


Brain Complexity and Parametrization of Power Spectral Density in Children with Specific Language Impairment
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Psychological Test
2.3. EEG Recording
2.4. Data Analysis
2.4.1. EEG Pre-Processing
2.4.2. Multiscale Entropy
2.4.3. Parameterization of Fitting Oscillations and One over F (FOOOF)
2.5. Statistical Analysis
2.5.1. Multiscale Entropy
2.5.2. Parametrization of Fitting Oscillations and One over F (FOOOF)
3. Results
3.1. Demographic, Cognitive and Technical Results of Participants
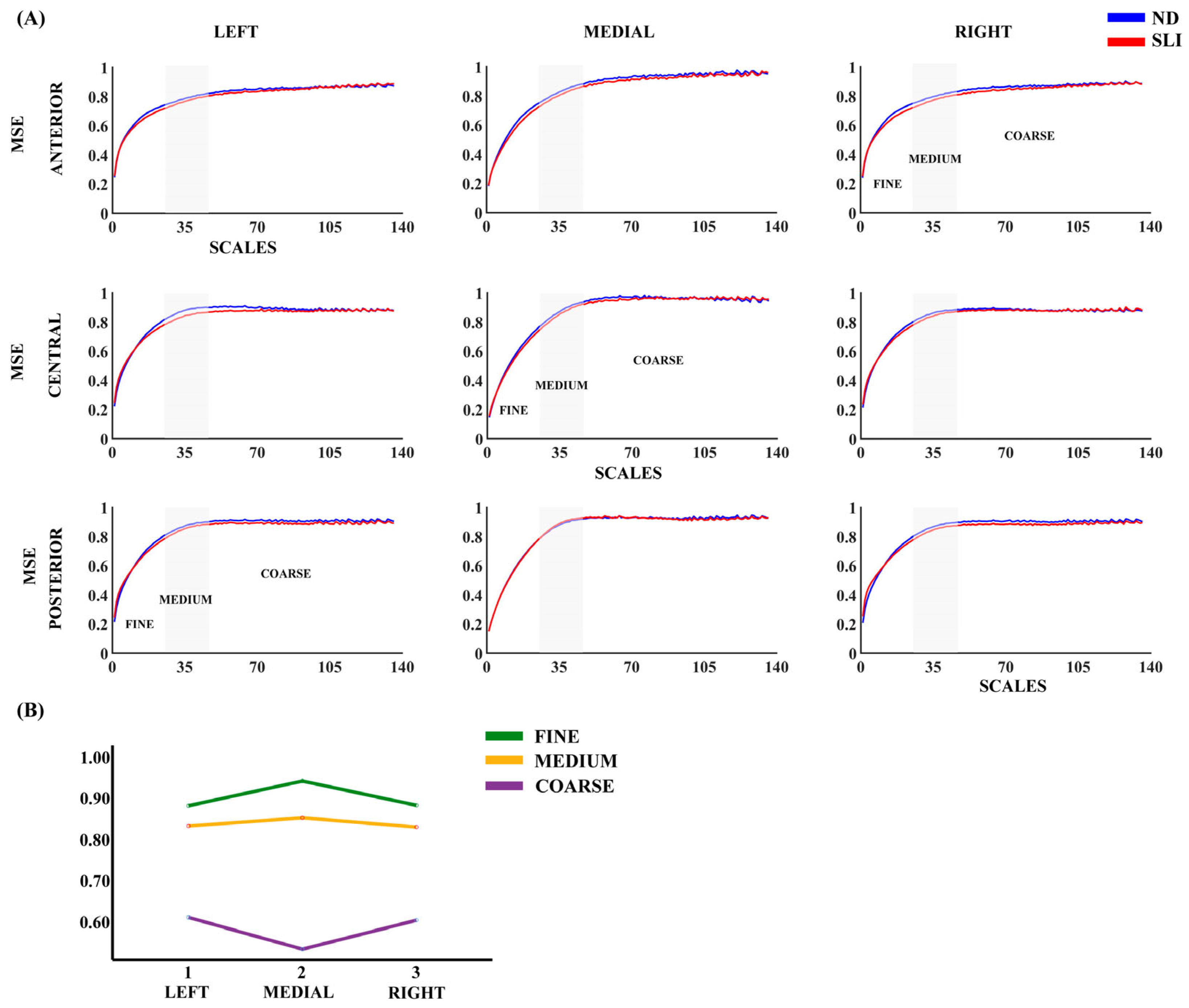
3.2. Multiscale Entropy (MSE)
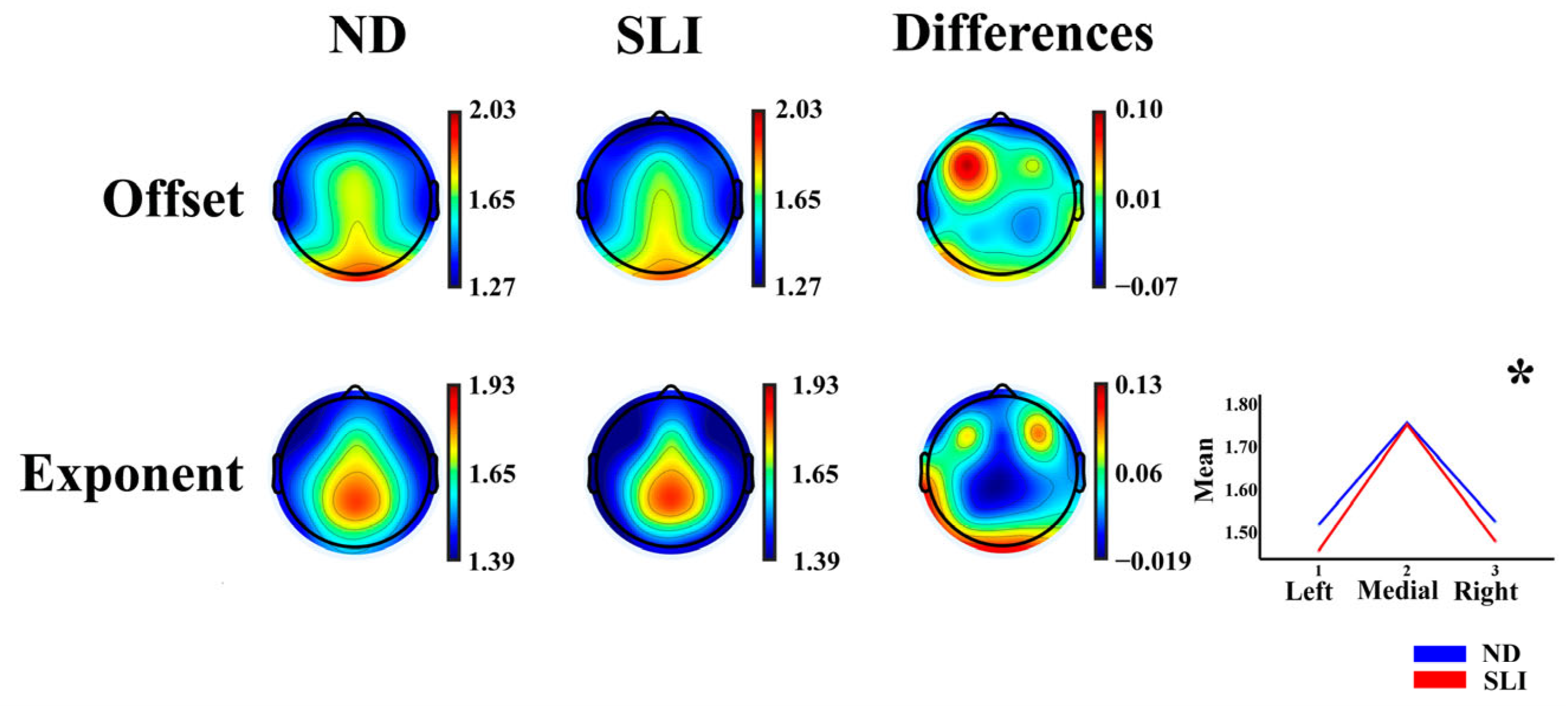
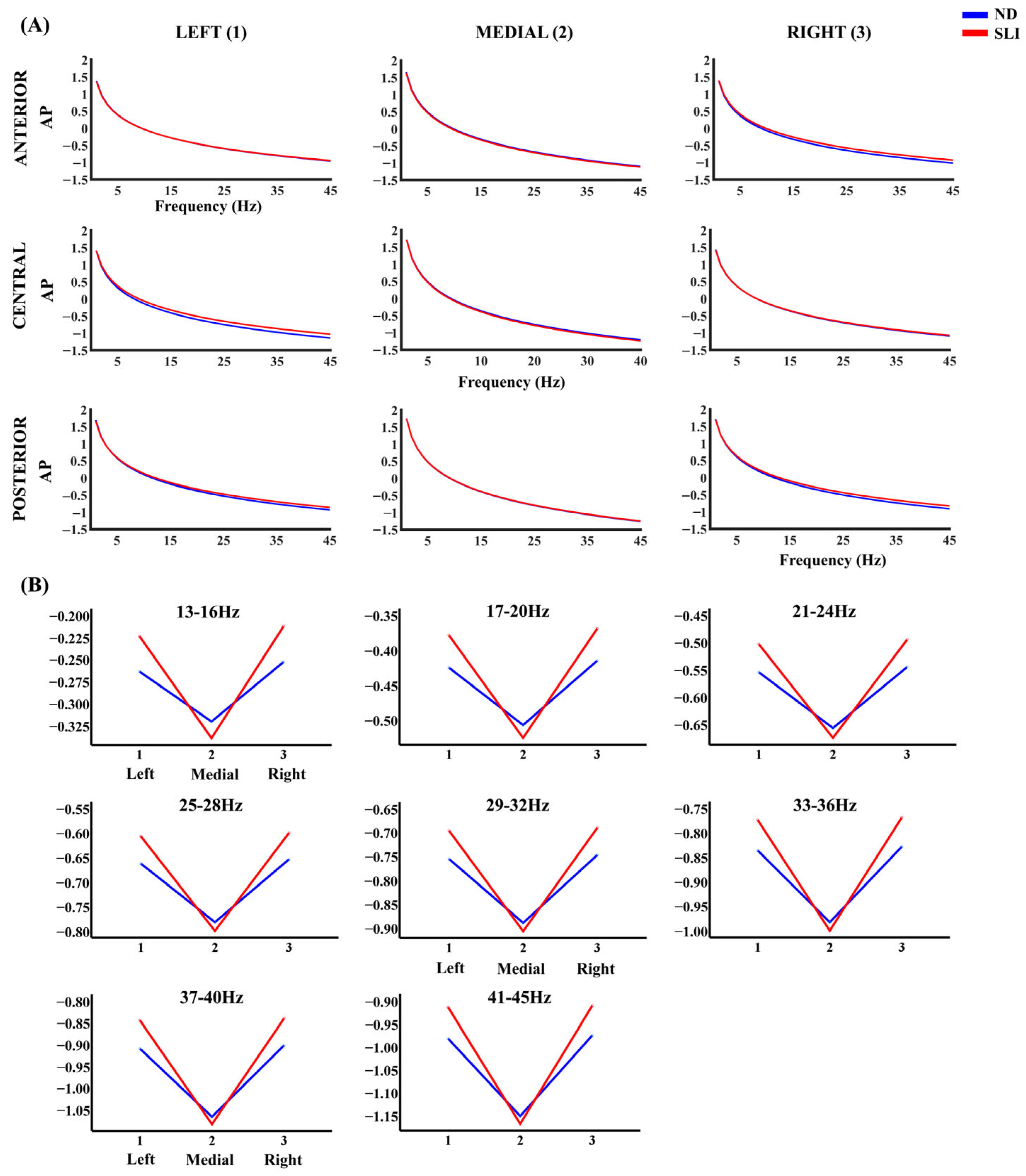
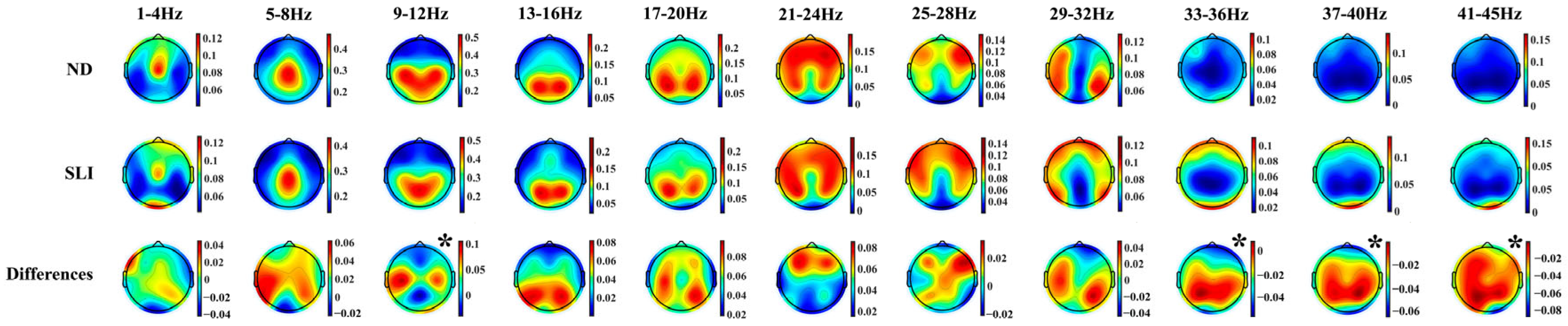
3.3. Parametrization of Fitting Oscillations and One over F (FOOOF)
3.3.1. Aperiodic Component (AP)
3.3.2. Periodic Component (P)
4. Discussion
4.1. Multiscale Entropy (MSE)
4.2. Parametrization of Fitting Oscillations and One over F (FOOOF)
4.2.1. Topographies of the Exponent and Offset Parameters of the Aperiodic Component
4.2.2. Aperiodic Component (AP)
4.2.3. Topographies of the Periodic Component
4.2.4. Periodic Component (P)
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADHD | Attention-Deficit Hyperactivity Disorder |
| AP | Aperiodic |
| ASD | Autism Spectrum Disorder |
| ASR | Artifact Subspace Reconstruction |
| CELF | Clinical Evaluation of Language Fundamentals |
| DTI | Diffusion Tensor Imaging |
| EEG | Electroencephalogram |
| ERPs | Event-Related Potentials |
| FDR | False Discovery Rate |
| fMRI | Functional Magnetic Resonance Imaging |
| FOOOF | Fitting Oscillations and One Over F |
| fTCD | Functional Transcranial Doppler |
| ICA | Independent Component Analysis |
| ITPA | Illinois Test of Psycholinguistic Abilities |
| KBIT | Kaufman Brief Intelligence Test |
| M | Mean |
| MAE | Mean Absolute Error |
| MMN | Mismatch Negativity |
| MSE | Multiscale Entropy |
| ND | Normo-Development |
| P | Periodic |
| PLON-R | Navarre Oral Language Test—Revised |
| PSD | Power Spectral Density |
| PPVT-5 | Peabody Picture Vocabulary Test |
| RM-ANOVA | Repeated-Measures Analysis of Variance |
| SD | Standard Deviation |
| SE | Sample Entropy |
| SLI | Specific Language Impairment |
| SPECT | Single-Photon Emission Computed Tomography |
| UDIATE | Unidad de Desarrollo Infantil y Atención Temprana |
References
- American Psychiatric Association. Diagnostics and Statistical Manual of Mental Disorders, IV Text Revision; American Psychiatric Association: Arlington, VA, USA, 2000. [Google Scholar]
- American Psychiatric Association. DSM-V Development. 2012. Available online: https://www.federaciocatalanatdah.org/wp-content/uploads/2018/12/dsm5-manualdiagnsticoyestadisticodelostrastornosmentales-161006005112.pdf (accessed on 1 December 2024).
- Villegas, L.F. Specific language impairment in Andalusia, Spain: Prevelance by subtype and gender. Rev. Logop. Foniatría Audiol. 2022, 42, 147–157. [Google Scholar] [CrossRef]
- Bishop, D.V. The role of genes in the etiology of specific language impairment. J. Commun. Disord. 2002, 35, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.V. Genes, cognition, and communication: Insights from neurodevelopmental disorders. Ann. N. Y. Acad. Sci. 2009, 1156, 1–18. [Google Scholar] [CrossRef]
- Bishop, D.V. Cerebral asymmetry and language development: Cause, correlate, or consequence? Science 2013, 340, 1230531. [Google Scholar] [CrossRef]
- Li, N.; Bartlett, C.W. Defining the genetic architecture of human developmental language impairment. Life Sci. 2012, 90, 469–475. [Google Scholar] [CrossRef]
- Tomblin, J.B.; Records, N.L.; Buckwalter, P.; Zhang, X.; Smith, E.; O’Brien, M. Prevalence of specific language impairment in kindergarten children. J. Speech Lang. Hear. Res. 1997, 40, 1245–1260. [Google Scholar] [CrossRef]
- Pennington, B.F.; Bishop, D.V. Relations among speech, language, and reading disorders. Annu. Rev. Psychol. 2009, 60, 283–306. [Google Scholar] [CrossRef] [PubMed]
- Young, A.R.; Beitchman, J.H.; Johnson, C.; Douglas, L.; Atkinson, L.; Escobar, M.; Wilson, B. Young adult academic outcomes in a longitudinal sample of early identified language impaired and control children. J. Child Psychol. Psychiatry Allied Discip. 2002, 43, 635–645. [Google Scholar] [CrossRef]
- Wadman, R.; Botting, N.; Durkin, K.; Conti-Ramsden, G. Changes in emotional health symptoms in adolescents with specific language impairment. Int. J. Lang. Commun. Disord. 2011, 46, 641–656. [Google Scholar] [CrossRef]
- Wadman, R.; Durkin, K.; Conti-Ramsden, G. Close relationships in adolescents with and without a history of specific language impairment. Lang. Speech Hear. Serv. Sch. 2011, 42, 41–51. [Google Scholar] [CrossRef]
- Wadman, R.; Durkin, K.; Conti-Ramsden, G. Social stress in young people with specific language impairment. J. Adolesc. 2011, 34, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Arkkila, E.; Rasanen, P.; Roine, R.P.; Vilkman, E. Specific language impairment in childhood is associated with impaired mental and social well-being in adulthood. Logop. Phoniatr. Vocol. 2008, 33, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Tallal, P. Improving language and literacy is a matter of time. Nat. Rev. Neurosci. 2004, 5, 721–728. [Google Scholar] [CrossRef]
- Hsu, H.J.; Tomblin, J.B.; Christiansen, M.H. Impaired statistical learning of non-adjacent dependencies in adolescents with specific language impairment. Front. Psychol. 2014, 5, 175. [Google Scholar] [CrossRef]
- Goswami, U. Speech rhythm and language acquisition: An amplitude modulation phase hierarchy perspective. Ann. N. Y. Acad. Sci. 2019, 1453, 67–78. [Google Scholar] [CrossRef]
- Abbott, N.; Love, T. Bridging the Divide: Brain and Behavior in Developmental Language Disorder. Brain Sci. 2023, 13, 1606. [Google Scholar] [CrossRef]
- Gauger, L.M.; Lombardino, L.J.; Leonard, C.M. Brain morphology in children with specific language impairment. J. Speech Lang. Hear. Res. 1997, 40, 1272–1284. [Google Scholar] [CrossRef]
- Herbert, M.R.; Ziegler, D.A.; Makris, N.; Filipek, P.A.; Kemper, T.L.; Normandin, J.J.; Sanders, H.A.; Kennedy, D.N.; Caviness, V.S., Jr. Localization of white matter volume increase in autism and developmental language disorder. Ann. Neurol. 2004, 55, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Bahar, N.; Cler, G.J.; Krishnan, S.; Asaridou, S.S.; Smith, H.J.; Willis, H.E.; Healy, M.P.; Watkins, K.E. Differences in Cortical Surface Area in Developmental Language Disorder. Neurobiol. Lang. 2024, 5, 288–314. [Google Scholar] [CrossRef]
- Badcock, N.A.; Bishop, D.V.; Hardiman, M.J.; Barry, J.G.; Watkins, K.E. Co-localisation of abnormal brain structure and function in specific language impairment. Brain Lang. 2012, 120, 310–320. [Google Scholar] [CrossRef]
- Soriano-Mas, C.; Pujol, J.; Ortiz, H.; Deus, J.; López-Sala, A.; Sans, A. Age-related brain structural alterations in children with specific language impairment. Hum. Brain Mapp. 2009, 30, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Nopoulos, P.C.; Bruce Tomblin, J. Abnormal subcortical components of the corticostriatal system in young adults with DLI: A combined structural MRI and DTI study. Neuropsychologia 2013, 51, 2154–2161. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Cler, G.J.; Smith, H.J.; Willis, H.E.; Asaridou, S.S.; Healy, M.P.; Papp, D.; Watkins, K.E. Quantitative MRI reveals differences in striatal myelin in children with DLD. eLife 2022, 11, e74242. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.R.; Ziegler, D.A.; Makris, N.; Bakardjiev, A.; Hodgson, J.; Adrien, K.T.; Kennedy, D.N.; Filipek, P.A.; Caviness, V.S., Jr. Largerbrain and white matter volumes in children with developmental language disorder. Dev. Sci. 2003, 6, F11–F22. [Google Scholar] [CrossRef]
- Hugdahl, K.; Gundersen, H.; Brekke, C.; Thomsen, T.; Rimol, L.M.; Ersland, L.; Niemi, J. FMRI brain activation in a finnish family with specific language impairment compared with a normal control group. J. Speech Lang. Hear. Res. 2004, 47, 162–172. [Google Scholar] [CrossRef]
- de Guibert, C.; Maumet, C.; Jannin, P.; Ferré, J.C.; Tréguier, C.; Barillot, C.; Le Rumeur, E.; Allaire, C.; Biraben, A. Abnormal functional lateralization and activity of language brain areas in typical specific language impairment (developmental dysphasia). Brain 2011, 134, 3044–3058. [Google Scholar] [CrossRef]
- Ors, M.; Ryding, E.; Lindgren, M.; Gustafsson, P.; Blennow, G.; Rosén, I. SPECT findings in children with specific language impairment. Cortex J. Devoted Study Nerv. Syst. Behav. 2005, 41, 316–326. [Google Scholar] [CrossRef]
- Whitehouse, A.J.; Bishop, D.V. Cerebral dominance for language function in adults with specific language impairment or autism. Brain J. Neurol. 2008, 131, 3193–3200. [Google Scholar] [CrossRef]
- Vydrova, R.; Komarek, V.; Sanda, J.; Sterbova, K.; Jahodova, A.; Maulisova, A.; Zackova, J.; Reissigova, J.; Krsek, P.; Kyncl, M. Structural alterations of the language connectome in children with specific language impairment. Brain Lang. 2015, 151, 35–41. [Google Scholar] [CrossRef]
- Roberts, T.P.; Heiken, K.; Zarnow, D.; Dell, J.; Nagae, L.; Blaskey, L.; Solot, C.; Levy, S.E.; Berman, J.I.; Edgar, J.C. Left hemisphere diffusivity of the arcuate fasciculus: Influences of autism spectrum disorder and language impairment. Am. J. Neuroradiol. 2014, 35, 587–592. [Google Scholar] [CrossRef]
- Lee, J.C.; Dick, A.S.; Tomblin, J.B. Altered brain structures in the dorsal and ventral language pathways in individuals with and without developmental language disorder (DLD). Brain Imaging Behav. 2020, 14, 2569–2586. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.V.; McArthur, G.M. Individual differences in auditory processing in specific language impairment: A follow-up study using event-related potentials and behavioural thresholds. Cortex J. Devoted Study Nerv. Syst. Behav. 2005, 41, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Shafer, V.L.; Morr, M.L.; Datta, H.; Kurtzberg, D.; Schwartz, R.G. Neurophysiological indexes of speech processing deficits in children with specific language impairment. J. Cogn. Neurosci. 2005, 17, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Datta, H.; Shafer, V.L.; Morr, M.L.; Kurtzberg, D.; Schwartz, R.G. Electrophysiological indices of discrimination of long-duration, phonetically similar vowels in children with typical and atypical language development. J. Speech Lang. Hear. Res. 2010, 53, 757–777. [Google Scholar] [CrossRef]
- Kujala, T.; Leminen, M. Low-level neural auditory discrimination dysfunctions in specific language impairment-A review on mismatch negativity findings. Dev. Cogn. Neurosci. 2017, 28, 65–75. [Google Scholar] [CrossRef]
- Sabisch, B.; Hahne, A.; Glass, E.; von Suchodoletz, W.; Friederici, A.D. Lexical–semantic processes in children with specific language impairment. NeuroReport 2006, 17, 1511–1514. [Google Scholar] [CrossRef]
- Haebig, E.; Leonard, L.; Usler, E.; Deevy, P.; Weber, C. An Initial Investigation of the Neural Correlates of Word Processing in Preschoolers with Specific Language Impairment. J. Speech Lang. Hear. Res. 2018, 61, 729–739. [Google Scholar] [CrossRef]
- Shafer, V.L.; Ponton, C.; Datta, H.; Morr, M.L.; Schwartz, R.G. Neurophysiological indices of attention to speech in children with specific language impairment. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2007, 118, 1230–1243. [Google Scholar] [CrossRef]
- Shafer, V.L.; Schwartz, R.G.; Martin, B. Evidence of deficient central speech processing in children with specific language impairment: The T-complex. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2011, 122, 1137–1155. [Google Scholar] [CrossRef]
- Albrecht, R.; Suchodoletz, W.; Uwer, R. The development of auditory evoked dipole source activity from childhood to adulthood. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2000, 111, 2268–2276. [Google Scholar] [CrossRef]
- Shaheen, E.A.; Shohdy, S.S.; Abd Al Raouf, M.; Mohamed El Abd, S.; Abd Elhamid, A. Relation between language, audio-vocal psycholinguistic abilities and P300 in children having specific language impairment. Int. J. Pediatr. Otorhinolaryngol. 2011, 75, 1117–1122. [Google Scholar] [CrossRef]
- Evans, J.L.; Selinger, C.; Pollak, S.D. P300 as a measure of processing capacity in auditory and visual domains in specific language impairment. Brain Res. 2011, 1389, 93–102. [Google Scholar] [CrossRef]
- Gasser, T.; Verleger, R.; Bächer, P.; Sroka, L. Development of the EEG of school-age children and adolescents. I. Analysis of band power. Electroencephalogr. Clin. Neurophysiol. 1988, 69, 91–99. [Google Scholar] [CrossRef]
- Whitford, T.J.; Rennie, C.J.; Grieve, S.M.; Clark, C.R.; Gordon, E.; Williams, L.M. Brain maturation in adolescence: Concurrent changes in neuroanatomy and neurophysiology. Hum. Brain Mapp. 2007, 28, 228–237. [Google Scholar] [CrossRef]
- Segalowitz, S.J.; Santesso, D.L.; Jetha, M.K. Electrophysiological changes during adolescence: A review. Brain Cogn. 2010, 72, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Miskovic, V.; Ma, X.; Chou, C.A.; Fan, M.; Owens, M.; Sayama, H.; Gibb, B.E. Developmental changes in spontaneous electrocortical activity and network organization from early to late childhood. NeuroImage 2015, 118, 237–247. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, E.I.; Ruiz-Martínez, F.J.; Barriga Paulino, C.I.; Gómez, C.M. Frequency shift in topography of spontaneous brain rhythms from childhood to adulthood. Cogn. Neurodyn. 2017, 11, 23–33. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, E.I.; Angulo-Ruiz, B.Y.; Arjona-Valladares, A.; Rufo, M.; Gómez-González, J.; Gómez, C.M. Frequency coupling of low and high frequencies in the EEG of ADHD children and adolescents in closed and open eyes conditions. Res. Dev. Disabil. 2020, 96, 103520. [Google Scholar] [CrossRef] [PubMed]
- Lea-Carnall, C.A.; Montemurro, M.A.; Trujillo-Barreto, N.J.; Parkes, L.M.; El-Deredy, W. Cortical Resonance Frequencies Emerge from Network Size and Connectivity. PLoS Comput. Biol. 2016, 12, e1004740. [Google Scholar] [CrossRef] [PubMed]
- Szostakiwskyj, J.M.H.; Willatt, S.E.; Cortese, F.; Protzner, A.B. The modulation of EEG variability between internally- and externally-driven cognitive states varies with maturation and task performance. PLoS ONE 2017, 12, e0181894. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; Johnstone, S.J.; McCarthy, R.; Selikowitz, M. Electroencephalogram theta/beta ratio and arousal in attention-deficit/hyperactivity disorder: Evidence of independent processes. Biol. Psychiatry 2009, 66, 398–401. [Google Scholar] [CrossRef]
- Newson, J.J.; Thiagarajan, T.C. EEG Frequency Bands in Psychiatric Disorders: A Review of Resting State Studies. Front. Hum. Neurosci. 2019, 12, 521. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; Johnstone, S. Resting state EEG power research in Attention-Deficit/Hyperactivity Disorder: A review update. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2020, 131, 1463–1479. [Google Scholar] [CrossRef]
- Dawson, G.; Klinger, L.G.; Panagiotides, H.; Lewy, A.; Castelloe, P. Subgroups of autistic children based on social behavior display distinct patterns of brain activity. J. Abnorm. Child Psychol. 1995, 23, 569–583. [Google Scholar] [CrossRef]
- Daoust, A.M.; Limoges, E.; Bolduc, C.; Mottron, L.; Godbout, R. EEG spectral analysis of wakefulness and REM sleep in high functioning autistic spectrum disorders. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2004, 115, 1368–1373. [Google Scholar] [CrossRef]
- Chan, A.S.; Sze, S.L.; Cheung, M.C. Quantitative electroencephalographic profiles for children with autistic spectrum disorder. Neuropsychology 2007, 21, 74–81. [Google Scholar] [CrossRef]
- Pop-Jordanova, N.; Zorcec, T.; Demerdzieva, A.; Gucev, Z. QEEG characteristics and spectrum weighted frequency for children diagnosed as autistic spectrum disorder. Nonlinear Biomed. Phys. 2010, 4, 4. [Google Scholar] [CrossRef]
- Wang, J.; Barstein, J.; Ethridge, L.E.; Mosconi, M.W.; Takarae, Y.; Sweeney, J.A. Resting state EEG abnormalities in autism spectrum disorders. J. Neurodev. Disord. 2013, 5, 24. [Google Scholar] [CrossRef]
- Sohn, H.; Kim, I.; Lee, W.; Peterson, B.S.; Hong, H.; Chae, J.H.; Hong, S.; Jeong, J. Linear and non-linear EEG analysis of adolescents with attention-deficit/hyperactivity disorder during a cognitive task. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2010, 121, 1863–1870. [Google Scholar] [CrossRef]
- Sokunbi, M.O.; Fung, W.; Sawlani, V.; Choppin, S.; Linden, D.E.; Thome, J. Resting state fMRI entropy probes complexity of brain activity in adults with ADHD. Psychiatry Res. 2013, 214, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Rezaeezadeh, M.; Shamekhi, S.; Shamsi, M. Attention Deficit Hyperactivity Disorder Diagnosis using non-linear univariate and multivariate EEG measurements: A preliminary study. Phys. Eng. Sci. Med. 2020, 43, 577–592. [Google Scholar] [CrossRef]
- Angulo-Ruiz, B.Y.; Muñoz, V.; Rodríguez-Martínez, E.I.; Cabello-Navarro, C.; Gómez, C.M. Multiscale entropy of ADHD children during resting state condition. Cogn. Neurodyn. 2022, 17, 869–891. [Google Scholar] [CrossRef]
- Chu, Y.J.; Chang, C.F.; Shieh, J.S.; Lee, W.T. The Potential Application of Multiscale Entropy Analysis of Electroencephalography in Children with Neurological and Neuropsychiatric Disorders. Entropy 2017, 19, 428. [Google Scholar] [CrossRef]
- Liu, T.; Chen, Y.; Chen, D.; Li, C.; Qiu, Y.; Wang, J. Altered electroencephalogram complexity in autistic children shown by the multiscale entropy approach. NeuroReport 2017, 28, 169–173. [Google Scholar] [CrossRef]
- Milne, E.; Gomez, R.; Giannadou, A.; Jones, M. Atypical EEG in autism spectrum disorder: Comparing a dimensional and a categorical approach. J. Abnorm. Psychol. 2019, 128, 442–452. [Google Scholar] [CrossRef]
- Angulo-Ruiz, B.Y.; Ruiz-Martínez, F.J.; Rodríguez-Martínez, E.I.; Ionescu, A.; Saldaña, D.; Gómez, C.M. Linear and Non-linear Analyses of EEG in a Group of ASD Children During Resting State Condition. Brain Topogr. 2023, 36, 736–749. [Google Scholar] [CrossRef]
- DiStefano, C.; Dickinson, A.; Baker, E.; Jeste, S.S. EEG Data Collection in Children with ASD: The Role of State in Data Quality and Spectral Power. Res. Autism Spectr. Disord. 2019, 57, 132–144. [Google Scholar] [CrossRef]
- Pierce, S.; Kadlaskar, G.; Edmondson, D.A.; McNally Keehn, R.; Dydak, U.; Keehn, B. Associations between sensory processing and electrophysiological and neurochemical measures in children with ASD: An EEG-MRS study. J. Neurodev. Disord. 2021, 13, 5. [Google Scholar] [CrossRef]
- Takahashi, T.; Yoshimura, Y.; Hiraishi, H.; Hasegawa, C.; Munesue, T.; Higashida, H.; Minabe, Y.; Kikuchi, M. Enhanced brain signal variability in children with autism spectrum disorder during early childhood. Hum. Brain Mapp. 2016, 37, 1038–1050. [Google Scholar] [CrossRef]
- Bosl, W.J.; Loddenkemper, T.; Nelson, C.A. Nonlinear EEG biomarker profiles for autism and absence epilepsy. Neuropsychiatr. Electrophysiol. 2017, 3, 1. [Google Scholar] [CrossRef]
- Nenadović, V.; Stokić, M.; Vuković, M.; Đoković, S.; Subotić, M. Cognitive and electrophysiological characteristics of children with specific language impairment and subclinical epileptiform electroencephalogram. J. Clin. Exp. Neuropsychol. 2014, 36, 981–991. [Google Scholar] [CrossRef]
- Chutko, L.S.; Surushkina, S.Y.; Yakovenko, E.A.; Sergeev, A.V.; Rozhkova, A.V.; Anosova, L.V.; Chistyakova, N.P. Clinical and electroencephalographic characteristics of specific language impairment in children and an evaluation of the efficacy of cerebrolysin. Zhurnal Nevrologii i Psikhiatrii Imeni SS Korsakova 2015, 115, 98–102. [Google Scholar] [CrossRef]
- Fatić, S.; Stanojević, N.; Stokić, M.; Nenadović, V.; Jeličić, L.; Bilibajkić, R.; Gavrilović, A.; Maksimović, S.; Adamović, T.; Subotić, M. Electroencephalography correlates of word and non-word listening in children with specific language impairment: An observational study20F0. Medicine 2022, 101, e31840. [Google Scholar] [CrossRef]
- Stanojevic, N.; Fatic, S.; Jelicic, L.; Nenadovic, V.; Stokic, M.; Bilibajkic, R.; Subotic, M.; Boskovic Matic, T.; Konstantinovic, L.; Cirovic, D. Resting-state EEG alpha rhythm spectral power in children with specific language impairment: A cross-sectional study. J. Appl. Biomed. 2023, 21, 113–120. [Google Scholar] [CrossRef]
- Donoghue, T.; Haller, M.; Peterson, E.J.; Varma, P.; Sebastian, P.; Gao, R.; Noto, T.; Lara, A.H.; Wallis, J.D.; Knight, R.T.; et al. Parameterizing neural power spectra into periodic and aperiodic components. Nat. Neurosci. 2020, 23, 1655–1665. [Google Scholar] [CrossRef]
- McSweeney, M.; Morales, S.; Valadez, E.A.; Buzzell, G.A.; Yoder, L.; Fifer, W.P.; Pini, N.; Shuffrey, L.C.; Elliott, A.J.; Isler, J.R.; et al. Age-related trends in aperiodic EEG activity and alpha oscillations during early- to middle-childhood. NeuroImage 2023, 269, 119925. [Google Scholar] [CrossRef]
- Buzsáki, G.; Anastassiou, C.A.; Koch, C. The origin of extracellular fields and currents--EEG, ECoG, LFP and spikes. Nature reviews. Neuroscience 2012, 13, 407–420. [Google Scholar] [CrossRef]
- Gao, R.; Peterson, E.J.; Voytek, B. Inferring synaptic excitation/inhibition balance from field potentials. NeuroImage 2017, 158, 70–78. [Google Scholar] [CrossRef]
- Robertson, M.M.; Furlong, S.; Voytek, B.; Donoghue, T.; Boettiger, C.A.; Sheridan, M.A. EEG power spectral slope differs by ADHD status and stimulant medication exposure in early childhood. J. Neurophysiol. 2019, 122, 2427–2437. [Google Scholar] [CrossRef]
- Pertermann, M.; Bluschke, A.; Roessner, V.; Beste, C. The Modulation of Neural Noise Underlies the Effectiveness of Methylphenidate Treatment in Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2019, 4, 743–750. [Google Scholar] [CrossRef]
- Mamiya, P.C.; Arnett, A.B.; Stein, M.A. Precision Medicine Care in ADHD: The Case for Neural Excitation and Inhibition. Brain Sci. 2021, 11, 91. [Google Scholar] [CrossRef]
- Ostlund, B.D.; Alperin, B.R.; Drew, T.; Karalunas, S.L. Behavioral and cognitive correlates of the aperiodic (1/f-like) exponent of the EEG power spectrum in adolescents with and without ADHD. Dev. Cogn. Neurosci. 2021, 48, 100931. [Google Scholar] [CrossRef]
- Levin, A.R.; Naples, A.J.; Scheffler, A.W.; Webb, S.J.; Shic, F.; Sugar, C.A.; Murias, M.; Bernier, R.A.; Chawarska, K.; Dawson, G.; et al. Day-to-Day Test-Retest Reliability of EEG Profiles in Children With Autism Spectrum Disorder and Typical Development. Front. Integr. Neurosci. 2020, 14, 21. [Google Scholar] [CrossRef]
- Wiig, E.H.; Semel, E.; Secord, W.A. Clinical Evaluation of Language Fundamentals—Fifth Edition (CELF-5); NCS Pearson: Bloomington, MN, USA, 2013. [Google Scholar]
- Aguinaga Ayerra, G. PLON-R: Prueba de Lenguaje Oral Navarra Revisada, 2nd ed.; TEA Ediciones, S.A.U.: Madrid, España, 2005. [Google Scholar]
- Kirk, S.A.; McCarthy, J.J. The Illinois test of psycholinguistic abilities--an approach to differential diagnosis. Am. J. Ment. Defic. 1961, 66, 399–412. [Google Scholar]
- Dunn, L.M. Peabody Picture Vocabulary Test; American Guidance Service: Miami, FL, USA, 1959. [Google Scholar]
- Kaufman, A.S.; Kaufman, N.L. KBIT: Kaufman Brief Intelligence Test (KBIT Spanish Version); ASD Editions: Madrid, Spain, 2004. [Google Scholar]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Mullen, T.R.; Kothe, C.A.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.P.; Cauwenberghs, G. Real-Time Neuroimaging and Cognitive Monitoring Using Wearable Dry EEG. IEEE Trans. Bio-Med. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef]
- Bell, A.J.; Sejnowski, T.J. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995, 7, 1129–1159. [Google Scholar] [CrossRef]
- Amari, S.; Cichocki, A.; Yang, H.H. A New Learning Algorithm for Blind Signal Separation; NIPS: San Diego, CA, USA, 1995. [Google Scholar]
- Pion-Tonachini, L.; Kreutz-Delgado, K.; Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage 2019, 198, 181–197. [Google Scholar] [CrossRef]
- Malik, J. Multiscale Sample Entropy. 2022. Available online: https://www.mathworks.com/matlabcentral/fileexchange/62706-multiscale-sample-entropy (accessed on 1 December 2024).
- Costa, M.; Goldberger, A.L.; Peng, C.K. Multiscale entropy analysis of biological signals. Phys. Rev. E 2005, 71, 021906. [Google Scholar] [CrossRef]
- Richman, J.S.; Moorman, J.R. Physiological time-series analysis using approximate entropy and sample entropy. Am. J. physiology. Heart Circ. Physiol. 2000, 278, H2039–H2049. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Kovacevic, N.; Itier, R.J. Increased brain signal variability accompanies lower behavioral variability in development. PLoS Comput. Biol. 2008, 4, e1000106. [Google Scholar] [CrossRef]
- Miskovic, V.; Owens, M.; Kuntzelman, K.; Gibb, B.E. Charting moment-to-moment brain signal variability from early to late childhood. Cortex J. Devoted Study Nerv. Syst. Behav. 2016, 83, 51–61. [Google Scholar] [CrossRef]
- Kloosterman, N.A.; Kosciessa, J.Q.; Lindenberger, U.; Fahrenfort, J.J.; Garrett, D.D. Boosting brain signal variability underlies liberal shifts in decision bias. bioRxiv 2019. [Google Scholar] [CrossRef]
- Kosciessa, J.Q.; Kloosterman, N.A.; Garrett, D.D. Standard multiscale entropy reflects neural dynamics at mismatched temporal scales: What’s signal irregularity got to do with it? PLoS Comput. Biol. 2020, 16, e1007885. [Google Scholar] [CrossRef]
- Papaioannou, A.G.; Kalantzi, E.; Papageorgiou, C.C.; Korombili, K.; Βokou, A.; Pehlivanidis, A.; Papageorgiou, C.C.; Papaioannou, G. Complexity analysis of the brain activity in Autism Spectrum Disorder (ASD) and Attention Deficit Hyperactivity Disorder (ADHD) due to cognitive loads/demands induced by Aristotle’s type of syllogism/reasoning. A Power Spectral Density and multiscale entropy (MSE) analysis. Heliyon 2021, 7, e07984. [Google Scholar] [CrossRef]
- Garrett, D.D.; Samanez-Larkin, G.R.; MacDonald, S.W.; Lindenberger, U.; McIntosh, A.R.; Grady, C.L. Moment-to-moment brain signal variability: A next frontier in human brain mapping? Neurosci. Biobehav. Rev. 2013, 37, 610–624. [Google Scholar] [CrossRef]
- Bosl, W.J.; Loddenkemper, T.; Vieluf, S. Coarse-graining and the Haar wavelet transform for multiscale analysis. Bioelectron. Med. 2022, 8, 3. [Google Scholar] [CrossRef]
- Angulo-Ruiz, B.Y.; Rodríguez-Martínez, E.I.; Muñoz, V.; Gómez, C.M. Unveiling the hidden electroencephalographical rhythms during development: Aperiodic and Periodic activity in healthy subjects. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2024, 169, 53–64. [Google Scholar] [CrossRef]
- Ostlund, B.; Donoghue, T.; Anaya, B.; Gunther, K.E.; Karalunas, S.L.; Voytek, B.; Pérez-Edgar, K.E. Spectral parameterization for studying neurodevelopment: How and why. Dev. Cogn. Neurosci. 2022, 54, 101073. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates: Hillsdale, MI, USA, 1988. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Jacob, M.S.; Roach, B.J.; Sargent, K.; Mathalon, D.H.; Ford, J.M. Aperiodic measures of neural excitability are associated with anticorrelated hemodynamic networks at rest: A combined EEG-fMRI study. NeuroImage 2021, 245, 1187052021. [Google Scholar] [CrossRef]
- Hill, A.T.; Clark, G.M.; Bigelow, F.J.; Lum, J.A.G.; Enticott, P.G. Periodic and aperiodic neural activity displays age-dependent changes across early-to-middle childhood. Dev. Cogn. Neurosci. 2022, 54, 101076. [Google Scholar] [CrossRef]
- Manning, J.R.; Jacobs, J.; Fried, I.; Kahana, M.J. Broadband shifts in local field potential power spectra are correlated with single-neuron spiking in humans. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 13613–13620. [Google Scholar] [CrossRef]
- Miller, K.J.; Honey, C.J.; Hermes, D.; Rao, R.P.; denNijs, M.; Ojemann, J.G. Broadband changes in the cortical surface potential track activation of functionally diverse neuronal populations. NeuroImage 2014, 85 Pt 2, 711–720. [Google Scholar] [CrossRef]
- Voytek, B.; Knight, R.T. Dynamic network communication as a unifying neural basis for cognition, development, aging, and disease. Biol. Psychiatry 2015, 77, 1089–1097. [Google Scholar] [CrossRef]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef]
- Proal, E.; Álvarez-Segura, M.; De la Iglesia-Vayá, M.; Martí-Bonmatí, L.; Castellanos, F.X.; Spanish Resting State Network. Actividad funcional cerebral en estado de reposo: Redes en conexion [Functional cerebral activity in a state of rest: Connectivity networks]. Rev. Neurol. 2011, 52 (Suppl. S1), S3–S10. [Google Scholar] [CrossRef]
- Wilke, M.; Hauser, T.K.; Krägeloh-Mann, I.; Lidzba, K. Specific impairment of functional connectivity between language regions in former early preterms. Hum. Brain Mapp. 2014, 35, 3372–3384. [Google Scholar] [CrossRef]
- Barnes-Davis, M.E.; Williamson, B.J.; Merhar, S.L.; Holland, S.K.; Kadis, D.S. Rewiring the extremely preterm brain: Altered structural connectivity relates to language function. NeuroImage Clin. 2020, 25, 102194. [Google Scholar] [CrossRef]
- Chen, A.C.; Feng, W.; Zhao, H.; Yin, Y.; Wang, P. EEG default mode network in the human brain: Spectral regional field powers. NeuroImage 2008, 41, 561–574. [Google Scholar] [CrossRef]
- Gómez, C.M.; Marco-Pallarés, J.; Grau, C. Location of brain rhythms and their modulation by preparatory attention estimated by current density. Brain Res. 2006, 1107, 151–160. [Google Scholar] [CrossRef]
- Rodríguez-Martinez, E.I.; Barriga-Paulino, C.I.; Zapata, M.I.; Chinchilla, C.; López-Jiménez, A.M.; Gómez, C.M. Narrow band quantitative and multivariate electroencephalogram analysis of peri-adolescent period. BMC Neurosci. 2012, 13, 104. [Google Scholar] [CrossRef]
- Finneran, D.A.; Francis, A.L.; Leonard, L.B. Sustained attention in children with specific language impairment (SLI). J. Speech Lang. Hear. Res. 2009, 52, 915–929. [Google Scholar] [CrossRef]
- Verche, B.E.; Hernández, E.S.; Quintero, F.I.; Acosta, R.V.M. Alteraciones en la memoria en el Trastorno Específico del Lenguaje. Una perspectiva neuropsicológica. Rev. Logop. Foniatría Audiol. 2013, 33, 179–185. [Google Scholar] [CrossRef]
- Quintero, I.; Hernández, S.; Verche, E.; Acosta, V.; Hernández, A. Disfunción ejecutiva en el Trastorno Específico del Lenguaje. Rev. Logop. Foniatría Audiol. 2013, 33, 172–178. [Google Scholar] [CrossRef]
- Rodríguez, A.; Ramírez, S.G.M.; Expósito, H. Funciones ejecutivas y lenguaje en subtipos de niños con trastorno específico del lenguaje. Neurología 2017, 32, 355–362. [Google Scholar] [CrossRef]
- Bosman, C.A.; Lansink, C.S.; Pennartz, C.M. Functions of gamma-band synchronization in cognition: From single circuits to functional diversity across cortical and subcortical systems. Eur. J. Neurosci. 2014, 39, 1982–1999. [Google Scholar] [CrossRef]
- Friedman, N.P.; Robbins, T.W. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2022, 47, 72–89. [Google Scholar] [CrossRef]
- Mazza, A.; Dal Monte, O.; Schintu, S.; Colombo, S.; Michielli, N.; Sarasso, P.; Törlind, P.; Cantamessa, M.; Montagna, F.; Ricci, R. Beyond alpha-band: The neural correlate of creative thinking. Neuropsychologia 2023, 179, 108446. [Google Scholar] [CrossRef]
- Hindriks, R.; van Putten, M.J.A.M.; Deco, G. Intra-cortical propagation of EEG alpha oscillations. NeuroImage 2014, 103, 444–453. [Google Scholar] [CrossRef]
- Klimesch, W. α-band oscillations, attention, and controlled access to stored information. Trends Cogn. Sci. 2012, 16, 606–617. [Google Scholar] [CrossRef] [PubMed]
- London, R.E.; Benwell, C.S.Y.; Cecere, R.; Quak, M.; Thut, G.; Talsma, D. EEG alpha power predicts the temporal sensitivity of multisensory perception. Eur. J. Neurosci. 2022, 55, 3241–3255. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SLI | ND | |
|---|---|---|
| Age | M = 6.38, SD = 1.75 | M = 6.90, SD = 2.08 |
| Males | M = 6.52, SD = 1.71 | M = 7.05, SD = 2.09 |
| Females | M = 6.09, SD = 1.87 | M = 6.29, SD = 2.05 |
| KBIT | M = 22.52, SD = 7.49 | M = 26.63, SD = 6.49 |
| Components | M = 12.93, SD = 0.254 | M = 12.56, SD = 0.652 |
| Epochs | M = 27.87, SD = 2.45 | M = 32.58, SD = 7.51 |
| Frequency | Within Subjects |
|---|---|
| 13–16 Hz | Laterality × group p = 0.034 F(1.89,114.96) = 3.58, np2 = 0.055, power = 0.636 |
| 17–20 Hz | Laterality × group p = 0.025 F(1.88,114.93) = 4.52, np2 = 0.060, power = 0.676 |
| 21–24 Hz | Laterality × group p = 0.021 F(1.89,115.11) = 4.12, np2 = 0.063, power = 0.701 |
| 25–28 Hz | Laterality × group p = 0.018 F(1.89,115.36) = 4.27, np2 = 0.065, power = 0.719 |
| 29–32 Hz | Laterality × group p = 0.016 F(1.89,115.63) = 4.38, np2 = 0.067, power = 0.731 |
| 33–36 Hz | Laterality × group p = 0.015 F(1.90,115.90) = 4.47, np2 = 0.068, power = 0.741 |
| 37–40 Hz | Laterality × group p = 0.014 F(1.90,116.16) = 4.54, np2 = 0.069, power = 0.748 |
| 41–45 Hz | Laterality × group p = 0.013 F(1.91,116.43) = 4.60, np2 = 0.070, power = 0.755 |
| RM-ANOVA | ||
|---|---|---|
| Frequency | Within Subjects | Between Subjects |
| 1–4 Hz | Laterality × group p = 0.050 F(1.98,120.93) = 3.07, np2 = 0.048, power = 0.581 | - |
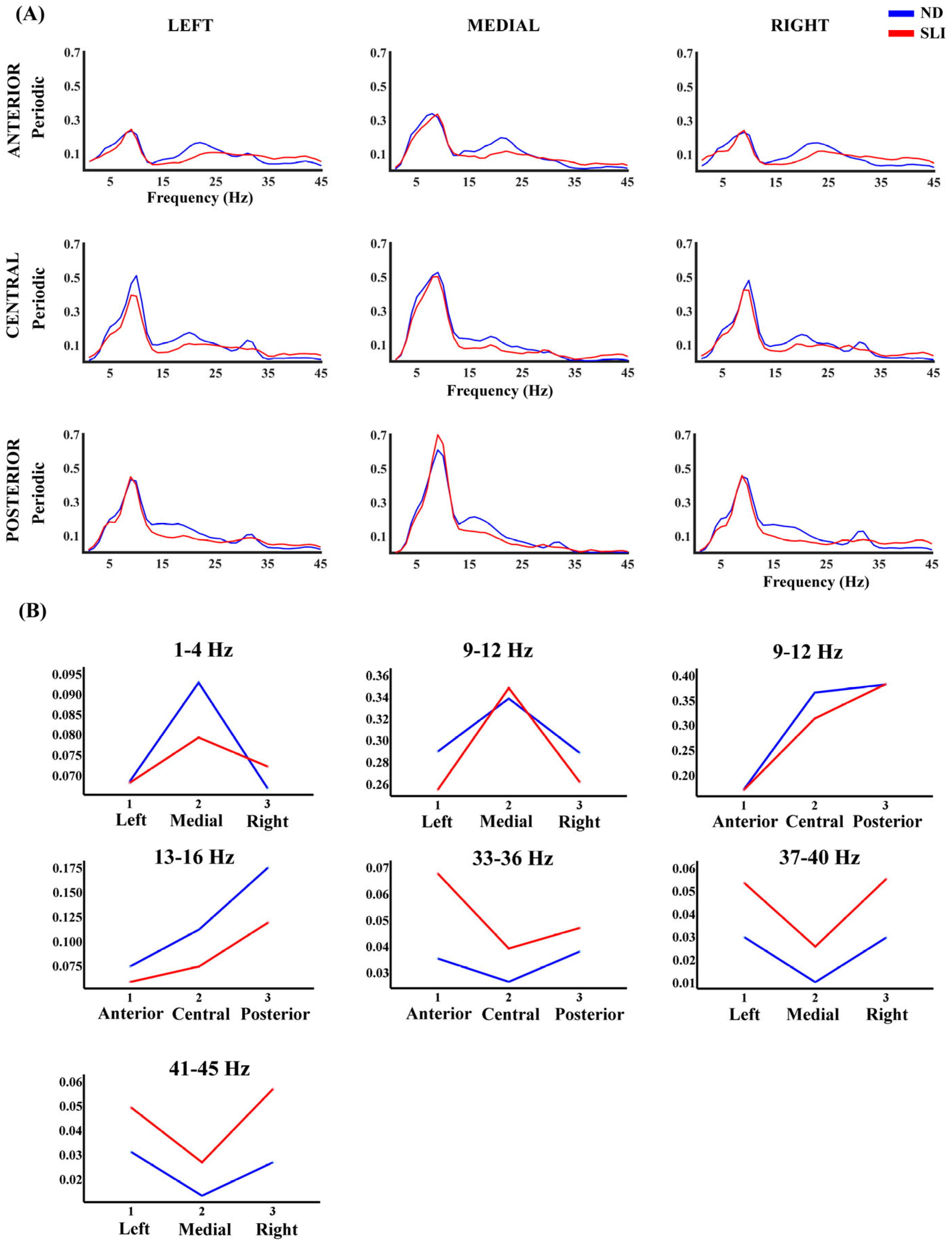
| 9–12 Hz | Laterality × group p = 0.017 F(1.57,95.48) = 4.78, np2 = 0.073, power = 0.710 Antero-posterior × group p = 0.031 F(1.97,120.13) = 3.59, np2 = 0.056, power = 0.650 | - |
| 13–16 Hz | Antero-posterior × group p = 0.030 F(1.89,115.23) = 3.69, np2 = 0.057, power = 0.650 | - |
| 33–36 Hz | Antero-posterior × group p = 0.036 F(1.53,93.23) = 3.84, np2 = 0.059, power = 0.603 | - |
| 37–40 Hz | - | Group p = 0.005 F(1,61) = 8.61, np2 = 0.124, power = 0.823 |
| 41–45 Hz | Laterality × group p = 0.037 F(1.97,120.09) = 3.40, np2 = 0.053, power = 0.626 | Group p = 0.006 F(1,61) = 8.03, np2 = 0.116, power = 0.796 |
| Aperiodic | ||
|---|---|---|
| Frequency Range | Between Subjects | Within Subjects |
| 13–16 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND) |
| 17–20 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND) |
| 21–24 Hz | Left–Medial (SLI > ND) Right–Medial (SLI > ND) | |
| 25–28 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND) |
| 29–32 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND) |
| 33–36 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND)) |
| 37–40 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND) |
| 41–45 Hz | - | Left–Medial (SLI > ND) Right–Medial (SLI > ND) |
| Periodic | ||
| 9–12 Hz | - | Central–Anterior (ND>SLI) Posterior–Central (SLI > ND) |
| 33–36 Hz | - | Anterior (SLI > ND) |
| 37–40 Hz | SLI > ND | - |
| 41–45 Hz | SLI > ND | Left (SLI > ND) Medial (SLI > ND) Right (SLI > ND) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angulo-Ruiz, B.Y.; Rodríguez-Martínez, E.I.; Ruiz-Martínez, F.J.; Gómez-Treviño, A.; Muñoz, V.; Andalia Crespo, S.; Gómez, C.M. Brain Complexity and Parametrization of Power Spectral Density in Children with Specific Language Impairment. Entropy 2025, 27, 572. https://doi.org/10.3390/e27060572
Angulo-Ruiz BY, Rodríguez-Martínez EI, Ruiz-Martínez FJ, Gómez-Treviño A, Muñoz V, Andalia Crespo S, Gómez CM. Brain Complexity and Parametrization of Power Spectral Density in Children with Specific Language Impairment. Entropy. 2025; 27(6):572. https://doi.org/10.3390/e27060572
Chicago/Turabian StyleAngulo-Ruiz, Brenda Y., Elena I. Rodríguez-Martínez, Francisco J. Ruiz-Martínez, Ana Gómez-Treviño, Vanesa Muñoz, Sheyla Andalia Crespo, and Carlos M. Gómez. 2025. "Brain Complexity and Parametrization of Power Spectral Density in Children with Specific Language Impairment" Entropy 27, no. 6: 572. https://doi.org/10.3390/e27060572
APA StyleAngulo-Ruiz, B. Y., Rodríguez-Martínez, E. I., Ruiz-Martínez, F. J., Gómez-Treviño, A., Muñoz, V., Andalia Crespo, S., & Gómez, C. M. (2025). Brain Complexity and Parametrization of Power Spectral Density in Children with Specific Language Impairment. Entropy, 27(6), 572. https://doi.org/10.3390/e27060572