Selective Transport of Protein-Bound Uremic Toxins in Erythrocytes


 , , and
, , and
Abstract
1. Introduction
2. Results
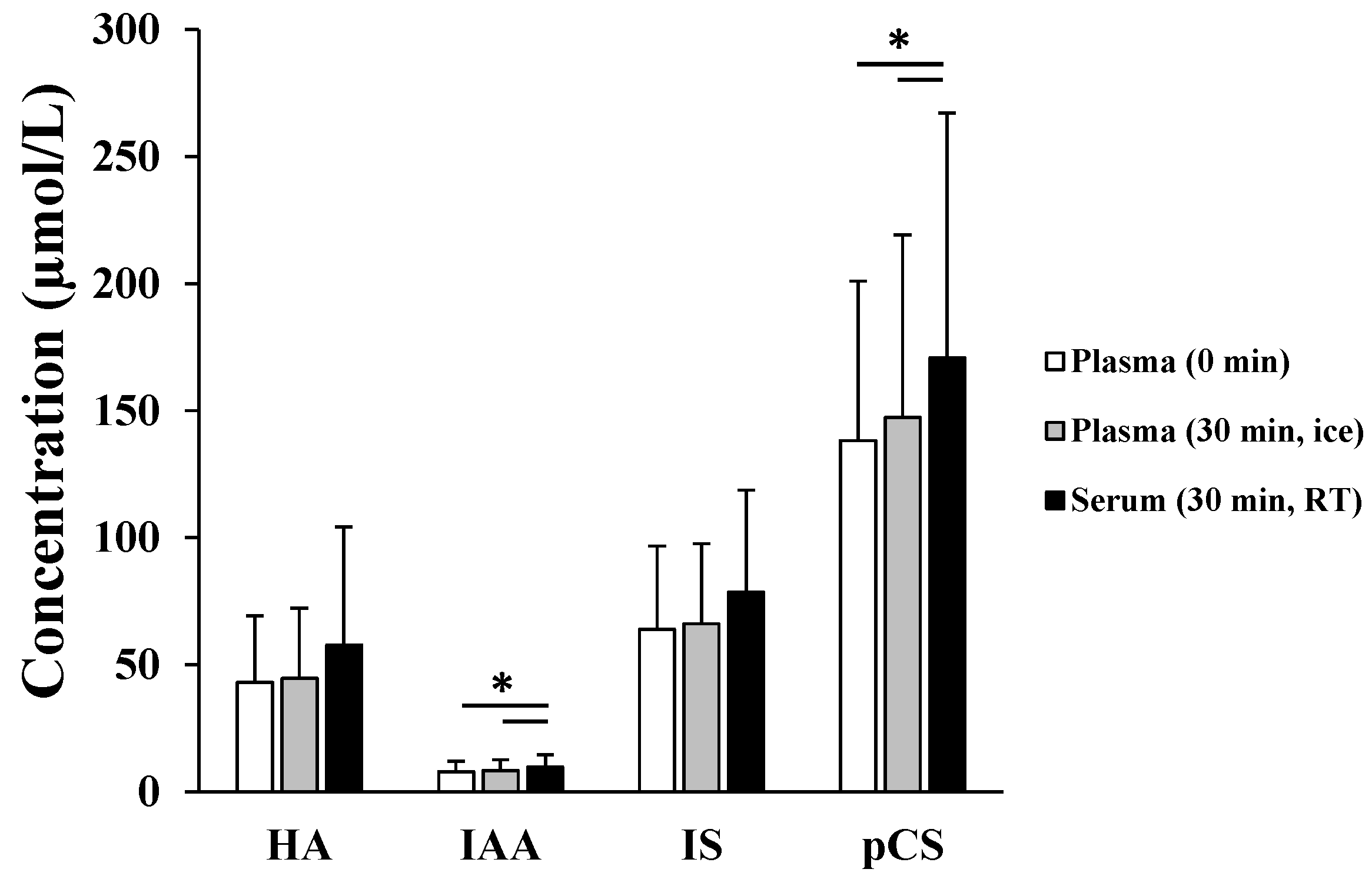
2.1. Presence of Protein-Bound Uremic Toxins (PBUTs) in Erythrocytes
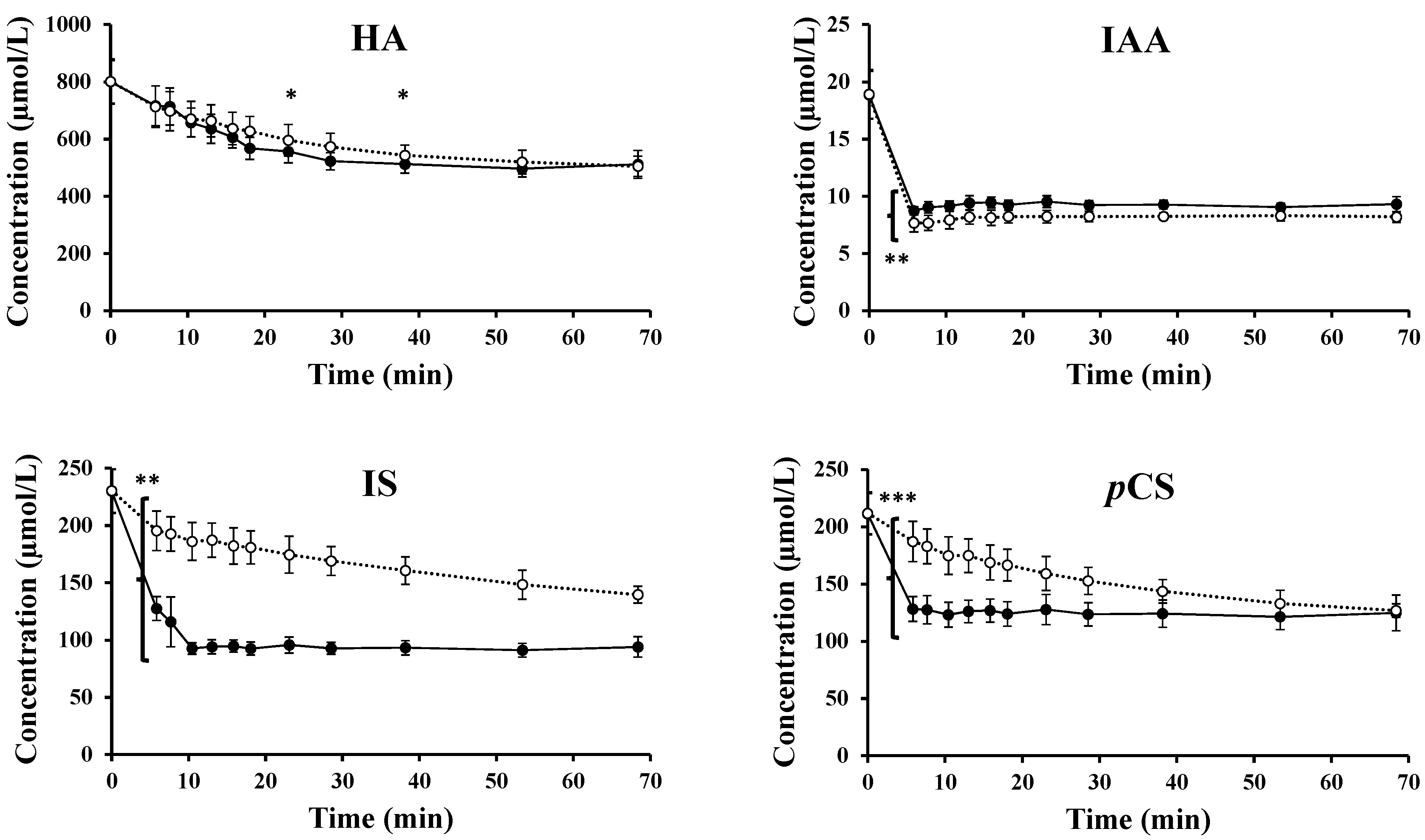
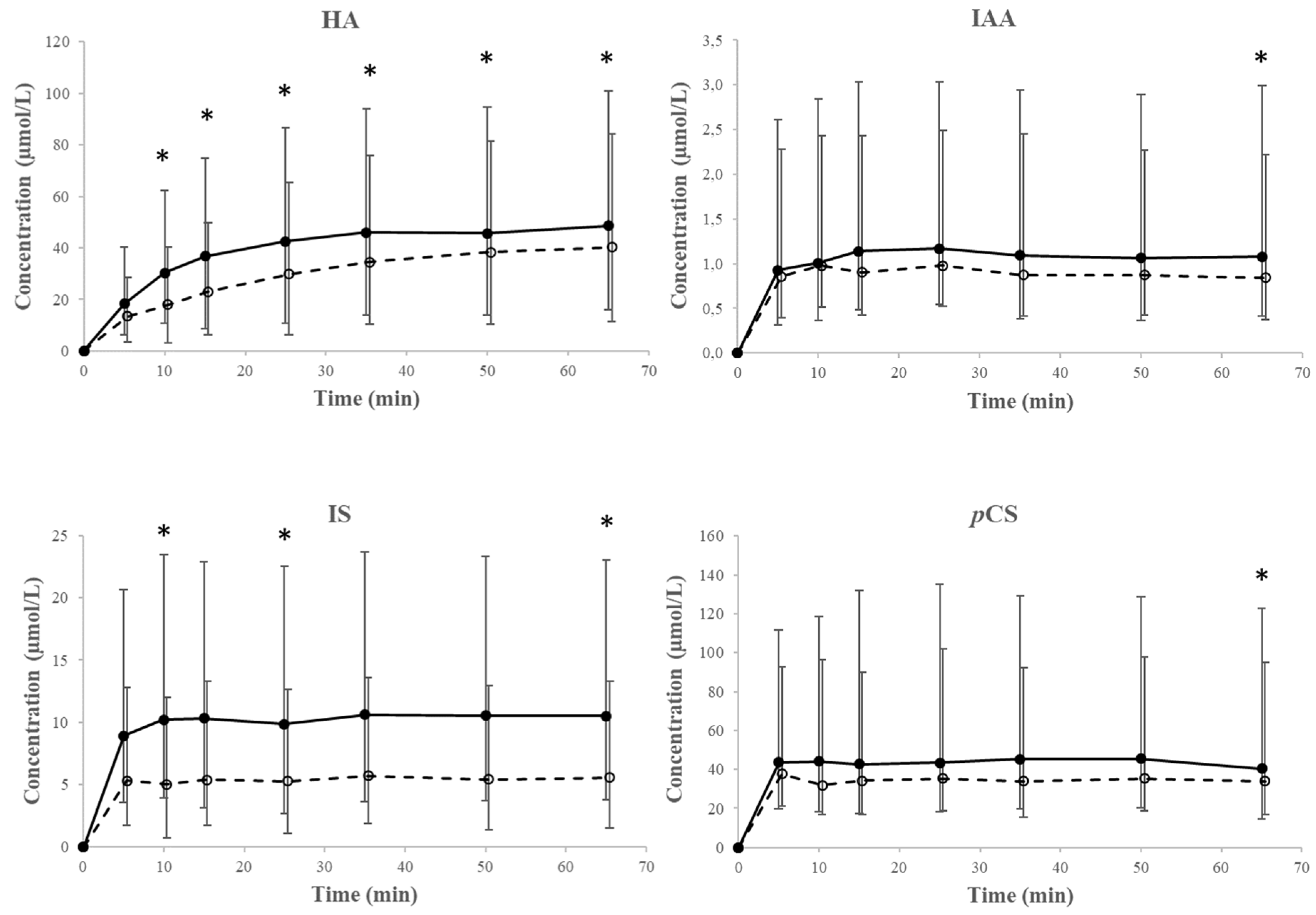
2.2. Experimental Data for Loading and Unloading Experiments
2.3. Transport across the Erythrocyte Membrane—Influx
2.4. Transport Across the Erythrocyte Membrane—Efflux
2.5. Kinetic Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Collection
5.2. Chemicals
5.3. In Vitro Protocols
5.3.1. Proof of Concept—Presence of PBUTs in Erythrocytes?
5.3.2. Transport of PBUTs across the Erythrocyte Membrane
5.4. Analyses
5.5. Kinetic Model
5.6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
Appendix B.1. Berkeley Madonna Script for Loading Experiments for Hippuric Acid (HA) without Inhibitor
Appendix B.2. Berkeley Madonna Script for Unloading Experiments for Hippuric Acid (HA)
References
- Vanholder, R.; De Smet, R.; Glorieux, G.; Argilés, A.; Baurmeister, U.; Brunet, P.; Clark, W.; Cohen, G.; De Deyn, P.P.; Deppisch, R.; et al. Review on uremic toxins: Classification, concentration and interindividual variability. Am. J. Soc. Nephrol. 2003, 63, 1934–1943. [Google Scholar] [CrossRef] [PubMed]
- Duranton, F.; Cohen, G.; De Smet, R.; Rodriguez, M.; Jankowski, J.; Vanholder, R.; Argiles, A. Normal and Pathologic Concentrations of Uremic Toxins. J. Am. Soc. Nephrol. 2012, 23, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Deltombe, O.; de Loor, H.; Glorieux, G.; Dhondt, A.; Van Biesen, W.; Meijers, B.; Eloot, S. Exploring binding characteristics and the related competition of different protein-bound uremic toxins. Biochimie 2017, 139, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Ezawa, A.; Kikuchi, K.; Tsuruta, Y.; Niwa, T. Protein-bound uremic toxins in hemodialysis patients measured by liquid chromatography/tandem mass spectrometry and their effects on endothelial ROS production. Anal. Bioanal. Chem. 2012, 403, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Deltombe, O.; van Biesen, W.; Glorieux, G.; Massy, Z.; Dhondt, A.; Eloot, S. Exploring Protein Binding of Uremic Toxins in Patients with Different Stages of Chronic Kidney Disease and during Hemodialysis. Toxins 2015, 7, 3933–3946. [Google Scholar] [CrossRef] [PubMed]
- Masereeuw, R.; Mutsaers, H.A.; Toyohara, T.; Abe, T.; Jhawar, S.; Sweet, D.H.; Lowenstein, J. The Kidney and Uremic Toxin Removal: Glomerulus or Tubule? Semin. Nephrol. 2014, 34, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; Jankowski, J.; Gajjala, P.R.; Wetzels, J.F.M.; Masereeuw, R. Disposition and clinical implications of protein-bound uremic toxins. Clin. Sci. 2017, 1631–1647. [Google Scholar] [CrossRef]
- Vanholder, R.; de Smet, R.; Lesaffer, G. Dissociation between dialysis adequacy and Kt/V. Semin. Dial. 2002, 15, 3–7. [Google Scholar] [CrossRef]
- Eloot, S.; Van Biesen, W.; Dhondt, A.; Van de Wynkele, H.; Glorieux, G.; Verdonck, P.; Vanholder, R. Impact of hemodialysis duration on the removal of uremic retention solutes. Kidney Int. 2008, 73, 765–770. [Google Scholar] [CrossRef]
- Eloot, S.; Schneditz, D.; Cornelis, T.; Van Biesen, W.; Glorieux, G.; Dhondt, A.; Kooman, J.; Vanholder, R. Protein-bound uremic toxin profiling as a tool to optimize hemodialysis. PLoS ONE 2016, 11, e0147159. [Google Scholar] [CrossRef]
- Meert, N.; Eloot, S.; Schepers, E.; Lemke, H.D.; Dhondt, A.; Glorieux, G.; Van Landschoot, M.; Waterloos, M.A.; Vanholder, R. Comparison of removal capacity of two consecutive generations of high-flux dialysers during different treatment modalities. Nephrol. Dial. Transpl. 2011, 26, 2624–2630. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, T.; Eloot, S.; Vanholder, R.; Glorieux, G.; van der Sande, F.M.; Scheijen, J.L.; Leunissen, K.M.; Kooman, J.P.; Schalkwijk, C.G. Protein-bound uraemic toxins, dicarbonyl stress and advanced glycation end products in conventional and extended haemodialysis and haemodiafiltration. Nephrol. Dial. Transplant. 2015, 30, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Böhringer, F.; Jankowski, V.; Gajjala, P.R.; Zidek, W.; Jankowski, J. Release of Uremic Retention Solutes from Protein Binding by Hypertonic Predilution Hemodiafiltration. ASAIO J. 2015, 61, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Meijers, B.K.; Weber, V.; Bammens, B.; Dehaen, W.; Verbeke, K.; Falkenhagen, D.; Evenepoel, P. Removal of the uremic retention solute p-cresol using fractionated plasma separation and adsorption. Artif. Organs 2008, 32, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Brettschneider, F.; Tölle, M.; von der Giet, M.; Passlick-Deetjen, J.; Steppan, S.; Peter, M.; Jankowski, V.; Krause, A.; Kühne, S.; Zidek, W.; et al. Removal of Protein-Bound, Hydrophobic Uremic Toxins by a Combined Fractionated Plasma Separation and Adsorption Technique. Artif. Organs 2013, 37, 409–416. [Google Scholar] [CrossRef]
- Meyer, T.W.; Peattie, J.W.; Miller, J.D.; Dinh, D.C.; Recht, N.S.; Walther, J.L.; Hostetter, T.H. Increasing the Clearance of Protein-Bound Solutes by Addition of a Sorbent to the Dialysate. J. Am. Soc. Nephrol. 2007, 18, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Sandeman, S.R.; Howell, C.A.; Phillips, G.J.; Zheng, Y.; Standen, G.; Pletzenauer, R.; Davenport, A.; Basnayake, K.; Boyd, O.; Holt, S.; et al. An adsorbent monolith device to augment the removal of uraemic toxins during haemodialysis. J. Mater. Sci. Mater. Med. 2014, 25, 1589–1597. [Google Scholar] [CrossRef]
- Maheshwari, V.; Thijssen, S.; Tao, X.; Fuertinger, D.; Kappel, F.; Kotanko, P. A novel mathematical model of protein-bound uremic toxin kinetics during hemodialysis. Sci. Rep. 2017, 7, 10371. [Google Scholar] [CrossRef]
- Eloot, S.; Torremans, A.; De Smet, R.; Marescau, B.; De Deyn, P.P.; Verdonck, P.; Vanholder, R. Complex Compartmental Behavior of Small Water-Soluble Uremic Retention Solutes: Evaluation by Direct Measurements in Plasma and Erythrocytes. Am. J. Kidney Dis. 2007, 50, 279–288. [Google Scholar] [CrossRef]
- Gary-Bobo, C.; Lindenberg, A.B. Velocity of the penetration of creatinine into human erythrocytes as a function of temperature. J. Physiol. 1960, 52, 106–107. Available online: http://www.ncbi.nlm.nih.gov/pubmed/13826714 (accessed on 30 November 2017).
- Langsdorf, L.J.; Zydney, A.L. Effect of Uremia on the Membrane Transport Characteristics of Red Blood Cells. Blood 1993, 81, 820–827. Available online: http://www.bloodjournal.org/content/bloodjournal/81/3/820.full.pdf (accessed on 10 May 2018). [PubMed]
- Sands, J.; Timmer, R.; Gunn, R. Urea transporters in kidney and erythrocytes. Am. J. Physiol. 1997, 273, F321–F339. [Google Scholar] [CrossRef] [PubMed]
- Schneditz, D.; Yang, Y.; Christopoulos, G.; Kellner, J. Rate of creatinine equilibration in whole blood. Hemodial. Int. 2009, 13, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Brahm, J. The permeability of red blood cells to chloride, urea and water. J. Exp. Biol. 2013, 216, 2238–2246. [Google Scholar] [CrossRef] [PubMed]
- Soszynski, M.; Bartosz, G. Penetration of erythrocyte membrane by peroxynitrite: Participation of the anion exchange protein. Biochem. Mol. Biol. Int. 1997, 43, 319–325. [Google Scholar] [CrossRef]
- Salhany, J.M. Stilbenedisulfonate binding kinetics to band 3 (AE 1): Relationship between transport and stilbenedisulfonate binding sites and role of subunit interactions in transport. Blood Cells Mol. Dis. 2001, 27, 127–134. [Google Scholar] [CrossRef]
- Salhany, J.M.; Schopfer, L.M. Kinetic mechanism of DIDS binding to band 3 (AE1) in human erythrocyte membranes. Blood Cells Mol. Dis. 2001, 27, 844–849. [Google Scholar] [CrossRef]
- Reithmeier, R.A.F.; Casey, J.R.; Kalli, A.C.; Sansom, M.S.P.; Alguel, Y.; Iwata, S. Band 3, the human red cell chloride/bicarbonate anion exchanger (AE1, SLC4A1), in a structural context. Biochim. Biophys. Acta 2016, 1858, 1507–1532. [Google Scholar] [CrossRef]
- Renes, J.; de Vries, E.G.; Nienhuis, E.F.; Jansen, P.L.; Müller, M. ATP- and glutathione-dependent transport of chemotherapeutic drugs by the multidrug resistance protein MRP1. Br. J. Pharmacol. 1999, 126, 681–688. [Google Scholar] [CrossRef]
- Allen, J.D.; van Loevezijn, A.; Lakhai, J.M.; van der Valk, M.; van Tellingen, O.; Reid, G.; Schellens, J.H.; Koomen, G.J.; Schinkel, A.H. Potent and specific inhibition of the breast cancer resistance protein multidrug transporter in vitro and in mouse intestine by a novel analogue of fumitremorgin C. Mol. Cancer Ther. 2002, 1, 417–425. [Google Scholar]
- Wu, W.; Bush, K.T.; Nigam, S.K. Key Role for the Organic Anion Transporters, OAT1 and OAT3, in the in vivo Handlingof Uremic Toxins and Solutes. Sci. Rep. 2017, 10, 4939. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, B.; Balcerczyk, A.; Klimczak, A.; Bartosz, G. Membrane Biology The Role of Multidrug Resistance Protein 1 (MRP1) in Transport of Fluorescent Anions across the Human Erythrocyte Membrane. J. Membr. Biol. 2003, 193, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Wesoaowska, O.; Mosiadz, D.; Motohashi, N.; Kawase, M.; Michalak, K. Phenothiazine maleates stimulate MRP1 transport activity in human erythrocytes. Biochim. Biophys. Acta 2005, 1720, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Homolya, L.; Váradi, A.; Sarkadi, B. Multidrug resistance-associated proteins: Export pumps for conjugates with glutathione, glucuronate or sulfate. BioFactors 2003, 17, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Deeley, R.G.; Cole, S.P.C. Substrate recognition and transport by multidrug resistance protein 1 (ABCC1). FEBS Lett. 2005, 580, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Hegedűs, T.; Chaubey, P.M.; Várady, G.; Szabó, E.; Sarankó, H.; Hofstetter, L.; Roschitzki, B.; Stieger, B.; Sarkadi, B. Inconsistencies in the red blood cell membrane proteome analysis: Generation of a database for research and diagnostic applications. Database 2015, 2015. [Google Scholar] [CrossRef]
- Caetano-Pinto, P.; Janssen, M.J.; Gijzen, L.; Verscheijden, L.; Wilmer, M.J.; Masereeuw, R. Fluorescence Based Transport Assays Revisited in a Human REnal Proximal Tubule Cell Line. Mol. Pharm. 2016, 13, 933–944. [Google Scholar] [CrossRef]
- Descombes, E.; Perriard, F.; Fellay, G. Diffusion kinetics of urea, creatinine and uric acid in blood during hemodialysis. Clinical implications. Clin. Nephrol. 1993, 40, 286–295. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8281718 (accessed on 8 March 2018).
- Schneditz, D.; Platzer, D.; Daugirdas, J.T. A diffusion-adjusted regional blood flow model to predict solute kinetics during haemodialysis. Nephrol. Dial. Transplant. 2009, 24, 2218–2224. [Google Scholar] [CrossRef][Green Version]
- Deltombe, O.; Dhondt, A.; van Biesen, W.; Glorieux, G.; Eloot, S. Effect of sample temperature, pH, and matrix on the percentage protein binding of protein-bound uraemic toxins. Anal. Methods 2017, 9, 1935–1940. [Google Scholar] [CrossRef]
- Kety, S.S. The theory and applications of the exchange of inert gas at the lungs and tissues. Pharmacol. Rev. 1951, 3, 1–41. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Experiment | CBUFFER(0),c (µmol/L) | CBUFFER(eq),m (µmol/L) | Hwb (%) | Hsusp (%) |
|---|---|---|---|---|
| Loading (n = 8) | 47 ± 5 | 46 ± 5 | ||
| HA | 802.9 ± 72.3 | 504.3 ± 33.1 | ||
| IAA | 19.2 ± 1.7 | 9.2 ± 0.5 | ||
| IS | 227.6 ± 20.5 | 92.5 ± 7.0 | ||
| pCS | 209.6 ± 18.9 | 122.9 ± 13.1 | ||
| Loading + inhibitor (n = 8) | 47 ± 5 | 46 ± 5 | ||
| HA | 804.3 ± 76.4 | 518.6 ± 42.4 | ||
| IAA | 19.2 ± 1.8 | 8.3 ± 0.5 * | ||
| IS | 228.0 ± 21.7 | 146.5 ± 14.5 * | ||
| pCS | 210.0 ± 20.0 | 131.8 ± 12.5 | ||
| Unloading (n = 6) | 36 ± 3 | 32 ± 3 | ||
| HA | 0 | 48.6 (32.6;52.4) | ||
| IAA | 0 | 1.08 (0.66;1.91) | ||
| IS | 0 | 10.5 (6.76;12.5) | ||
| pCS | 0 | 40.4 (25.8;82.5) | ||
| Unloading + inhibitor (n = 6) | 36 ± 3 | 33 ± 3 | ||
| HA | 0 | 40.2 (28.8;43.9) * | ||
| IAA | 0 | 0.84 (0.47;1.37) * | ||
| IS | 0 | 5.55 (4.05;7.71) * | ||
| pCS | 0 | 33.8 (16.7;61.3) * | ||
| Type of Experiment | a (1/min) | ks (1/min) | KC (mL/min) | γ |
|---|---|---|---|---|
| Loading (n = 8) | ||||
| HA | 0.06 ± 0.02 | 0.03 ± 0.01 | 0.16 ± 0.04 | 1.24 ± 0.20 |
| IAA | 1.46 ± 0.02a | 0.82 ± 0.10a | 4.66 ± 0.23a | 1.54 ± 0.13a |
| IS | 0.27 ± 0.07b | 0.19 ± 0.05b | 1.10 ± 0.27b | 2.72 ± 0.39a,b |
| pCS | 0.85 ± 0.56a,b,c | 0.40 ± 0.23a,b,c | 2.38 ± 1.46a,b,c | 1.30 ± 0.24b,c |
| Loading + inhibitor (n = 8) | ||||
| HA | 0.04 ± 0.01d | 0.02 ± 0.01d | 0.12 ± 0.04d | 1.24 ± 0.201 |
| IAA | 1.47 ± 0.001a | 0.83 ± 0.10a | 4.68 ± 0.19a | 1.54 ± 0.131,a |
| IS | 0.03 ± 0.004a,d | 0.02 ± 0.003b,d | 0.10 ± 0.01b,d | 2.72 ± 0.391,a,b |
| pCS | 0.05 ± 0.007b,c,d | 0.02 ± 0.004b,d | 0.13 ± 0.02b,d | 1.30 ± 0.241,b,c |
| Unloading (n = 6) | ||||
| HA | 0.10 (0.08;0.13)e | 0.06 (0.05;0.08)e | 0.38 (0.33;0.52)e | 1.242 |
| IAA | 0.51 (0.39;3.29)a | 0.35 (0.28;2.39)a | 2.36 (1.58;15.0)a | 1.542 |
| IS | 0.39 (0.35;1.30)a,e | 0.38 (0.34;1.33)a,e | 2.53 (1.98;8.40)a,e | 2.722 |
| pCS | 0.48 (0.40;2.28)a,e | 0.29 (0.25;1.46)a,b,e | 2.00 (1.42;9.31)a,b,e | 1.302 |
| Unloading + inhibitor (n = 6) | ||||
| HA | 0.07 (0.05;0.09)e | 0.04 (0.03;0.06)e | 0.25 (0.21;0.36)e | 1.242 |
| IAA | 2.19 (0.71;3.66)a | 1.52 (0.50;2.61)a | 8.79 (3.21;16.5)a | 1.542 |
| IS | 0.77 (0.43;3.35)a,e | 0.73 (0.43;3.49)a,e | 4.93 (2.74;19.7)a,e | 2.722 |
| pCS | 2.08 (0.45;4.18)a,c,e | 1.35 (0.28;2.62)a,e | 8.03 (1.83;15.0)a,e | 1.302 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deltombe, O.; Glorieux, G.; Marzouki, S.; Masereeuw, R.; Schneditz, D.; Eloot, S. Selective Transport of Protein-Bound Uremic Toxins in Erythrocytes. Toxins 2019, 11, 385. https://doi.org/10.3390/toxins11070385
Deltombe O, Glorieux G, Marzouki S, Masereeuw R, Schneditz D, Eloot S. Selective Transport of Protein-Bound Uremic Toxins in Erythrocytes. Toxins. 2019; 11(7):385. https://doi.org/10.3390/toxins11070385
Chicago/Turabian StyleDeltombe, Olivier, Griet Glorieux, Sami Marzouki, Rosalinde Masereeuw, Daniel Schneditz, and Sunny Eloot. 2019. "Selective Transport of Protein-Bound Uremic Toxins in Erythrocytes" Toxins 11, no. 7: 385. https://doi.org/10.3390/toxins11070385
APA StyleDeltombe, O., Glorieux, G., Marzouki, S., Masereeuw, R., Schneditz, D., & Eloot, S. (2019). Selective Transport of Protein-Bound Uremic Toxins in Erythrocytes. Toxins, 11(7), 385. https://doi.org/10.3390/toxins11070385