
Synergistic Effect of the Long-Term Overexpression of Bcl-2 and BDNF Lentiviral in Cell Protecting against Death and Generating TH Positive and CHAT Positive Cells from MSC

 , and
, and
Abstract
:
1. Introduction
2. Results
- (a)
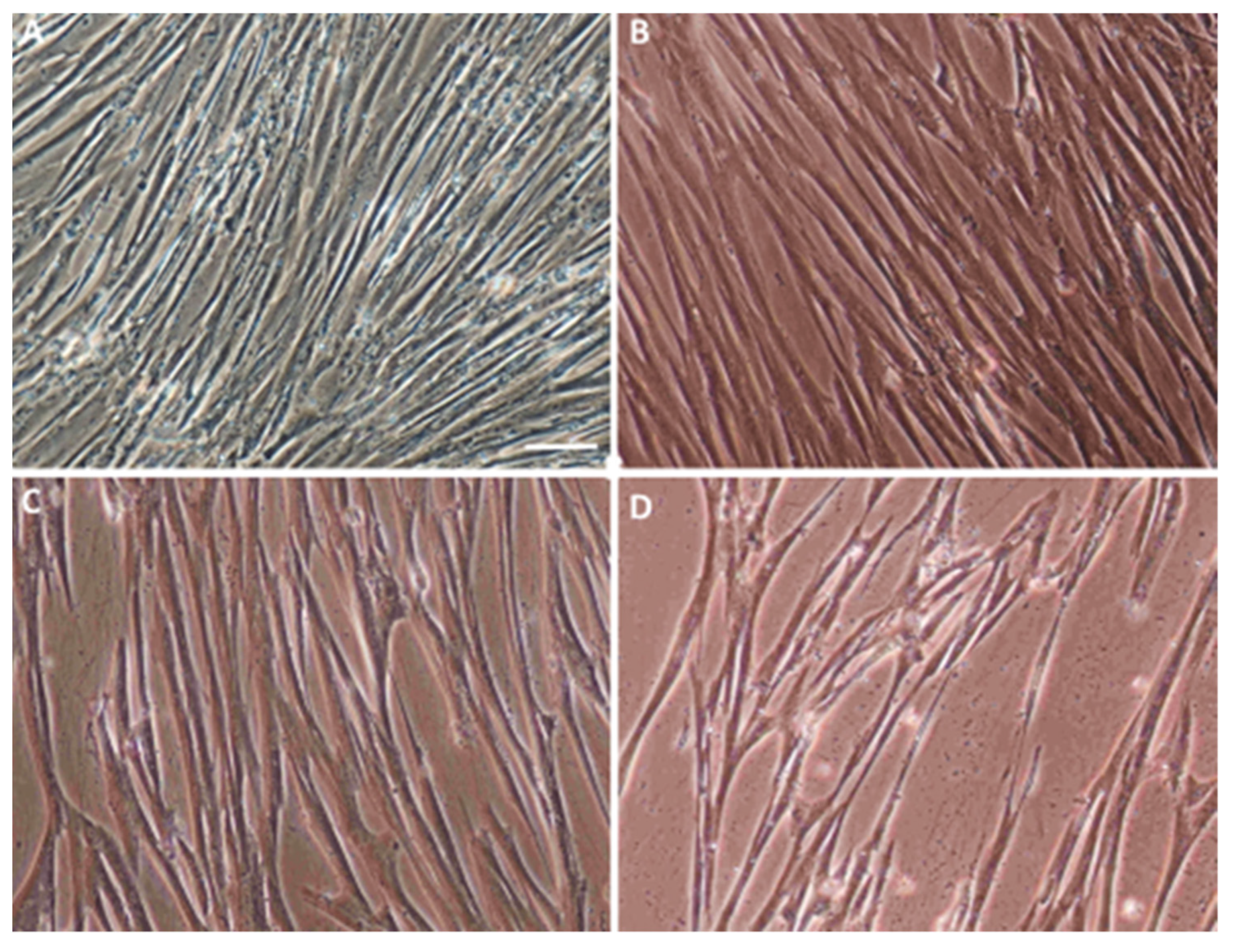
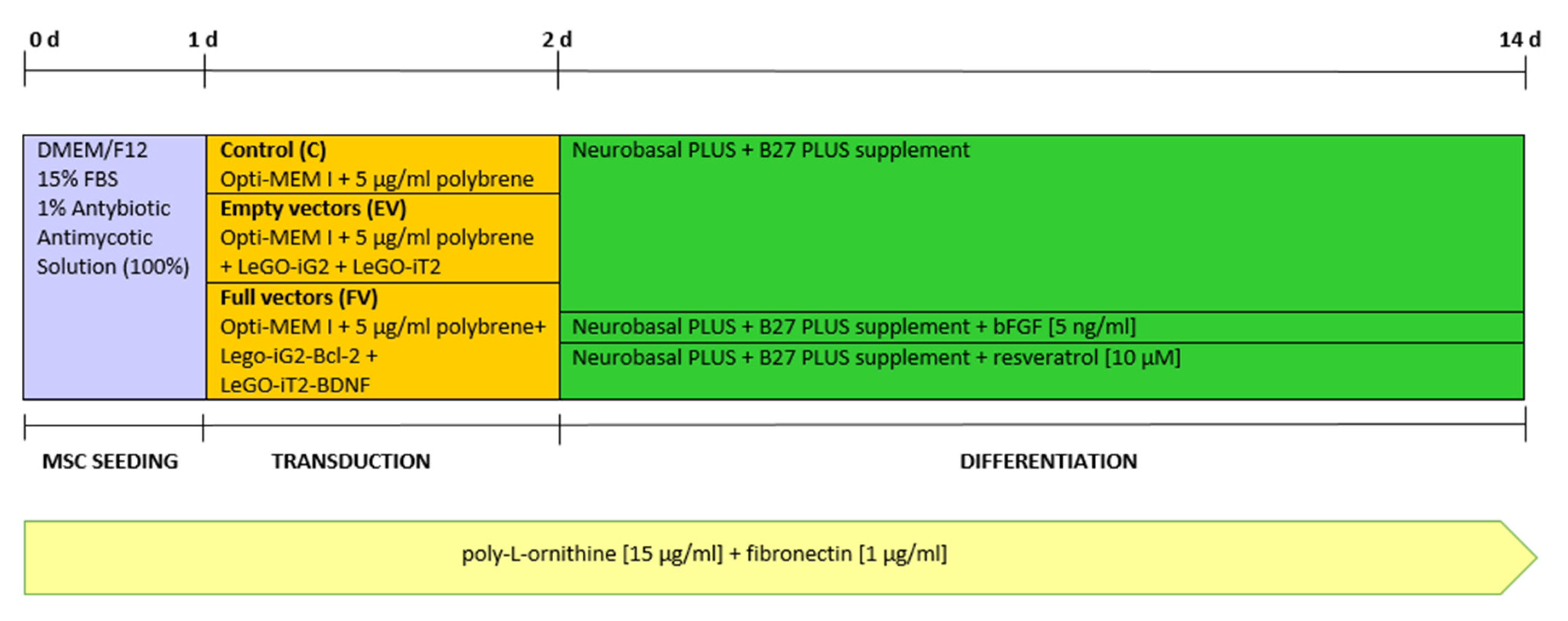
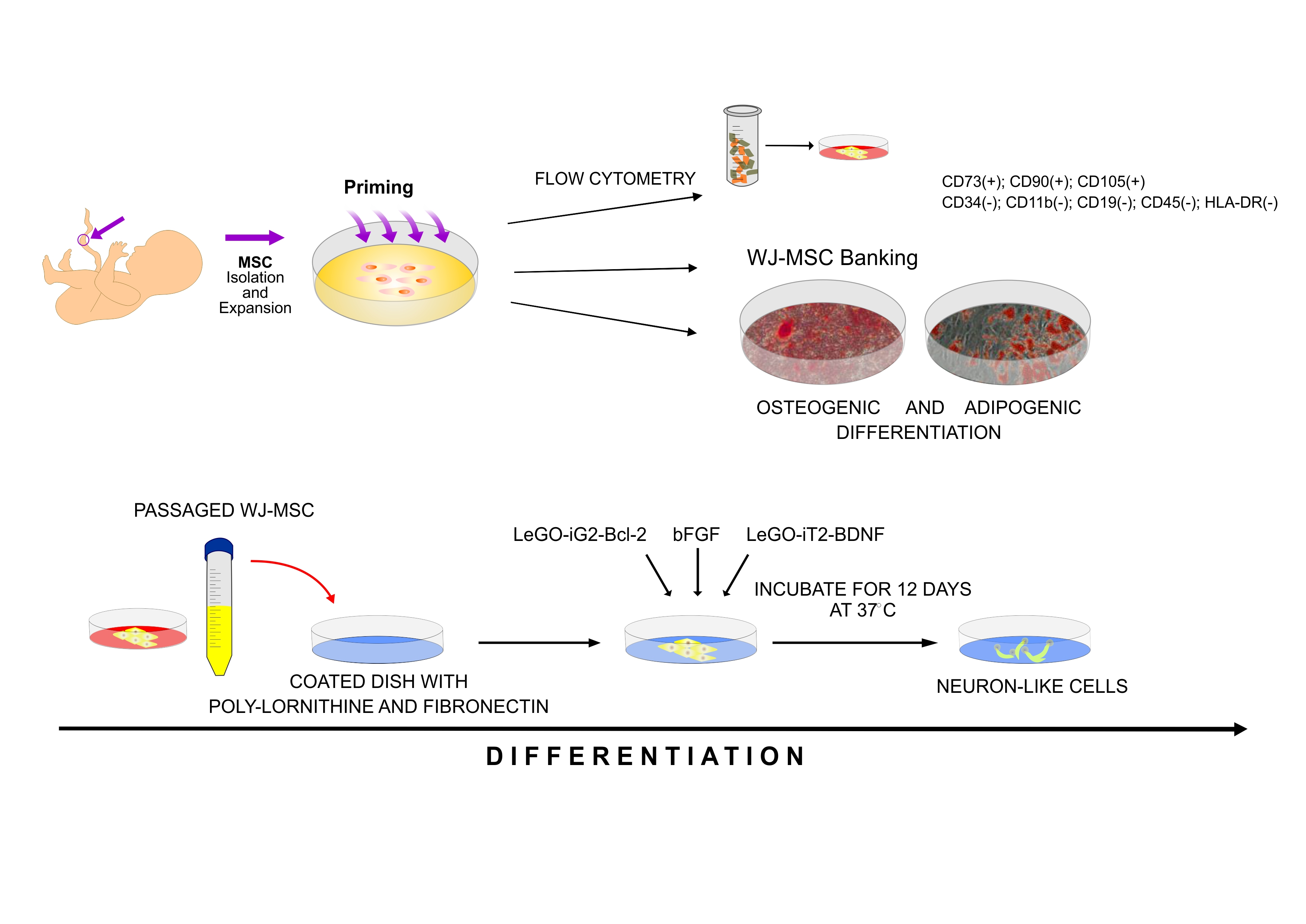
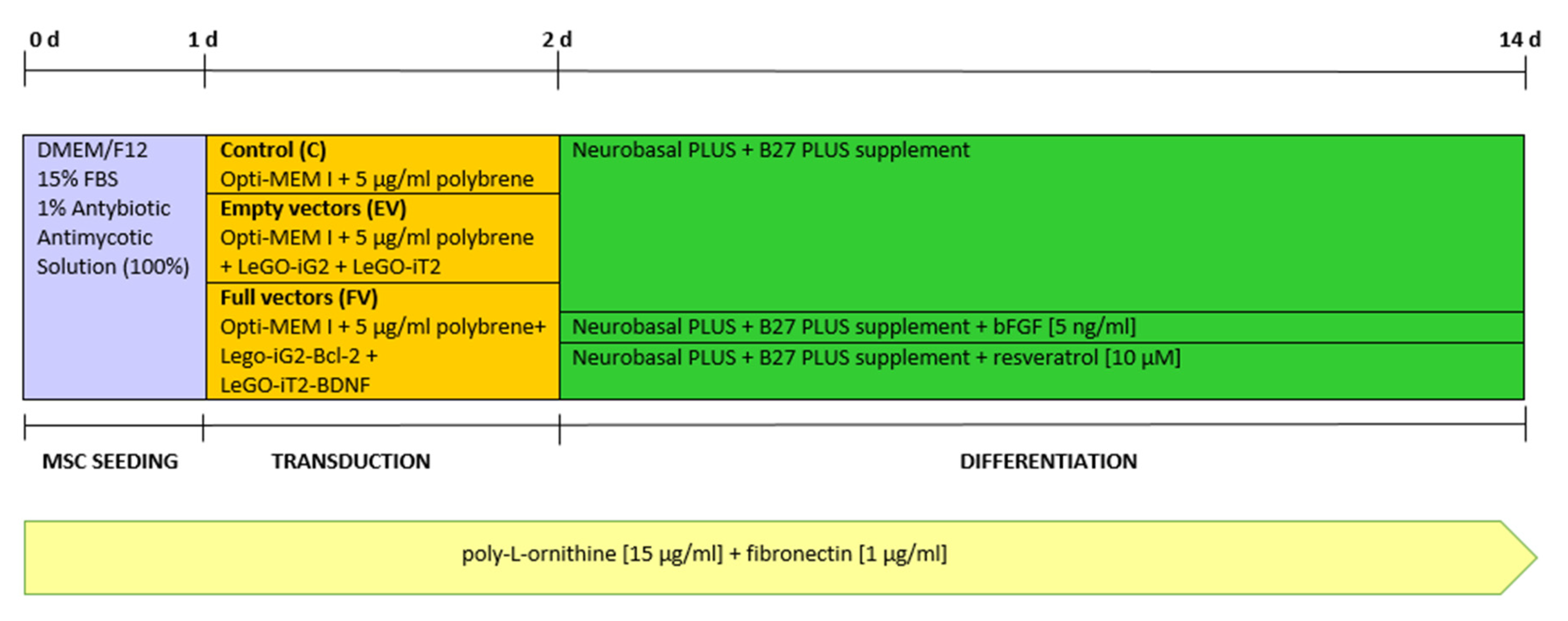
- The control, marked with the working symbol (C), was the WJ-MSC culture. These cells were not subjected to lentiviral transduction or any additional substances. Their culture was carried out in parallel with the cultures of the cells from the other two groups in an analogous culture medium, but without any additional stimulators (Figure 1B).
- (b)
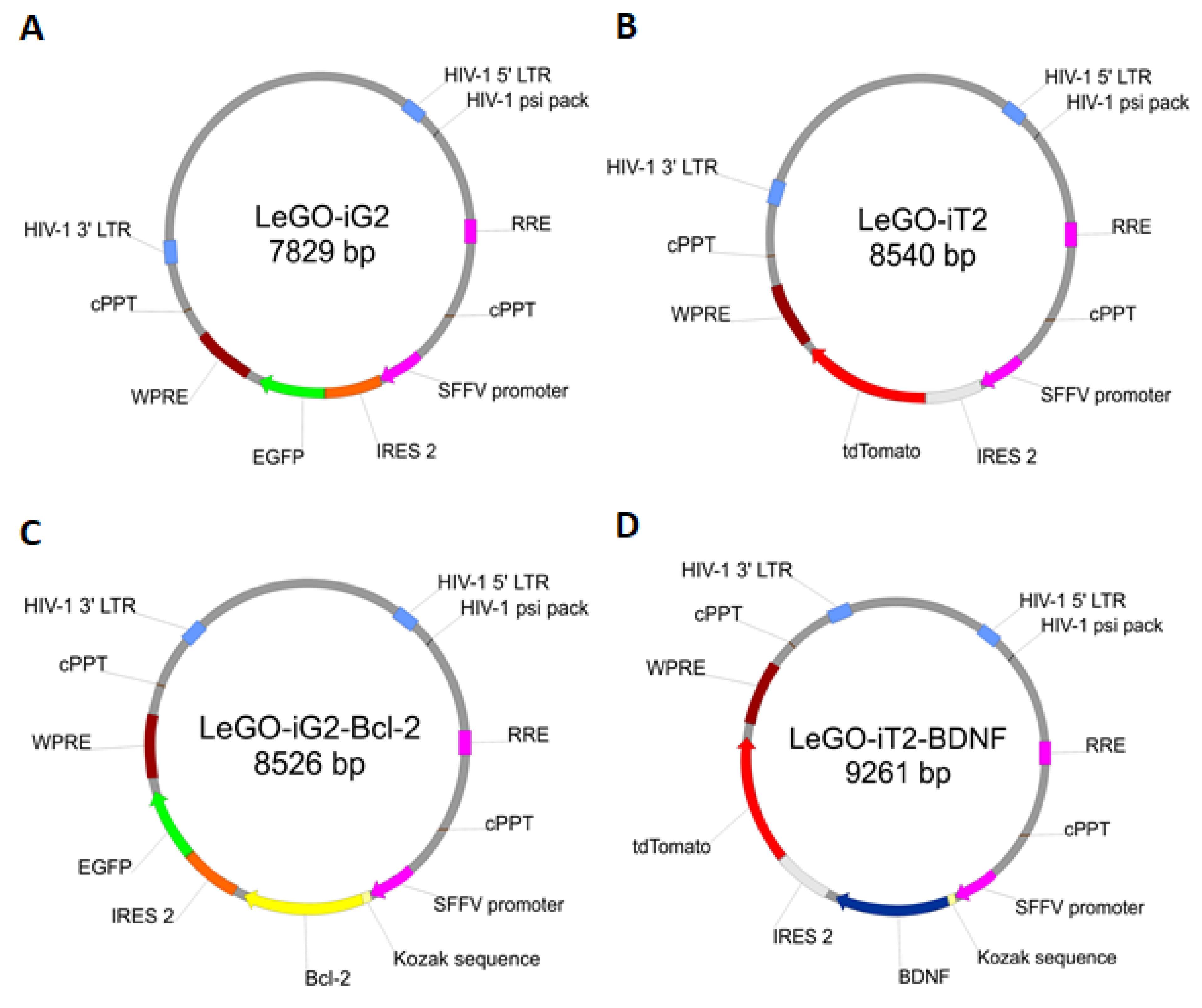
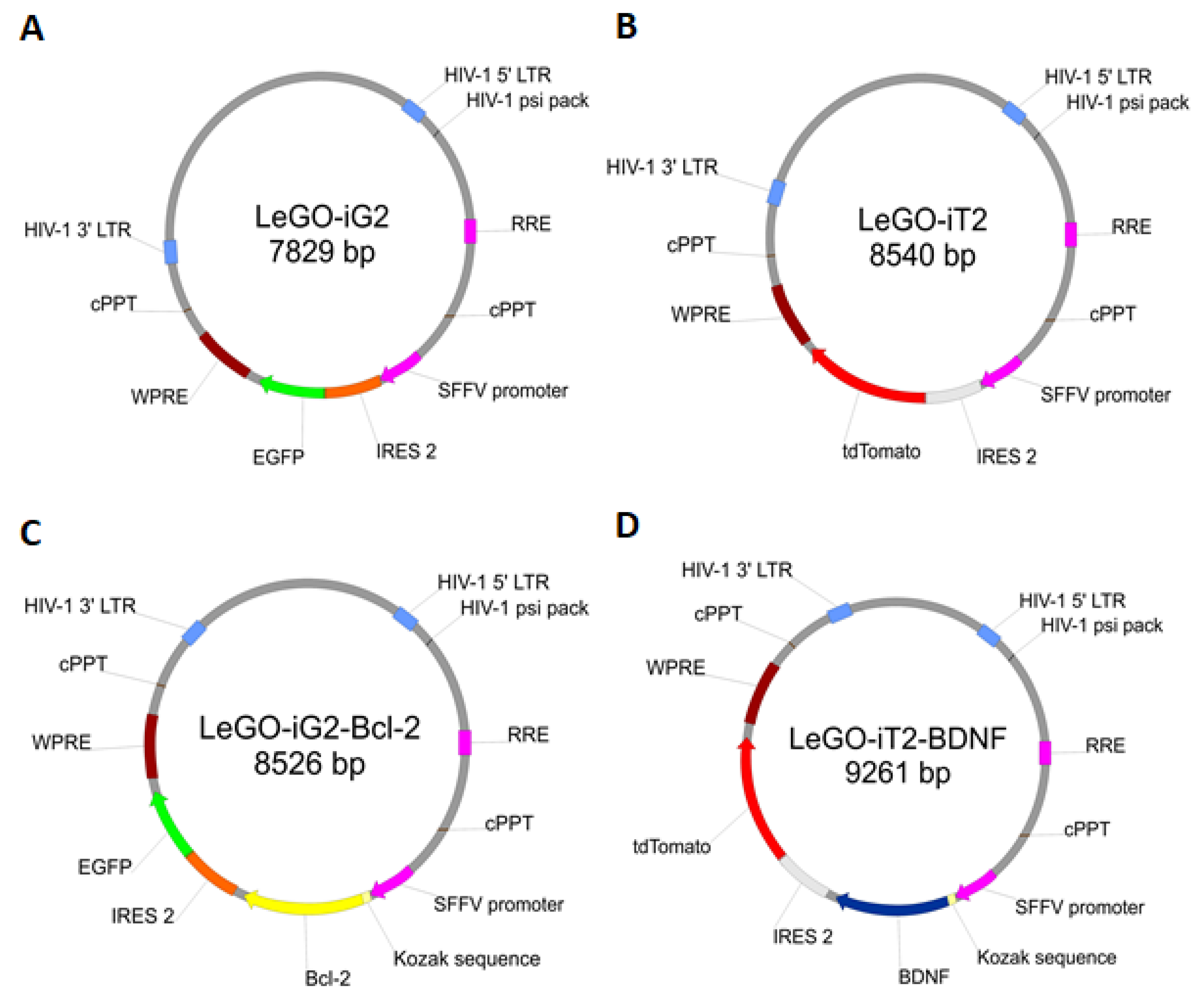
- A control was transduced with the LeGO-iG2 and LeGO-iT2 “empty” vectors (EV) that contained the green or red fluorescent reporter proteins (Figure 2A,B) but no cloned genes that were to be overexpressed. That culture was carried out in parallel with the culture of the cells from the other two groups in an analogous culture medium, and the only additional stimulator was transduction with the empty skeletons of the lentiviruses (Figure 1C).
- (c)
- The study group was transduced with a pair of LeGO-iG2-Bcl-2 and LeGO-iT2-BDNF vectors (full vectors—FV) in order to undergo a synergistic overexpression (Figure 2C,D). That culture was carried out in parallel with the culture of the cells from the other two groups in an analogous culture medium, and the only additional stimulator was BCL2 and BDNF overexpression (Figure 1D). In some cases, the addition of bFGF or resveratrol was investigated, and then the group was indicated as FV + bFGF or FV + resveratrol.
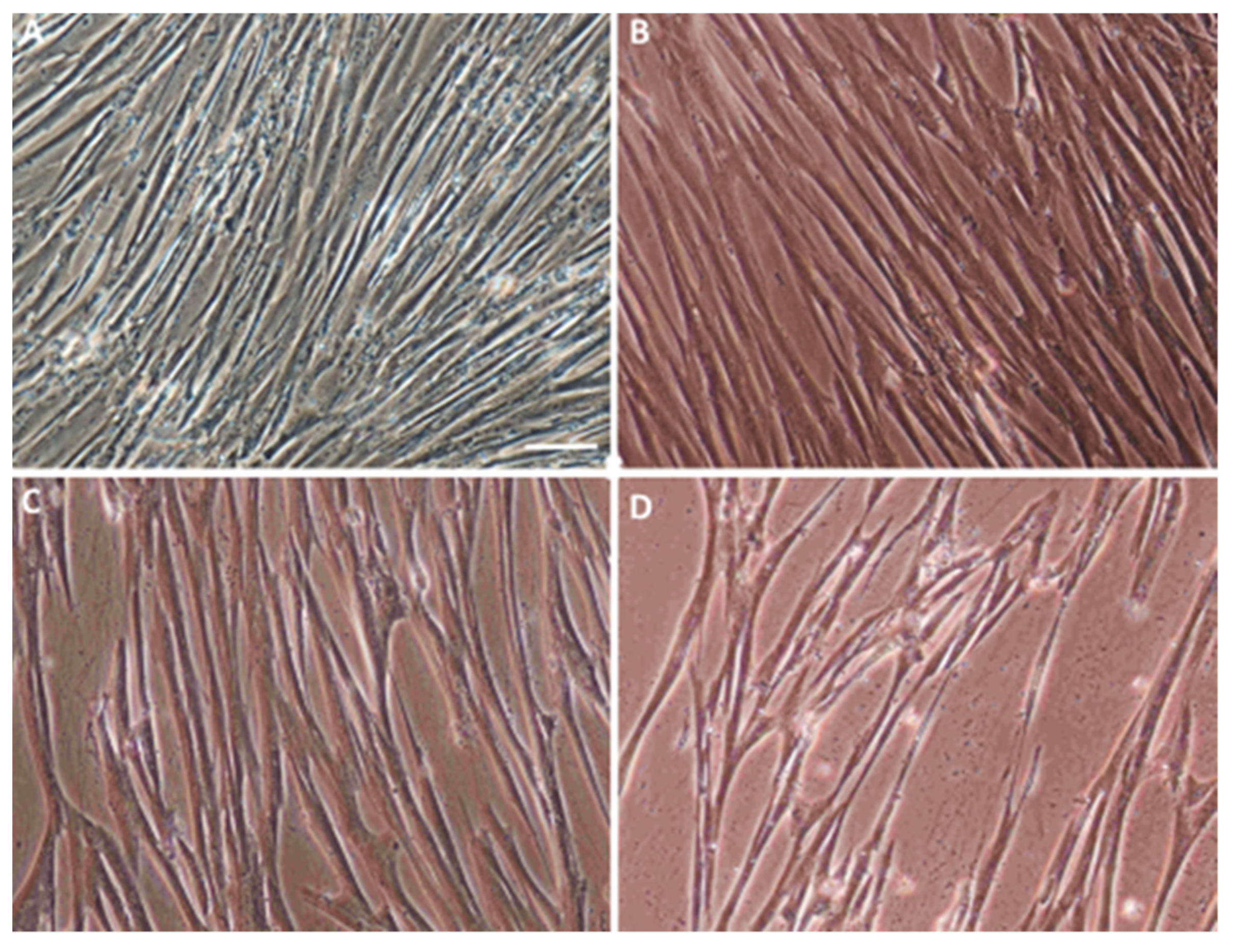
2.1. Characterization of the Isolated WJ-MSC
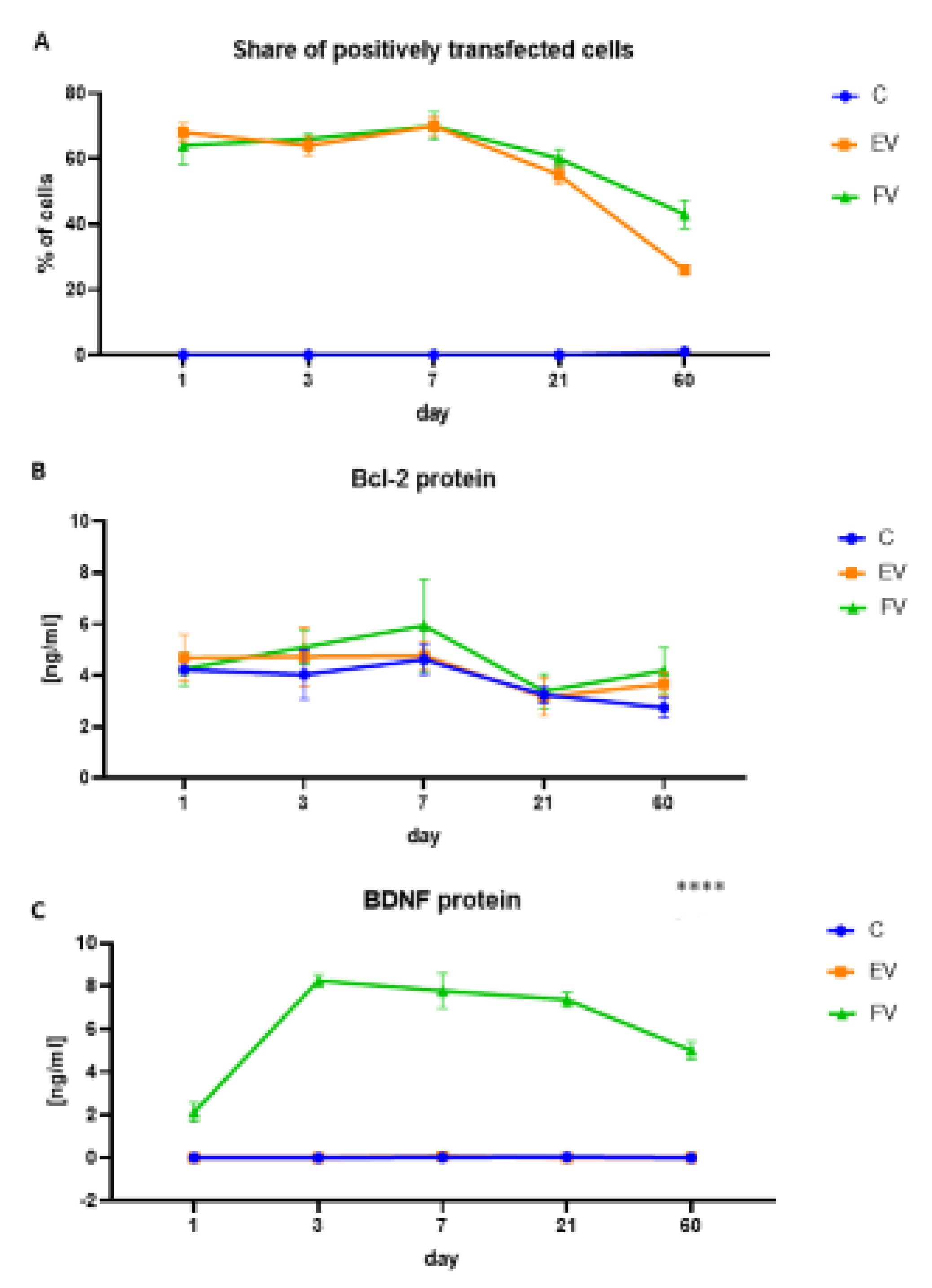
2.2. Efficiency of WJ-MSC Transduction and Overexpressed Protein Levels
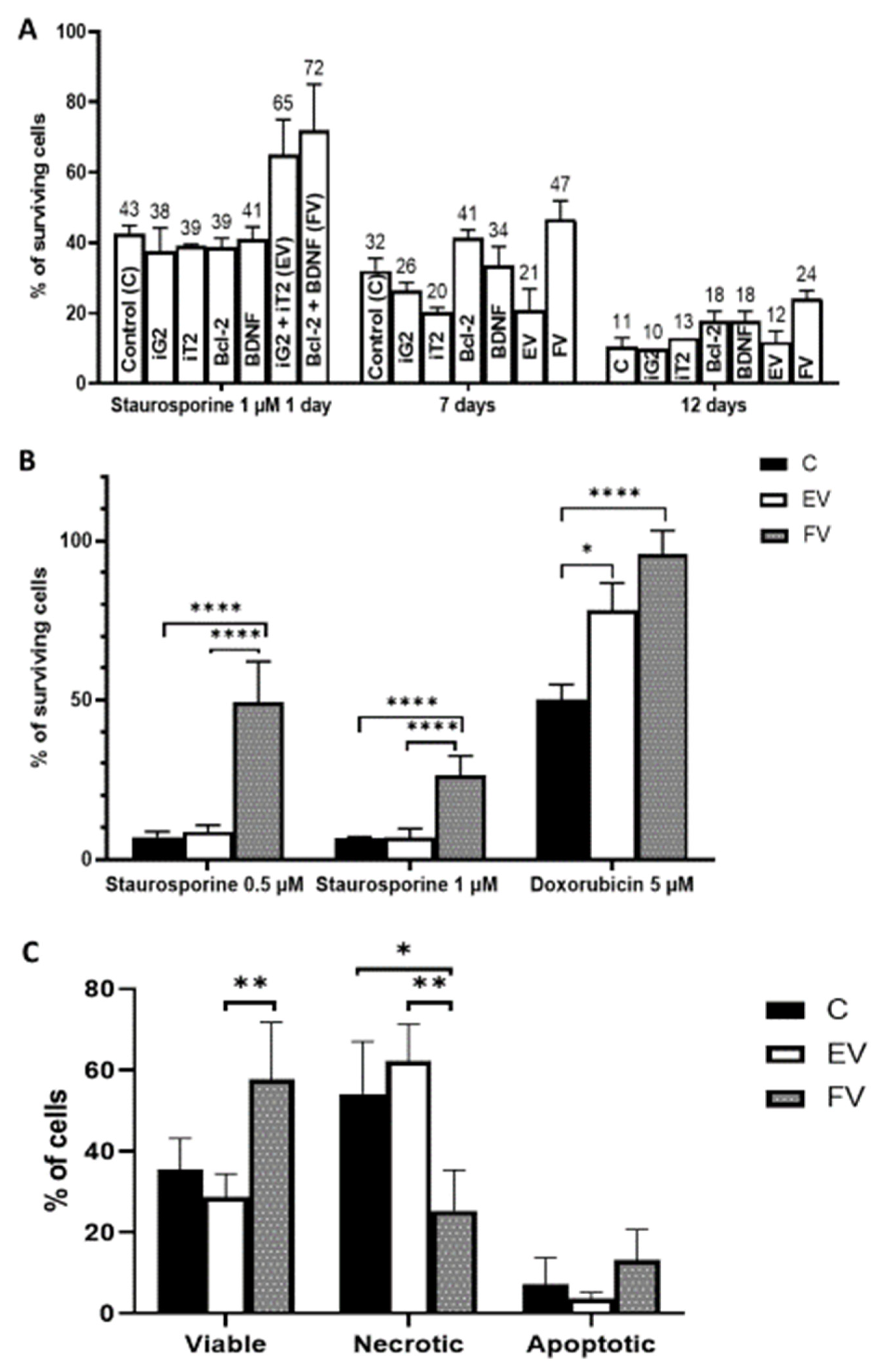
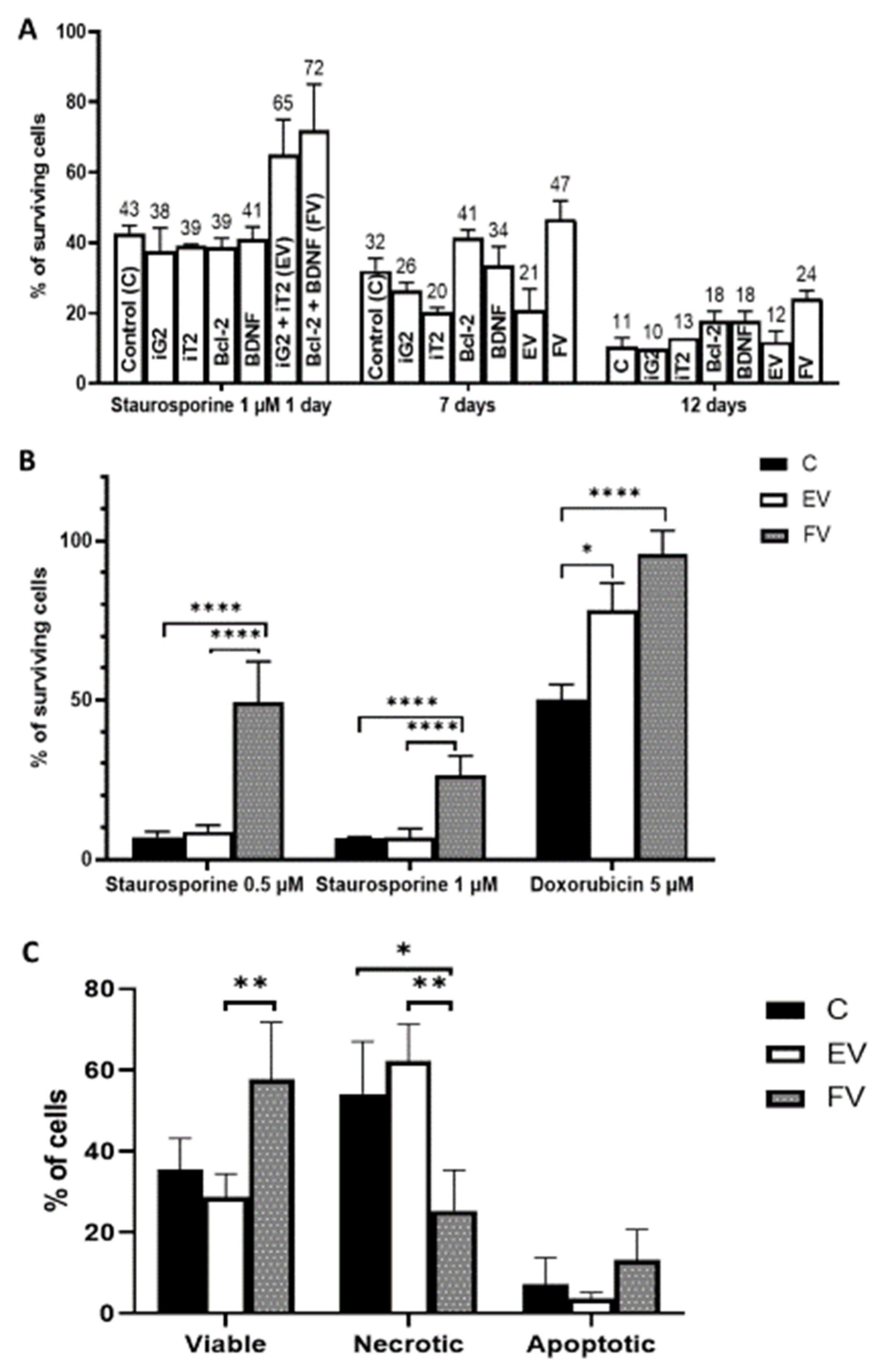
2.3. Cytoprotective Effect Depending on the Duration of BCL2 and BDNF Overexpression
2.4. Analysis of Cell Death
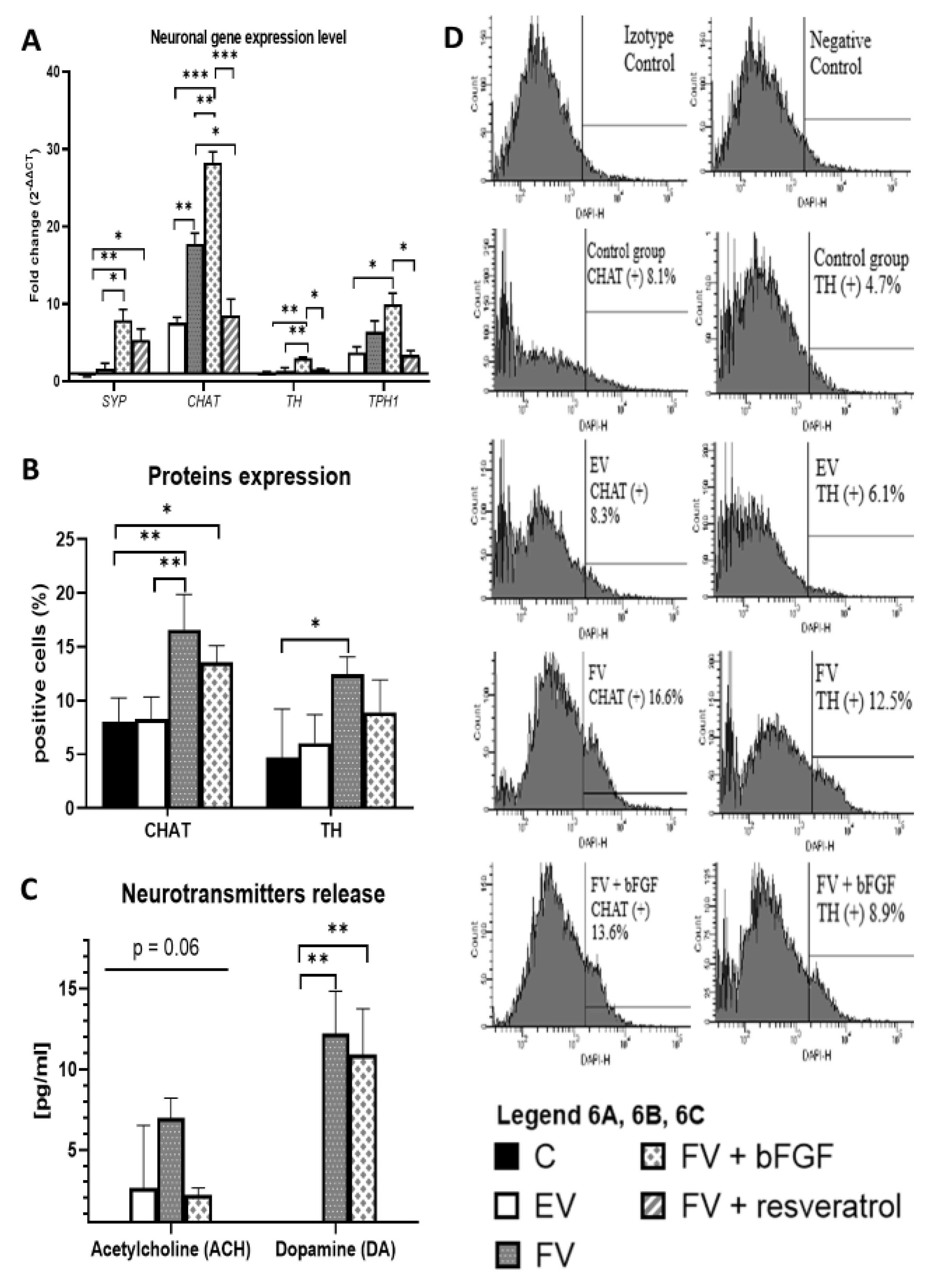
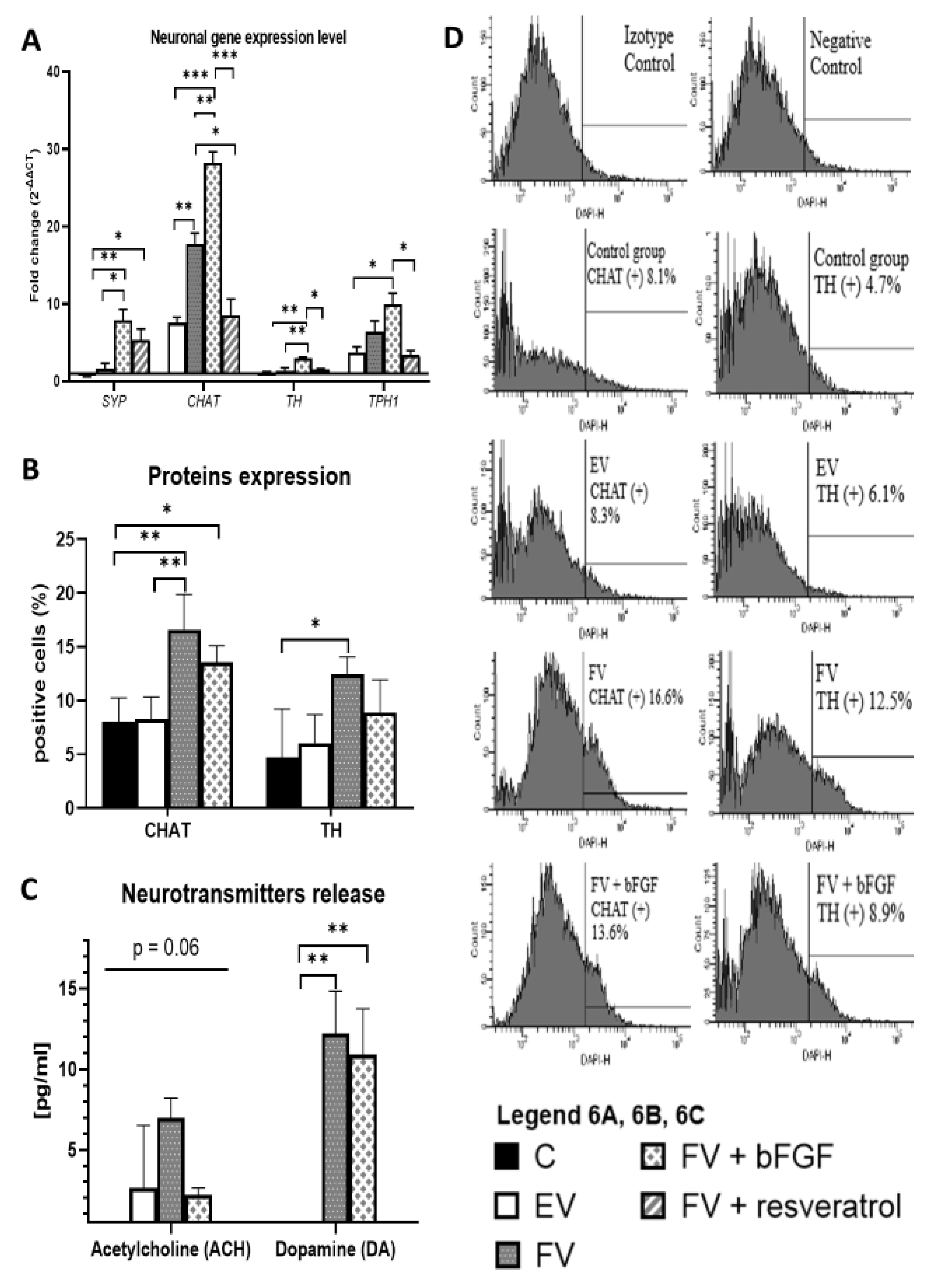
2.5. Neuronal Gene Expression Level
2.6. CHAT and TH Protein Expression
2.7. Release of the Neurotransmitters
3. Discussion
4. Materials and Methods
4.1. Isolating, Culturing, and Characterizing the WJ-MSC
4.2. Lentiviral Vectors
4.3. Flow Cytometry Analysis
4.4. WJ-MSC Transduction, Protein Extractions
4.5. ELISA Analysis of the Bcl-2 and BDNF Proteins
4.6. Cell Viability WST-1 Colorimetric Assay
4.7. Cell Death Analysis
4.8. Neuronal Differentiation
4.9. RT-qPCR
4.10. Evaluating the TH and CHAT Expression of the Proteins
4.11. Neurotransmitters Release Analysis
4.12. Statistical Analysis
5. Conclusions
- increased the survival rate of the transduced cells under toxic conditions
- directed the transduced cells into the neuronal differentiation pathway
- indicated that the cells that are obtained as a result of the process produce neurotransmitters, and thus one can infer their functionality.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Picinich, S.C.; Mishra, P.J.; Mishra, P.J.; Glod, J.; Banerjee, D. The therapeutic potential of mesenchymal stem cells. Expert Opin. Biol. Ther. 2007, 7, 965–973. [Google Scholar] [CrossRef]
- Zipori, D. Mesenchymal stem cells: Harnessing cell plasticity to tissue and organ repair. Blood Cells Mol. Dis. 2004, 33, 211–215. [Google Scholar] [CrossRef]
- Borkowska-Kuczkowska, A.; Sługocka, D.; Świątkowska-Flis, B.; Boruczkowski, D. The use of mesenchymal stem cells for the treatment of progressive retinal diseases: A review. Regen. Med. 2019, 14, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Galderisi, U.; Marino, I.R. From the laboratory bench to the patient’s bedside: An update on clinical trials with mesenchymal stem cells. J. Cell Physiol. 2007, 211, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Isakova, I. Plasticity and therapeutic potential of mesenchymal stem cells in the nervous system. Curr. Pharm. Des. 2005, 11, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.; Riminucci, M.; Gronthos, S.; Robey, P. Bone marrow stromal stem cells: Nature, biology, and potential applications. Stem Cells 2001, 19, 180–192. [Google Scholar] [CrossRef] [Green Version]
- Potian, J.A.; Aviv, H.; Ponzio, N.M.; Harrison, J.S.; Rameshwar, P. Veto-like activity of mesenchymal stem cells: Functional discrimination between cellular responses to alloantigens and recall antigens. J. Immunol. 2003, 171, 3426–3434. [Google Scholar] [CrossRef] [Green Version]
- Krampera, M.; Pasini, A.; Pizzolo, G.; Cosmi, L.; Romagnani, S.; Annunziato, F. Regenerative and immunomodulatory potential of mesenchymal stem cells. Curr. Opin. Pharm. 2006, 6, 435–441. [Google Scholar] [CrossRef]
- Greco, S.J.; Zhou, C.; Ye, J.H.; Rameshwar, P. An interdisciplinary approach and characterization of neuronal cells transdifferentiated from human mesenchymal stem cells. Stem Cells Dev. 2007, 16, 811–826. [Google Scholar] [CrossRef]
- Sato, Y.; Araki, H.; Kato, J.; Nakamura, K.; Kawano, Y.; Kobune, M.; Sato, T.; Miyanishi, K.; Takayama, T.; Takahashi, M.; et al. Human mesenchymal stem cells xenografted directly to rat liver are differentiated into human hepatocytes without fusion. Blood 2005, 106, 756–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzaska, K.A.; Rameshwar, P. Dopaminergic neuronal differentiation protocol for human mesenchymal stem cells. Methods Mol. Biol. 2011, 698, 295–303. [Google Scholar]
- Singh, M.; Kakkar, A.; Sharma, R.; Kharbanda, O.P.; Monga, N.; Kumar, M.; Chowdhary, S.; Airan, B.; Mohanty, S. Synergistic effect of BDNF and FGF2 in efficient generation of functional dopaminergic neurons from human mesenchymal stem cells. Sci. Rep. 2017, 4, 10378. [Google Scholar] [CrossRef] [Green Version]
- Nandy, S.B.; Mohanty, S.; Singh, M.; Behari, M.; Airan, B. Fibroblast Growth Factor-2 alone as an efficient inducer for differentiation of human bone marrow mesenchymal stem cells into dopaminergic neurons. J. Biomed. Sci. 2014, 24, 21–83. [Google Scholar] [CrossRef] [Green Version]
- Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Direct transdifferentiation of human Wharton’s jelly mesenchymal stromal cells into cholinergic-like neurons. J. Neurosci. Methods 2019, 312, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Feng, Z.; Xu, J.; Jiang, Z.; Feng, M. Brain-derived neurotrophic factor modified human umbilical cord mesenchymal stem cells-derived cholinergic-like neurons improve spatial learning and memory ability in Alzheimer’s disease rats. Brain Res. 2019, 1710, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Sortwell, C.E.; Pitzer, M.R.; Collier, T.J. Time course of apoptotic cell death within mesencephalic cell suspension grafts: Implications for improving grafted dopamine neuron survival. Exp. Neurol. 2000, 165, 268–277. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Kluck, R.M.; Bossy-Wetzel, E.; Green, D.; Newmeyer, D.D. The release of cytochrome c from mitochondria: A primary site for Bcl-2 regulation of apoptosis. Science 1997, 275, 1132–1136. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ma, N.; Ong, L.-L.; Nesselmann, C.; Klopsch, C.; Ladilov, Y.; Furlani, D.; Piechaczek, C.; Moebius, J.M.; Lützow, K.; et al. Bcl-2 Engineered MSCs Inhibited Apoptosis and Improved Heart Function. Stem Cells 2007, 25, 2118–2127. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Law, H.K.; Lau, Y.L.; Chan, G.C. Differential damage and recovery of human mesenchymal stem cells after exposure to chemotherapeutic agents. Br. J. Haematol. 2004, 127, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Mueller, L.P.; Luetzkendorf, J.; Mueller, T.; Reichelt, K.; Simon, H.; Schmoll, H.-J. Presence of Mesenchymal Stem Cells in Human Bone Marrow After Exposure to Chemotherapy: Evidence of Resistance to Apoptosis Induction. Stem Cells 2006, 24, 2753–2765. [Google Scholar] [CrossRef] [PubMed]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Borkowska, P.; Zielińska, A.; Paul-Samojedny, M.; Stojko, R.; Kowalski, J. Evaluation of reference genes for quantitative real-time PCR in Wharton’s Jelly-derived mesenchymal stem cells after lentiviral transduction and differentiation. Mol. Biol. Rep. 2019, 47, 1107–1115. [Google Scholar] [CrossRef]
- Martín, F.; Tristán-Manzano, M.; Maldonado-Pérez, N.; Sánchez-Hernández, S.; Benabdellah, K.; Cobo, M. Stable genetic modification of mesenchymal stromal cells using lentiviral vectors. Methods Mol. Biol. 2019, 1937, 267–280. [Google Scholar]
- Yin, X.; Xu, J.-C.; Cho, G.-S.; Kwon, C.; Dawson, T.M.; Dawson, V.L. Neurons derived from human induced pluripotent stem cells integrate into rat brain circuits and maintain both excitatory and inhibitory synaptic activities. Eneuro 2019, 22, 6. [Google Scholar] [CrossRef] [Green Version]
- Simão, D.; Silva, M.M.; Terrasso, A.P.; Arez, F.; Sousa, M.F.Q.; Mehrjardi, N.Z.; Šarić, T.; Gomes-Alves, P.; Raimundo, N.; Alves, P.M.; et al. Recapitulation of human neural microenvironment signatures in iPSC-derived NPC 3D differentiation. Stem Cell Rep. 2018, 14, 552–564. [Google Scholar] [CrossRef] [Green Version]
- Ni, X.; Ou, C.; Guo, J.; Liu, B.; Zhang, J.; Wu, Z.; Li, H.; Chen, M. Lentiviral vector-mediated co-overexpression of VEGF and Bcl-2 improves mesenchymal stem cell survival and enhances paracrine effects in vitro. Int. J. Mol. Med. 2017, 40, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Yuste, V.J.; Sánchez-López, I.; Solé, C.; Encinas, M.; Bayascas, J.R.; Boix, J.; Comella, J.X. The prevention of the staurosporine-induced apoptosis by Bcl-X(L), but not by Bcl-2 or caspase inhibitors, allows the extensive differentiation of human neuroblastoma cells. J. Neurochem. 2002, 80, 126–139. [Google Scholar] [CrossRef]
- Oliver, L.; Hue, E.; Rossignol, J.; Bougras, G.; Hulin, P.; Naveilhan, P.; Heymann, D.; Lescaudron, L.; Vallette, F.M. Distinct roles of Bcl-2 and Bcl-Xl in the apoptosis of human bone marrow mesenchymal stem cells during differentiation. PLoS ONE 2011, 6, e19820. [Google Scholar] [CrossRef]
- Lejkowska, R.; Kawa, M.P.; Pius-Sadowska, E.; Rogińska, D.; Łuczkowska, K.; Machaliński, B.; Machalińska, A. Preclinical evaluation of long-term neuroprotective effects of BDNF-engineered mesenchymal stromal cells as intravitreal therapy for chronic retinal degeneration in Rd6 mutant mice. Int. J. Mol. Sci. 2019, 12, 777. [Google Scholar] [CrossRef] [Green Version]
- Scheper, V.; Schwieger, J.; Hamm, A.; Lenarz, T.; Hoffmann, A. BDNF-overexpressing human mesenchymal stem cells mediate increased neuronal protection in vitro. J. Neurosci. Res. 2019, 97, 1414–1429. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Qiu, X.C.; Ma, Y.H.; Duan, J.J.; Chen, Y.F.; Gu, H.Y.; Wang, J.M.; Ling, E.A.; Wu, J.L.; Wu, W.; et al. Integration of donor mesenchymal stem cell-derived neuron-like cells into host neural network after rat spinal cord transection. Biomaterials 2015, 53, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Elias, G.; Marcus, A.J.; Coyne, T.M.; Woodbury, D.; Black, I.B. Adult Bone Marrow Stromal Cells in the Embryonic Brain: Engraftment, Migration, Differentiation, and Long-Term Survival. J. Neurosci. 2004, 24, 4585–4595. [Google Scholar] [CrossRef]
- Chou, F.C.; Sytwu, H.K. Overexpression of thioredoxin in islets transduced by a lentiviral vector prolongs graft survival in autoimmune diabetic NOD mice. J. Biomed. Sci. 2009, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathina, S.; Srinivas, N.; Das, U.N. BDNF protects pancreatic β cells (RIN5F) against cytotoxic action of alloxan, streptozotocin, doxorubicin and benzo(a)pyrene in vitro. Metabolism 2016, 65, 667–684. [Google Scholar] [CrossRef]
- Takeda, K.; Kermani, P.; Anastasia, A.; Obinata, Y.; Hempstead, B.L.; Kurihara, H. BDNF protects human vascular endothelial cells from TNFα-induced apoptosis. Biochem. Cell Biol. 2013, 91, 341–349. [Google Scholar] [CrossRef]
- Lipsky, R.H.; Marini, A.M. Brain-derived neurotrophic factor in neuronal survival and behavior-related plasticity. Ann. N. Y. Acad. Sci. 2007, 1122, 130–143. [Google Scholar] [CrossRef]
- Chang, W.; Song, B.-W.; Moon, J.-Y.; Cha, M.-J.; Ham, O.; Lee, S.-Y.; Choi, E.; Choi, E.; Hwang, K.-C. Anti-death strategies against oxidative stress in grafted mesenchymal stem cells. Histol. Histopathol. 2013, 28, 1529–1536. [Google Scholar]
- Peruzzaro, S.T.; Andrews, M.M.M.; Al-Gharaibeh, A.; Pupiec, O.; Resk, M.; Story, D.; Maiti, P.; Rossignol, J.; Dunbar, G.L. Transplantation of mesenchymal stem cells genetically engineered to overexpress interleukin-10 promotes alternative inflammatory response in rat model of traumatic brain injury. J. Neuroinflamm. 2019, 16, 2. [Google Scholar] [CrossRef]
- Jang, S.; Kang, Y.H.; Ullah, I.; Shivakumar, S.B.; Rho, G.J.; Cho, Y.C.; Sung, I.Y.; Park, B.W. Cholinergic nerve differentiation of mesenchymal stem cells derived from long-term cryopreserved human dental pulp in vitro and analysis of their motor nerve regeneration potential in vivo. Int. J. Mol. Sci. 2018, 19, 2434. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Sun, C.; Su, L.; Zhao, J.; Zhang, S.; Miao, J. Phosphatidylcholine-specific phospholipase C/heat shock protein 70 (Hsp70)/transcription factor B-cell translocation gene 2 signaling in rat bone marrow stromal cell differentiation to cholinergic neuron-like cells. Int. J. Biochem. Cell Biol. 2012, 44, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Guo, S.; Liao, H.; Yu, P.; Wang, L.; Song, X.; Chen, J.Q. Resveratrol enhances neurite outgrowth and synaptogenesis via sonic hedgehog signaling following oxygen-glucose deprivation/reoxygenation injury. Cell Physiol. Biochem. 2017, 43, 852–869. [Google Scholar] [CrossRef] [Green Version]
- Kopach, O. Monitoring maturation of neural stem cell grafts within a host microenvironment. World J. Stem Cells 2019, 11, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Kopach, O.; Pivneva, T. Cell-based therapies for neural replacement strategies in stroke-related neurodegeneration: Neurophysiological insights into stem progenitor cell neurogenesis within a host environment. Neural. Regen. Res. 2018, 13, 1350–1351. [Google Scholar] [CrossRef]
- Morgan, P.J.; Liedmann, A.; Hübner, R.; Hovakimyan, M.; Rolfs, A.; Frech, M.J. Human neural progenitor cells show functional neuronal differentiation and regional preference after engraftment onto hippocampal slice cultures. Stem Cells Dev. 2012, 21, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- El Sayed Marei, H.; El-Gamal, A.; Althani, A.; Afifi, N.; Abd-Elmaksoud, A.; Farag, A.; Cenciarelli, C.; Thomas, C.; Anwarul, H. Cholinergic and dopaminergic neuronal differentiation of human adipose tissue derived mesenchymal stem cells. J. Cell Physiol. 2018, 233, 936–945. [Google Scholar] [CrossRef]
- Alizadeh, R.; Bagher, Z.; Kamrava, S.K.; Falah, M.; Hamidabadi, H.G.; Boroujeni, M.E.; Mohammadi, F.; Khodaverdi, S.; Sadeghi, A.Z.; Olya, A.; et al. Differentiation of human mesenchymal stem cells (MSC) to dopaminergic neurons: A comparison between Wharton’s Jelly and olfactory mucosa as sources of MSCs. J. Chem. Neuroanat. 2019, 96, 126–133. [Google Scholar] [CrossRef]
- Weber, K.; Bartsch, U.; Stocking, C.; Fehse, B. A multicolor panel of novel lentiviral “gene ontology” (LeGO) vectors for functional gene analysis. Mol. Ther. 2008, 16, 698–706. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figure 4A Left | Figure 4A Middle | Figure 4A Right | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Tukey’s Multiple Comparisons Test | Significant? | Summary | Adjusted p Value | Significant? | Summary | Adjusted p Value | Significant? | Summary | Adjusted p Value |
| Control (C) (1d) vs. iG2 (1d) | No | ns | 0.9834 | No | ns | 0.9847 | No | ns | >0.9999 |
| Control (C) (1d) vs. iT2 (1d) | No | ns | 0.9969 | No | ns | 0.3946 | No | ns | 0.7552 |
| Control (C) (1d) vs. Bcl-2 (1d) | No | ns | 0.995 | No | ns | 0.3541 | No | ns | 0.1392 |
| Control (C) (1d) vs. BDNF (1d) | No | ns | >0.9999 | No | ns | 0.9973 | No | ns | 0.1553 |
| Control (C) (1d) vs. iG2+iT2 (EV) (1d) | Yes | * | 0.0309 | No | ns | 0.6398 | No | ns | 0.9997 |
| Control (C) (1d) vs. Bcl-2+BDNF (FV) (1d) | Yes | ** | 0.0036 | Yes | ** | 0.0049 | Yes | **** | <0.0001 |
| iG2 (1d) vs. iT2 (1d) | No | ns | >0.9999 | No | ns | 0.8561 | No | ns | 0.9566 |
| iG2 (1d) vs. Bcl-2 (1d) | No | ns | >0.9999 | Yes | * | 0.0488 | No | ns | 0.5076 |
| iG2 (1d) vs. BDNF (1d) | No | ns | 0.9984 | No | ns | 0.7783 | No | ns | 0.5348 |
| iG2 (1d) vs. iG2+iT2 (EV) (1d) | Yes | * | 0.0121 | No | ns | 0.976 | No | ns | >0.9999 |
| iG2 (1d) vs. Bcl-2+BDNF (FV) (1d) | Yes | ** | 0.0017 | Yes | *** | 0.0002 | Yes | **** | <0.0001 |
| iT2 (1d) vs. Bcl-2 (1d) | No | ns | >0.9999 | Yes | *** | 0.0006 | No | ns | 0.9783 |
| iT2 (1d) vs. BDNF (1d) | No | ns | >0.9999 | No | ns | 0.0926 | No | ns | 0.9832 |
| iT2 (1d) vs. iG2+iT2 (EV) (1d) | Yes | * | 0.0178 | No | ns | 0.9992 | No | ns | 0.8891 |
| iT2 (1d) vs. Bcl-2+BDNF (FV) (1d) | Yes | ** | 0.0024 | Yes | **** | <0.0001 | Yes | ** | 0.0054 |
| Bcl-2 (1d) vs. BDNF (1d) | No | ns | 0.9998 | No | ns | 0.6515 | No | ns | >0.9999 |
| Bcl-2 (1d) vs. iG2+iT2 (EV) (1d) | Yes | * | 0.0161 | Yes | ** | 0.002 | No | ns | 0.2446 |
| Bcl-2 (1d) vs. Bcl-2+BDNF (FV) (1d) | Yes | ** | 0.0022 | No | ns | 0.406 | No | ns | 0.0602 |
| BDNF (1d) vs. iG2+iT2 (EV) (1d) | Yes | * | 0.0314 | No | ns | 0.2117 | No | ns | 0.2688 |
| BDNF (1d) vs. Bcl-2+BDNF (FV) (1d) | Yes | ** | 0.0042 | Yes | * | 0.0135 | No | ns | 0.0528 |
| iG2+iT2 (EV) (1d) vs. Bcl-2+BDNF (FV) (1d) | No | ns | 0.9307 | Yes | **** | <0.0001 | Yes | **** | <0.0001 |
| Markers | Gene Name | Primer Pair ID | Forward Primer | Reverse Primer | |
|---|---|---|---|---|---|
| RPS17 | endogenous control | Ribosomal Protein S17 | H_RPS17_1 | CCATTATCCCCAGCAAAAAG | GAGACCTCAGGAACATAATTG |
| TH | dopaminergic neurons | Tyrosine Hydroxylase | H_TH_1 | CAAAATCCACCATCTAGAGAC | CTGACACTTTTCTTGGGAAC |
| CHAT | cholinergic neurons | Choline O-Acetyltransferase | H_CHAT_1 | TCATTTCTTTGTCTTGGATG | TGGAAGCCATTTTGACTATC |
| TPH1 | serotoninergic neurons | Tryptophan Hydroxylase 1 | H_TPH1_1 | AAAGAGCGTACAGGTTTTTC | GTCTCACATATTGAGTGCAG |
| SYP | synaptogenesis marker | Synaptophysin | H_SYP_1 | CCCTTCGGTATTGTTCAAAG | TTTGACTAGGTGGTTAAGGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borkowska, P.; Zielinska, A.; Paul-Samojedny, M.; Stojko, R.; Kowalski, J. Synergistic Effect of the Long-Term Overexpression of Bcl-2 and BDNF Lentiviral in Cell Protecting against Death and Generating TH Positive and CHAT Positive Cells from MSC. Int. J. Mol. Sci. 2021, 22, 7086. https://doi.org/10.3390/ijms22137086
Borkowska P, Zielinska A, Paul-Samojedny M, Stojko R, Kowalski J. Synergistic Effect of the Long-Term Overexpression of Bcl-2 and BDNF Lentiviral in Cell Protecting against Death and Generating TH Positive and CHAT Positive Cells from MSC. International Journal of Molecular Sciences. 2021; 22(13):7086. https://doi.org/10.3390/ijms22137086
Chicago/Turabian StyleBorkowska, Paulina, Aleksandra Zielinska, Monika Paul-Samojedny, Rafał Stojko, and Jan Kowalski. 2021. "Synergistic Effect of the Long-Term Overexpression of Bcl-2 and BDNF Lentiviral in Cell Protecting against Death and Generating TH Positive and CHAT Positive Cells from MSC" International Journal of Molecular Sciences 22, no. 13: 7086. https://doi.org/10.3390/ijms22137086
APA StyleBorkowska, P., Zielinska, A., Paul-Samojedny, M., Stojko, R., & Kowalski, J. (2021). Synergistic Effect of the Long-Term Overexpression of Bcl-2 and BDNF Lentiviral in Cell Protecting against Death and Generating TH Positive and CHAT Positive Cells from MSC. International Journal of Molecular Sciences, 22(13), 7086. https://doi.org/10.3390/ijms22137086