
Morphological, Molecular and Genomic Characterization of Two Inter-Subspecific Hybrids between Olive Cultivars and Olive Subspecies


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Characterization
2.3. DNA Extraction and Microsatellite Amplification
2.4. Parentage Analysis Using SSR Markers
2.5. Genomic DNA Isolation and DNA Sequencing of SCNL
2.6. Data Analysis
3. Results and Discussions
3.1. Morphological Characterization of Two Inter-Subspecific Progenies and Their Parents
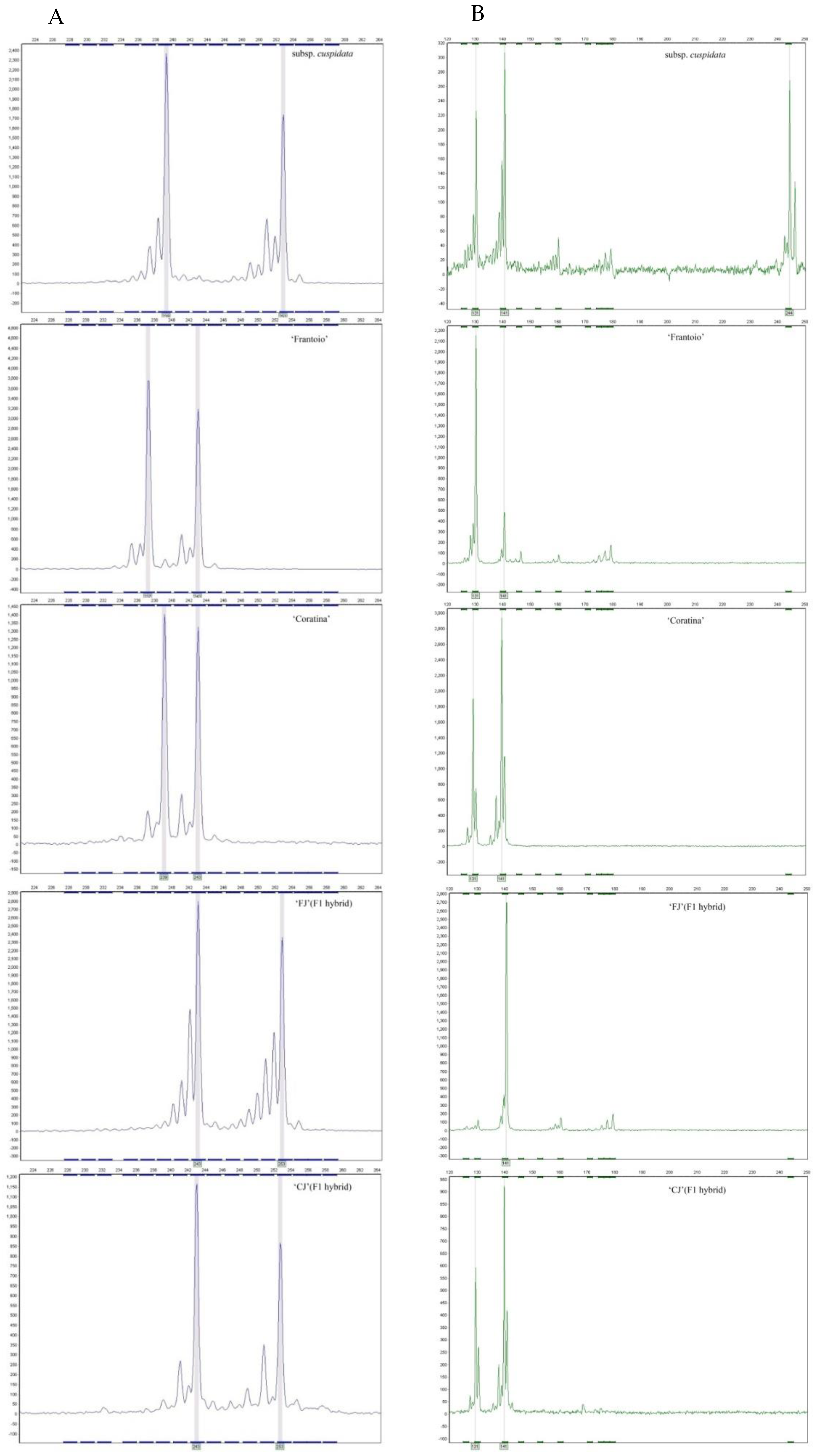
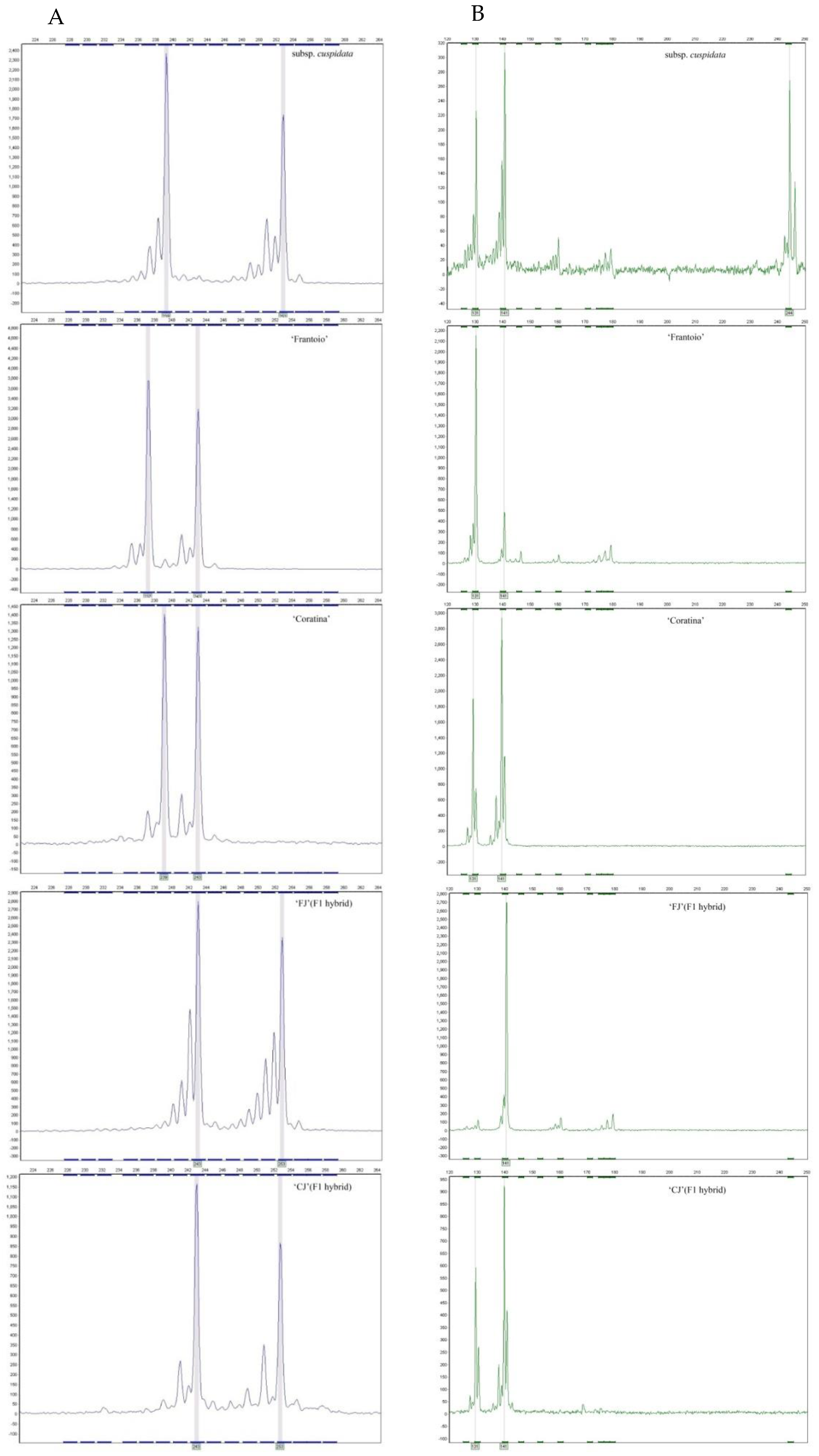
3.2. Parentage Analysis and Progeny Discrimination by SSR
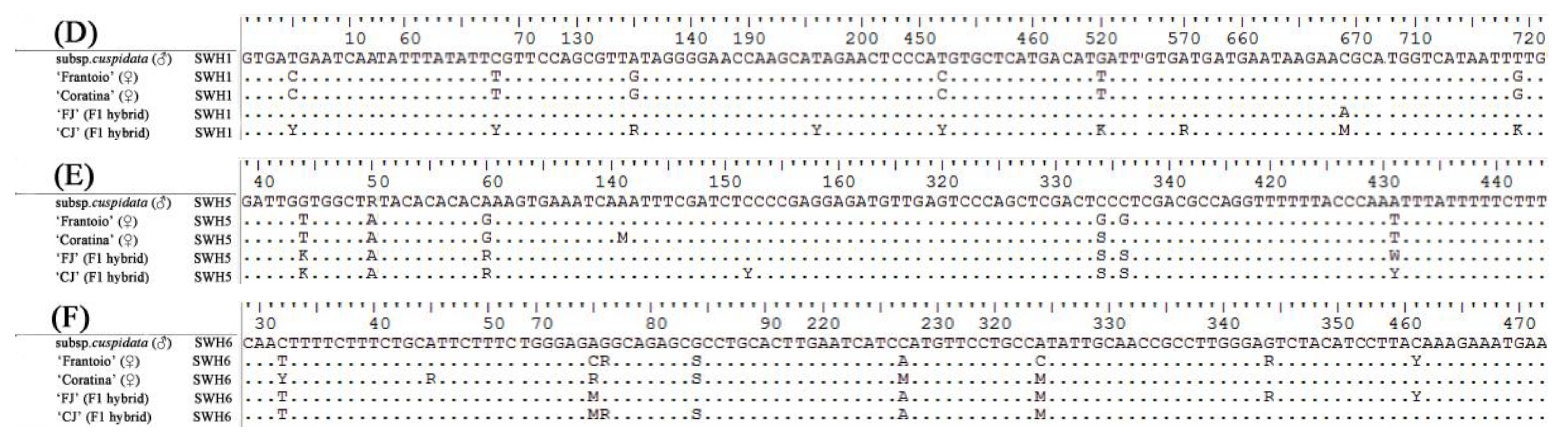
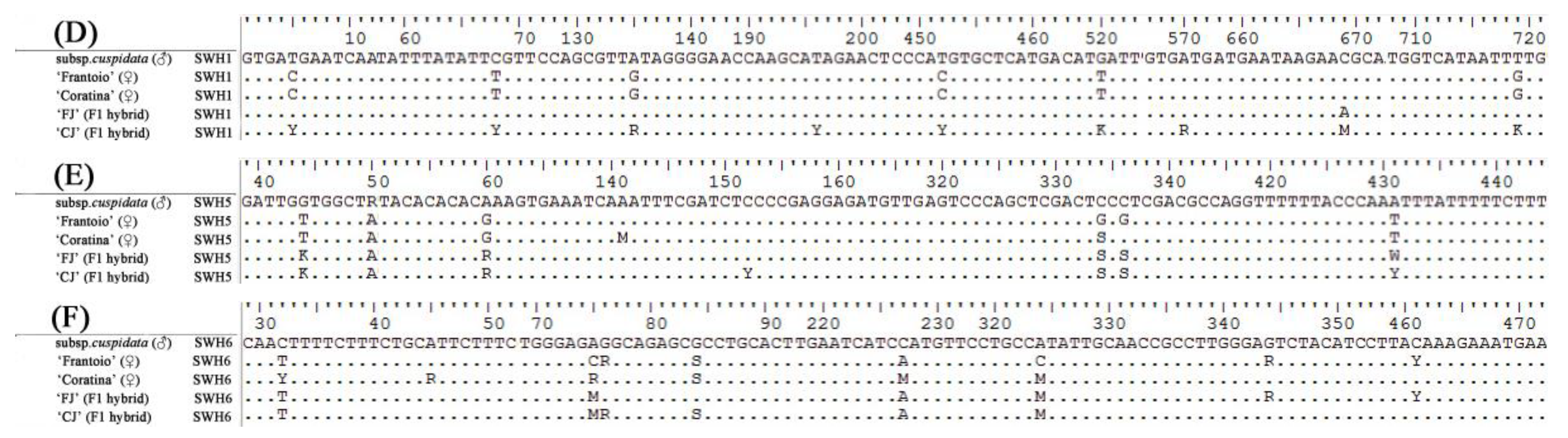
3.3. DNA Sequence Alignment of Progenies and Their Parents by SCNL
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Haouane, H.; El Bakkali, A.; Moukhli, A.; Tollon-Cordet, C.; Santoni, S.; Oukabli, A.; El Modafar, C.; Khadari, B. Genetic structure and core collection of the World Olive Germplasm Bank of Marrakech: Towards the optimised management and use of Mediterranean olive genetic resources. Genetica 2011, 139, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Belaj, A.; Dominguez-García, M.D.C.; Atienza, S.G.; Urdíroz, N.M.; De La Rosa, R.; Satovic, Z.; Martín, A.; Kilian, A.; Trujillo, I.; Valpuesta, V.; et al. Developing core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Díez, C.M.; Imperato, A.; Rallo, L.; Barranco, D.; Trujillo, I. World-wide core collection of olive cultivars based on simple sequence repeatand morphological markers. Crop Sci. 2012, 52, 211–221. [Google Scholar] [CrossRef]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; de Caraffa, V.B.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From late quaternary diversification of mediterranean lineages to primary domestication in the northern levant. Proc. R. Soc. Lond. Ser. B 2013, 280, 20122833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Mariotti, R.; Bagnoli, F.; Costantini, L.; Cultrera, N.G.M.; Arzani, K.; Pandolfi, S.; Vendramin, G.G.; Torkzaban, B.; Hosseini-Mazinani, M.; et al. The eastern part of the Fertile Crescent concealed an unexpected route of olive (Olea europaea L.) differentiation. Ann. Bot. 2017, 119, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Besnard, G.; Terral, J.-F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S.; Ruiz, P.F.M. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Kaniewski, D.; van Campo, E.; Boiy, T.; Terral, J.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeo botanical, historical and molecular evidences from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rallo, L.; Barranco, D.; Díez, C.M.; Rallo, P.; Suárez, M.P.; Trapero, C.; Pliego-Alfaro, F. Strategies for olive (Olea europaea L.) breeding: Cultivated genetic resources and crossbreeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Lavee, S.; Avidan, B.; Ben-Ari, G. Trends in breeding new olive varieties in Israel for quality and economic management. Agric. Sci. 2014, 5, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Hannachi, H.; Sommerlatte, H.; Breton, C.; Msallem, M.; El Gazzah, M.; Ben El Hadj, S.; Bervillé, A. Oleaster (var. sylvestris) and subsp. cuspidata are suitable genetic resources for improvement of the olive (Olea europaea subsp. europaea var. europaea). Genet. Resour. Crop Evol. 2008, 56, 393–403. [Google Scholar] [CrossRef]
- Klepo, T.; De La Rosa, R.; Satovic, Z.; León, L.; Belaj, A. Utility of wild germplasm in olive breeding. Sci. Hortic. 2013, 152, 92–101. [Google Scholar] [CrossRef]
- De La Rosa, R.; Klepo, T.; Arias-Calderón, R.; Toumi, A.; Domínguez-García, M.; Leon, L.; Belaj, A.; Mariotti, R.; Baldoni, L.; Satovic, Z.; et al. Current status of conservation, evaluation and usefulness of wild olive germplasm. Acta Hortic. 2014, 1057, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Belaj, A.; León, L.; Satovic, Z.; de la Rosa, R. Variability of wild olives (Olea europaea subsp. europaea var. sylvestris) analyzed by agromorphological traits and SSR markers. Sci. Hortic. 2011, 129, 561–569. [Google Scholar] [CrossRef]
- Klepo, T.; Toumi, A.; De La Rosa, R.; León, L.; Belaj, A. Agronomic evaluation of seedlings from crosses between the main Spanish olive cultivar ‘Picual’ and two wild olive trees. J. Hortic. Sci. Biotechnol. 2014, 89, 508–512. [Google Scholar] [CrossRef]
- León, L.; De La Rosa, R.; Velasco, L.; Belaj, A. Using wild olives in breeding programs: Implications on oil quality composition. Front. Plant Sci. 2018, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- León, L.; Díaz-Rueda, P.; Belaj, A.; De la Rosa, R.; Carrascosa, C.; Colmenero-Flores, J. Evaluation of early vigor traits in wild olive germplasm. Sci. Hortic. 2020, 264, 109157. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Rueda, P.; Franco-Navarro, J.D.; Messora, R.; Espartero, J.; Rivero-Núñez, C.M.; Aleza, P.; Capote, N.; Cantos, M.; García-Fernández, J.L.; De Cires, A.; et al. SILVOLIVE, a germplasm collection of wild subspecies with high genetic variability as a source of rootstocks and resistance genes for olive breeding. Front. Plant Sci. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, M.E.; Ceccarelli, M.; Pupilli, F.; Sarri, V.; Mencuccini, M. Obtainment of inter-subspecific hybrids in olive (Olea europaea L.). Euphytica 2014, 201, 307–319. [Google Scholar] [CrossRef]
- Lavee, S. Evaluation of the need and present potential of olive breeding indicating the nature of the available genetic resources involved. Sci. Hortic. 2013, 161, 333–339. [Google Scholar] [CrossRef]
- Lavee, S.; Zohary, D. The potential of genetic diversity and the effect of geographically isolated resources in olive breeding. Isr. J. Plant Sci. 2011, 59, 3–13. [Google Scholar] [CrossRef]
- Green, P.S. A revision of Olea, L. (Oleaceae). Kew Bull. 2002, 57, 91–140. [Google Scholar] [CrossRef]
- Trapero, C.; Rallo, L.; López-Escudero, F.J.; Barranco, D.; Díez, C.M. Variability and selection of Verticillium wilt resistant genotypes in cultivated olive and in the Olea genus. Plant Pathol. 2014, 64, 890–900. [Google Scholar] [CrossRef]
- Shi, Z.M.; Luo, F.S.; Li, Y.; Yang, F.Y.; Xie, K.M.; Yang, W.M. Study on the rootstock (Olea ferruginea) for grafting olive. Acta Bolanica Yunnanica 1991, 13, 65–74. (In Chinese) [Google Scholar]
- Arias-Calderón, R.; Rodríguez-Jurado, D.; León, L.; Bejarano-Alcázar, J.; De La Rosa, R.; Belaj, A. Pre-breeding for resistance to Verticillium wilt in olive: Fishing in the wild relative gene pool. Crop Prot. 2015, 75, 25–33. [Google Scholar] [CrossRef]
- Wei-Ying, H. Introduction of olive trees in China. Sci. Silvae Sin. 1981, 17, 78–83. (In Chinese) [Google Scholar]
- Pan, L.; Li, Y.J.; Ma, T.; Wu, T.; Ning, D.L. SSR Identification of authenticity of interspecific hybrids between Olea europaea × cuspidata. Mol. Plant Breed. 2019, 17, 2279–2284. (In Chinese) [Google Scholar]
- Niu, E.; Jiang, C.; Wang, W.; Zhang, Y.S.Z. Comparative study on the growth and morphology of new olive variety 'Jinyefoxilan' and its parents. Nanfang Yuanyi South. Hortic. 2015, 26, 1–3. (In Chinese) [Google Scholar]
- Tian, X.; Yang, W.; Ting, M.; Na, H.; Yongjie, L.; Delu, N. Preliminary study on the characteristics of hybrid of Olea europaea L. J. West China For. Sci. 2020, 49, 130–135. (In Chinese) [Google Scholar]
- Barranco, D.; Cimato, A.; Fiorino, P.; Rallo, L.; Touzani, A.; Castaneda, C.; Serafin, F.; Truijillo, I. World Catalogue of Olive Varieties; International Olive Council: Madrid, Spain, 2000. [Google Scholar]
- de la Rosa, R.; James, C.M.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellites in olive (Olea europaea L.) and their transferability to other genera in the Oleaceae. Mol. Ecol. Notes 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Sefc, K.M.; Lopes, M.S.; Mendonça, D.; Santos, M.R.D.; Machado, M.L.D.; Machado, A.D. Identification of microsatellite loci in olive (Olea europaea) and their characterization in Italian and Iberian olive trees. Mol. Ecol. 2000, 9, 1171–1173. [Google Scholar] [CrossRef]
- Cipriani, G.; Marrazzo, M.T.; Marconi, R.; Cimato, A.; Testolin, R. Microsatellite markers isolated in olive (Olea europaea L.) are suitable for individual fingerprinting and reveal polymorphism within ancient cultivars. Theor. Appl. Genet. 2002, 104, 223–228. [Google Scholar] [CrossRef]
- de la Rosa, R.; Belaj, A.; Muñoz-Mérida, A.; Trelles, O.; Ortíz-Martín, I.; González-Plaza, J.J.; Valpuesta, V.; Beuzón, C.R. Development of EST-derived SSR markers with long-core repeat in olive and their use for paternity testing. J. Am. Soc. Hortic. Sci. 2013, 138, 290–296. [Google Scholar] [CrossRef]
- Jihène, D.; Mahdi, F.; Breton, C.M.; Msallem, M. Characterization of olive progenies derived from a Tunisian breeding program by morphological traits and SSR markers. Sci. Hortic. 2018, 236, 127–136. [Google Scholar]
- Shao, W.H. Study on Molecular Identification and Genetic Evaluation of Olive Cultivars in China. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2019. [Google Scholar]
- Koubouris, G.C.; Avramidou, E.; Metzidakis, I.T.; Petrakis, P.V.; Sergentani, C.K.; Doulis, A.G. Phylogenetic and evolutionary applications of analyzing endocarp morphological characters by classification binary tree and leaves by SSR markers for the characterization of olive germplasm. Tree Genet. Genomes 2019, 15, 26. [Google Scholar] [CrossRef]
- D’Imperio, M.; Viscosi, V.; Scarano, M.T.; D’Andrea, M.; Zullo, B.A.; Pilla, F. Integration between molecular and morphological markers for the exploitation of olive germoplasm (Olea europaea). Sci. Hortic. 2001, 130, 229–240. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the worldwide olive germplasm bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2013, 10, 141–155. [Google Scholar] [CrossRef]
- Fendri, M.; Trujillo, I.; Trigui, A.; Rodríguez-García, M.I.; Ramírez, J.D.A. Simple sequence repeat identification and endocarp characterization of olive tree accessions in a Tunisian germplasm collection. Hortscience 2010, 45, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Doveri, S.; Gil, F.S.; Díaz, A.; Reale, S.; Busconi, M.; Machado, A.D.C.; Martín, A.; Fogher, C.; Donini, P.; Lee, D. Standardization of a set of microsatellite markers for use in cultivar identification studies in olive (Olea europaea L.). Sci. Hortic. 2008, 116, 367–373. [Google Scholar] [CrossRef]
- Besnard, G.; El Bakkali, A. Sequence analysis of single-copy genes in two wild olive subspecies: Nucleotide diversity and potential use for testing admixture. Genome 2014, 57, 145–153. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| F1 Hybrid | Crossing | Collecting Region |
|---|---|---|
| ‘FJ’ | ‘Frantoio’ × subsp. cuspidata | Yunnan Province, Southern China |
| ‘CJ’ | ‘Coratina’ × subsp. cuspidata | Sichuan Province, Southern China |
| Qualitative Trait | Subsp. cuspidata (♂) | ‘Frantoio’ (♀) | ‘Coratina’ (♀) | ‘FJ’ (F1 Hybrid) | ‘CJ’ (F1 Hybrid) | |
|---|---|---|---|---|---|---|
| Leaf | Underside color | Brown | Silvery-grey | Silvery-grey | Silvery-grey | Light brown |
| Shape | Lanceolate | Elliptic | Elliptic-lanceolate | Elliptic-lanceolate | Elliptic-lanceolate | |
| Fruit | Shape | Spherical | Ovoid | Elongated | Ovoid | Ovoid |
| Apex | Rounded | Rounded | Rounded | Rounded | Rounded | |
| Base | Rounded | Truncated | Rounded | Truncated | Rounded | |
| Endocarp | Shape | Ovoid | Elliptic | Elongated | Elliptic | Ovoid |
| Apex | Rounded | Pointed | Pointed | Rounded | Rounded | |
| Base | Rounded | Pointed | Pointed | Rounded | Rounded | |
| Number of grooves | High | Medium | Medium | High | High | |
| Tip of apex | Without mucro | With mucro | With mucro | Without mucro | Without mucro | |
| Quantitative Trait | Subsp. caspidata (♂) | ‘Frantoio’ (♀) | ‘Coratina’ (♀) | ‘FJ’ (F1 Hybrid) | ‘CJ’ (F1 Hybrid) | |
|---|---|---|---|---|---|---|
| Leaf | Length/cm | 7.62 (6.66–8.63) | 6.43 (5–8.9) | 6.27 (5.1–7.1) | 5.29 (4.57–6.12) | 6.41 (4.8–8.1) |
| Width/cm | 1.21 (0.87–1.51) | 1.69 (1.28–2.09) | 1.32 (1.05–1.6) | 1.15 (0.97–1.42) | 1.23 (0.9–1.5) | |
| Shape index | 6.30 (4.41–10.65) | 3.86 (3.03–5.26) | 4.83 (3.27–6.38) | 4.6 (3.22–6.31) | 5.21 (3.2–9) | |
| Fruit | Length/cm | 1.17 | 1.64 (1.5–1.9) | 1.41 (1.3–1.5) | 1.55 (1.39–1.7) | 1.54 (1.37–1.72) |
| Width/cm | 0.96 | 2.33 (2.1–2.9) | 2.2 (2–2.4) | 1.13 (1.03–1.25) | 1.21 (1.07–1.34) | |
| Shape index | 1.22 | 1.42 (1.11–1.93) | 1.56 (1.5–1.62) | 1.37 (1.24–1.37) | 1.28 (1.18–1.45) | |
| Endocarp | Length/mm | 7–8 | 17.5 (16–19) | 18.3 (17–20) | 12.82 (11.65–14.18) | 12.64 (11.45–13.9) |
| Width/mm | 5–6 | 8.36 (7.85–9.78) | 7.81 (7.58–8) | 7.09 (6.54–7.77) | 7.48 (6.82–8.24) | |
| Shape index | 1.17–1.6 | 2.1 (1.84–2.27) | 2.34 (2.23–2.57) | 1.81 (1.67–1.93) | 1.69 (1.58–1.83) | |
| Source | SSR | Primer Sequence (5′–3′) | Repeat Motif |
|---|---|---|---|
| Sefc et al., 2000 [32] | DCA3 | F-CCCAAGCGGAGGTGTATATTGTTAC R-TGCTTTTGTCGTGTTTGAGATGTTG | (GA)19 |
| DCA11 | F-GATCAAACTACTGCACGAGAGAG R-TTGTCTAGTGAACCCTTAAACC | (GA)26(GGGA)4 | |
| DCA18 | F-AAGAAAGAAAAAGGCAGAATTAAGC R-GTTTTCGTCTCTCTACATAAGTGAC | (CA)4CT(CA)3 (GA)19 | |
| Cipriani et al., 2002 [33] | UDO99-11 | F-TGACTCCCTTTAAACTCATCAGG R-TGCGCATGTAGATGTGAATATG | (CT)7(CA)10(CT)2(CA)2 CT(CA)2CT(CA)9 |
| UDO99-19 | F-TCCCTTGTAGCCTCGTCTTG R-GGCCTGATCATCGATACCTC | (GT)20(AT)5 | |
| De la Rosa et al., 2013 [34] | EMO90 | F-CATCCGGATTTCTTGCTTTT R-AGCGAATGTAGCTTTGCATGT | (CA)10 |
| Locus | Primer Sequence (5′–3′) | Tm (°C) | GenBank Accession No. |
|---|---|---|---|
| SWH1 | F-ACTTCATTTTACACCCATTTTTA R-CACTTGATGCTTTTGTTTCTTTT | 50 | MG725069–MG725089 |
| SWH5 | F-CAAGAAAAAACAAAGAAAGAGCA R-GTAAAACTACCTGGGAACAAACC | 52 | MG725153–MG725173 |
| SWH6 | F-GGCAGAGGATTACAGGACAGG R-AGAGGGAGAAAGGGGTAGCAG | 58 | MG725174–MG725194 |
| Hybrid/Cultivars | SSR Locus | |||||
|---|---|---|---|---|---|---|
| DCA3 | DCA11 | DCA18 | UDO11 | UDO19 | EMO90 | |
| subsp.cuspidata (♂) | 239/253 | 131/141/244 | 172/196 | 112/114 | 130/130 | 182/186 |
| ‘Frantoio’ (♀) | 237/243 | 131/141 | 174/176 | 110/120 | 130/166 | 186/192 |
| ‘Coratina’ (♀) | 239/243 | 131/141 | 174/178 | 110/126 | 112/112 | 186/192 |
| ‘FJ’ (F1 hybrid) | 243/253 | 141/141 | 172/174 | 114/120 | 130/166 | 186/186 |
| ‘CJ’ (F1 hybrid) | 243/253 | 131/141 | 172/178 | 112/126 | 112/112 | 182/186 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ji, X.; Wang, Z.; Zeng, Y.; Zhang, J. Morphological, Molecular and Genomic Characterization of Two Inter-Subspecific Hybrids between Olive Cultivars and Olive Subspecies. Horticulturae 2021, 7, 138. https://doi.org/10.3390/horticulturae7060138
Li J, Ji X, Wang Z, Zeng Y, Zhang J. Morphological, Molecular and Genomic Characterization of Two Inter-Subspecific Hybrids between Olive Cultivars and Olive Subspecies. Horticulturae. 2021; 7(6):138. https://doi.org/10.3390/horticulturae7060138
Chicago/Turabian StyleLi, Jinhua, Xinyue Ji, Zhaoshan Wang, Yanfei Zeng, and Jianguo Zhang. 2021. "Morphological, Molecular and Genomic Characterization of Two Inter-Subspecific Hybrids between Olive Cultivars and Olive Subspecies" Horticulturae 7, no. 6: 138. https://doi.org/10.3390/horticulturae7060138
APA StyleLi, J., Ji, X., Wang, Z., Zeng, Y., & Zhang, J. (2021). Morphological, Molecular and Genomic Characterization of Two Inter-Subspecific Hybrids between Olive Cultivars and Olive Subspecies. Horticulturae, 7(6), 138. https://doi.org/10.3390/horticulturae7060138