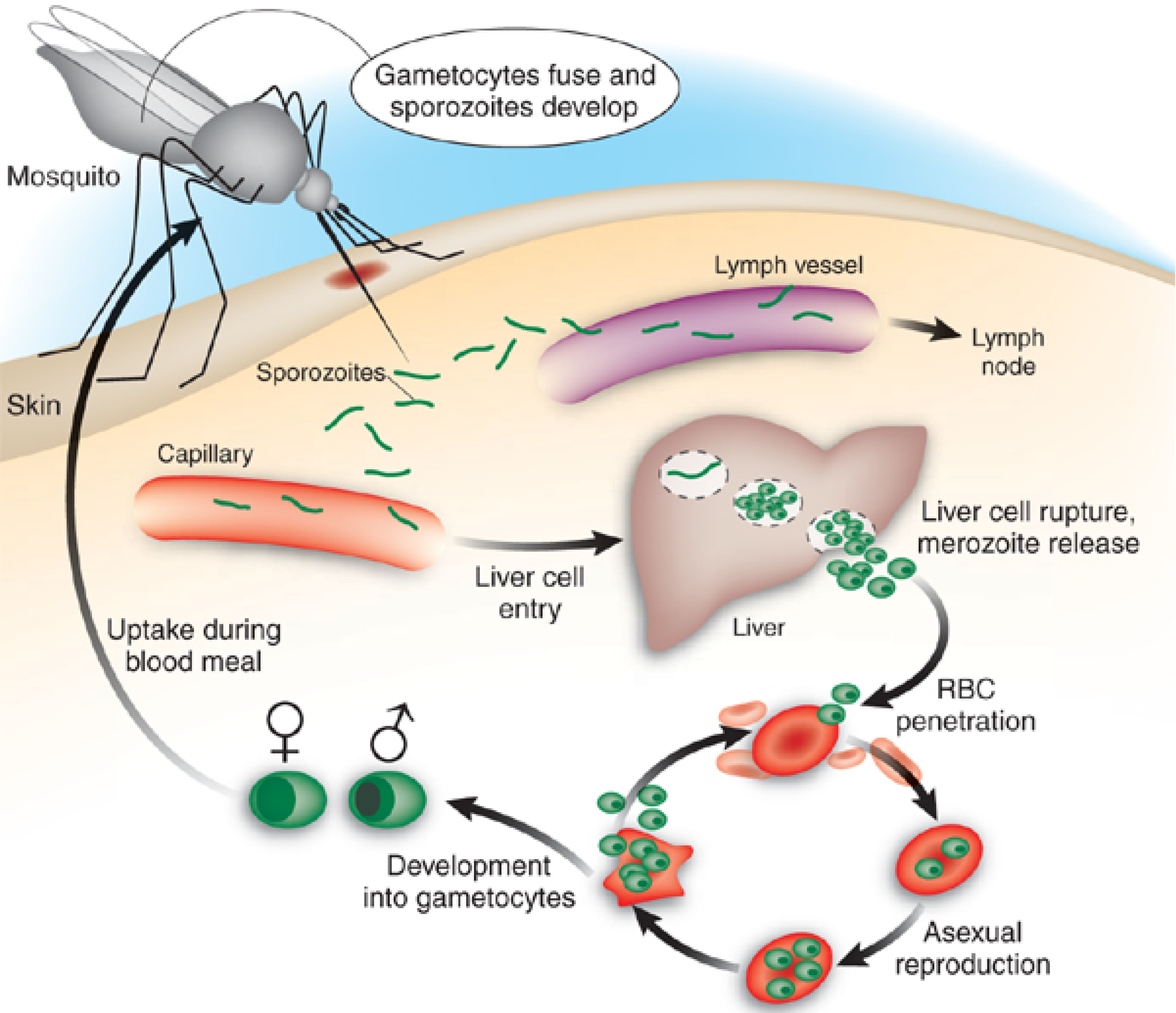
2. Recently-Published Crude Extracts with Antiplasmodial Activity

{kind=link}
{kind=link}
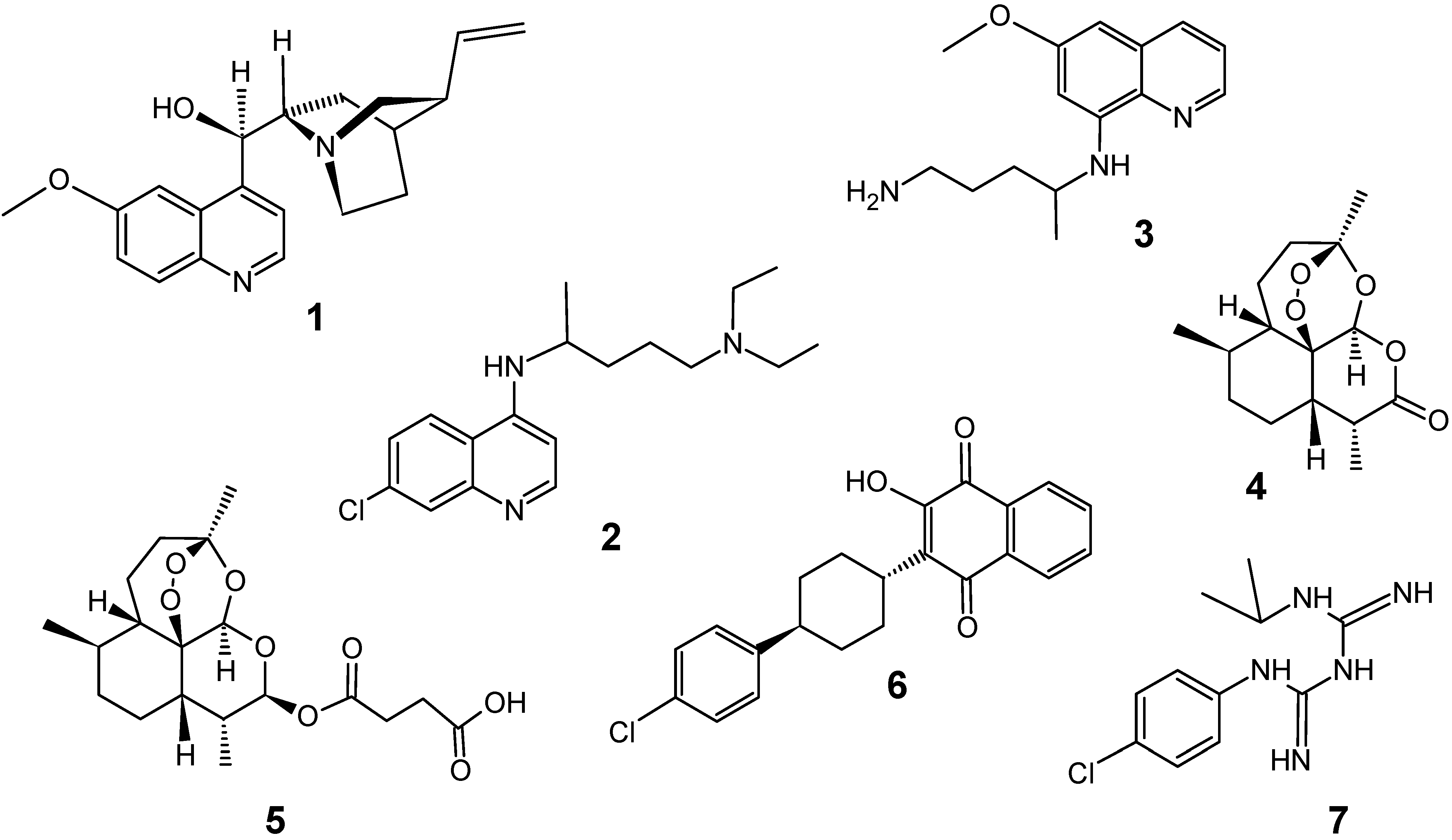{kind=link}
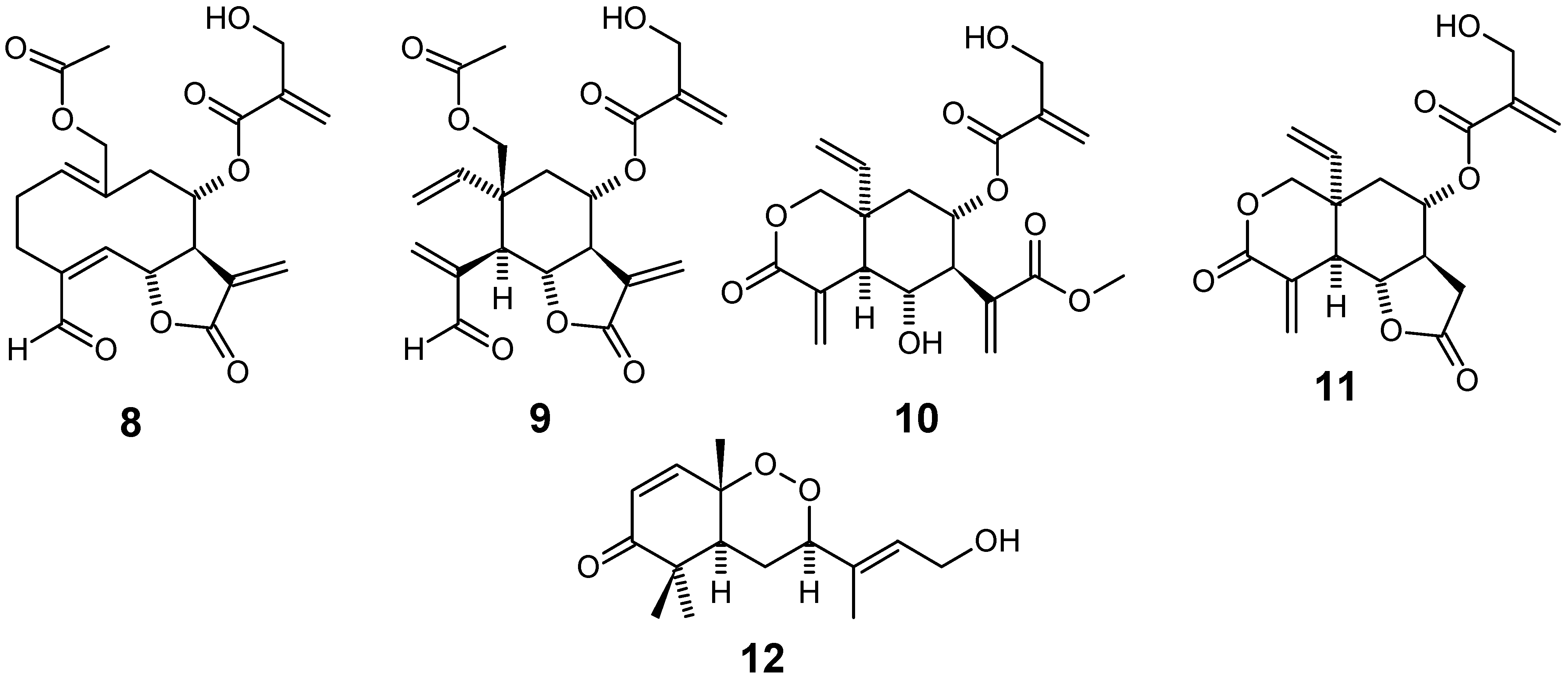{kind=link}
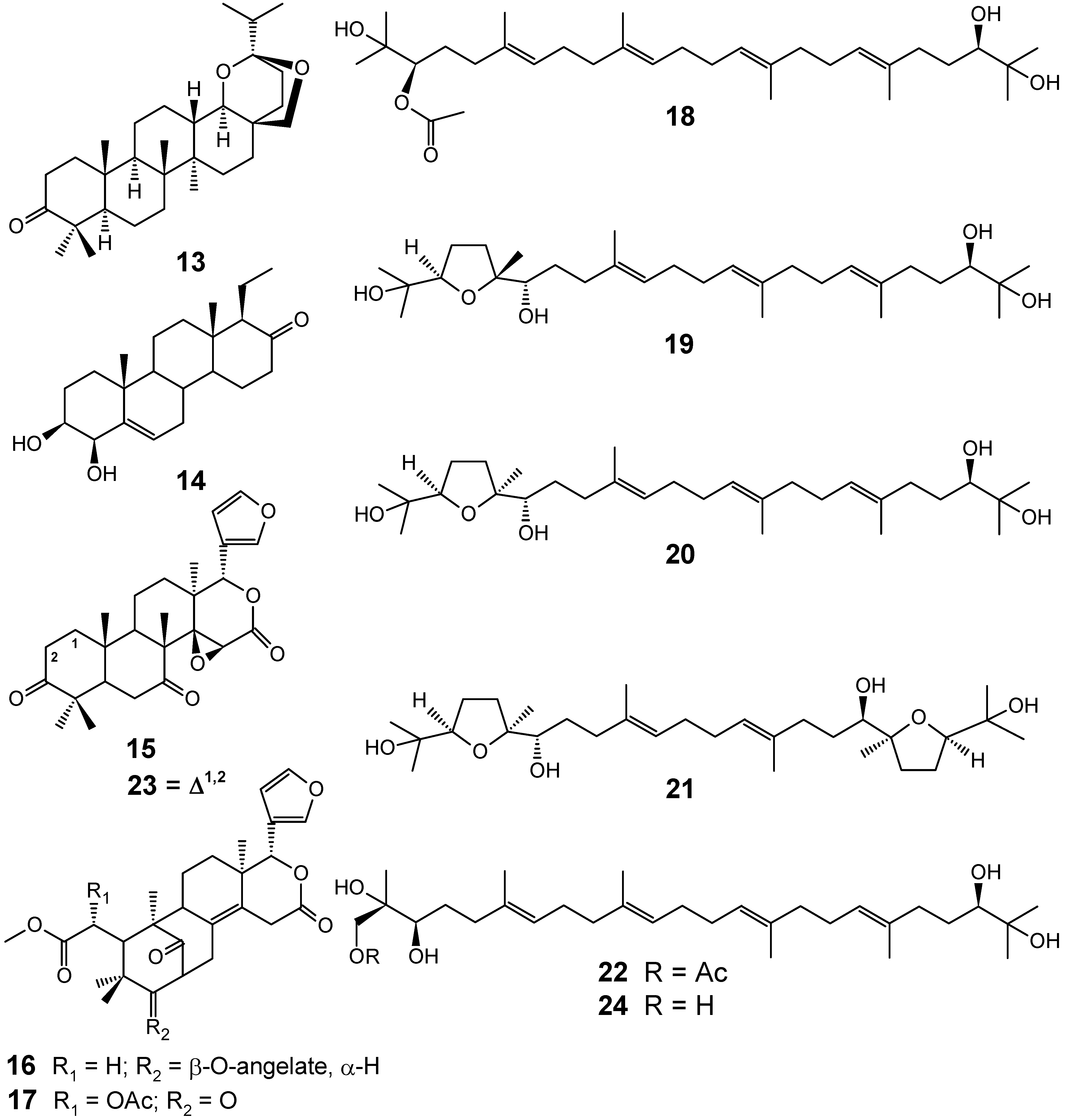{kind=link}
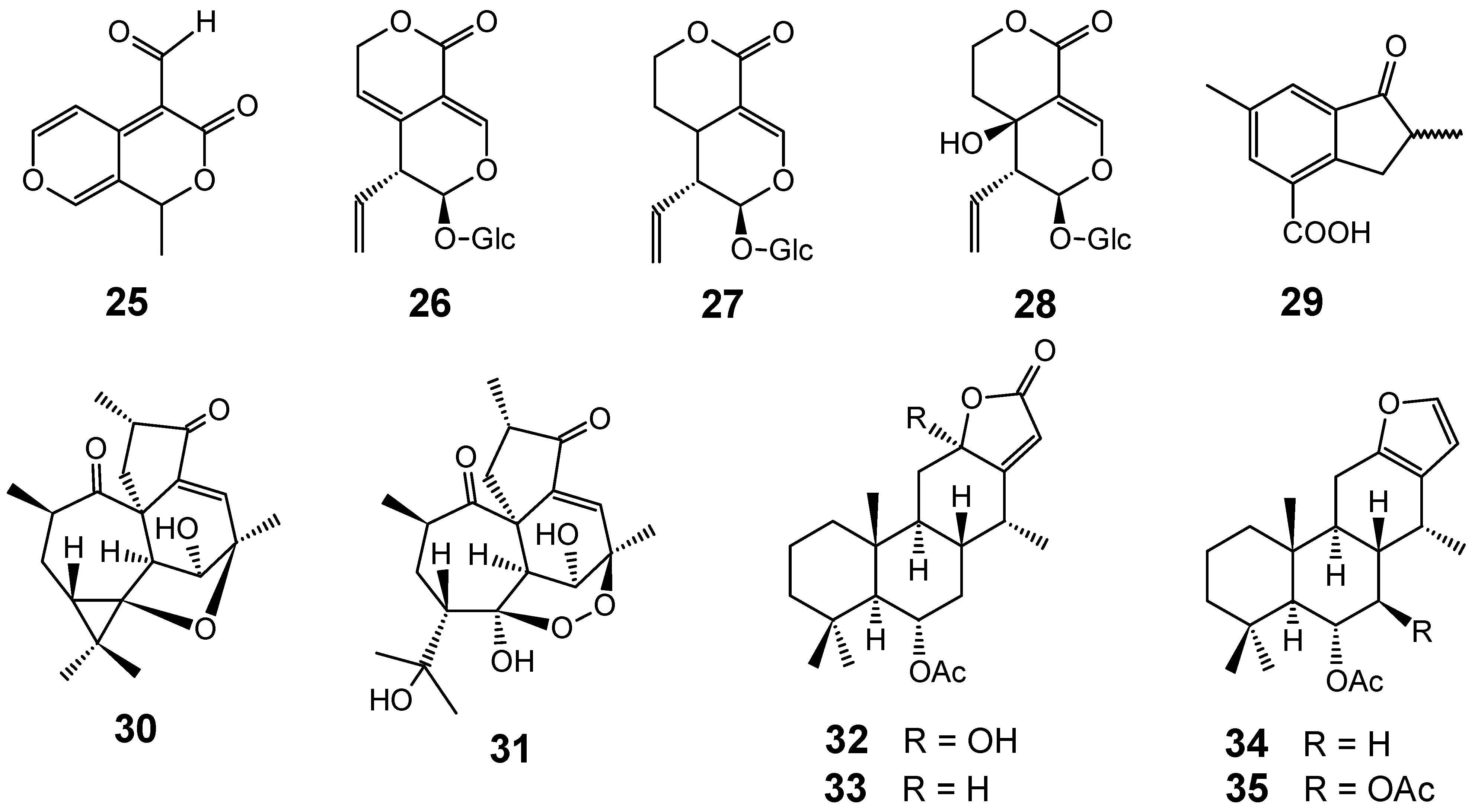{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
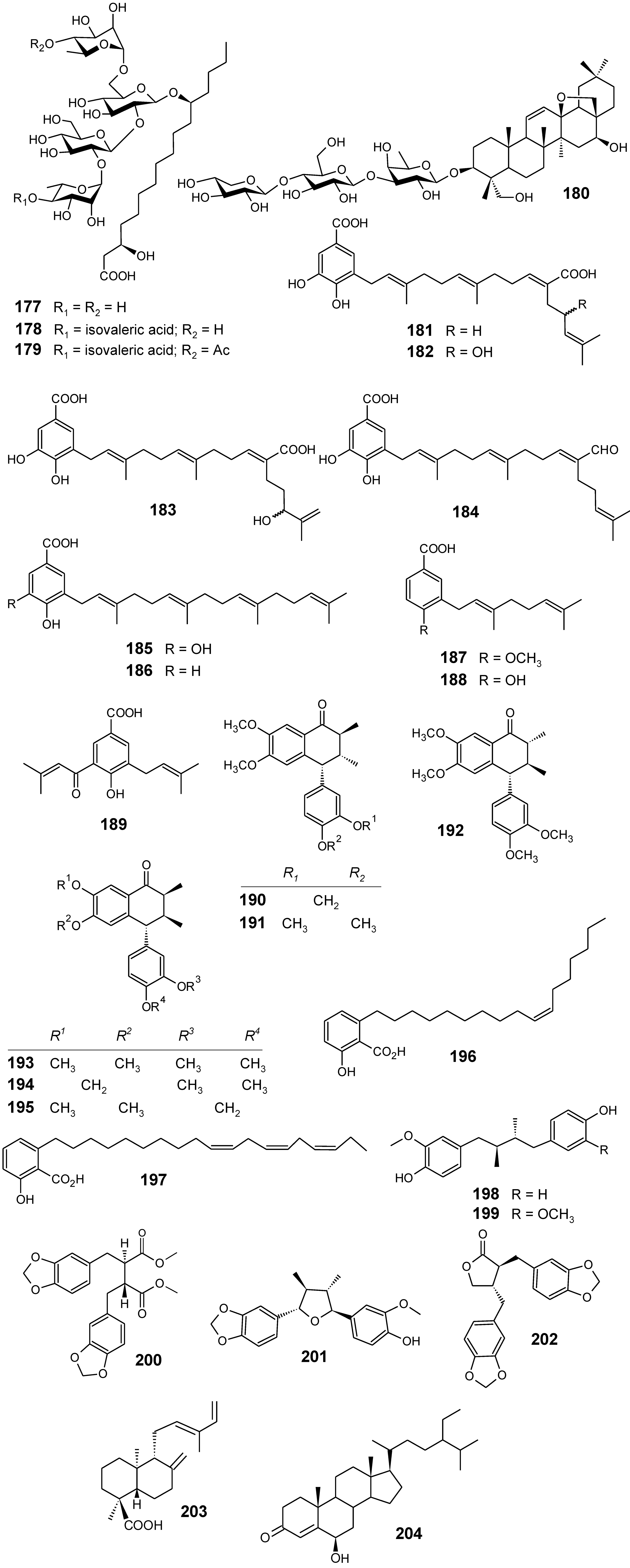{kind=link}
| Family | Species | Extract (part) | Antiplasmodial Activity (IC50, μg/mL) | Strain | Ref. | |
|---|---|---|---|---|---|---|
| Amaranthaceae | Amaranthus spinosus | Water (stem bark) | 789.36 ± 7.19 ¥ | (N.S.) ≠ S | [44] | |
| Annonaceae | Uvariopsis congolana | MeOH* (stem) | 4.47 ± 0.45 | W2 R | [45] | |
| MeOH* (leaves) | 4.57 ± 0.76 | W2 R | ||||
| Polyalthia oliveri | MeOH* (stem bark) | 4.30 ± 0.31 | W2 R | |||
| Enantia chlorantha | MeOH* (stem) | 4.79 ± 1.09 | W2 R | |||
| MeOH* (stem bark) | 2.06 ± 0.01 | W2 R | ||||
| Aphloiaceae | Aphloia theiforms | MeOH (bark) | 13.3 ± 0.8 | 3D7 S | [46] | |
| 11.0 ± 3.1 | W2 R | |||||
| CH2Cl2 (bark) | 16.1 ± 2.7 | 3D7 S | ||||
| 18.2 ± 2.7 | W2 R | |||||
| CH2Cl2 (leaves) | 21.0 ± 1.6 | 3D7S | ||||
| 22.7 ± 2.9 | W2 R | |||||
| Apiaceae | Ferula oopoda | MeOH (roots) | 26.6 | K1 R | [47] | |
| 24.9 | 3D7 S | |||||
| Astrodaucus orientalis | MeOH (aerial parts) | 46.1 | K1 R | |||
| 42.6 | 3D7 S | |||||
| Apocynaceae | Picralima nitida | EtOH (seeds) [115 mg/Kg] | 73.0%** | (N.S.) ≠ S | [48] | |
| Asclepiadaceae | Caralluma tuberculata | Pet. Ether (aerial parts) | 7.94 | K1 R | [49] | |
| Asteraceae | Vernonia amygdalina | EtOH (leaves) | 9.7 ± 2.6 | (N.S.) S | [50] | |
| Pet. Ether (leaves) | 2.5 ± 0.7 | (N.S.) S | ||||
| Isoamyl alcohol (leaves) | 2.7 ± 0.6 | (N.S.) S | ||||
| Psiadia arguta | CH2Cl2 (leaves) | 10.1 ± 2.2 | 3D7 S | [46] | ||
| 8.4 ± 1.1 | W2 R | |||||
| MeOH (leaves) | 22.4 ± 2.2 | 3D7 S | ||||
| 26.1 ± 6.5 | W2 R | |||||
| Centaurea bruguieriana | MeOH (aerial parts) | 36.9 | 3D7 S | [47] | ||
| Centaurea golestanica | MeOH (aerial parts) | 35.6 | K1 R | |||
| 31.6 | 3D7 S | |||||
| Boraginaceae | Heliotropium zeylanicum | MeOH (aerial parts) | 8.41 | K1 R | [49] | |
| Buxaceae | Buxus hyrcana | MeOH (aerial parts) | 4.7 | K1 R | [47] | |
| 7.7 | 3D7 S | |||||
| Caesalpiniaceae | Cassia occidentalis | EtOH (leaves) | 2.8 ± 0.5 | (N.S.) S | [50] | |
| Pet. Ether (leaves) | 1.5 ± 0.7 | (N.S.) S | ||||
| Isoamyl alcohol (leaves) | 18.6 ± 3.6 | (N.S.) S | ||||
| CHCl3 [pH 2-3] (leaves) | 2.9 ± 0.3 | (N.S.) S | ||||
| Capparaceae | Boscia angustifolia | CH2Cl2 (leaves) | 107.9 | 3D7 S | [51] | |
| MeOH (leaves) | 37.6 | 3D7 S | ||||
| Caryophyllaceae | Minuartia lineata | MeOH (aerial parts) | 44.0 | 3D7 S | [47] | |
| Clusiaceae | Croton zambesicus | EtOH (roots) [81 mg/Kg] | 86.18% ** | ANKA ≠ | [52] | |
| n-hexane (root) [57 mg/Kg] | 57.88% ** | ANKA ≠ | ||||
| CHCl 3 (root) [57 mg/Kg] | 75.39% ** | ANKA ≠ | ||||
| AcOEt (root) [57 mg/Kg] | 76.89% ** | ANKA ≠ | ||||
| MeOH (root) [57 mg/Kg] | 77.27% ** | ANKA ≠ | ||||
| Garcinia kola | EtOH (stem bark) | 2.9 ± 0.7 | (N.S.) S | [50] | ||
| Pet. Ether (stem bark) | 1.6 ± 0.2 | (N.S.) S | ||||
| Isoamyl alc. (stem bark) | 41.7 ± 3.2 | (N.S.) S | ||||
| CHCl3 [pH 2-3] (stem bark) | 27.1 ± 2.7 | (N.S.) S | ||||
| Symphonia globulifera | MeOH (leaves) | 4.1±0.5 | K1 R | [53] | ||
| Combretaceae | Terminalia bentzoe L. | MeOH (fresh leaves) | 12.8 ± 2.9 | 3D7 S | [46] | |
| 12.8 ± 3.5 | W2 R | |||||
| CH2Cl2 (leaves) | 42.7 ± 3.2 | 3D7 S | ||||
| 21.0 ± 2.1 | W2 R | |||||
| Cucurbitaceae | Momordica foetida | Water (shoots) | 40.7 ± 11.20 | D10 S | [54] | |
| 50.8 ± 3.00 | K1 R | |||||
| AcOEt (shoots) | 30.0 ± 1.70 | D10 S | ||||
| 29.30 ± 1.47 | K1 R | |||||
| MeOH (shoots) | 75.4 ± 17.50 | D10 S | ||||
| 68.80 ± 5.40 | K1 R | |||||
| Dilleniaceae | Tefracera pogge | EtOH (leaves) | 36.9± 4.2 | (N.S.) S | [50] | |
| Pet. Ether (leaves) | 1.7 ± 0.4 | (N.S.) S | ||||
| Isoamyl alcohol (leaves) | 21.8 ± 5.2 | (N.S.) S | ||||
| Euphorbiaceae | Euphorbia hirta | EtOH (whole plant) | 2.4 ± 0.2 | (N.S.) S | ||
| Pet. Ether (whole plant) | 1.2 ± 0.3 | (N.S.) S | ||||
| Isoamyl alc. (whole plant) | 2.6 ± 1.2 | (N.S.) S | ||||
| Neoboutonia. glabracens | MeOH (leaves) | 5.50 ± 0.20 | W2 R | [45] | ||
| Croton zambesicus | MeOH (stem bark) | 5.69 ± 0.06 | W2 R | |||
| Phyllantus niruri | EtOH (whole plant) | 2.5± 0.1 | (N.S.) S | [50] | ||
| Pet. Ether (whole plant) | 1.3 ± 0.3 | (N.S.) S | ||||
| Isoamyl alc. (whole plant) | 2.3 ± 0.5 | (N.S.) S | ||||
| Fabaceae | Glycyrrhiza glabra | MeOH (aerial parts) | 17.5 | K1 R | [47] | |
| Erythrina fusca | EtOAc (stem bark) | 7.5 | K1 R | [55] | ||
| Stylosanthes erecta | CH2Cl2 (aerial parts) | 21.9 | 3D7 S | [51] | ||
| MeOH (aerial parts) | 23.3 | 3D7 S | ||||
| Tetrapleura tetraptera | EtOH (fruits) [900 mg/Kg] | 76.37%** | (N.S.) ≠ S | [56] | ||
| Geraniaceae | Erodium oxyrrhnchum | MeOH (aerial parts) | 40.3 | K1 R | [47] | |
| 13.0 | 3D7 S | |||||
| Hypericaceae | Harungana madagascariensis | EtOH (stem bark) | 0.052-0.517 | (N.S.) | [57] | |
| MeOH (seeds) | 3.6 ± 0.3 | K1 R | [53] | |||
| Lamiaceae | Otostegia persica | MeOH (fruits + leaves) | 31.1 | K1 R | [47] | |
| Otostegia michauxii | MeOH (aerial parts) | 44.6 | K1 R | |||
| Perovskia abrotanoides | MeOH (aerial parts) | 37.3 | K1 R | |||
| Loganiaceae | Nuxia verticillata | CH2Cl2 (leaves) | 10.9 ± 1.8 | 3D7 S | [46] | |
| 8.8 ± 1.2 | W2 R | |||||
| MeOH (leaves) | 32.7 ± 7.4 | 3D7 S | ||||
| CH2Cl2 (bark) | 27.4 ± 6.6 | 3D7 S | ||||
| MeOH (bark) | 36.9 ± 5.7 | 3D7 S | ||||
| Buddleja salviifolia | CH2Cl2 (bark) | 49.9 ± 9.6 | 3D7 S | |||
| CH2Cl2 (leaves) | 29.7 ± 12.6 | 3D7 S | ||||
| 18.6 ± 5.8 | W2 R | |||||
| Strychnos angolensis | EtOAc (roots) | 17.0 ± 7.6 | FCA20 S | [58] | ||
| Strychnos cocculoides | EtOAc (leaves) | 20.0 ± 11.9 | FCA20 S | |||
| Strychnos gossweileri | EtOAc (roots) | 12.4 ± 4.1 | FCA20 S | |||
| Strychnos henningsii | EtOAc (leaves) | 15.9 ± 3.0 | FCA20 S | |||
| Strychnos johnsonii | EtOAc (stem) | 16.4 ± 1.8 | FCA20 S | |||
| Strychnos mellodora | EtOAc (leaves) | 13.4 | FCA20 S | |||
| MeOH (leaves) | 29.5 | FCA20 S | ||||
| EtOAc (stem) | 14.5 ± 1.5 | FCA20 S | ||||
| EtOAc (roots) | 11.2 ± 3.6 | FCA20 S | ||||
| MeOH (roots) | 25.4 ± 11.0 | FCA20 S | ||||
| Strychnos scheffleri | EtOAc (leaves) | 21.2 | FCA20 S | |||
| Strychnos variabilis | EtOAc (roots) | 2.5 ± 0.2 | FCA20 S | |||
| MeOH (roots) | 2.3 ± 0.5 | FCA20 S | ||||
| Meliaceae | Trichilia emetica | CH2Cl2 (leaves) | 11.9 | 3D7 S | [51] | |
| MeOH (leaves) | 47.6 | 3D7 S | ||||
| Mimosaceae | Cylicodiscus gabunensis | EtOH (stem bark) [60 mg/Kg] | 83.3% ¥ | (N.S.) ≠ S | [59] | |
| Albizia zygia | MeOH (stem bark) | 1.0 ± 0.1 | K1 R | [53] | ||
| Moraceae | Artocarpus communis | MeOH (leaves) | 4.00 ± 0.37 | W2 R | [45] | |
| Nyctagynaceae | Boerhaavia erecta | Water (stem bark) | 564.95 ± 6.23 ¥ | (N.S.) ≠ S | [44] | |
| Polygalaceae | Securidaca longipedunculata | CH2Cl2 (leaves) | 6.9 | 3D7 S | [51] | |
| Rubiaceae | Vangueria infausta | CHCl3 [fr.] (root bark) | 3.8 ± 1.5 | D6 S | [60] | |
| 4.5 ± 2.3 | W2 R | |||||
| Morinda morindoides | EtOH (leaves) | 94.2 ± 3.4 | (N.S.) S | [50] | ||
| Pet. Ether (leaves) | 1.8 ± 0.2 | (N.S.) S | ||||
| Isoamyl alcohol (leaves) | 15.3 ± 3.6 | (N.S.) S | ||||
| CHCl3 [pH 2-3] (leaves) | 8.8 ± 2.5 | (N.S.) S | ||||
| Sapindaceae | Cardiospermum halicacabum | AcOEt (shoots) | 28.60 ± 4.20 | D10 S | [54] | |
| 32.60 ± 2.60 | K1 R | |||||
| MeOH (shoots) | 62.60 ± 9.40 | D10 S | ||||
| 79.00 ± 5.20 | K1 R | |||||
| Solanaceae | Licium shawii | MeOH (aerial parts) | 7.75 | K1 R | [49] | |
| Tamaricaceae | Tamarix aralensis | MeOH (aerial parts) | 43.8 | 3D7 S | [47] | |
| Verbenaceae | Lantana camara | CH2Cl2 (leaves) | 8.7 ± 1.0 | 3D7 S | [46] | |
| 5.7 ± 1.6 | W2 R | |||||
| Vitaceae | Cissus quadrangulari | CH2Cl2 (whole plant) | 23.9 | 3D7 S | [51] | |
| MeOH (whole plant) | 52.8 | 3D7 S | ||||
| Zingiberaceae | Siphonochilus aethiopicus | EtOAc (rhizomes) | 2.90 ± 0.28 | D10 S | [61] | |
| 1.4 | K1 R | |||||
3. Antiplasmodial Non-Alkaloidal Natural Products
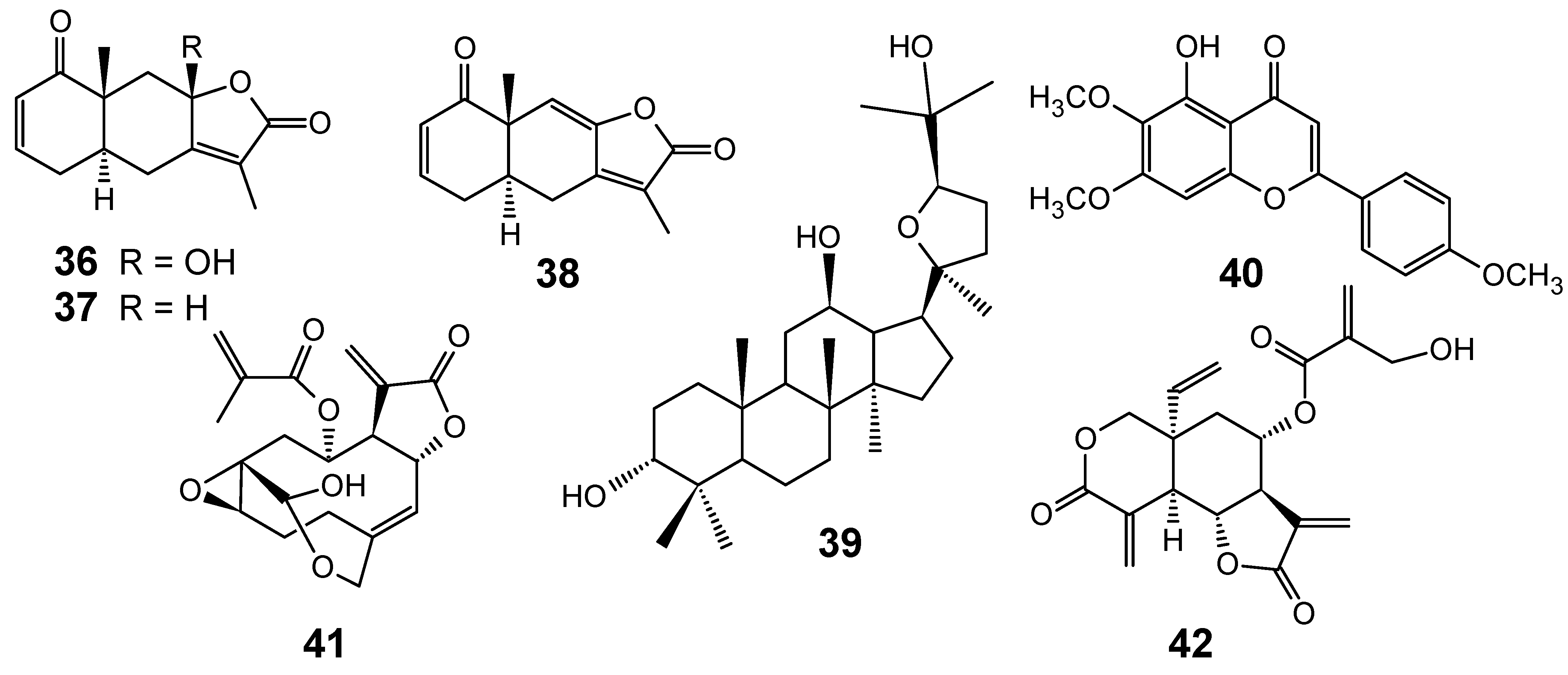
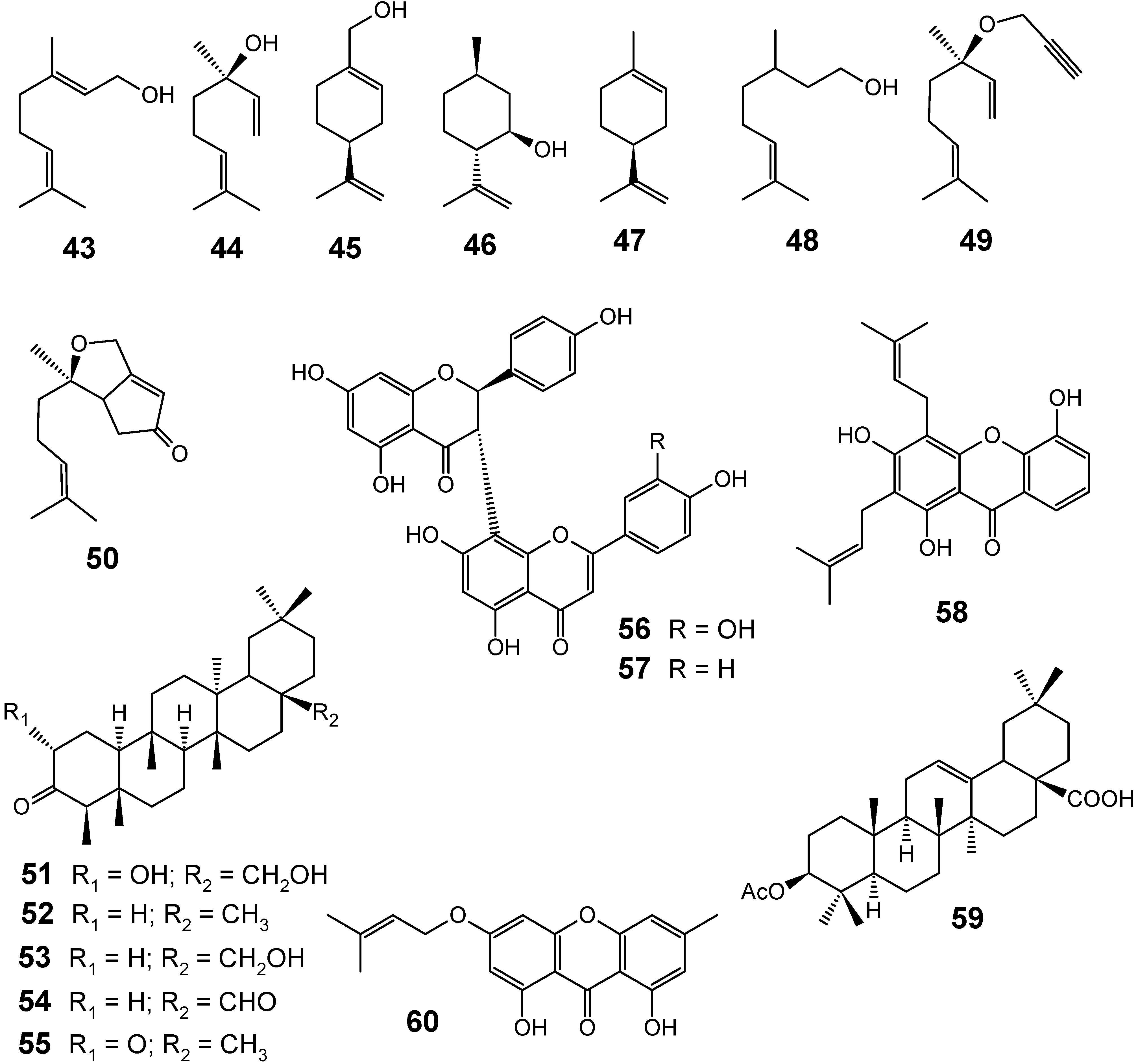
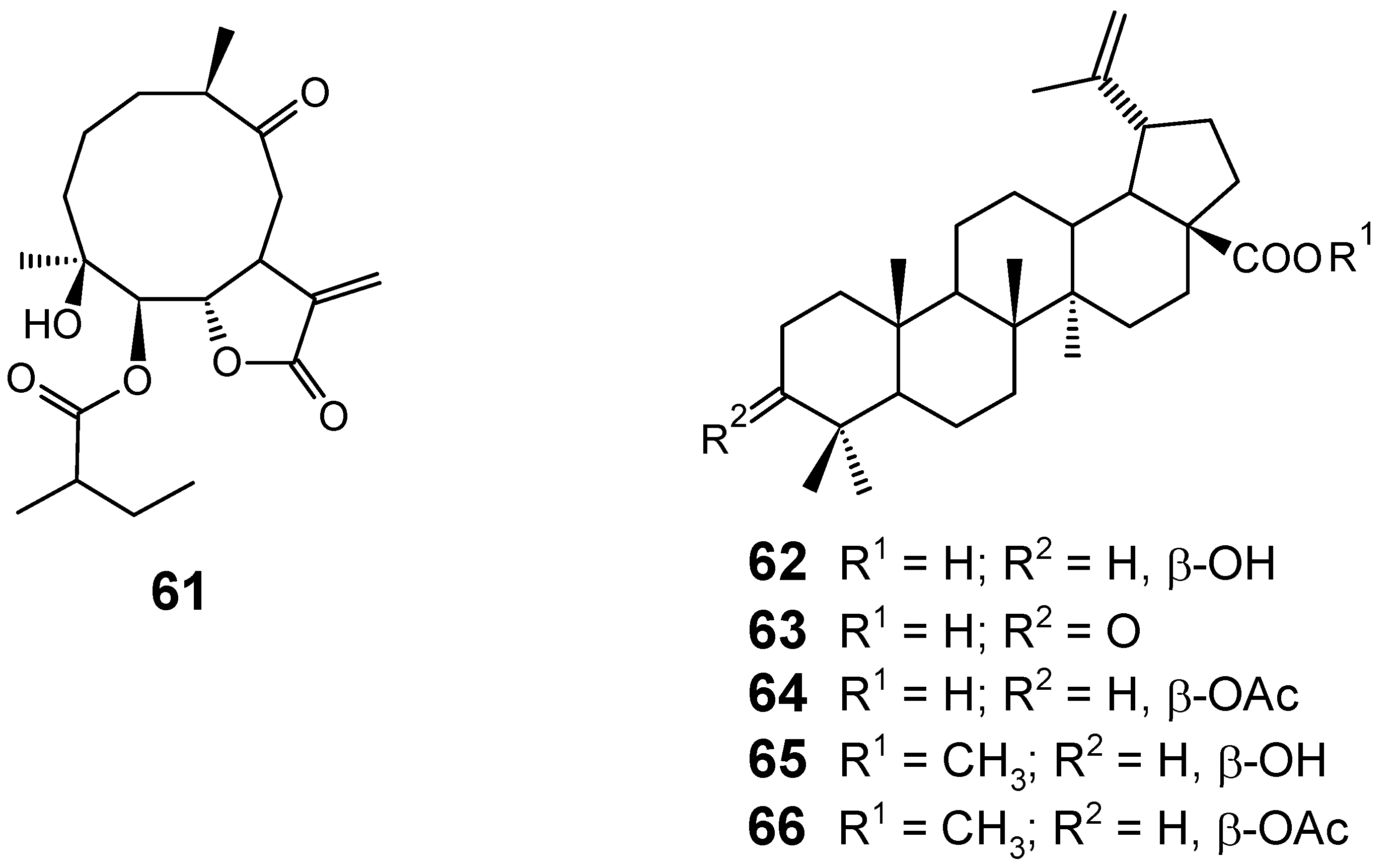
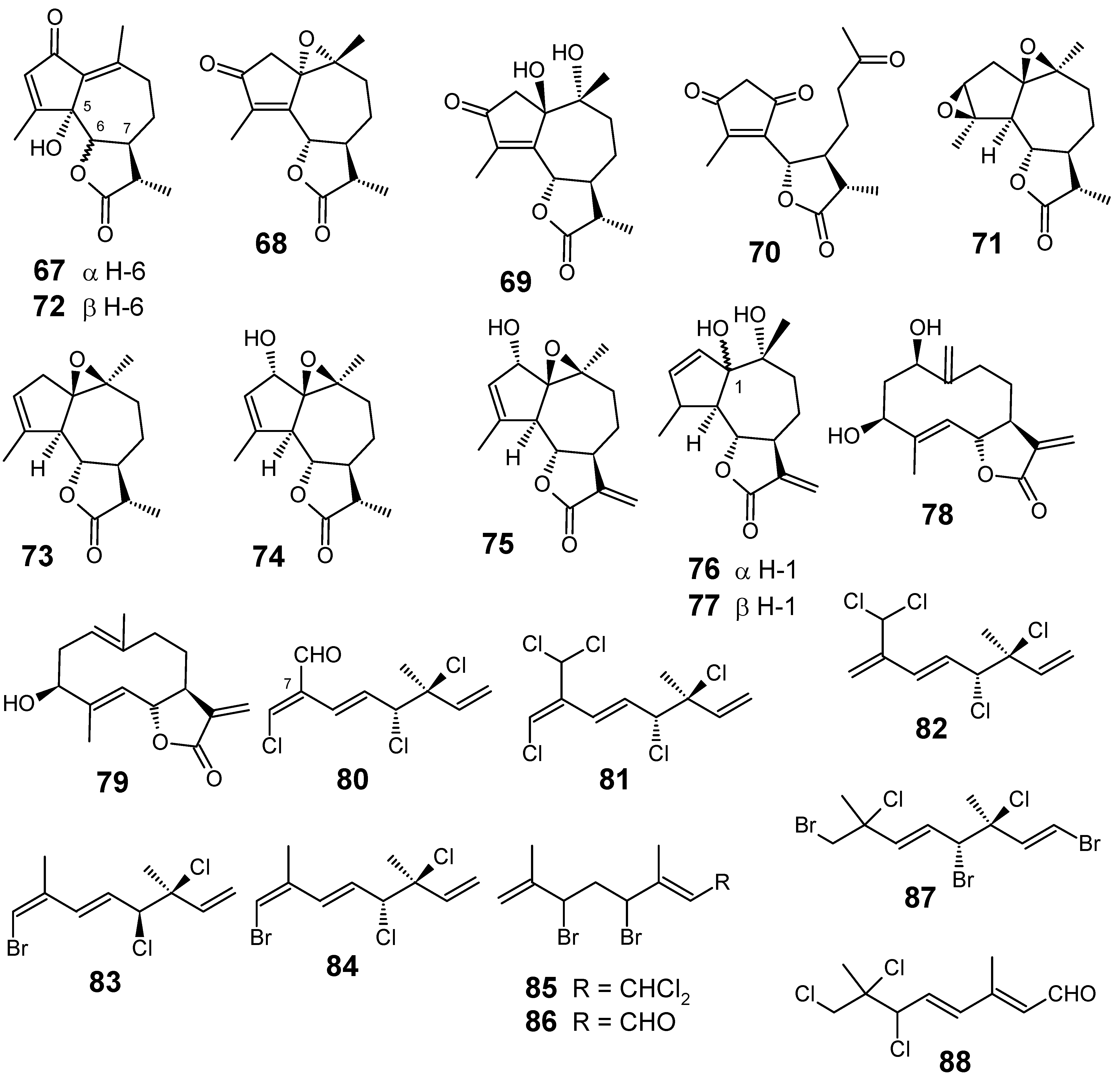
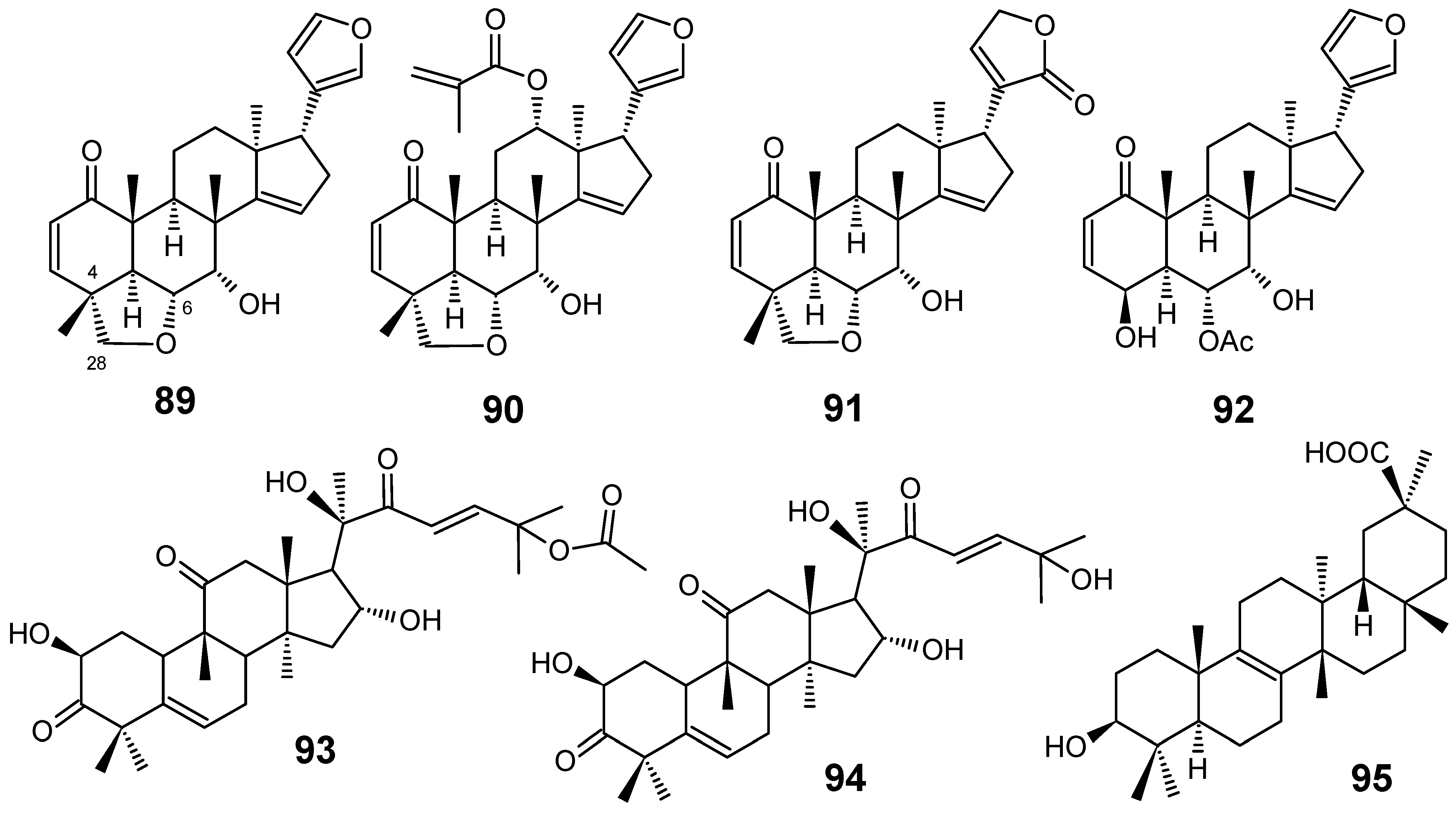
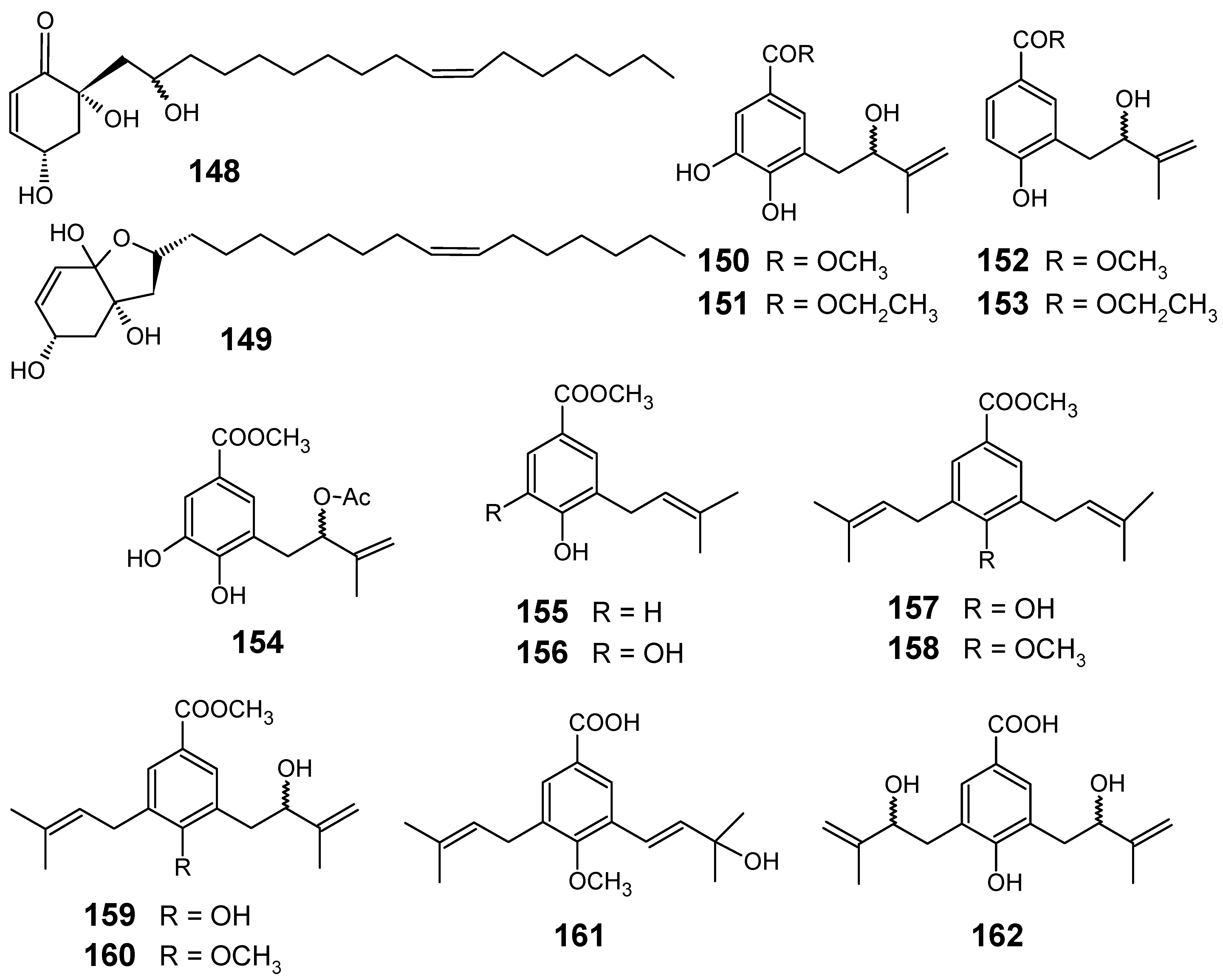
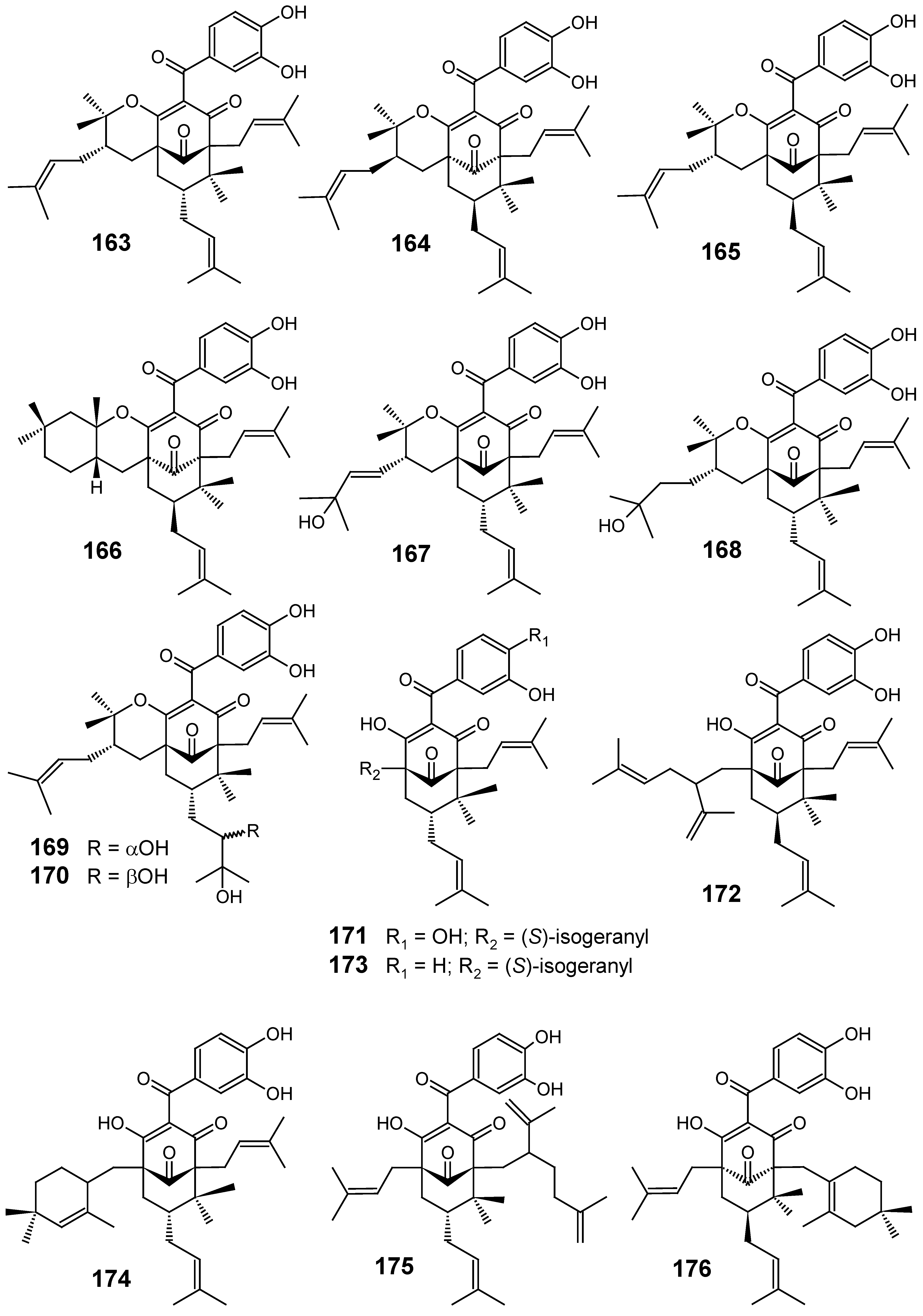
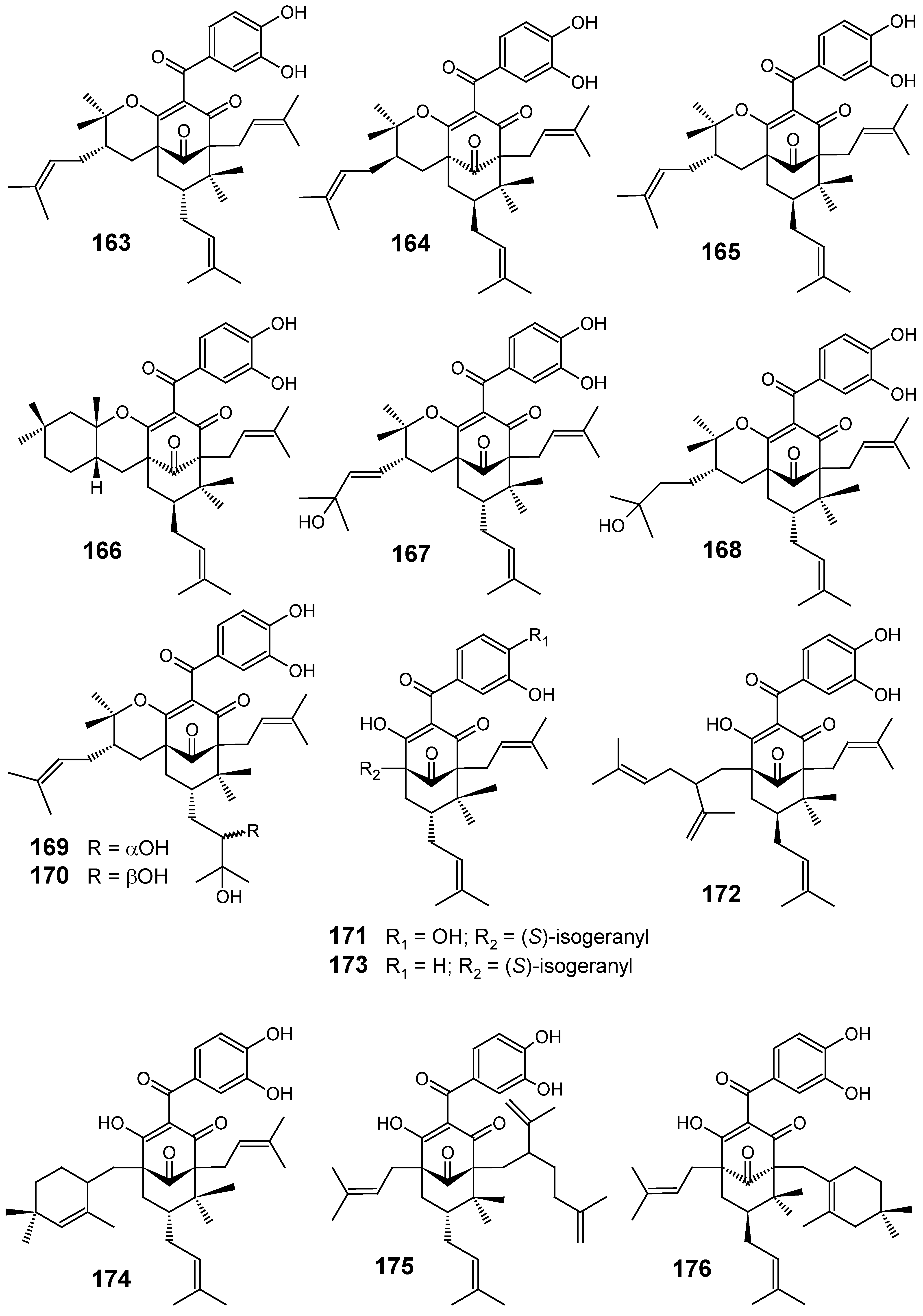
3.1. Terpenes and related compounds

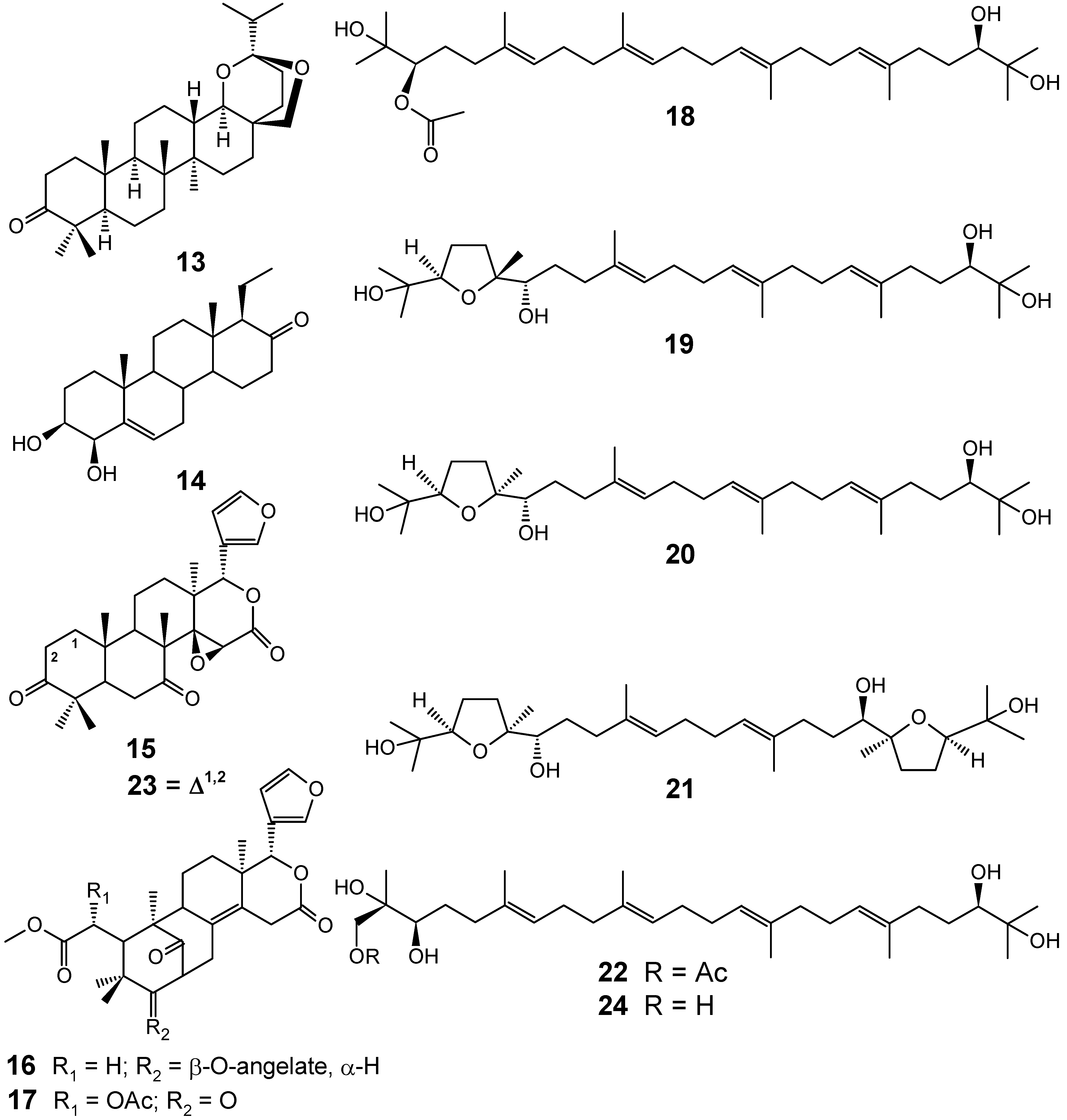
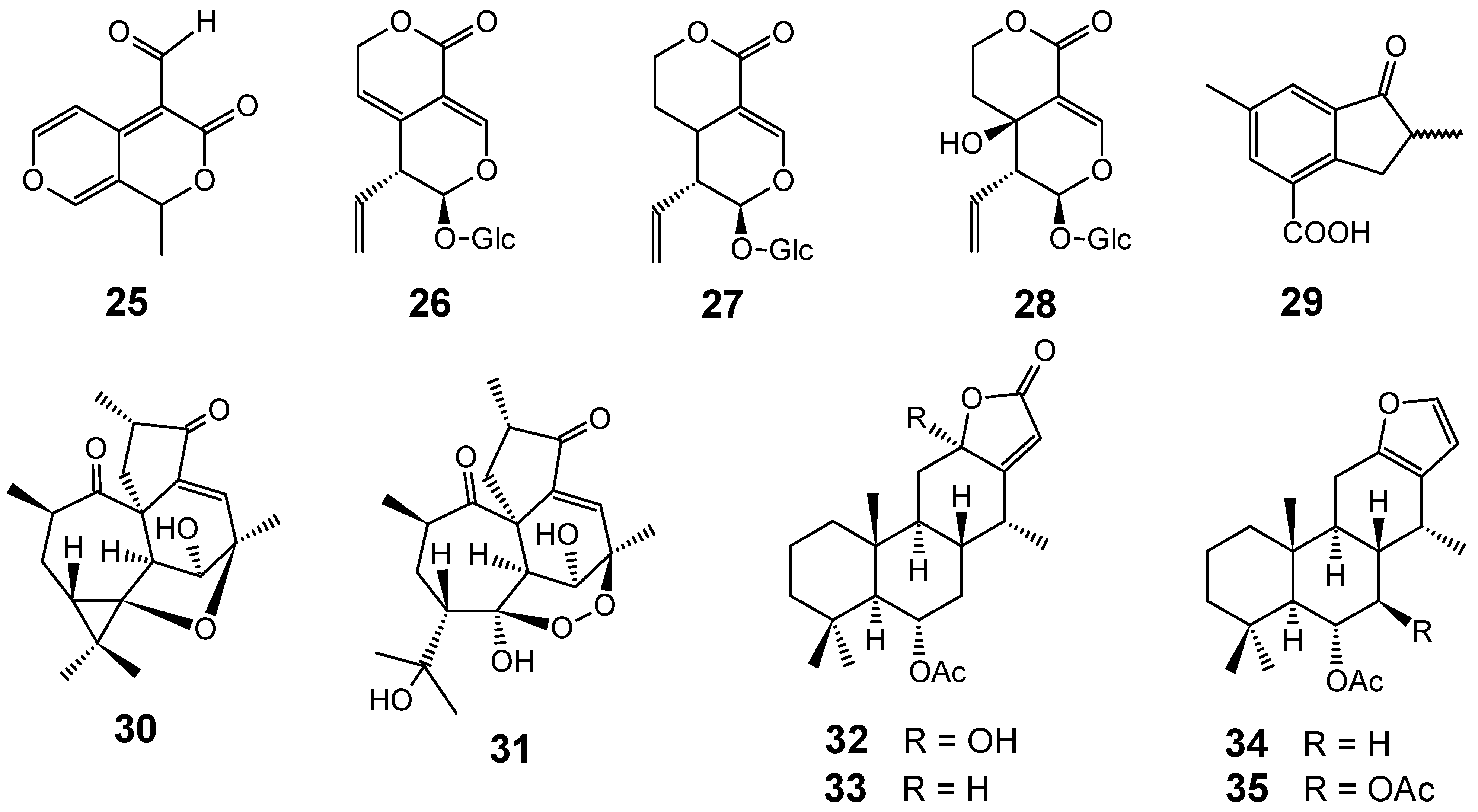


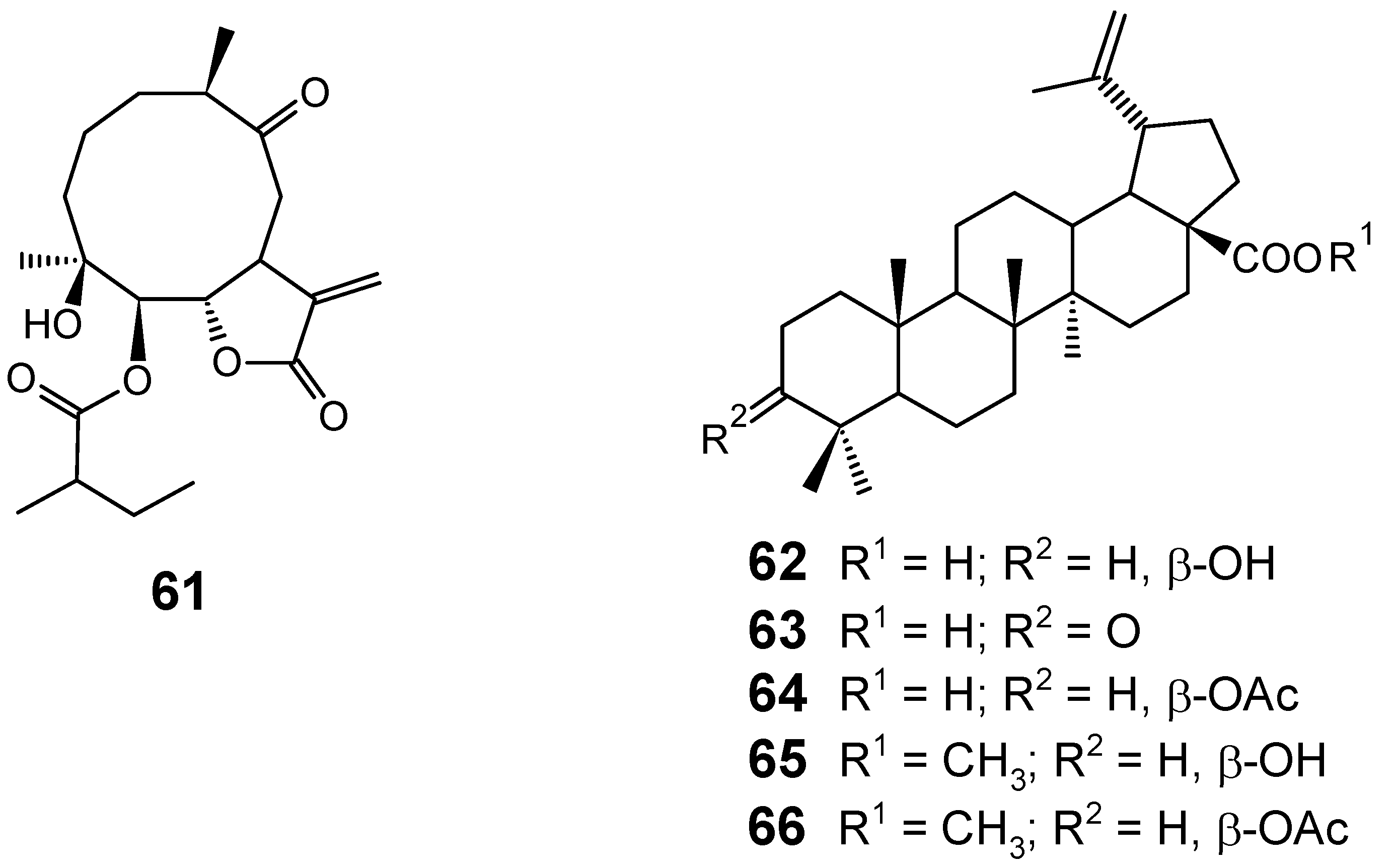
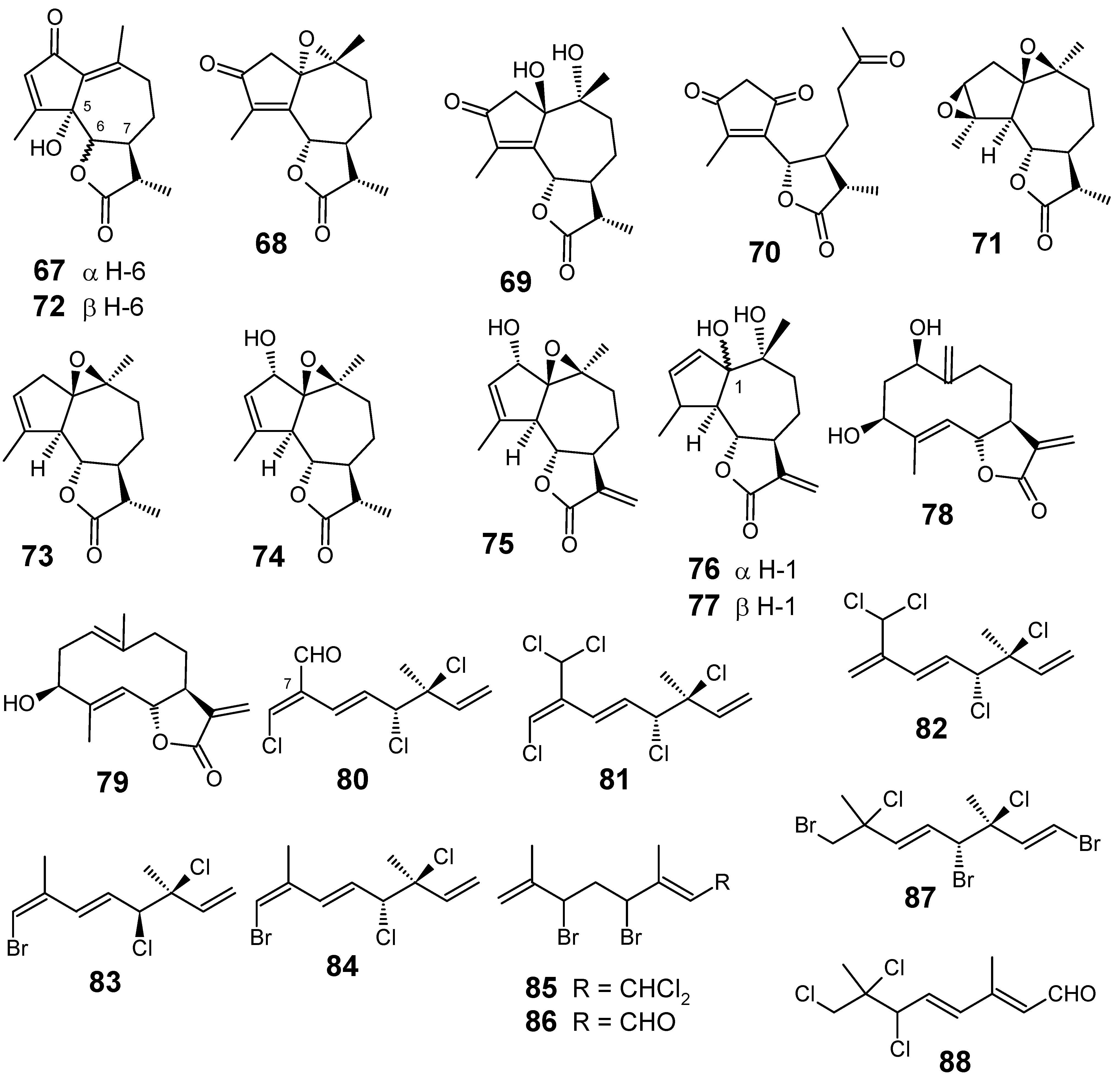
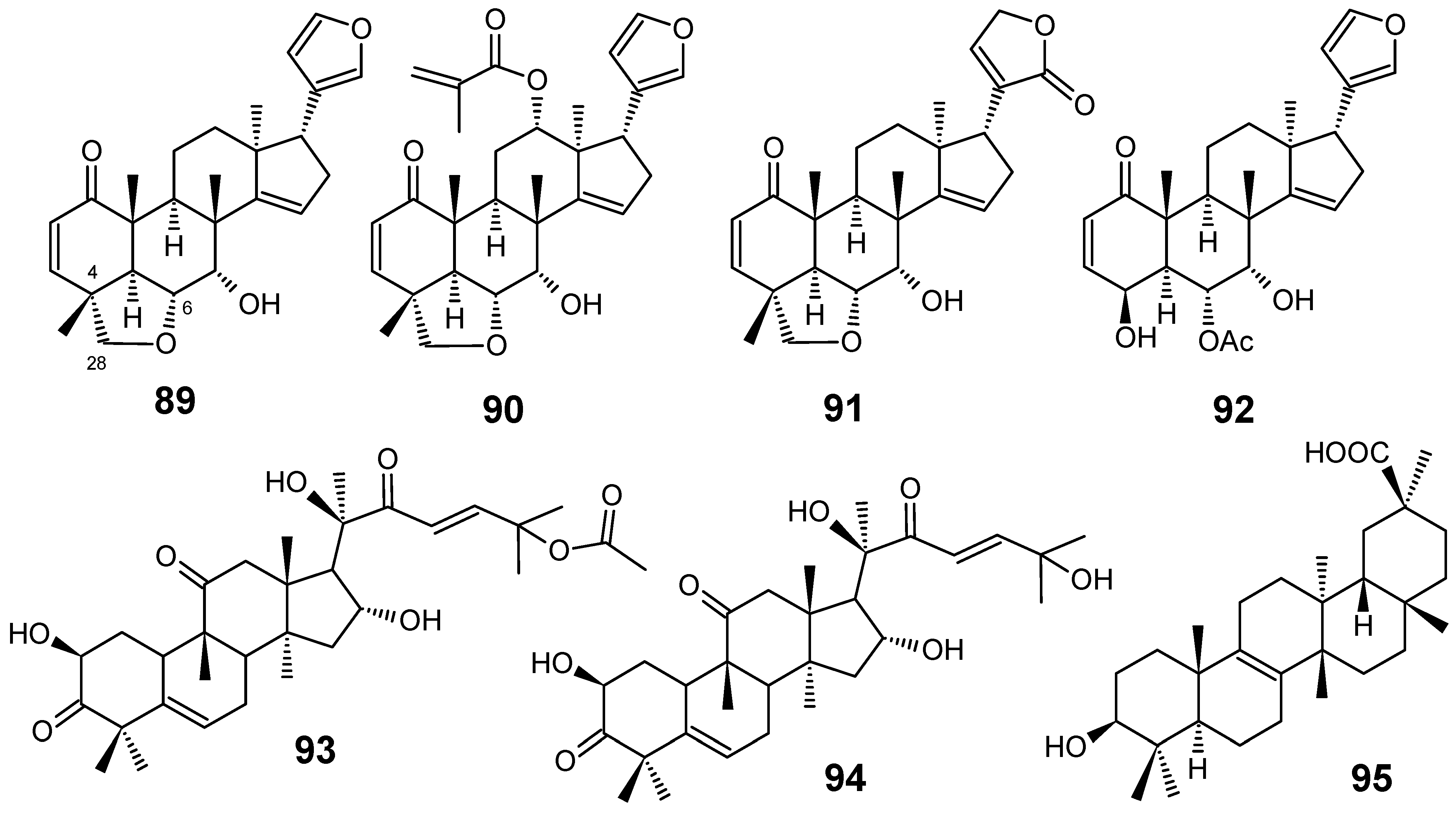
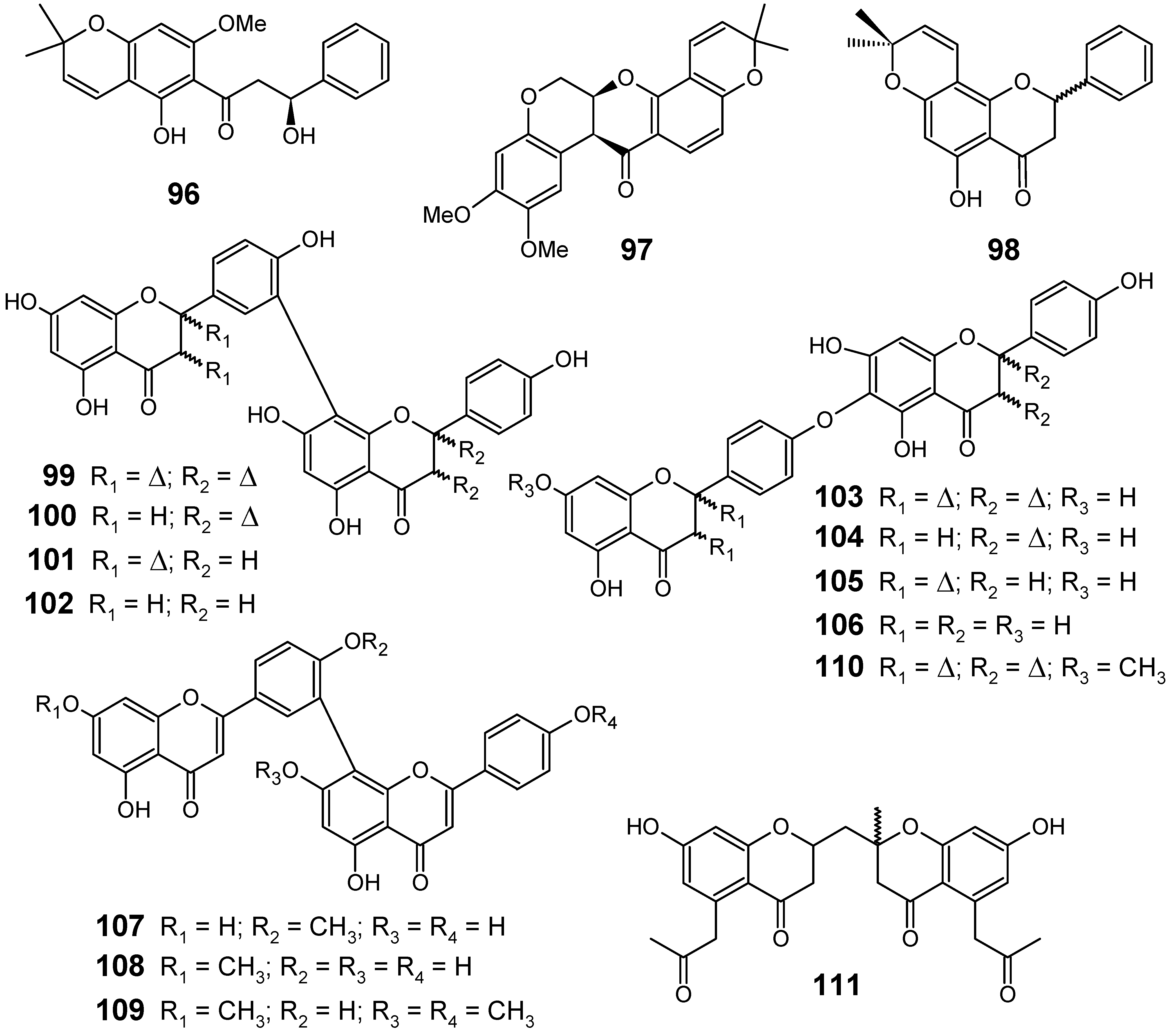
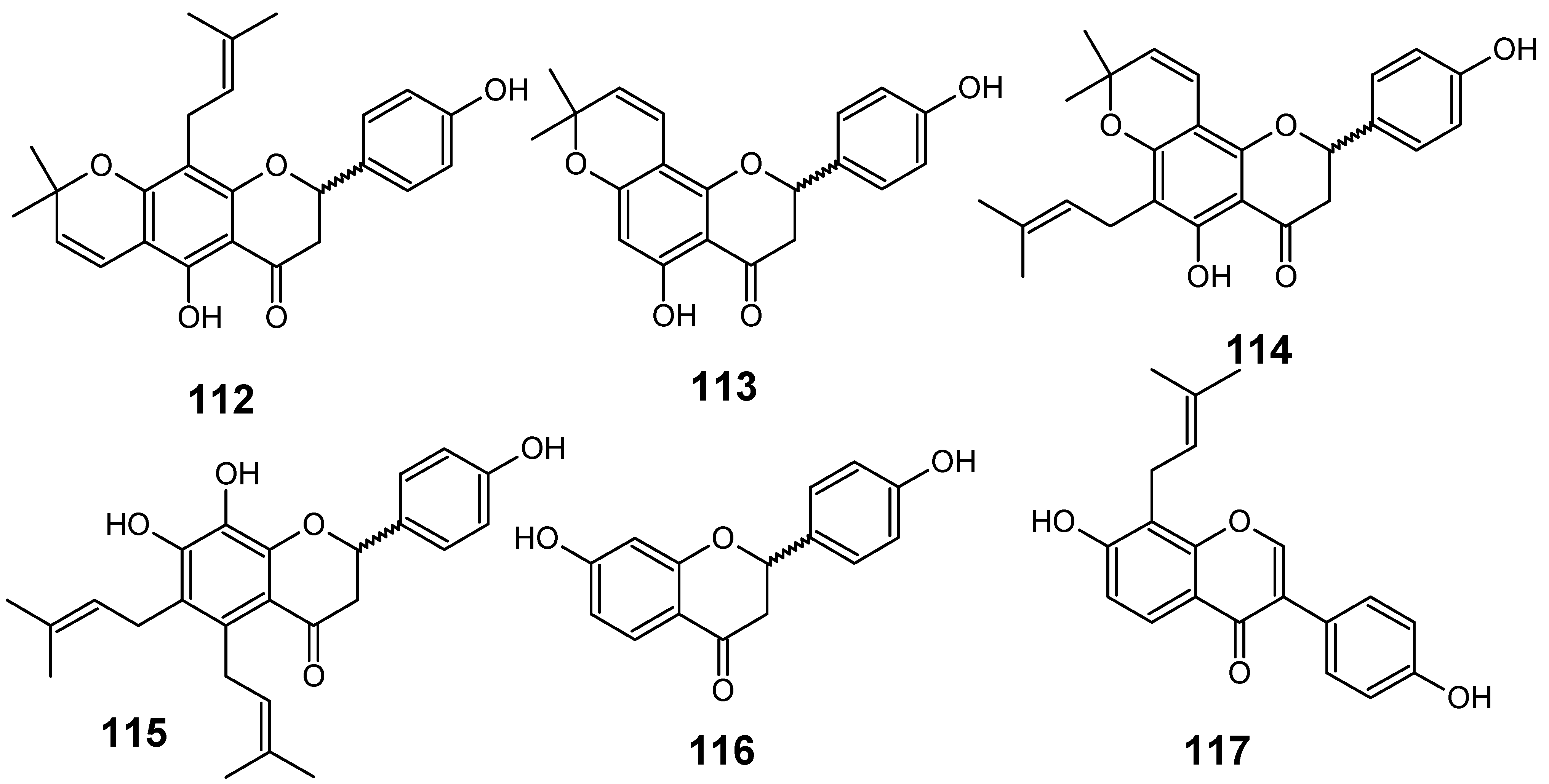
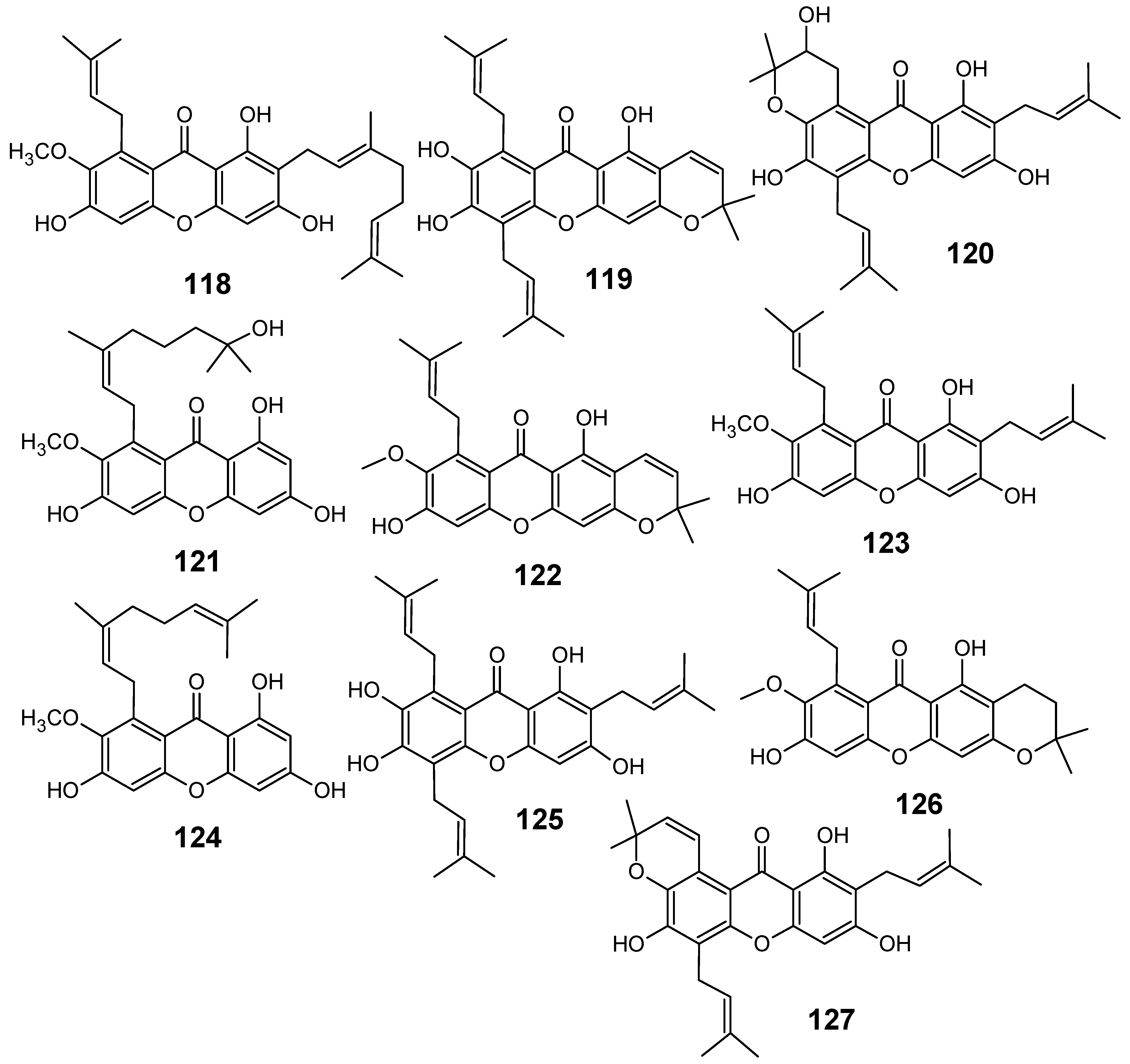
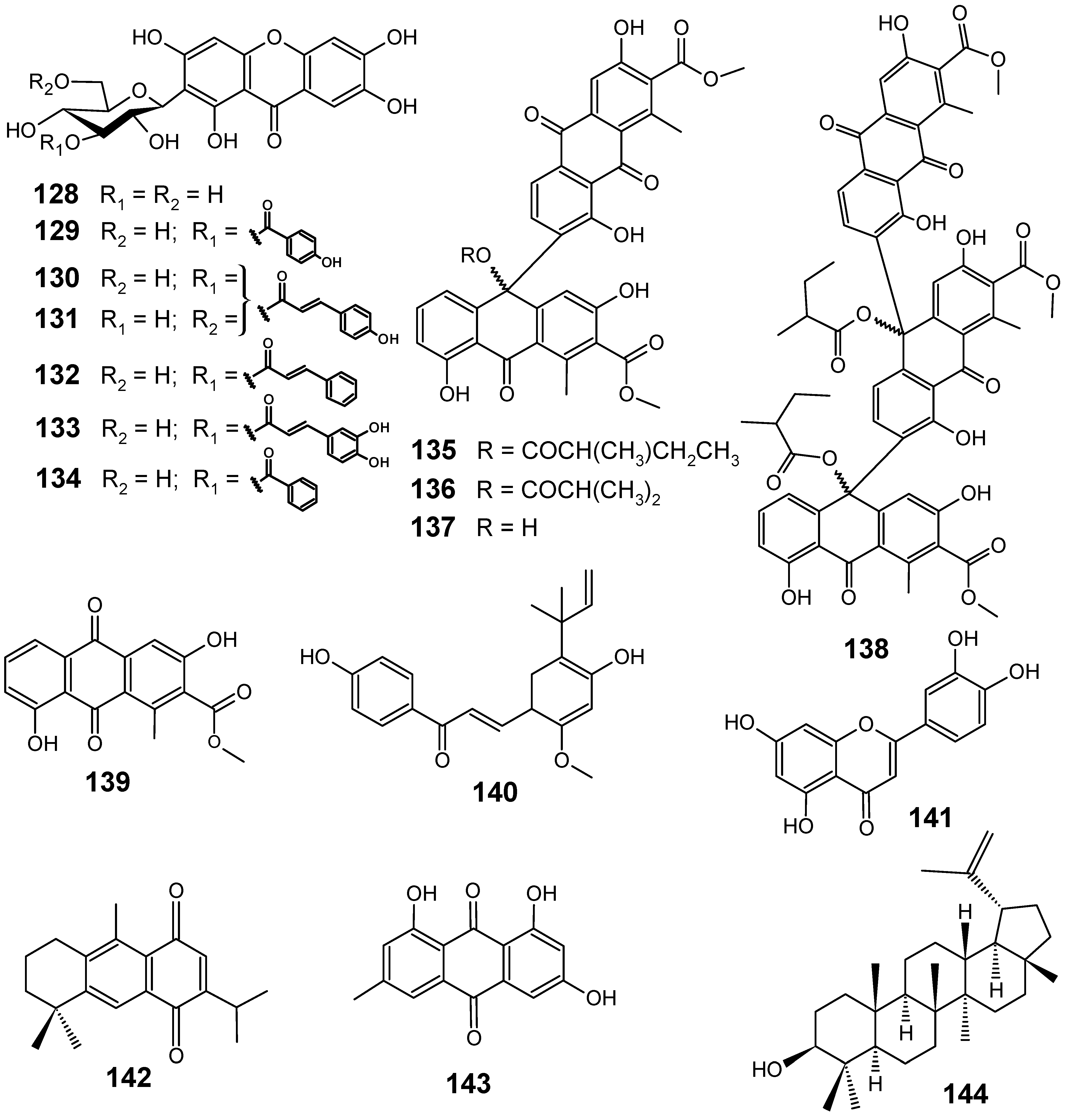
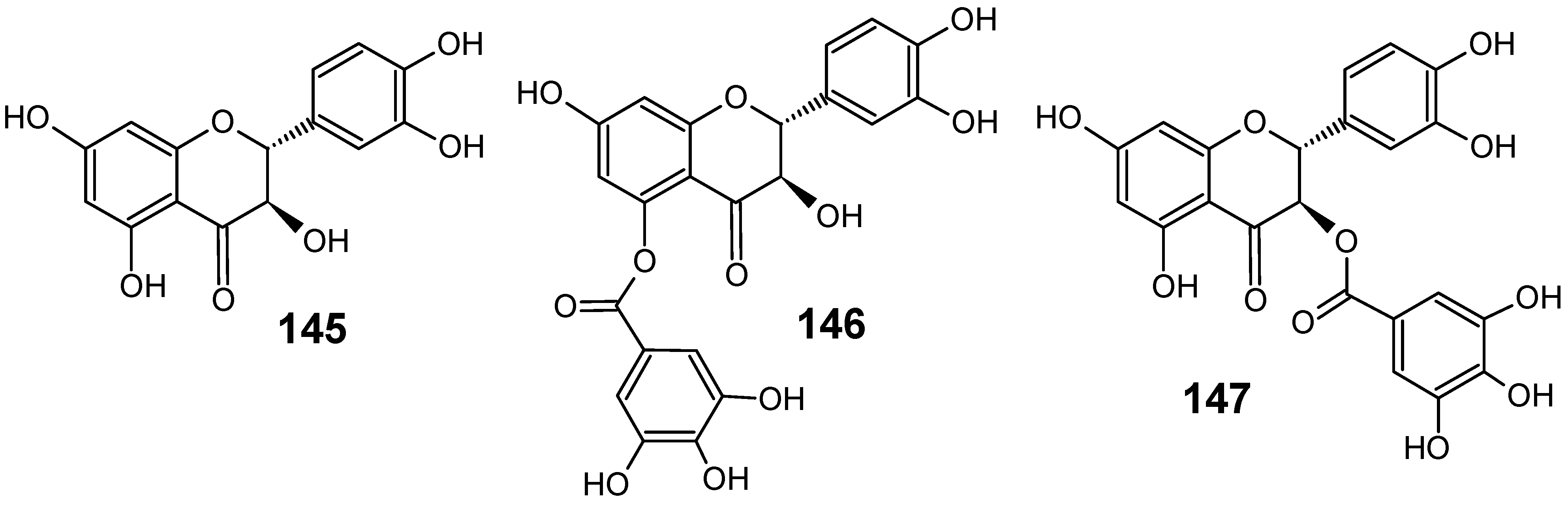
3.2. Flavonoids, chromones, xanthones, anthraquinones and related compounds
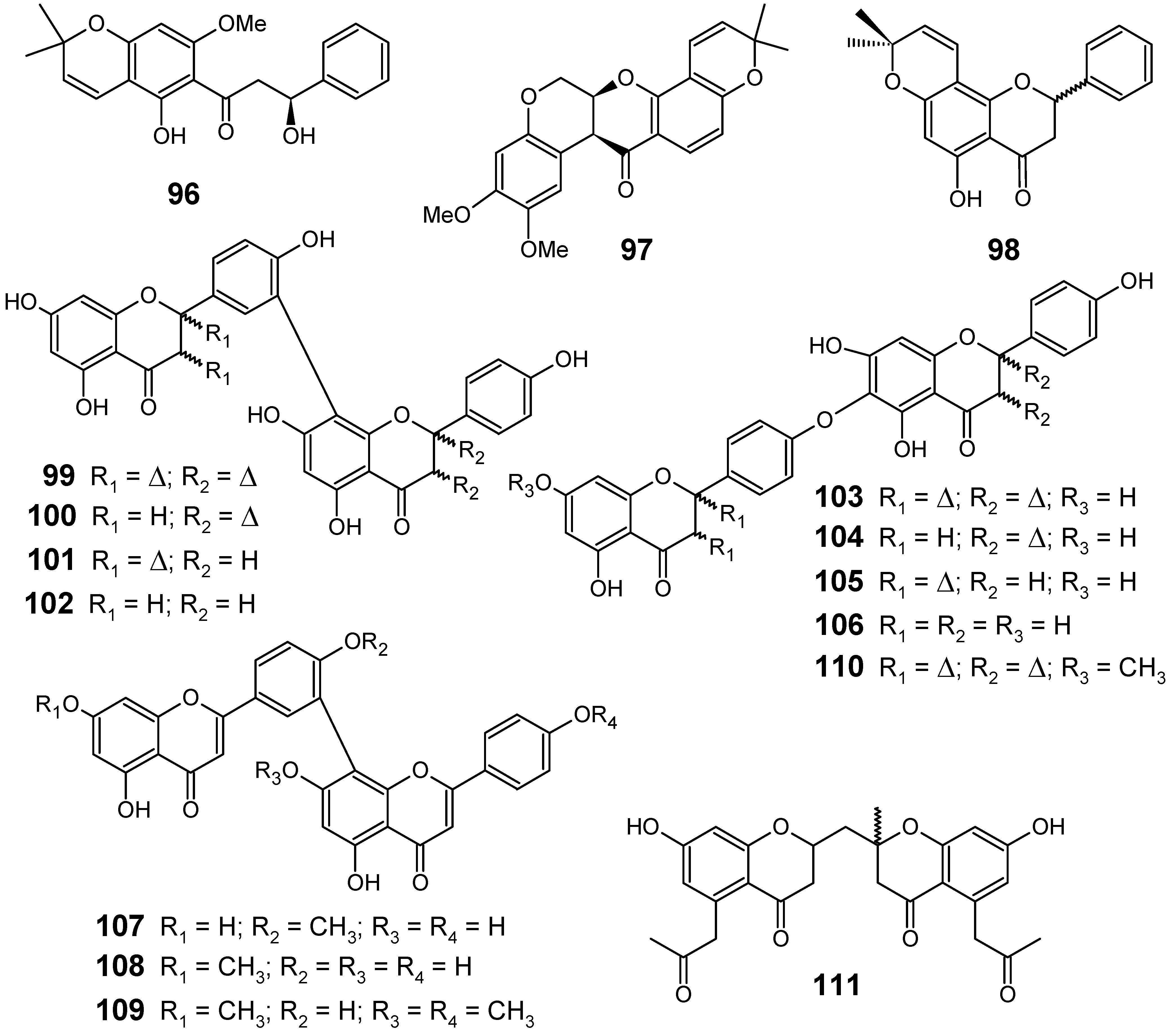

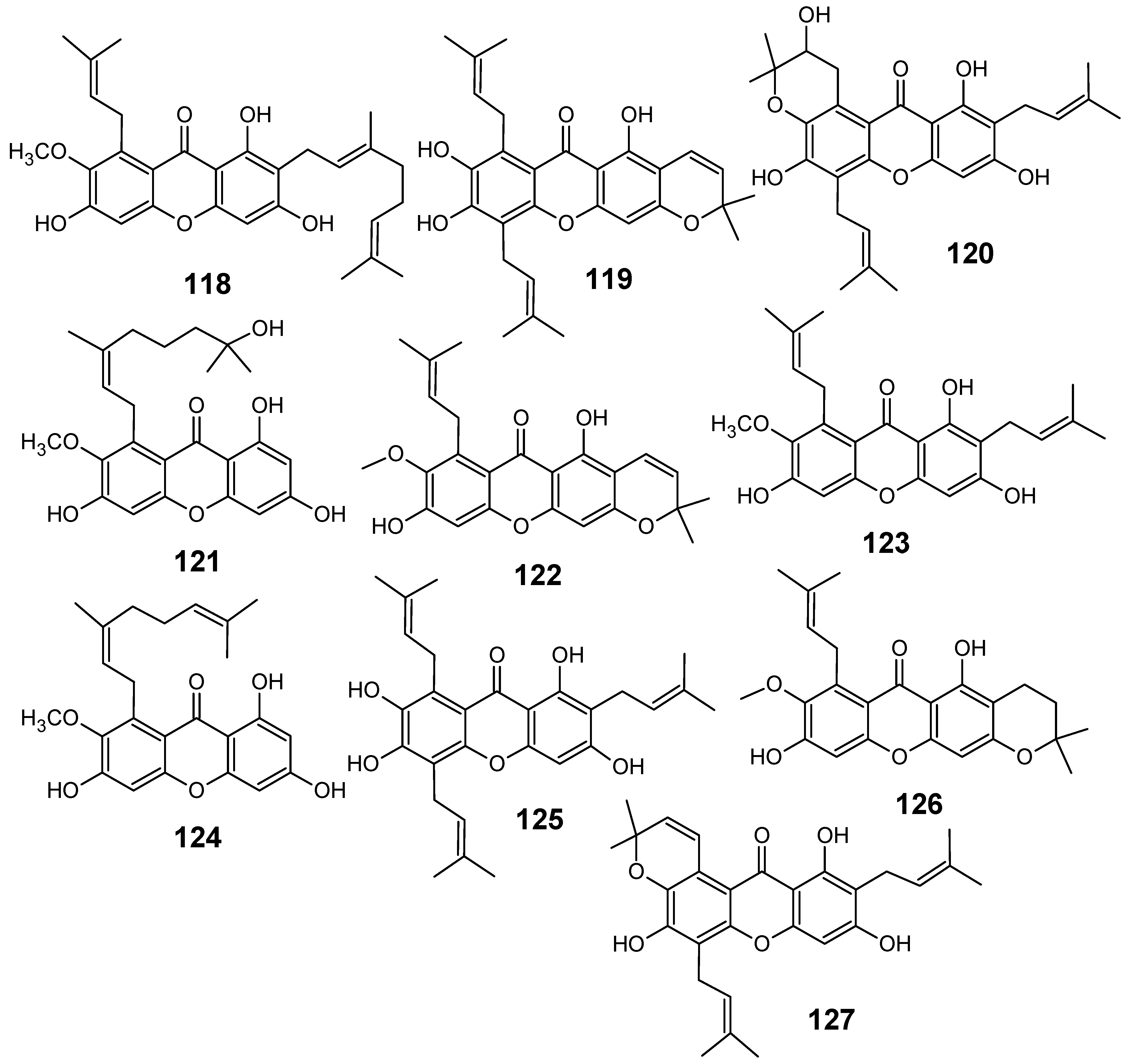
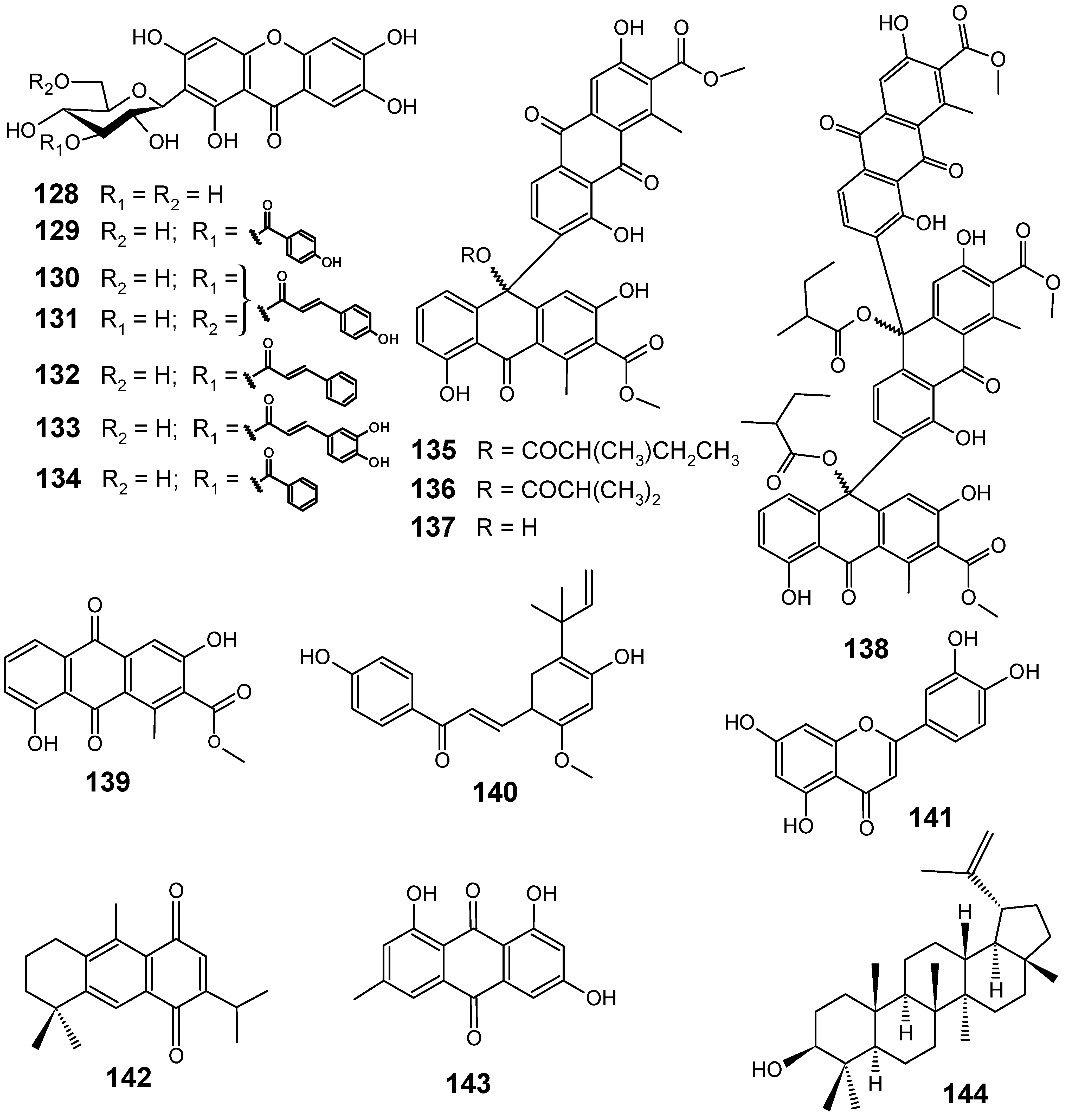
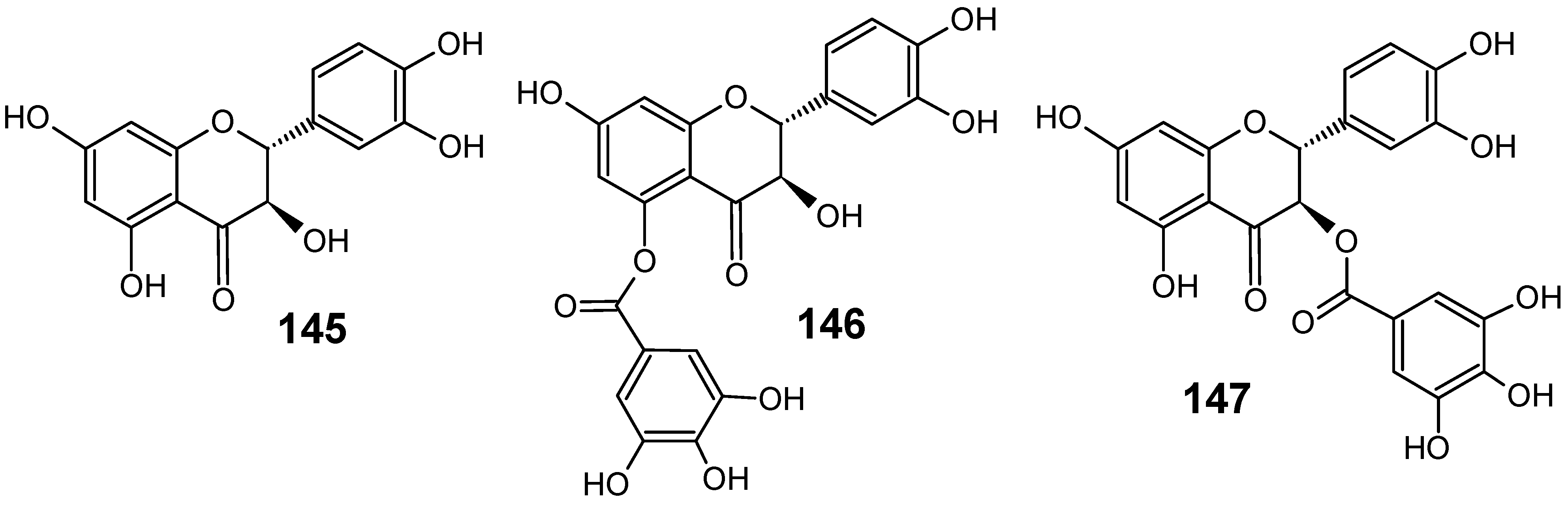
3.3. Miscellaneous compounds

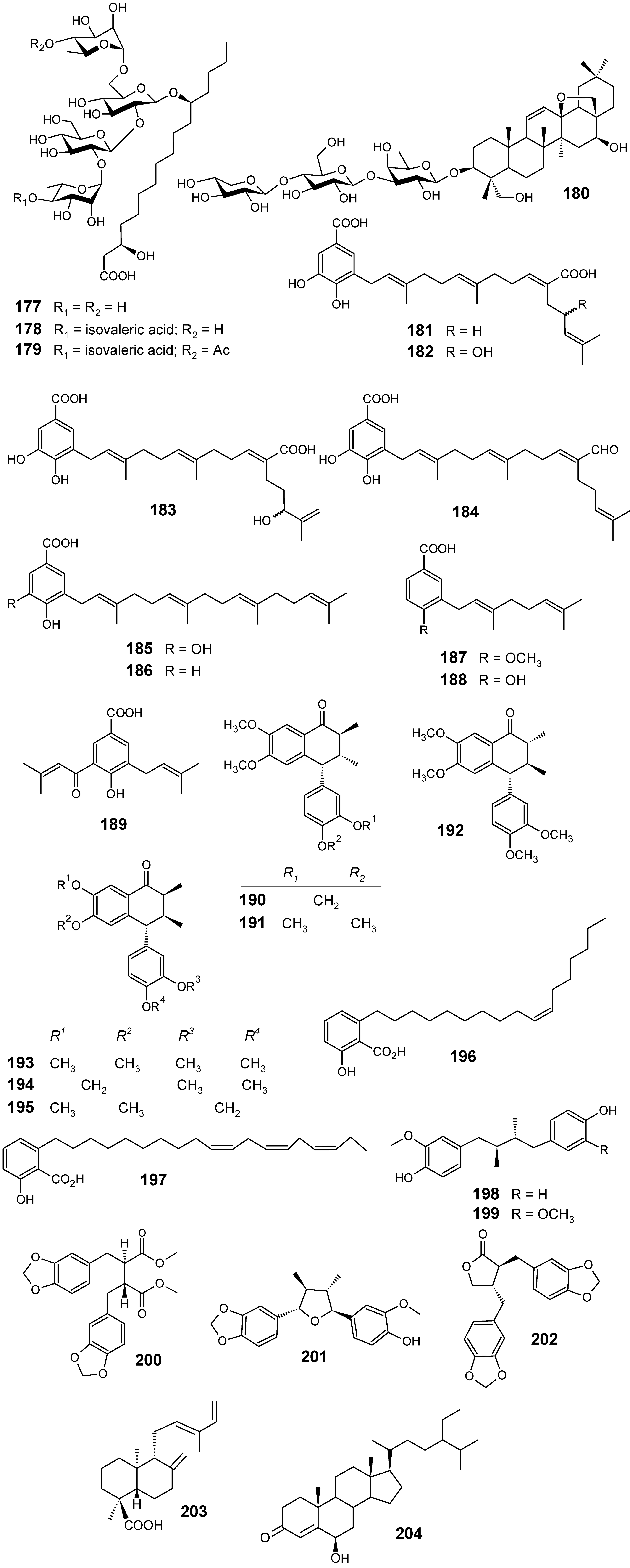
4. Perspectives and Remarks on the Development of New Drugs and Phytomedicines for Malaria
5. Conclusions
Acknowledgements
References
- Greenwood, B.M.; Bojang, K.; Whitty, C.J.; Targett, G.A. Malaria. Lancet 2005, 365, 1487–1498. [Google Scholar] [CrossRef]
- Winter, R.W.; Kelly, J.X.; Smilkstein, M.J.; Dodean, R.; Bagby, G.C.; Rathbun, R.K.; Levin, J.I.; Hinrichs, D.; Riscoe, M.K. Evaluation and lead optimization of anti-malarial acridones. Exp. Parasitol. 2006, 114, 47–56. [Google Scholar] [CrossRef]
- Sachs, J.; Malaney, P. The economic and social burden of malaria. Nature 2002, 415, 680–685. [Google Scholar] [CrossRef]
- WHO. World Malaria Report. World Health Organization: Geneva, Switzerland, 2008. Available online: http://apps.who.int/malaria/wmr2008/.
- Fidock, D.A.; Rosenthal, P.J.; Croft, S.L.; Brun, R.; Nwaka, S. Antimalarial drug discovery: efficacy models for compound screening. Nat. Rev. Drug Discov. 2004, 3, 509–520. [Google Scholar] [CrossRef]
- Deprez-Poulain, R.; Melnyk, P. 1,4-Bis(3-aminopropyl)piperazine libraries: from the discovery of classical chloroquine-like antimalarials to the identification of new targets. Comb. Chem. High Throughput Screen. 2005, 8, 39–48. [Google Scholar] [CrossRef]
- Jones, M.K.; Good, M.F. Malaria parasites up close. Nat. Med. 2006, 12, 170–171. [Google Scholar] [CrossRef]
- Saxena, S.; Pant, N.; Jain, D.C.; Bhakuni, R.S. Antimalarial agents from plant sources. Curr. Sci. 2003, 85, 1314–1329. [Google Scholar]
- Viegas Júnior, C.; Bolzani, V.S.; Barreiro, E.J. Os produtos naturais e a química medicinal moderna. Quím. Nova 2006, 29, 326–337. [Google Scholar]
- Taylor, W.R.; White, N.J. Antimalarial drug toxicity: a review. Drug Saf. 2004, 27, 25–61. [Google Scholar] [CrossRef]
- Jambou, R.; Legrand, E.; Niang, M.; Khim, N.; Lim, P.; Volney, B.; Ekala, M.T.; Bouchier, C.; Esterre, P.; Fandeur, T.; Mercereau-Puijalon, O. Resistance of Plasmodium falciparum field isolates to in vitro artemether and point mutations of the SERCA-type PfATPase6. Lancet 2005, 366, 1960–1963. [Google Scholar] [CrossRef]
- Wichmann, O.; Muhlen, M.; Grub, H.; Mockenhaupt, F.P.; Suttorp, N.; Jelinek, T. Malarone treatment failure not associated with previously described mutations in the cytochrome b gene. Malaria J. 2004, 3, 1–3. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981-2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef]
- Ziegler, H.L.; Staerk, D.; Christensen, J.; Hviid, L.; Hagerstrand, H.; Jaroszewski, J.W. In vitro Plasmodium falciparum drug sensitivity assay: inhibition of parasite growth by incorporation of stomatocytogenic amphiphiles into the erythrocyte membrane. Antimicrob. Agents Chemother. 2002, 46, 1441–1446. [Google Scholar] [CrossRef]
- Kalauni, S.K.; Awale, S.; Tezuka, Y.; Banskota, A.H.; Linn, T.Z.; Asih, P.B.; Syafruddin, D.; Kadota, S. Antimalarial activity of cassane- and norcassane-type diterpenes from Caesalpinia crista and their structure-activity relationship. Biol. Pharm. Bull. 2006, 29, 1050–1052. [Google Scholar] [CrossRef]
- Portet, B.; Fabre, N.; Roumy, V.; Gornitzka, H.; Bourdy, G.; Chevalley, S.; Sauvain, M.; Valentin, A.; Moulis, C. Activity-guided isolation of antiplasmodial dihydrochalcones and flavanones from Piper hostmannianum var. berbicense. Phytochemistry 2007, 68, 1312–1320. [Google Scholar] [CrossRef]
- Frederich, M.; Tits, M.; Angenot, L. Potential antimalarial activity of indole alkaloids. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 11–19. [Google Scholar] [CrossRef]
- Kaur, K.; Jain, M.; Kaur, T.; Jain, R. Antimalarials from nature. Bioorg. Med. Chem. 2009, 17, 3229–3256. [Google Scholar] [CrossRef]
- Oliveira, A.B.; Dolabela, M.F.; Braga, F.C.; Jácome, R.L.R.P.; Varotti, F.P.; Póvoa, M.M. Plant-derived antimalarial agents: new leads and efficient phytomedicines. Part I. Alkaloids. Ann. Braz. Acad. Sci. 2009, in press. [Google Scholar]
- Basco, L.; Mitaku, S.; Skaltsounis, A.L.; Ravelomanantsoa, N.; Tillequin, R.; Koch, M.; Le Bras, J. In vitro activities of furoquinoline and acridone alkaloids against Plasmodium falciparum. Antimicrob. Agents Chemother. 1994, 38, 1169–1171. [Google Scholar] [CrossRef]
- Dolabela, M.F.; Oliveira, S.G.; Nascimento, J.M.; Peres, J.M.; Wagner, H.; Póvoa, M.M.; Oliveira, A.B. In vitro antiplasmodial activity of extract and constituents from Esenbeckia febrifuga, a plant traditionally used to treat malaria in the Brazilian Amazon. Phytomedicine 2008, 15, 367–372. [Google Scholar] [CrossRef]
- Muriithi, M.W.; Abraham, W.R.; Addae-Kyereme, J.; Scowen, I.; Croft, S.L.; Gitu, P.M.; Kendrick, H.; Njagi, E.N.M.; Wright, C.W. Isolation and in vitro antiplasmodial activities of alkaloids from Teclea trichocarpa: in vivo antimalarial activity and X-ray crystal structure of normeliopicine. J. Nat. Prod. 2002, 65, 956–959. [Google Scholar] [CrossRef]
- Pillay, P.; Maharaj, V.J.; Smith, P.J. Investigation South African plants as a source of new antimalarial drugs. J. Ethnopharmacol. 2008, 119, 438–454. [Google Scholar] [CrossRef]
- Mariath, I.R.; Falcão, H.S.; Barbosa-Filho, J.M.; Sousa, L.C.F.; Tomaz, A.C.A.; Batista, L.M.; Diniz, M.F.M.F.; Athayde-Filho, P.F.; Tavares, J.F.; Silva, M.S.; Cunha, E.V.L. Plants of the American continent with antimalarial activity. Braz. J. Pharmacogn. 2009, 19, 158–192. [Google Scholar]
- Zirihi, G.N.; Mambu, L.; Guédé-Guina, F.; Bodo, B.; Grellier, P. In vitro antiplasmodial activity and cytotoxicity of 33 West African plants used for treatment of malaria. J. Ethnopharmacol. 2005, 98, 281–285. [Google Scholar] [CrossRef]
- Koch, A.; Tamez, P.; Pezzuto, J.; Soejarto, D. Evaluation of plants used for antimalarial treatment by the Maasai of Kenya. J. Ethnopharmacol. 2005, 101, 95–99. [Google Scholar] [CrossRef]
- Mbatchi, S.F.; Mbatchi, B.; Banzouzi, J.T.; Bansimba, T.; Nsonde Ntandou, G.F.; Ouamba, J.M.; Berry, A.; Benoit-Vical, F. In vitro antiplasmodial activity of 18 plants used in Congo Brazzaville traditional medicine. J. Ethnopharmacol. 2006, 104, 168–174. [Google Scholar] [CrossRef]
- Menan, H.; Banzouzi, J.T.; Hocquette, A.; Pelissier, Y.; Blache, Y.; Kone, M.; Mallie, M.; Assi, L.A.; Valentin, A. Antiplasmodial activity and cytotoxicity of plants used in West African traditional medicine for the treatment of malaria. J. Ethnopharmacol. 2006, 105, 131–136. [Google Scholar] [CrossRef]
- Kvist, L.P.; Christensen, S.B.; Rasmussen, H.B.; Mejia, K.; Gonzalez, A. Identification and evaluation of Peruvian plants used to treat malaria and leishmaniasis. J. Ethnopharmacol. 2006, 106, 390–402. [Google Scholar] [CrossRef]
- Hout, S.; Chea, A.; Bun, S.S.; Elias, R.; Gasquet, M.; Timon-David, P.; Balansard, G.; Azas, N. Screening of selected indigenous plants of Cambodia for antiplasmodial activity. J. Ethnopharmacol. 2006, 107, 12–18. [Google Scholar] [CrossRef]
- Nguyen-Pouplin, J.; Tran, H.; Phan, T.A.; Dolecek, C.; Farrar, J.; Tran, T.H.; Caron, P.; Bodo, B.; Grellier, P. Antimalarial and cytotoxic activities of ethnopharmacologically selected medicinal plants from South Vietnam. J. Ethnopharmacol. 2007, 109, 417–427. [Google Scholar] [CrossRef]
- de Mesquita, M.L.; Grellier, P.; Mambu, L.; de Paula, J.E.; Espindola, L.S. In vitro antiplasmodial activity of Brazilian Cerrado plants used as traditional remedies. J. Ethnopharmacol. 2007, 110, 165–170. [Google Scholar] [CrossRef]
- Muregi, F.W.; Ishih, A.; Miyase, T.; Suzuki, T.; Kino, H.; Amano, T.; Mkoji, G.M.; Terada, M. Antimalarial activity of methanolic extracts from plants used in Kenyan ethnomedicine and their interactions with chloroquine (CQ) against a CQ-tolerant rodent parasite, in mice. J. Ethnopharmacol. 2007, 111, 190–195. [Google Scholar] [CrossRef]
- Osorio, E.; Arango, G.J.; Jimenez, N.; Alzate, F.; Ruiz, G.; Gutierrez, D.; Paco, M.A.; Gimenez, A.; Robledo, S. Antiprotozoal and cytotoxic activities in vitro of Colombian Annonaceae. J. Ethnopharmacol. 2007, 111, 630–635. [Google Scholar] [CrossRef]
- Roumy, V.; Garcia-Pizango, G.; Gutierrez-Choquevilca, A.L.; Ruiz, L.; Jullian, V.; Winterton, P.; Fabre, N.; Moulis, C.; Valentin, A. Amazonian plants from Peru used by Quechua and Mestizo to treat malaria with evaluation of their activity. J. Ethnopharmacol. 2007, 112, 482–489. [Google Scholar] [CrossRef]
- Muthaura, C.N.; Rukunga, G.M.; Chhabra, S.C.; Omar, S.A.; Guantai, A.N.; Gathirwa, J.W.; Tolo, F.M.; Mwitari, P.G.; Keter, L.K.; Kirira, P.G.; Kimani, C.W.; Mungai, G.M.; Njagi, E.N. Antimalarial activity of some plants traditionally used in treatment of malaria in Kwale district of Kenya. J. Ethnopharmacol. 2007, 112, 545–551. [Google Scholar] [CrossRef]
- Soh, P.N.; Benoit-Vical, F. Are West African plants a source of future antimalarial drugs? J. Ethnopharmacol. 2007, 114, 130–140. [Google Scholar] [CrossRef]
- Mesia, G.K.; Tona, G.L.; Nanga, T.H.; Cimanga, R.K.; Apers, S.; Cos, P.; Maes, L.; Pieters, L.; Vlietinck, A.J. Antiprotozoal and cytotoxic screening of 45 plant extracts from Democratic Republic of Congo. J. Ethnopharmacol. 2008, 115, 409–415. [Google Scholar] [CrossRef]
- Benoit-Vical, F.; Soh, P.N.; Salery, M.; Harguem, L.; Poupat, C.; Nongonierma, R. Evaluation of Senegalese plants used in malaria treatment: focus on Chrozophora senegalensis. J. Ethnopharmacol. 2008, 116, 43–48. [Google Scholar] [CrossRef]
- Kaou, A.M.; Mahiou-Leddet, V.; Hutter, S.; Ainouddine, S.; Hassani, S.; Yahaya, I.; Azas, N.; Ollivier, E. Antimalarial activity of crude extracts from nine African medicinal plants. J. Ethnopharmacol. 2008, 116, 74–83. [Google Scholar] [CrossRef]
- Bero, J.; Ganfon, H.; Jonville, M.C.; Frederich, M.; Gbaguidi, F.; DeMol, P.; Moudachirou, M.; Quetin-Leclercq, J. In vitro antiplasmodial activity of plants used in Benin in traditional medicine to treat malaria. J. Ethnopharmacol. 2009, 122, 439–444. [Google Scholar] [CrossRef]
- Celine, V.; Adriana, P.; Eric, D.; Joaquina, A.C.; Yannick, E.; Augusto, L.F.; Rosario, R.; Dionicia, G.; Michel, S.; Denis, C.; Genevieve, B. Medicinal plants from the Yanesha (Peru): evaluation of the leishmanicidal and antimalarial activity of selected extracts. J. Ethnopharmacol. 2009, 123, 413–422. [Google Scholar] [CrossRef]
- Wright, C.W. Plant derived antimalarials agents: new leads and challenges. Phytochem. Rev. 2005, 4, 55–61. [Google Scholar] [CrossRef]
- Hilou, A.; Nacoulma, O.G.; Guiguemde, T.R. In vivo antimalarial activities of extracts from Amaranthus spinosus L. and Boerhaavia erecta L. in mice. J. Ethnopharmacol. 2006, 103, 236–240. [Google Scholar] [CrossRef]
- Boyom, F.F.; Kemgne, E.M.; Tepongning, R.; Mbacham, W.F.; Tsamo, E.; Zollo, P.H.A.; Gut, J.; Rosenthal, P.J. Antiplasmodial activity of extracts from seven medicinal plants used in malaria treatment in Cameroon. J. Ethnopharmacol. 2009, 123, 483–488. [Google Scholar] [CrossRef]
- Jonville, M.C.; Kodja, H.; Humeau, L.; Fournel, J.; De Mol, P.; Cao, M.; Angenot, L.; Frederich, M. Screening of medicinal plants from Reunion Island for antimalarial and cytotoxic activity. J. Ethnopharmacol. 2008, 120, 382–386. [Google Scholar] [CrossRef]
- Esmaeili, S.; Naghibi, F.; Mosaddegh, M.; Sahranavard, S.; Ghafari, S.; Abdullah, N.R. Screening of antiplasmodial properties among some traditionally used Iranian plants. J. Ethnopharmacol. 2009, 121, 400–404. [Google Scholar] [CrossRef]
- Okokon, J.E.; Antia, B.S.; Igboasoiyi, A.C.; Essien, E.E.; Mbagwu, H.O. Evaluation of antiplasmodial activity of ethanolic seed extract of Picralima nitida. J Ethnopharmacol. 2007, 111, 464–467. [Google Scholar] [CrossRef]
- Abdel-Sattar, E.; Harraz, F.M.; Al-Ansari, S.M.; El-Mekkawy, S.; Ichino, C.; Kiyohara, H.; Otoguro, K.; Omura, S.; Yamada, H. Antiplasmodial and antitrypanosomal activity of plants from the Kingdom of Saudi Arabia. Nat. Med. (Tokyo) 2009, 63, 232–239. [Google Scholar]
- Tona, L.; Cimanga, R.K.; Mesia, K.; Musuamba, C.T.; De Bruyne, T.; Apers, S.; Hernans, N.; Van Miert, S.; Pieters, L.; Totté, J.; Vlietinck, A.J. In vitro antiplasmodial activity of extracts and fractions from seven medicinal plants used in the Democratic Republic of Congo. J. Ethnopharmacol. 2004, 93, 27–32. [Google Scholar] [CrossRef]
- Bah, S.; Jager, A.K.; Adsersen, A.; Diallo, D.; Paulsen, B.S. Antiplasmodial and GABA(A)-benzodiazepine receptor binding activities of five plants used in traditional medicine in Mali, West Africa. J. Ethnopharmacol. 2007, 110, 451–457. [Google Scholar] [CrossRef]
- Okokon, J.E.; Nwafor, P.A. Antiplasmodial activity of root extract and fractions of Croton zambesicus. J. Ethnopharmacol. 2009, 121, 74–78. [Google Scholar] [CrossRef]
- Lenta, B.N.; Vonthron-Senecheau, C.; Soh, R.F.; Tantangmo, F.; Ngouela, S.; Kaiser, M.; Tsamo, E.; Anton, R.; Weniger, B. In vitro antiprotozoal activities and cytotoxicity of some selected Cameroonian medicinal plants. J. Ethnopharmacol. 2007, 111, 8–12. [Google Scholar] [CrossRef]
- Waako, P.J.; Gumede, B.; Smith, P.; Folb, P.I. The in vitro and in vivo antimalarial activity of Cardiospermum halicacabum L. and Momordica foetida Schumch. Et Thonn. J. Ethnopharmacol. 2005, 99, 137–143. [Google Scholar] [CrossRef]
- Khaomek, P.; Ichino, C.; Ishiyama, A.; Sekiguchi, H.; Namatame, M.; Ruangrungsi, N.; Saifah, E.; Kiyohara, H.; Otoguro, K.; Omura, S.; Yamada, H. In vitro antimalarial activity of prenylated flavonoids from Erythrina fusca. J. Nat. Med. 2008, 62, 217–220. [Google Scholar] [CrossRef]
- Okokon, J.E.; Udokpoh, A.E.; Antia, B.S. Antimalaria activity of ethanolic extract of Tetrapleura tetraptera fruit. J. Ethnopharmacol. 2007, 111, 537–540. [Google Scholar] [CrossRef]
- Iwalewa, E.O.; Omisore, N.O.; Adewunmi, C.O.; Gbolade, A.A.; Ademowo, O.G. Anti-protozoan activities of Harungana madagascariensis stem bark extract on trichomonads and malaria. J. Ethnopharmacol. 2008, 117, 507–511. [Google Scholar] [CrossRef]
- Phillipe, G.; Angenot, L.; De Mol, P.; Goffin, E.; Hayette, M.P.; Tits, M.; Frédérich, M. In vitro screening of some Strychnos species for antiplasmodial activity. J. Ethnopharmacol. 2005, 97, 535–539. [Google Scholar] [CrossRef]
- Okokon, J.E.; Ita, B.N.; Udokpoh, A.E. Antiplasmodial activity of Cylicodiscus gabunensis. J. Ethnopharmacol. 2006, 107, 175–178. [Google Scholar] [CrossRef]
- Abosl, A.O.; Mbukwa, E.; Majinda, R.R.; Raserok, B.H.; Yenesew, A.; Midiwo, J.O.; Akala, H.; Liyala, P.; Waters, N.C. Vangueria infausta root bark: in vivo and in vitro antiplasmodial activity. Br. J. Biomed. Sci. 2006, 63, 129–133. [Google Scholar]
- Lategan, C.A.; Campbell, W.E.; Seaman, T.; Smith, P.J. The bioactivity of novel furanoterpenoids isolated from Siphonochilus aethiopicus. J. Ethnopharmacol. 2009, 121, 92–97. [Google Scholar] [CrossRef]
- Pedersen, M.M.; Chukwujekwu, J.C.; Lategan, C.A.; Staden, J.; Smith, P.J.; Staerk, D. Antimalarial sesquiterpene lactones from Distephanus angulifolius. Phytochemistry 2009, 70, 601–607. [Google Scholar] [CrossRef]
- Efange, S.M.; Brun, R.; Wittlin, S.; Connolly, J.D.; Hoye, T.R.; McAkam, T.; Makolo, F.L.; Mbah, J.A.; Nelson, D.P.; Nyongbela, K.D.; Wirmum, C.K. Okundoperoxide, a bicyclic cyclo-farnesylsesquiterpene endoperoxide from Scleria striatinux with antiplasmodial activity. J. Nat. Prod. 2009, 72, 280–283. [Google Scholar] [CrossRef]
- Murata, T.; Miyase, T.; Muregi, F.W.; Naoshima-Ishibashi, Y.; Umehara, K.; Warashina, T.; Kanou, S.; Mkoji, G.M.; Terada, M.; Ishih, A. Antiplasmodial triterpenoids from Ekebergia capensis. J. Nat. Prod. 2008, 71, 167–174. [Google Scholar] [CrossRef]
- Jonville, M.C.; Capel, M.; Frederich, M.; Angenot, Luc; Dive, G.; Faure, R.; Azas, N.; Ollivier, E. Fagraldehyde, a secoiridoid isolated from Fagraea fragrans. J. Nat. Prod. 2008, 71, 2038–2040. [Google Scholar] [CrossRef]
- Adelekan, A.M.; Prozesky, E.A.; Hussein, A.A.; Urena, L.D.; van Rooyen, P.H.; Liles, D.C.; Meyer, J.J.; Rodriguez, B. Bioactive diterpenes and other constituents of Croton steenkampianus. J. Nat. Prod. 2008, 71, 1919–1922. [Google Scholar] [CrossRef]
- Matsuno, Y.; Deguchi, J.; Hirasawa, Y.; Ohyama, K.; Toyoda, H.; Hirobe, C.; Ekasari, W.; Widyawaruyanti, A.; Zaini, N.C.; Morita, H. Sucutiniranes A and B, new cassane-type diterpenes from Bowdichia nitida. Bioorg. Med. Chem. Lett. 2008, 18, 3774–3777. [Google Scholar] [CrossRef]
- Kamatou, G.P.P.; Van Zyl, R.L.; Davids, H.; Heerden, F.R.V.; Lourens, A.C.U.; Viljoen, A.M. Antimalarial and anticancer activities of selected South African Salvia species and isolated compounds from S. radula. S. Afr. J. Bot. 2008, 74, 238–243. [Google Scholar] [CrossRef]
- Chukwujekwu, J.C.; Lategan, C.A.; Smith, P.J.; Van Heerden, F.R.; Van Staden, J. Antiplasmodial and cytotoxic activity of isolated sesquiterpene lactones from the acetone leaf extract of Vernonia colorata. S. Afr. J. Bot. 2009, 75, 176–179. [Google Scholar]
- Olagnier, D.; Costes, P.; Berry, A.; Linas, M.D.; Urrutigoity, M.; Dechy-Cabaret, O.; Benoit-Vical, F. Modifications of the chemical structure of terpenes in antiplasmodial and antifungal drug research. Bioorg. Med. Chem. Lett. 2007, 17, 6075–6078. [Google Scholar] [CrossRef]
- Ngouamegne, E.T.; Fongang, R.S.; Ngouela, S.; Boyom, F.F.; Rohmer, M.; Tsamo, E.; Gut, J.; Rosenthal, P.J. Endodesmiadiol, a friedelane triterpenoid, and other antiplasmodial compounds from Endodesmia calophylloides. Chem. Pharm. Bull. (Tokyo) 2008, 56, 374–377. [Google Scholar] [CrossRef]
- Moon, H.I. Antiplasmodial activity of ineupatorolides A from Carpesium rosulatum. Parasitol. Res. 2007, 100, 1147–1149. [Google Scholar] [CrossRef]
- Chung, I.M.; Kim, M.Y.; Moon, H.I. Antiplasmodial activity of sesquiterpene lactone from Carpesium rosulatum in mice. Parasitol. Res. 2008, 103, 341–344. [Google Scholar] [CrossRef]
- de Sa, M.S.; Costa, J.F.; Krettli, A.U.; Zalis, M.G.; Maia, G.L.; Sette, I.M.; Camara, C.D.; Filho, J.M.; Giulietti-Harley, A.M.; Ribeiro Dos Santos, R.; Soares, M.B. Antimalarial activity of betulinic acid and derivatives in vitro against Plasmodium falciparum and in vivo in P. berghei-infected mice. Parasitol. Res. 2009, 105, 275–279. [Google Scholar] [CrossRef]
- Ortet, R.; Prado, S.; Mouray, E.; Thomas, O.P. Sesquiterpene lactones from the endemic Cape Verdean Artemisia gorgonum. Phytochemistry 2008, 69, 2961–2965. [Google Scholar] [CrossRef]
- Afolayan, A.F.; Mann, M.G.; Lategan, C.A.; Smith, P.J.; Bolton, J.J.; Beukes, D.R. Antiplasmodial halogenated monoterpenes from the marine red alga Plocamium cornutum. Phytochemistry 2009, 70, 597–600. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Chen, F.; Androulakis, X.M.; Wargovich, M.J. Anticancer activity of limonoid from Khaya senegalensis. Phytother. Res. 2007, 21, 731–734. [Google Scholar] [CrossRef]
- Roy, A.; Saraf, S. Limonoids: Overview of significant bioactive triterpenes distributed in plant kingdom. Biol. Pharm. Bull. 2006, 29, 191–201. [Google Scholar] [CrossRef]
- Mohamad, K.; Hirasawa, Y.; Litaudon, M.; Awang, K.; Hadi, A.H.; Takeya, K.; Ekasari, W.; Widyawaruyanti, A.; Zaini, N.C.; Morita, H. Ceramicines B-D, new antiplasmodial limonoids from Chisocheton ceramicus. Bioorg Med Chem 2009, 17, 727–730. [Google Scholar] [CrossRef]
- Banzouzi, J.T.; Soh, P.N.; Mbatchi, B.; Cave, A.; Ramos, S.; Retailleau, P.; Rakotonandrasana, O.; Berry, A.; Benoit-Vical, F. Cogniauxia podolaena: bioassay-guided fractionation of defoliated stems, isolation of active compounds, antiplasmodial activity and cytotoxicity. Planta Med. 2008, 74, 1453–1456. [Google Scholar] [CrossRef]
- Muiva, L.M.; Yenesew, A.; Derese, S.; Heydenreich, M.; Peter, M.G.; Akala, H.M.; Eyase, F.; Waters, N.C.; Mutai, C.; Keriko, J.M.; Walsh, D. Antiplasmodial β-hydroxydihydrochalcone from seedpods of Tephrosia elata. Phytochem. Lett. 2009, in press. [Google Scholar] [CrossRef]
- Kunert, O.; Swamy, R.C.; Kaiser, M.; Presser, A.; Buzzi, S.; Appa-Rao, A.V.N.; Shuhly, W. Antiplasmodial and leishmanicidal activity of biflavonoids from Indian Selaginella bryopteris. Phytochem. Lett. 2008, 1, 171–174. [Google Scholar] [CrossRef]
- Oshimi, S.; Tomizawa, Y.; Hirasawa, Y.; Honda, T.; Ekasari, W.; Widyawaruyanti, A.; Rudyanto, M.; Indrayanto, G.; Zaini, N.C.; Morita, H. Chrobisiamone A, a new bischromone from Cassia siamea and a biomimetic transformation of 5-acetonyl-7-hydroxy-2-methylchromone into cassiarin A. Bioorg. Med. Chem. Lett. 2008, 18, 3761–3763. [Google Scholar] [CrossRef]
- Zelefack, F.; Guilet, D.; Fabre, N.; Bayet, C.; Chevalley, S.; Ngouela, S.; Lenta, B.N.; Valentin, A.; Tsamo, E.; Dijoux-Franca, M.G. Cytotoxic and antiplasmodial xanthones from Pentadesma butyracea. J. Nat. Prod. 2009, 72, 954–957. [Google Scholar] [CrossRef]
- Martin, F.; Hay, A.E.; Cressende, D.; Reist, M.; Vivas, L.; Gupta, M.P.; Carrupt, P.A.; Hostettmann, K. Antioxidant c-glucosylxanthones from the leaves of Arrabidaea patellifera. J. Nat. Prod. 2008, 71, 1887–1890. [Google Scholar] [CrossRef]
- Hou, Y.; Cao, S.; Brodie, P.J.; Callmander, M.W.; Ratovoson, F.; Rakotobe, E.A.; Rasamison, V.E.; Ratsimbason, M.; Alumasa, J.N.; Roepe, P.D.; Kingston, D.G. Antiproliferative and antimalarial anthraquinones of Scutia myrtina from the Madagascar forest. Bioorg. Med. Chem. 2009, 17, 2871–2876. [Google Scholar] [CrossRef]
- Ziegler, H.L.; Hansen, H.S.; Staerk, D.; Christensen, S.B.; Hägerstrand, H.; Jaroszewski, J.W. The antiparasitic compound licochalcone A is a potent echinocytogenic agent that modifies the erythrocyte membrane in the concentration range where antiplasmodial activity is observed. Antimicrob. Agents Chemother. 2004, 48, 4067–4071. [Google Scholar] [CrossRef]
- Mishra, L.C.; Bhattacharya, A.; Bhasin, V.K. Phytochemical licochalcone A enhances antimalarial activity of artemisinin in vitro. Acta Trop. 2009, 109, 194–198. [Google Scholar] [CrossRef]
- Lehane, A.M.; Saliba, K.J. Common dietary flavonoids inhibit the growth of the intraerythrocytic malaria parasite. BMC Res. Notes 2008, 1, 26. [Google Scholar] [CrossRef]
- Moein, M.R.; Pawar, R.S.; Khan, S.I.; Tekwani, B.L.; Khan, I.A. Antileishmanial, antiplasmodial and cytotoxic activities of 12,16-dideoxy aegyptinone B from Zhumeria majdae Rech.f. & Wendelbo. Phytother. Res. 2008, 22, 283–285. [Google Scholar] [CrossRef]
- Ajaiyeoba, E.O.; Oladepo, O.; Fawole, O.I.; Bolaji, O.M.; Akinboye, D.O.; Ogundahunsi, O.A.T.; Falade, C.O.; Gbotosho, G.O.; Itiola, O.A.; Happi, T.C.; Ebong, O.O.; Ononiwu, I.M.; Osowole, O.S.; Oduola, O.O.; Ashidi, J.S.; Oduola, A.M.J. Cultural categorization of febrile illnesses in correlation with herbal remedies for treatment in Southwestern Nigeria. J. Ethnopharmacol. 2003, 85, 179–185. [Google Scholar] [CrossRef]
- Ajaiyeoba, E.O.; Bolaji, O.M.; Akinboye, D.O.; Falade, C.O.; Gbotosho, G.O.; Ashidi, J.S.; Okpako, L.C.; Oduola, O.O.; Falade, M.O.; Itiola, O.A.; Houghton, P.J.; Wright, C.W.; Ogundahunsi, O.A.; Oduola, A.M.J. In vitro antiplasmodial and cytotoxic activities of plants used as antimalarial agents in the southwest Nigerian ethnomedicine. J. Nat. Rem. 2005, 5, 1–6. [Google Scholar]
- Ajaiyeoba, E.O.; Ashidi, J.S.; Okpako, L.C.; Houghton, P.J.; Wright, C.W. Antiplasmodial compounds from Cassia siamea stem bark extract. Phytother. Res. 2008, 22, 254–255. [Google Scholar] [CrossRef]
- Ramanandraibe, V.; Grellier, P.; Martin, M.T.; Deville, A.; Joyeau, R.; Ramanitrahasimbola, D.; Mouray, E.; Rasoanaivo, P.; Mambu, L. Antiplasmodial phenolic compounds from Piptadenia pervillei. Planta Med. 2008, 74, 417–421. [Google Scholar] [CrossRef]
- Roumy, V.; Fabre, N.; Portet, B.; Bourdy, G.; Acebey, L.; Vigor, C.; Valentin, A.; Moulis, C. Four anti-protozoal and anti-bacterial compounds from Tapirira guianensis. Phytochemistry 2009, 70, 305–311. [Google Scholar] [CrossRef]
- Flores, N.; Jimenez, I.A.; Gimenez, A.; Ruiz, G.; Gutierrez, D.; Bourdy, G.; Bazzocchi, I.L. Benzoic acid derivatives from Piper species and their antiparasitic activity. J. Nat. Prod. 2008, 71, 1538–1543. [Google Scholar] [CrossRef]
- Marti, G.; Eparvier, V.; Moretti, C.; Susplugas, S.; Prado, S.; Grellier, P.; Retailleau, P.; Guéritte, F.; Litaudon, M. Antiplasmodial benzophenones from the trunk latex of Moronobea coccinea (Clusiaceae). Phytochemistry 2009, 70, 75–85. [Google Scholar] [CrossRef]
- Tasdemir, D.; Brun, R.; Franzblau, S.G.; Sezgin, Y.; Çalis, I. Evaluation of antiprotozoal and antimycobacterial activities of the resin glycosides and the other metabolites of Scrophularia cryptophila. Phytomedicine 2008, 15, 209–215. [Google Scholar] [CrossRef]
- Flores, N.; Jimenez, I.A.; Gimenez, A.; Ruiz, G.; Gutierrez, D.; Bourdy, G.; Bazzocchi, I.L. Antiparasitic activity of prenylated benzoic acid derivatives from Piper species. Phytochemistry 2009, 70, 621–627. [Google Scholar] [CrossRef]
- de Andrade-Neto, V.F.; da Silva, T.; Lopes, L.M.; do Rosario, V.E.; de Pilla Varotti, F.; Krettli, A.U. Antiplasmodial activity of aryltetralone lignans from Holostylis reniformis. Antimicrob. Agents Chemother. 2007, 51, 2346–2350. [Google Scholar] [CrossRef]
- Lee, S.J.; Park, W.H.; Moon, H.I. Bioassay-guided isolation of antiplasmodial anacardic acids derivatives from the whole plants of Viola websteri Hemsl. Parasitol. Res. 2009, 104, 463–466. [Google Scholar] [CrossRef]
- Abrantes, M.; Mil-Homens, T.; Duarte, N.; Lopes, D.; Cravo, P.; Madureira, M.; Ferreira, M.J. Antiplasmodial activity of lignans and extracts from Pycnanthus angolensis. Planta Med. 2008, 74, 1408–1412. [Google Scholar] [CrossRef]
- Willcox, M.L.; Bodeker, G. Traditional herbal medicines for malaria. Brit. Med. J. 2004, 329, 1156–1159. [Google Scholar] [CrossRef]
- Harborne, J.B. Twenty-five years of chemical ecology. Nat. Prod. Rep. 2001, 18, 361–379. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Batista, R.; De Jesus Silva Júnior, A.; De Oliveira, A.B. Plant-Derived Antimalarial Agents: New Leads and Efficient Phytomedicines. Part II. Non-Alkaloidal Natural Products. Molecules 2009, 14, 3037-3072. https://doi.org/10.3390/molecules14083037
Batista R, De Jesus Silva Júnior A, De Oliveira AB. Plant-Derived Antimalarial Agents: New Leads and Efficient Phytomedicines. Part II. Non-Alkaloidal Natural Products. Molecules. 2009; 14(8):3037-3072. https://doi.org/10.3390/molecules14083037
Chicago/Turabian StyleBatista, Ronan, Ademir De Jesus Silva Júnior, and Alaíde Braga De Oliveira. 2009. "Plant-Derived Antimalarial Agents: New Leads and Efficient Phytomedicines. Part II. Non-Alkaloidal Natural Products" Molecules 14, no. 8: 3037-3072. https://doi.org/10.3390/molecules14083037
APA StyleBatista, R., De Jesus Silva Júnior, A., & De Oliveira, A. B. (2009). Plant-Derived Antimalarial Agents: New Leads and Efficient Phytomedicines. Part II. Non-Alkaloidal Natural Products. Molecules, 14(8), 3037-3072. https://doi.org/10.3390/molecules14083037