
Targeting the Renin–Angiotensin–Aldosterone System to Prevent Hypertension and Kidney Disease of Developmental Origins



Abstract
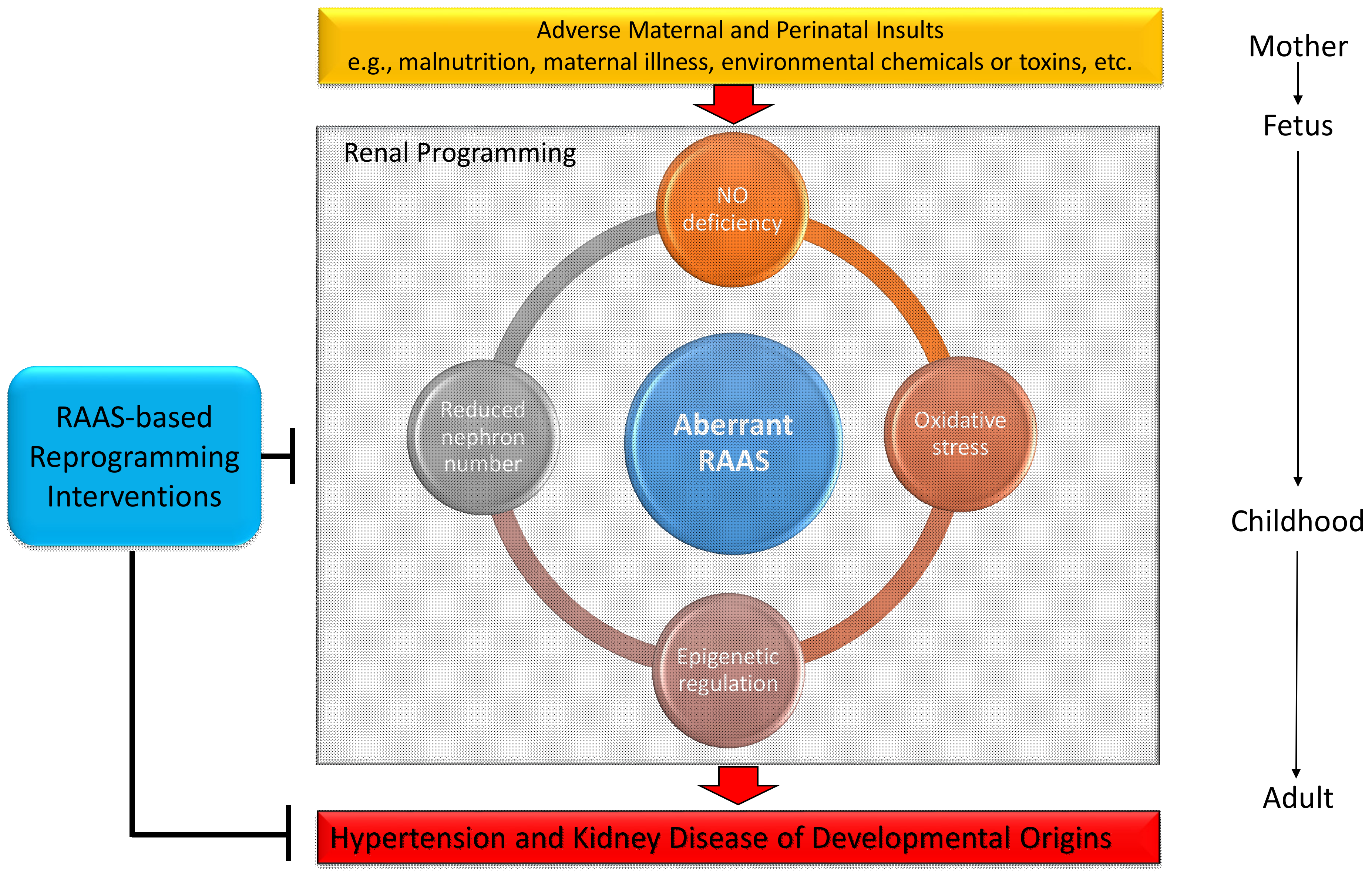
1. Introduction
2. RAAS and the Programmed Kidney
2.1. Intrarenal RAAS
2.2. The Programmed Kidney: Cause for Adult Hypertension and Kindey Disease?
2.3. Impact of RAAS in Renal Programming
3. Animal Models of Renal Programming: Impact of the RAAS
3.1. RAAS-Related Renal Programming in Animal Models
3.2. Renin, (Pro)renin, and Their Receptor in Renal Programming
3.3. Classical RAAS Axis in Renal Programming
3.4. Non-Classical RAAS Axis in Renal Programming
3.5. Aldosterone in Renal Programming
4. The Central Role of the RAAS on Mediating Common Mechanisms Underlying Renal Programming
4.1. Oxidative Stress
4.2. Nitric Oxide Deficiency
4.3. Reduced Nephron Number
4.4. Epigenetic Regulation
4.5. Others
5. Targeting on the RAAS as Reprogramming Strategies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- World Health Organization. Hypertension. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/hypertension (accessed on 12 January 2021).
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Weir, M.R. Hypertension and the kidney: Perspectives on the relationship of kidney disease and cardiovascular disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, S.M.; Textor, S.C. Renovascular Hypertension. Endocrinol. Metab. Clin. N. Am. 2019, 48, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Basso, N.; Terragno, N.A. History about the discovery of the renin-angiotensin system. Hypertension 2001, 38, 1246–1249. [Google Scholar] [CrossRef]
- Zandi-Nejad, K.; Luyckx, V.A.; Brenner, B.M. Adult hypertension and kidney disease: The role of fetal programming. Hypertension 2006, 47, 502–508. [Google Scholar] [CrossRef]
- Chong, E.; Yosypiv, I.V. Developmental programming of hypertension and kidney disease. Int. J. Nephrol. 2012, 2012, 760580. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Developmental origins of chronic kidney disease: Should we focus on early life? Int. J. Mol. Sci. 2017, 18, 381. [Google Scholar] [CrossRef]
- Kett, M.M.; Denton, K.M. Renal programming: Cause for concern? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R791–R803. [Google Scholar] [CrossRef] [PubMed]
- Paixão, A.D.; Alexander, B.T. How the kidney is impacted by the perinatal maternal environment to develop hypertension. Biol. Reprod. 2013, 89, 144. [Google Scholar] [CrossRef]
- Barker, D.J. Fetal programming of coronary heart disease. Trends. Endocrinol. Metab. 2002, 13, 364–368. [Google Scholar] [CrossRef]
- Haugen, A.C.; Schug, T.T.; Collman, G.; Heindel, J.J. Evolution of DOHaD: The impact of environmental health sciences. J. Dev. Orig. Health Dis. 2015, 6, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Joles, J.A. Reprogramming: A preventive strategy in hypertension focusing on the kidney. Int. J. Mol. Sci. 2016, 17, 23. [Google Scholar] [CrossRef]
- Paauw, N.D.; van Rijn, B.B.; Lely, A.T.; Joles, J.A. Pregnancy as a critical window for blood pressure regulation in mother and child: Programming and reprogramming. Acta Physiol. 2017, 219, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.M.; Borkowski, K.R.; Leenen, F.H.; Tsoporis, J.; Coughlin, M. Combined effect of neonatal sympathectomy and adrenal demedullation on blood pressure and vascular changes in spontaneously hypertensive rats. Circ. Res. 1991, 69, 714–721. [Google Scholar] [CrossRef]
- Vavřínová, A.; Behuliak, M.; Bencze, M.; Vodička, M.; Ergang, P.; Vaněčková, I.; Zicha, J. Sympathectomy-induced blood pressure reduction in adult normotensive and hypertensive rats is counteracted by enhanced cardiovascular sensitivity to vasoconstrictors. Hypertens. Res. 2019, 42, 1872–1882. [Google Scholar] [CrossRef] [PubMed]
- Paz Ocaranza, M.; Riquelme, J.A.; García, L.; Jalil, J.E.; Chiong, M.; Santos, R.A.S.; Lavandero, S. Counter-regulatory renin-angiotensin system in cardiovascular disease. Nat. Rev. Cardiol. 2020, 17, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Bagby, S.P. Maternal nutrition, low nephron number, and hypertension in later life: Pathways of nutritional programming. J. Nutr. 2007, 137, 1066–1072. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Regulation of nitric oxide production in the developmental programming of hypertension and kidney disease. Int. J. Mol. Sci. 2019, 20, 681. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Early-life programming and reprogramming of adult kidney disease and hypertension: The interplay between maternal nutrition and oxidative stress. Int. J. Mol. Sci. 2020, 21, 3572. [Google Scholar] [CrossRef]
- Te Riet, L.; van Esch, J.H.; Roks, A.J.; van den Meiracker, A.H.; Danser, A.H. Hypertension: Renin-angiotensin-aldosterone system alterations. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef] [PubMed]
- Bogdarina, I.; Welham, S.; King, P.J.; Burns, S.P.; Clark, A.J. Epigenetic modification of the renin-angiotensin system in the fetal programming of hypertension. Circ. Res. 2007, 100, 520–526. [Google Scholar] [CrossRef]
- Chappell, M.C.; Marshall, A.C.; Alzayadneh, E.M.; Shaltout, H.A.; Diz, D.I. Update on the Angiotensin converting enzyme 2-Angiotensin (1-7)-MAS receptor axis: Fetal programing, sex differences, and intracellular pathways. Front. Endocrinol. 2014, 4, 201. [Google Scholar] [CrossRef] [PubMed]
- Cravedi, P.; Ruggenenti, P.; Remuzzi, G. Which antihypertensive drugs are the most nephroprotective and why? Expert Opin. Pharmacother. 2010, 11, 2651–2663. [Google Scholar] [CrossRef]
- Wilson, B.A.; Marshall, A.C.; Alzayadneh, E.M.; Chappell, M.C. The ins and outs of angiotensin processing within the kidney. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R487–R489. [Google Scholar] [CrossRef]
- Stanton, A. Potential of renin inhibition in cardiovascular disease. J. Renin-Angiotensin-Aldosterone Syst. 2003, 4, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.Z.; Xiao, H.D.; Li, P.; Billet, S.; Lin, C.X.; Fuchs, S.; Bernstein, K.E. Tissue specific expression of angiotensin converting enzyme: A new way to study an old friend. Int. Immunopharmacol. 2008, 8, 171–176. [Google Scholar] [CrossRef][Green Version]
- Navar, L.G.; Kobori, H.; Prieto, M.C.; Gonzalez-Villalobos, R.A. Intrarenal reninangiotensin system in hypertension. Hypertension 2011, 57, 355–362. [Google Scholar] [CrossRef]
- Schwacke, J.H.; Spainhour, J.C.; Ierardi, J.L.; Chaves, J.M.; Arthur, J.M.; Janech, M.G.; Velez, J.C. Network modeling reveals steps in angiotensin peptide processing. Hypertension 2013, 61, 690–700. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Donoghue, M.; Hsieh, F.; Baronas, E.; Godbout, K.; Gosselin, M.; Stagliano, N.; Donovan, M.; Woolf, B.; Robison, K.; Jeyaseelan, R.; et al. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9. Circ. Res. 2000, 87, E1–E9. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.F.; Douglas-Denton, R.N.; Diouf, B.; Hughson, M.D.; Hoy, W.E. Human nephron number: Implications for health and disease. Pediatr. Nephrol. 2011, 26, 1529–1533. [Google Scholar] [CrossRef]
- Rosenblum, S.; Pal, A.; Reidy, K. Renal development in the fetus and premature infant. Semin. Fetal Neonatal Med. 2017, 22, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.; Sampogna, R.V.; Sakurai, H.; Bush, K.T.; Nigam, S.K. Branching morphogenesis and kidney disease. Development 2004, 131, 1449–1462. [Google Scholar] [CrossRef]
- Sainio, K.; Suvanto, P.; Davies, J.; Wartiovaara, J.; Wartiovaara, K.; Saarma, M.; Arumae, U.; Meng, X.; Lindahl, M.; Pachnis, V.; et al. Glial-cell-line-derived neurotrophic factor is required for bud initiation from ureteric epithelium. Development 1997, 124, 4077–4087. [Google Scholar]
- Roseboom, T.; de Rooij, S.; Painter, R. The Dutch famine and its long-term consequences for adult health. Early Hum. Dev. 2006, 82, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.C.; Roseboom, T.J.; van Montfrans, G.A.; Bossuyt, P.M.; Krediet, R.T.; Osmond, C.; Barker, D.J.; Bleker, O.P. Microalbuminuria in adults after prenatal exposure to the Dutch famine. J. Am. Soc. Nephrol. 2005, 16, 189–194. [Google Scholar] [CrossRef]
- White, S.L.; Perkovic, V.; Cass, A.; Chang, C.L.; Poulter, N.R.; Spector, T.; Haysom, L.; Craig, J.C.; Salmi, I.A.; Chadban, S.J.; et al. Is low birth weight an antecedent of CKD in later life? A systematic review of observational studies. Am. J. Kidney Dis. 2009, 54, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Luh, H.; Lin, C.Y.; Hsu, C.N. Incidence and risks of congenital anomalies of kidney and urinary tract in newborns: A population-based case-control study in Taiwan. Medicine 2016, 95, e2659. [Google Scholar] [CrossRef] [PubMed]
- De Jong, F.; Monuteaux, M.C.; van Elburg, R.M.; Gillman, M.W.; Belfort, M.B. Systematic review and meta-analysis of preterm birth and later systolic blood pressure. Hypertension 2012, 59, 226–234. [Google Scholar] [CrossRef]
- Luyckx, V.A.; Brenner, B.M. The clinical importance of nephron mass. J. Am. Soc. Nephrol. 2010, 21, 898–910. [Google Scholar] [CrossRef] [PubMed]
- Beeman, S.C.; Cullen-McEwen, L.A.; Puelles, G.; Zhang, M.; Wu, T.; Baldelomar, E.J.; Dowling, J.; Charlton, J.R.; Forbes, M.S.; Ng, A.; et al. MRI-based glomerular morphology and pathology in whole human kidneys. Am. J. Physiol. Ren. Physiol. 2014, 306, F1381–F1390. [Google Scholar] [CrossRef]
- Gubler, M.C.; Antignac, C. Renin-angiotensin system in kidney development: Renal tubular dysgenesis. Kidney Int. 2010, 77, 400–406. [Google Scholar] [CrossRef]
- Yosypiv, I.V. Renin-angiotensin system in ureteric bud branching morphogenesis: Insights into the mechanisms. Pediatr. Nephrol. 2011, 26, 1499–1512. [Google Scholar] [CrossRef]
- Schreuder, M.F.; Bueters, R.R.; Huigen, M.C.; Russel, F.G.; Masereeuw, R.; van den Heuvel, L.P. Effect of drugs on renal development. Clin. J. Am. Soc. Nephrol. 2011, 6, 212–217. [Google Scholar] [CrossRef]
- Lee, M.A.; Bohm, M.; Paul, M.; Ganten, D. Tissue renin-angiotensin systems. Their role in cardiovascular disease. Circulation 1993, 87, IV7–IV13. [Google Scholar] [PubMed]
- Okubo, S.; Niimura, F.; Matsusaka, T.; Fogo, A.; Hogan, B.L.; Ichikawa, I. Angiotensinogen gene null-mutant mice lack homeostatic regulation of glomerular filtration and tubular reabsorption. Kidney Int. 1998, 53, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, S.; Matsusaka, T.; Chen, X.; Okubo, S.; Niimura, F.; Nishimura, H.; Fogo, A.; Utsunomiya, H.; Inagami, T.; Ichikawa, I. Murine double nullizygotes of the angiotensin type 1A and 1B receptor genes duplicate severe abnormal phenotypes of angiotensinogen nullizygotes. J. Clin. Investig. 1998, 101, 755–760. [Google Scholar] [CrossRef]
- Mullins, J.J.; Peters, J.; Ganten, D. Fulminant hypertension in transgenic rats harbouring the mouse Ren-2 gene. Nature 1990, 344, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Woods, L.L.; Rasch, R. Perinatal ANG II programs adult blood pressure, glomerular number and renal function in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 275, R1593–R1599. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Shi, A.; Zhu, D.; Bo, L.; Zhong, Y.; Wang, J.; Xu, Z.; Mao, C. High sucrose intake during gestation increases angiotensin II type 1 receptor-mediated vascular contractility associated with epigenetic alterations in aged offspring rats. Peptides 2016, 86, 133–144. [Google Scholar] [CrossRef]
- Hsu, C.N.; Wu, K.L.; Lee, W.C.; Leu, S.; Chan, J.Y.; Tain, Y.L. Aliskiren Administration during Early Postnatal Life Sex-Specifically Alleviates Hypertension Programmed by Maternal High Fructose Consumption. Front. Physiol. 2016, 7, 299. [Google Scholar] [CrossRef]
- Tain, Y.L.; Wu, K.L.; Lee, W.C.; Leu, S.; Chan, J.Y. Maternal fructose-intake-induced renal programming in adult male offspring. J. Nutr. Biochem. 2015, 26, 642–650. [Google Scholar] [CrossRef]
- Sahajpal, V.; Ashton, N. Increased glomerular angiotensin II binding in rats exposed to a maternal low protein diet in utero. J. Physiol. 2005, 563, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Sahajpal, V.; Ashton, N. Renal function and angiotensin AT1 receptor expression in young rats following intrauterine exposure to a maternal low-protein diet. Clin. Sci.(Lond.) 2003, 104, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Vehaskari, V.M.; Stewart, T.; Lafont, D.; Soyez, C.; Seth, D.; Manning, J. Kidney angiotensin and angiotensin receptor expression in prenatally programmed hypertension. Am. J. Physiol. Ren. Physiol. 2004, 287, F262–F267. [Google Scholar] [CrossRef] [PubMed]
- Vehaskari, V.M.; Aviles, D.H.; Manning, J. Prenatal programming of adult hypertension in the rat. Kidney Int. 2001, 59, 238–245. [Google Scholar] [CrossRef]
- Luzardo, R.; Silva, P.A.; Einicker-Lamas, M.; Ortiz-Costa, S.; do Carmo Mda, G.; Vieira-Filho, L.D.; Paixão, A.D.; Lara, L.S.; Vieyra, A. Metabolic programming during lactation stimulates renal Na+ transport in the adult offspring due to an early impact on local angiotensin II pathways. PLoS ONE 2011, 6, e21232. [Google Scholar] [CrossRef]
- Woods, L.L.; Ingelfinger, J.R.; Nyengaard, J.R.; Rasch, R. Maternal protein restriction suppresses the newborn renin-angiotensin system and programs adult hypertension in rats. Pediatr. Res. 2001, 49, 460–467. [Google Scholar] [CrossRef]
- Woods, L.L.; Weeks, D.A.; Rasch, R. Programming of adult blood pressure by maternal protein restriction: Role of nephrogenesis. Kidney Int. 2004, 65, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.; Van-Wickle, J.; Goyal, D.; Longo, L.D. Antenatal maternal low protein diet: ACE-2 in the mouse lung and sexually dimorphic programming of hypertension. BMC Physiol. 2015, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.; Vehaskari, V.M. Low birth weight-associated adult hypertension in the rat. Pediatr. Nephrol. 2001, 16, 417–422. [Google Scholar] [CrossRef]
- Manning, J.; Vehaskari, V.M. Postnatal modulation of prenatally programmed hypertension by dietary Na and ACE inhibition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R80–R84. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.S.; Lang, A.L.; Grant, A.R.; Nijland, M.J. Maternal nutrient restriction in sheep: Hypertension and decreased nephron number in offspring at 9 months of age. J. Physiol. 2005, 565, 137–147. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Chan, J.Y.H.; Lee, C.T.; Tain, Y.L. Hypertension Programmed by Perinatal High-Fat Diet: Effect of Maternal Gut Microbiota-Targeted Therapy. Nutrients 2019, 11, 2908. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lin, Y.J.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Chen, C.C.; Tsai, C.C.; Huang, L.T.; Hsu, C.N. High Fat Diets Sex-Specifically Affect the Renal Transcriptome and Program Obesity, Kidney Injury, and Hypertension in the Offspring. Nutrients 2017, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.A.; de Noronha, I.L.; de Oliveira, I.B.; Malheiros, D.M.; Heimann, J.C. Renin-angiotensin system function and blood pressure in adult rats after perinatal salt overload. Nutr. Metab. Cardiovasc. Dis. 2003, 13, 133–139. [Google Scholar] [PubMed]
- Battista, M.C.; Oligny, L.L.; St-Louis, J.; Brochu, M. Intrauterine growth restriction in rats is associated with hypertension and renal dysfunction in adulthood. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E124–E131. [Google Scholar] [CrossRef]
- Guo, Q.; Feng, X.; Xue, H.; Teng, X.; Jin, S.; Duan, X.; Xiao, L.; Wu, Y. Maternal renovascular hypertensive rats treatment with hydrogen sulfide increased the methylation of AT1b gene in offspring. Am. J. Hypertens. 2017, 30, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Denton, K.M.; Flower, R.L.; Stevenson, K.M.; Anderson, W.P. Adult rabbit offspring of mothers with secondary hypertension have increased blood pressure. Hypertension 2003, 41, 634–639. [Google Scholar] [CrossRef][Green Version]
- Hsu, C.N.; Yang, H.W.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal Adenine-Induced Chronic Kidney Disease Programs Hypertension in Adult Male Rat Offspring: Implications of Nitric Oxide and Gut Microbiome Derived Metabolites. Int. J. Mol. Sci. 2020, 21, 7237. [Google Scholar] [CrossRef]
- Chen, Y.W.; Chenier, I.; Tran, S.; Scotcher, M.; Chang, S.Y.; Zhang, S.L. Maternal diabetes programs hypertension and kidney injury in offspring. Pediatr. Nephrol. 2010, 25, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Wichi, R.B.; Souza, S.B.; Casarini, D.E.; Morris, M.; Barreto-Chaves, M.L.; Irigoyen, M.C. Increased blood pressure in the offspring of diabetic mothers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1129–R1133. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lin, Y.J.; Chan, J.Y.H.; Lee, C.T.; Hsu, C.N. Maternal melatonin or agomelatine therapy prevents programmed hypertension in male offspring of mother exposed to continuous light. Biol. Reprod. 2017, 97, 636–643. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and renal transcriptome to prevent prenatal NG-Nitro-L-arginine-methyl ester (L-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obstet. Gynecol. 2016, 215, 636.e1–636.e72. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.Q.; Zhang, H.G.; Yuan, Z.B.; Yang, D.L.; Hao, L.Y.; Li, X.H. Prenatal exposure to lipopolysaccharide alters the intrarenal renin-angiotensin system and renal damage in offspring rats. Hypertens. Res. 2010, 33, 76–82. [Google Scholar] [CrossRef]
- Grigore, D.; Ojeda, N.B.; Robertson, E.B.; Dawson, A.S.; Huffman, C.A.; Bourassa, E.A.; Speth, R.C.; Brosnihan, K.B.; Alexander, B.T. Placental insufficiency results in temporal alterations in the renin angiotensin system in male hypertensive growth restricted offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R804–R811. [Google Scholar] [CrossRef] [PubMed]
- Walton, S.L.; Bielefeldt-Ohmann, H.; Singh, R.R.; Li, J.; Paravicini, T.M.; Little, M.H.; Moritz, K.M. Prenatal hypoxia leads to hypertension, renal renin-angiotensin system activation and exacerbates salt-induced pathology in a sex-specific manner. Sci. Rep. 2017, 7, 8241. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Rodriguez, M.; Loyse, N.; Bourdon, C.; Arab, S.; Pausova, Z. Effect of prenatal exposure to nicotine on kidney glomerular mass and AT1R expression in genetically diverse strains of rats. Toxicol. Lett. 2012, 213, 228–234. [Google Scholar] [CrossRef]
- Xiao, D.; Xu, Z.; Huang, X.; Longo, L.D.; Yang, S.; Zhang, L. Prenatal gender-related nicotine exposure increases blood pressure response to angiotensin II in adult offspring. Hypertension 2008, 51, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Serapiao-Moraes, D.F.; Souza-Mello, V.; Aguila, M.B.; Mandarim-de-Lacerda, C.A.; Faria, T.S. Maternal caffeine administration leads to adverse effects on adult mice offspring. Eur. J. Nutr. 2013, 52, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Lin, Y.J.; Lu, P.C.; Tain, Y.L. Maternal Resveratrol Therapy Protects Male Rat Offspring against Programmed Hypertension Induced by TCDD and Dexamethasone Exposures: Is It Relevant to Aryl Hydrocarbon Receptor? Int. J. Mol. Sci. 2018, 19, 2459. [Google Scholar] [CrossRef] [PubMed]
- Canale, D.; Rodrigues, M.V.; Ferreira, D.N.; Machado, F.G.; Veras, M.M.; Malheiros, D.M.; Krieger, J.E.; Fujihara, C.K.; Venturini, G.; Zatz, R. Programmed hypertension in rats treated with a NF-κB inhibitor during nephrogenesis: Renal mechanisms. Hypertens. Res. 2011, 34, 693–700. [Google Scholar] [CrossRef][Green Version]
- Tain, Y.L.; Sheen, J.M.; Chen, C.C.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Maternal citrulline supplementation prevents prenatal dexamethasone-induced programmed hypertension. Free Radic. Res. 2014, 48, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Chen, C.C.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Melatonin attenuates prenatal dexamethasone-induced blood pressure increase in a rat model. J. Am. Soc. Hypertens. 2014, 8, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Lin, Y.J.; Yu, H.R.; Lin, I.C.; Sheen, J.M.; Huang, L.T.; Tain, Y.L. Protection of Male Rat Offspring against Hypertension Programmed by Prenatal Dexamethasone Administration and Postnatal High-Fat Diet with the Nrf2 Activator Dimethyl Fumarate during Pregnancy. Int. J. Mol. Sci. 2019, 20, 3957. [Google Scholar] [CrossRef]
- Wu, T.H.; Kuo, H.C.; Lin, I.C.; Chien, S.J.; Huang, L.T.; Tain, Y.L. Melatonin prevents neonatal dexamethasone induced programmed hypertension: Histone deacetylase inhibition. J. Steroid Biochem. Mol. Biol. 2014, 144, 253–259. [Google Scholar] [CrossRef]
- Shaltout, H.A.; Figueroa, J.P.; Rose, J.C.; Diz, D.I.; Chappell, M.C. Alterations in circulatory and renal angiotensin-converting enzyme and angiotensin-converting enzyme 2 in fetal programmed hypertension. Hypertension 2009, 53, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Hartman, H.A.; Lai, H.L.; Patterson, L.T. Cessation of renal morphogenesis in mice. Dev. Biol. 2007, 310, 379–387. [Google Scholar] [CrossRef]
- Sengupta, P. The Laboratory Rat: Relating Its Age with Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Song, R.; Yosypiv, I.V. (Pro)renin Receptor in Kidney Development and Disease. Int. J. Nephrol. 2011, 2011, 247048. [Google Scholar] [CrossRef]
- Sanoski, C.A. Aliskiren: An oral direct renin inhibitor for the treatment of hypertension. Pharmacotherapy 2009, 29, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G. Renin, (pro)renin and receptor: An update. Clin. Sci. 2011, 120, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Krop, M.; Lu, X.; Danser, A.H.; Meima, M.E. The (pro)renin receptor. A decade of research: What have we learned? Pflüg. Arch. 2013, 465, 87–97. [Google Scholar] [CrossRef]
- Mercure, C.; Prescott, G.; Lacombe, M.J.; Silversides, D.W.; Reudelhuber, T.L. Chronic increases in circulating prorenin are not associated with renal or cardiac pathologies. Hypertension 2009, 53, 1062–1069. [Google Scholar] [CrossRef][Green Version]
- Riediger, F.; Quack, I.; Qadri, F.; Hartleben, B.; Park, J.K.; Potthoff, S.A.; Sohn, D.; Sihn, G.; Rousselle, A.; Fokuhl, V.; et al. Prorenin receptor is essential for podocyte autophagy and survival. J. Am. Soc. Nephrol. 2011, 22, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.M.; Tain, Y.L.; Leu, S.; Wu, K.L.; Lee, W.C.; Chan, J.Y. Developmental programming of the metabolic syndrome: Next-generation sequencing analysis of transcriptome expression in a rat model of maternal high fructose intake. Sheng Li Xue Bao 2016, 68, 557–567. [Google Scholar]
- Tain, Y.L.; Huang, L.T.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal citrulline supplementation prevents prenatal NG-nitro-L-arginine-methyl ester (L-NAME)-induced programmed hypertension in rats. Biol. Reprod. 2015, 92, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Seong, H.Y.; Cho, H.M.; Kim, M.; Kim, I. Maternal High-Fructose Intake Induces Multigenerational Activation of the Renin-Angiotensin-Aldosterone System. Hypertension 2019, 74, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Zhang, F.; Luo, Y.; Wang, L.; Huang, S.; Jin, F. Serum and Glucocorticoid Regulated Kinase 1 in Sodium Homeostasis. Int. J. Mol. Sci. 2016, 17, 1307. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, L.A.; Quan, A.; Weinberg, A.; Baum, M. Effect of prenatal dexamethasone on rat renal development. Kidney Int. 2001, 59, 1663–1669. [Google Scholar] [CrossRef]
- Dagan, A.; Kwon, H.M.; Dwarakanath, V.; Baum, M. Effect of renal denervation on prenatal programming of hypertension and renal tubular transporter abundance. Am. J. Physiol. Ren. Physiol. 2008, 295, F29–F34. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, W.C.; Leu, S.; Wu, K.; Chan, J. High salt exacerbates programmed hypertension in maternal fructose-fed male offspring. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 1146–1151. [Google Scholar] [CrossRef]
- Vallon, V.; Lang, F. New insights into the role of serum- and glucocorticoid-inducible kinase SGK1 in the regulation of renal function and blood pressure. Curr. Opin. Nephrol. Hypertens. 2005, 14, 59–66. [Google Scholar] [CrossRef]
- Klein, A.V.; Kiat, H. The mechanisms underlying fructose-induced hypertension: A review. J. Hypertens. 2015, 33, 912–920. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Early Origins of Hypertension: Should Prevention Start Before Birth Using Natural Antioxidants? Antioxidants 2020, 9, 1034. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Developmental Origins of Kidney Disease: Why Oxidative Stress Matters? Antioxidants 2021, 10, 33. [Google Scholar] [CrossRef]
- Thompson, L.P.; Al-Hasan, Y. Impact of oxidative stress in fetal programming. J. Pregnancy 2012, 2012, 582748. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.; Wilcox, C.S. Oxidative stress in hypertension: Role of the kidney. Antioxid. Redox Signal. 2014, 20, 74–101. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, M.; Urushihara, M.; Morikawa, T.; Konishi, Y.; Imanishi, M.; Nishiyama, A.; Kobori, H. Oxidative stress/angiotensinogen/renin-angiotensin system axis in patients with diabetic nephropathy. Int. J. Mol. Sci. 2013, 14, 23045–23062. [Google Scholar] [CrossRef] [PubMed]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; Von Knethen, A. Nrf2, the Master Regulator of Anti-Oxidative Responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef] [PubMed]
- Rush, B.M.; Bondi, C.D.; Stocker, S.D.; Barry, K.M.; Small, S.A.; Ong, J.; Jobbagy, S.; Stolz, D.B.; Bastacky, S.I.; Chartoumpekis, D.V.; et al. Genetic or pharmacologic Nrf2 activation increases proteinuria in chronic kidney disease in mice. Kidney Int. 2021, 99, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Seki, S.; Hiramoto, K.; Naganuma, E.; Kobayashi, E.H.; Yamaoka, A.; Baird, L.; Takahashi, N.; Sato, H.; Yamamoto, M. Hyperactivation of Nrf2 in early tubular development induces nephrogenic diabetes insipidus. Nat. Commun. 2017, 8, 14577. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N.; Lee, C.T. Melatonin therapy prevents programmed hypertension and nitric oxide deficiency in offspring exposed to maternal caloric restriction. Oxid. Med. Cell. Longev. 2014, 2014, 283180. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Targeting on asymmetric dimethylarginine related nitric oxide-reactive oxygen species imbalance to reprogram the development of hypertension. Int. J. Mol. Sci. 2016, 17, 2020. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Toxic Dimethylarginines: Asymmetric Dimethylarginine (ADMA) and Symmetric Dimethylarginine (SDMA). Toxins 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Schulman, I.H.; Zhou, M.S.; Raij, L. Interaction between nitric oxide and angiotensin II in the endothelium: Role in atherosclerosis and hypertension. J. Hypertens. 2006, 24, S45–S50. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N.; Lin, C.Y.; Huang, L.T.; Lau, Y.T. Aliskiren prevents hypertension and reduces asymmetric dimethylarginine in young spontaneously hypertensive rats. Eur. J. Pharmacol. 2011, 670, 561–565. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lee, C.T.; Huang, L.T.; Tain, Y.L. Aliskiren in early postnatal life prevents hypertension and reduces asymmetric dimethylarginine in offspring exposed to maternal caloric restriction. J. Renin-Angiotensin-Aldosterone Syst. 2015, 16, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Murai-Takeda, A.; Kawabe, H.; Itoh, H. Low birth weight trends: Possible impacts on the prevalences of hypertension and chronic kidney disease. Hypertens. Res. 2020, 43, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.A.; Brenner, B.M. Birth weight, malnutrition and kidney-associated outcomes—A global concern. Nat. Rev. Nephrol. 2015, 11, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Starr, M.C.; Hingorani, S.R. Prematurity and future kidney health: The growing risk of chronic kidney disease. Curr. Opin. Pediatr. 2018, 30, 228–235. [Google Scholar] [CrossRef]
- Bianco-Miotto, T.; Craig, J.M.; Gasser, Y.P.; van Dijk, S.J.; Ozanne, S.E. Epigenetics and DOHaD: From basics to birth and beyond. J. Dev. Orig. Health Dis. 2017, 8, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Rees, W.D.; Hay, S.M.; Brown, D.S.; Antipatis, C.; Palmer, R.M. Maternal protein deficiency causes hypermethylation of DNA in the livers of rat fetuses. J. Nutr. 2000, 130, 1821–1826. [Google Scholar] [CrossRef]
- Suter, M.; Ma, J.; Harris, A.; Patterson, L.; Brown, K.A.; Shope, C.; Showalter, L.; Abramovici, A.; Aagaard-Tillery, K.M. Maternal tobacco use modestly alters correlated epigenome-wide placental DNA methylation and gene expression. Epigenetics 2011, 6, 1284–1294. [Google Scholar] [CrossRef]
- Sable, P.; Randhir, K.; Kale, A.; Chavan-Gautam, P.; Joshi, S. Maternal micronutrients and brain global methylation patterns in the offspring. Nutr. Neurosci. 2015, 18, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Liu, Y.; Duan, S.; Perry, B.; Li, W.; He, Y. DNA methylation and hypertension: Emerging evidence and challenges. Brief Funct. Genom. 2016, 15, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Pei, F.; Wang, X.; Yue, R.; Chen, C.; Huang, J.; Huang, J.; Li, X.; Zeng, C. Differential expression and DNA methylation of angiotensin type 1A receptors in vascular tissues during genetic hypertension development. Mol. Cell. Biochem. 2015, 402, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bogdarina, I.G.; King, P.J.; Clark, A.J. Characterization of the angiotensin (AT1b) receptor promoter and its regulation by glucocorticoids. J. Mol. Endocrinol. 2009, 43, 73–80. [Google Scholar] [CrossRef][Green Version]
- Raftopoulos, L.; Katsi, V.; Makris, T.; Tousoulis, D.; Stefanadis, C.; Kallikazaros, I. Epigenetics, the missing link in hypertension. Life Sci. 2015, 129, 22–26. [Google Scholar] [CrossRef]
- Song, R.; Van Buren, T.; Yosypiv, I.V. Histone deacetylases are critical regulators of the renin-angiotensin system during ureteric bud branching morphogenesis. Pediatr. Res. 2010, 67, 573–578. [Google Scholar] [CrossRef]
- Costa, F.F. Non-coding RNAs, epigenetics and complexity. Gene 2008, 410, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- SeneLde, B.; Mesquita, F.F.; deMoraes, L.N.; Santos, D.C.; Carvalho, R.; Gontijo, J.A.; Boer, P.A. Involvement of renal corpuscle microRNA expression on epithelial-to-mesenchymal transition in maternal low protein diet in adult programmed rats. PLoS ONE 2013, 8, e71310. [Google Scholar]
- Goyal, R.; Goyal, D.; Leitzke, A.; Gheorghe, C.P.; Longo, L.D. Brain renin-angiotensin system: Fetal epigenetic programming by maternal protein restriction during pregnancy. Reprod. Sci. 2010, 17, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Interplay between oxidative stress and nutrient sensing signaling in the developmental origins of cardiovascular disease. Int. J. Mol. Sci. 2017, 18, 841. [Google Scholar] [CrossRef]
- Efeyan, A.; Comb, W.C.; Sabatini, D.M. Nutrient-sensing mechanisms and pathways. Nature 2015, 517, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N.; Chan, J.Y. PPARs link early life nutritional insults to later programmed hypertension and metabolic syndrome. Int. J. Mol. Sci. 2015, 17, 20. [Google Scholar] [CrossRef]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Andrade, J.M.; de Farias Lelis, D.; Mafra, V.; Cota, J. The Angiotensin Converting Enzyme 2 (ACE2), Gut Microbiota, and Cardiovascular Health. Protein Pept. Lett. 2017, 24, 827–832. [Google Scholar] [CrossRef]
- Richards, E.M.; Pepine, C.J.; Raizada, M.K.; Kim, S. The Gut, Its Microbiome, and Hypertension. Curr. Hypertens. Rep. 2017, 19, 36. [Google Scholar] [CrossRef]
- Gilbert, J.S.; Nijland, M.J. Sex differences in the developmental origins of hypertension and cardiorenal disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1941–R1952. [Google Scholar] [CrossRef] [PubMed]
- Tomat, A.L.; Salazar, F.J. Mechanisms involved in developmental programming of hypertension and renal diseases. Gender differences. Horm. Mol. Biol. Clin. Investig. 2014, 18, 63–77. [Google Scholar] [CrossRef]
- Hilliard, L.M.; Sampson, A.K.; Brown, R.D.; Denton, K.M. The “his and hers” of the renin-angiotensin system. Curr. Hypertens. Rep. 2013, 15, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Kwekel, J.C.; Desai, V.G.; Moland, C.L.; Vijay, V.; Fuscoe, J.C. Sex differences in kidney gene expression during the life cycle of F344 rats. Biol. Sex Differ. 2013, 4, 14. [Google Scholar] [CrossRef]
- Tain, Y.L.; Wu, M.S.; Lin, Y.J. Sex differences in renal transcriptome and programmed hypertension in offspring exposed to prenatal dexamethasone. Steroids 2016, 115, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Zicha, J.; Dobesová, Z.; Kunes, J. Late blood pressure reduction in SHR subjected to transient captopril treatment in youth: Possible mechanisms. Physiol. Res. 2008, 57, 495–498. [Google Scholar] [PubMed]
- Sherman, R.C.; Langley-Evans, S.C. Antihypertensive treatment in early postnatal life modulates prenatal dietary influences upon blood pressure in the rat. Clin. Sci. (Lond.) 2000, 98, 269–275. [Google Scholar] [CrossRef]
- Mansuri, A.; Elmaghrabi, A.; Legan, S.K.; Gattineni, J.; Baum, M. Transient Exposure of Enalapril Normalizes Prenatal Programming of Hypertension and Urinary Angiotensinogen Excretion. PLoS ONE 2015, 10, e0146183. [Google Scholar] [CrossRef]
- Harrap, S.B.; Nicolaci, J.A.; Doyle, A.E. Persistent effects on blood pressure and renal haemodynamics following chronic angiotensin converting enzyme inhibition with perindopril. Clin. Exp. Pharmacol. Physiol. 1986, 13, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Sherman, R.C.; Langley-Evans, S.C. Early administration of angiotensin-converting enzyme inhibitor captopril, prevents the development of hypertension programmed by intrauterine exposure to a maternal low-protein diet in the rat. Clin. Sci. (Lond.) 1998, 94, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Klimas, J.; Olvedy, M.; Ochodnicka-Mackovicova, K.; Kruzliak, P.; Cacanyiova, S.; Kristek, F.; Krenek, P.; Ochodnicky, P. Perinatally administered losartan augments renal ACE2 expression but not cardiac or renal Mas receptor in spontaneously hypertensive rats. J. Cell. Mol. Med. 2015, 19, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.N.; Lu, D.; Katovich, M.J.; Raizada, M.K. Chronic control of high blood pressure in the spontaneously hypertensive rat by delivery of angiotensin type 1 receptor antisense. Proc. Natl. Acad. Sci. USA 1996, 93, 9960–9965. [Google Scholar] [CrossRef]
- Bessa, A.S.M.; Jesus, É.F.; Nunes, A.D.C.; Pontes, C.N.R.; Lacerda, I.S.; Costa, J.M.; Souza, E.J.; Lino-Júnior, R.S.; Biancardi, M.F.; Dos Santos, F.C.A.; et al. Stimulation of the ACE2/Ang-(1–7)/Mas axis in hypertensive pregnant rats attenuates cardiovascular dysfunction in adult male offspring. Hypertens. Res. 2019, 42, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, A.; Hayashi, M.; Kaneshiro, Y.; Suzuki, F.; Nakagawa, T.; Tada, Y.; Koura, Y.; Nishiyama, A.; Okada, H.; Uddin, M.N.; et al. Inhibition of diabetic nephropathy by a decoy peptide corresponding to the “handle” region for nonproteolytic activation of prorenin. J. Clin. Investig. 2004, 114, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sullivan, M.N.; Zhang, S.; Worker, C.J.; Xiong, Z.; Speth, R.C.; Feng, Y. Intracerebroventricular infusion of the (Pro)renin receptor antagonist PRO20 attenuates deoxycorticosterone acetate-salt-induced hypertension. Hypertension 2015, 65, 352–361. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Animal Models | Intervention Period | Species/Gender | Age at Evaluation | Renal Phenotype | Alterations of the RAAS | Ref. |
|---|---|---|---|---|---|---|
| 20% w/v sucrose in drinking water | Pregnancy | SD rat/M | 90 weeks | ↑BP | ↑AT1R mRNA and protein | [51] |
| High-fructose diet, 60% | Pregnancy and Lactation | SD rat/M | 12 weeks | ↑BP | ↓AT2R mRNA | [52] |
| High-fructose diet, 60% | Pregnancy and Lactation | SD rat/M | 12 weeks | ↑BP, altered renal transcriptome | ↑Renin mRNA | [53] |
| Protein restriction, 9% | Pregnancy | SD rat/M | 4 weeks | ↑BP, ↔GFR | ↑AT1R protein and ↓AT2R protein, ↔ANG level | [54,55] |
| Protein restriction, 6% | Pregnancy | SD rat/M and F | 4 weeks | ↑BP | ↓AT1R and AT2R protein at birth; ↑AT1R and AT2R protein at 4 wk | [56] |
| Protein restriction, 6% | Pregnancy | SD rat/M and F | 8 weeks | ↑BP, ↓nephron number | ↓PRA, ↓AT1R mRNA and protein, ↑Aldosterone | [57] |
| Protein restriction, 8% | Lactation | Wistar rat/M | 150 days | ↑BP, ↑GFR, ↑Proteinuria | ↑AT1R protein and↓AT2R protein | [58] |
| Protein restriction, 8.5% | Pregnancy | SD rat/M | 22 weeks | ↑BP, ↔GFR | ↓renin mRNA and protein; and ↓renal ANG II level at 1–5 days of age | [59,60] |
| Protein restriction, 9% | 1 week before conception and throughout pregnancy | FVB/NJ mouse/F | 24 weeks | ↑BP | ↓ACE2 protein | [61] |
| Protein restriction | Second half of pregnancy | SD and Wistar rat/M and F | 11 months | ↑BP | ↑PRA, ↑AT1R mRNA and protein | [62,63] |
| 50% caloric restriction | Day 28 to day 78 of gestation | Sheep/M and F | 9 months | ↑BP | ↑ACE protein | [64] |
| High-fat diet, 58% | Pregnancy and Lactation | SD rat/M | 16 weeks | ↑BP | ↑AGT and ACE mRNA, and↑AT1R protein | [65] |
| High-fat diet, 58% | 5 weeks before the delivery and throughout pregnancy and lactation | SD rat/M and F | 6 months | ↔BP, ↓GFR, ↑glomerular injury, ↑tubulointerstitial injury, altered renal transcriptome | ↑ACE and AT1R mRNA in F | [66] |
| High-salt diet, 8% | Pregnancy | Wistar rat/F | 12 weeks | ↑BP | ↑ANG II | [67] |
| 0.03% low-salt diet | Last 7 days of pregnancy | SD rat/M and F | 12 weeks | ↑BP | ↑PRA | [68] |
| Maternal renovascular hypertension | Pregnancy | SD rat/M | 16 weeks | ↑BP | ↑AT1R protein | [69] |
| Maternal renovascular hypertension | Pregnancy | Rabbit/F | 30 weeks | ↑BP | ↓PRA at 10 week | [70] |
| Maternal adenine-induced CKD | Pregnancy and lactation | SD rat/M | 12 weeks | ↑BP, renal hypertrophy | ↓AT2R and MAS receptor mRNA | [71] |
| Maternal streptozotocin-induced diabetes | Pregnancy | C57BL/6 mouse/M | 20 weeks | ↑BP, microalbuminuria | ↑AT1R and ACE mRNA, ↓ACE2 mRNA | [72] |
| Maternal streptozotocin-induced diabetes | Pregnancy | Wistar rat/M | 2 months | ↑BP | ↑ACE activity | [73] |
| Continuous light exposure | Pregnancy and lactation | SD rat/M | 12 weeks | ↑BP | ↑Renin, PRR, AGT, ACE, ACE2, and AT1R mRNA | [74] |
| Maternal L-NAME exposure | Pregnancy | SD rat/M | 12 weeks | ↑BP | ↑Renin and ACE mRNA | [75] |
| Maternal LPS exposure | Pregnancy | SD rat/M and F | 24 weeks | ↑BP, ↓nephron number and GFR | ↑ACE mRNA | [76] |
| Placenta insufficiency | Pregnancy | SD rat/M | 16 weeks | ↑BP | ↓Renin and AGT mRNA at birth, ↑Renin and AGT mRNA, ↑ACE activity at 16 week | [77] |
| Prenatal hypoxia | From embryonic day 14.5 until birth | CD1 mouse/M and F | 12 months | Microalbuminuria, glomerular hypertrophy and renal fibrosis, ↓nephron number | ↑Renin and AT1R mRNA | [78] |
| Maternal nicotine exposure | Pregnancy | SHR/M | 9 weeks | ↑BP, ↓Glomerular mass | ↑AT1R mRNA | [79] |
| Maternal nicotine exposure | Pregnancy | SD rat/M | 5 months | ↑BP | ↑AT1R protein, ↓AT2R protein | [80] |
| Maternal caffeine exposure | Pregnancy | C57BL/6 mouse/M | 3 months | ↑BP | ↑Renin and AT1R mRNA | [81] |
| Maternal TCDD and dexamethasone exposure | Pregnancy and Lactation | SD rat/M | 16 weeks | ↑BP | ↑ACE mRNA | [82] |
| Neonatal PDTC administration | Lactation | Munich-Wistar rat/M | 10 months | ↑BP | ↑Renin and AGT mRNA at 3 month; ↓Renin and AGT at 10 month | [83] |
| Prenstal DEX exposure | Pregnancy | SD rat/M | 16 weeks | ↑BP | ↑AGT and AT1R mRNA | [84] |
| Prenstal DEX exposure | Pregnancy | SD rat/M | 16 weeks | ↑BP | ↑Renin and PRR mRNA | [85] |
| Prenatal DEX plus post-weaning high-fat diet | Pregnancy | SD rat/M | 16 weeks | ↑BP | ↑Renin and ACE mRNA | [86] |
| Neonatal DEX administration | Day 1 to day 3 after birth | SD rat/M | 16 weeks | ↑BP | ↓AGT, ACE, and ACE2 mRNA | [87] |
| Prenatal betamethasone exposure | 2 doses, 24 h apart at gestational day 80 | Sheep/M | 1.8 years | ↑BP | ↑ACE activity, ↓ACE2 activity | [88] |
| Intervention | Animal Model | Species/Gender | Age at Evaluation | Effects | Protective Mechanism | Ref. |
|---|---|---|---|---|---|---|
| Renin inhibitor | ||||||
| Aliskiren (10 or 30 mg/kg/day) between 4–10 weeks of age | Genetic hypertension model | SHR/M | 10 weeks | Prevented or attenuated hypertension by 30 or 10 mg, respectively | Restoration of NO bioavailability | [118] |
| Aliskiren (10 mg/kg/day) between 2–4 weeks of age | Maternal 50% caloric restriction | SD rat/M | 12 weeks | Prevented hypertension | Decreased renal AGT mRNA; Increased renal ACE2 and MAS receptor protein levels | [119] |
| Aliskiren (10 mg/kg/day) between 2–4 weeks of age | Maternal high-fructose diet | SD rat/M and F | 12 weeks | Prevented hypertension in both sexes | Increased renal ACE2 and MAS receptor protein levels in F | [52] |
| ACEI | ||||||
| Captopril (100 mg/kg/day) between 2–4 weeks of age | Maternal protein restriction | Wistar rat/M | 12 weeks | Prevented hypertension | Not evaluated | [63] |
| Captopril (100 mg/kg b.w./day) between 4–10 weeks of age | Genetic hypertension model | SHR/M | 30 weeks | Attenuated hypertension | Not evaluated | [147] |
| Enalapril (100 mg/L) in drinking water between 3–6 weeks of age | Maternal protein restriction | SD rat/M | 16 weeks | Prevented hypertension | Not evaluated | [148] |
| Enalapril (100 mg/L) in drinking water between 3–6 weeks of age | Maternal protein restriction | SD rat/M | 6 months | Prevented hypertension and albuminuria | Reduced urinary AGT and ANG II levels | [149] |
| Perindopril (3 mg/kg/day) between 4–16 weeks of age | Genetic hypertension model | SHR/M | 28 weeks | Attenuated hypertension and renal dysfunction | Not evaluated | [150] |
| ARB | ||||||
| Losartan (100 mg/L) in drinking water between 2–4 weeks of age | Maternal protein restriction | Wistar rat/M | 12 weeks | Prevented hypertension | Not evaluated | [151] |
| Losartan (20 mg/kg/day) between 2–4 weeks of age | Maternal 50% caloric restriction | SD rat/M | 12 weeks | Prevented hypertension | Decreased renal AGT mRNA | [118] |
| Losartan (20 mg/kg/day) between 4–9 weeks of age | Genetic hypertension model | SHR/M | 9 weeks | Prevented hypertension | Increased renal ACE2 expression | [152] |
| AT1R antisense | ||||||
| AT1R antisense delivery at 5 days of age | Genetic hypertension model | SHR/M | 3 months | Prevented hypertension | Decreased AT1R mRNA | [153] |
| ACE2 activator | ||||||
| Diminazene aceturate in pregnancy | Maternal hypertension | SHR/M | 16 weeks | Attenuated hypertension and renal fibrosis | Not evaluated in the kidney | [154] |
| ANG-(1-7) in pregnancy | Maternal hypertension | SHR/M | 16 weeks | Attenuated hypertension and renal fibrosis | Not evaluated in the kidney | [154] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-N.; Tain, Y.-L. Targeting the Renin–Angiotensin–Aldosterone System to Prevent Hypertension and Kidney Disease of Developmental Origins. Int. J. Mol. Sci. 2021, 22, 2298. https://doi.org/10.3390/ijms22052298
Hsu C-N, Tain Y-L. Targeting the Renin–Angiotensin–Aldosterone System to Prevent Hypertension and Kidney Disease of Developmental Origins. International Journal of Molecular Sciences. 2021; 22(5):2298. https://doi.org/10.3390/ijms22052298
Chicago/Turabian StyleHsu, Chien-Ning, and You-Lin Tain. 2021. "Targeting the Renin–Angiotensin–Aldosterone System to Prevent Hypertension and Kidney Disease of Developmental Origins" International Journal of Molecular Sciences 22, no. 5: 2298. https://doi.org/10.3390/ijms22052298
APA StyleHsu, C.-N., & Tain, Y.-L. (2021). Targeting the Renin–Angiotensin–Aldosterone System to Prevent Hypertension and Kidney Disease of Developmental Origins. International Journal of Molecular Sciences, 22(5), 2298. https://doi.org/10.3390/ijms22052298