
Neither Beetroot Juice Supplementation nor Increased Carbohydrate Oxidation Enhance Economy of Prolonged Exercise in Elite Race Walkers

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview and Participants
2.2. Overview of Dietary Control and Interventions
2.3. Physiological Testing before and after the Studies
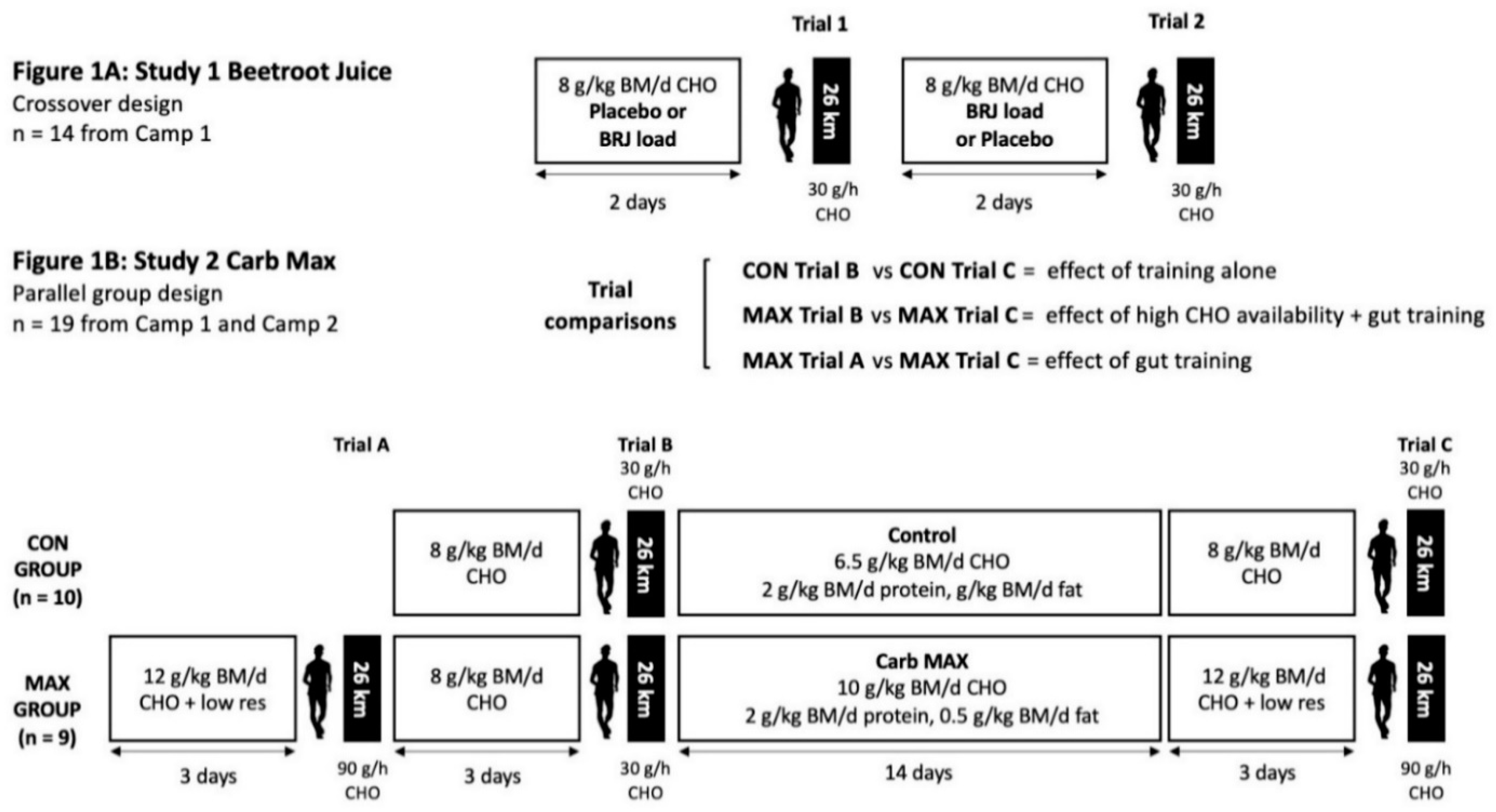
2.4. Study One (Beetroot Juice)
2.4.1. Overview
2.4.2. 26-km Exercise Protocol
2.4.3. Venous Blood Analysis—NO2− and NO3−
2.5. Study Two (Carb Max)
2.5.1. Overview
2.5.2. Summary of Dietary Intervention for Study Two
2.5.3. 26-km Exercise Protocol
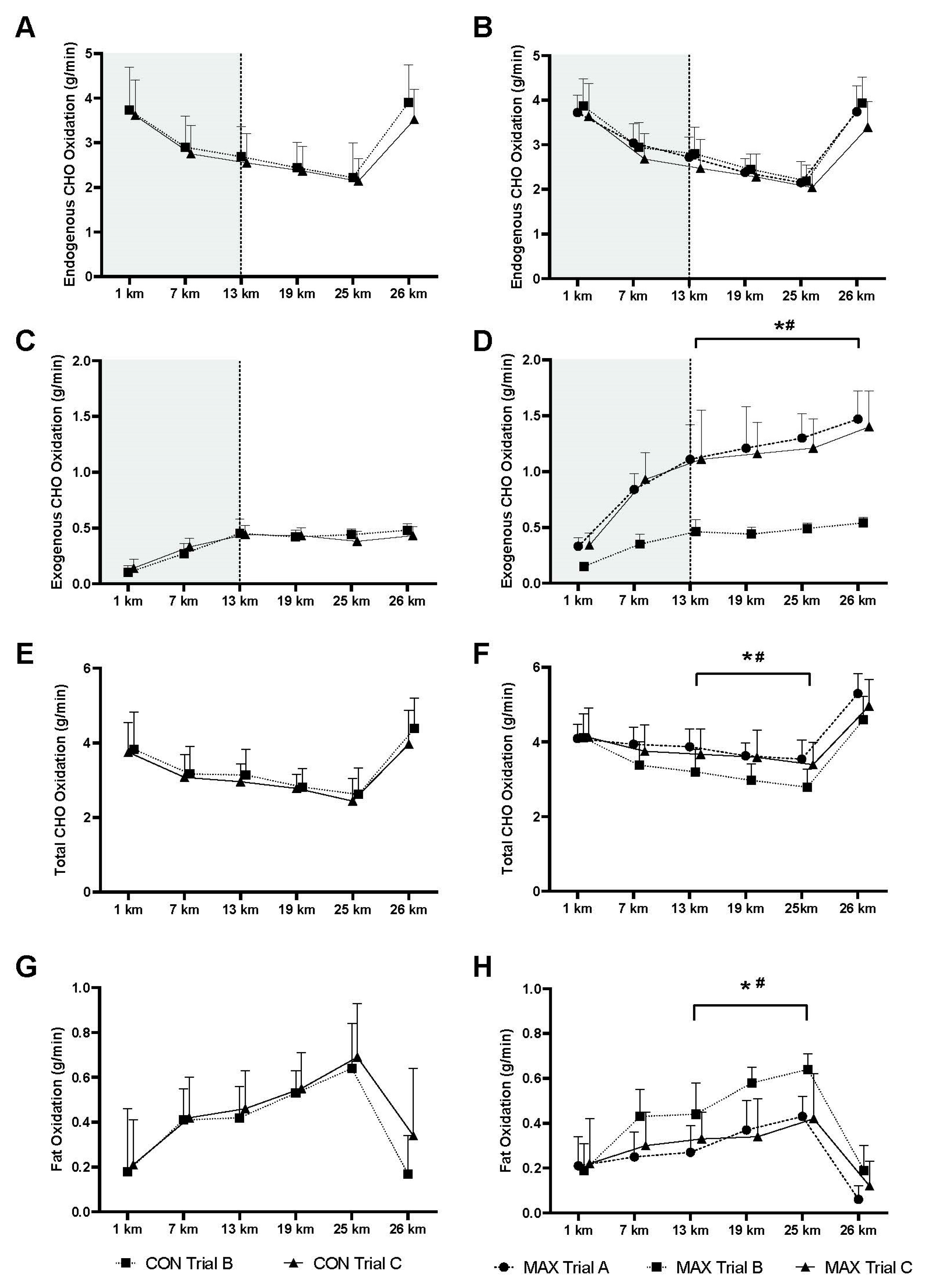
2.5.4. 13C Breath Analysis
2.6. Statistical Analysis
3. Results
3.1. Physiological Changes over the Training Camp
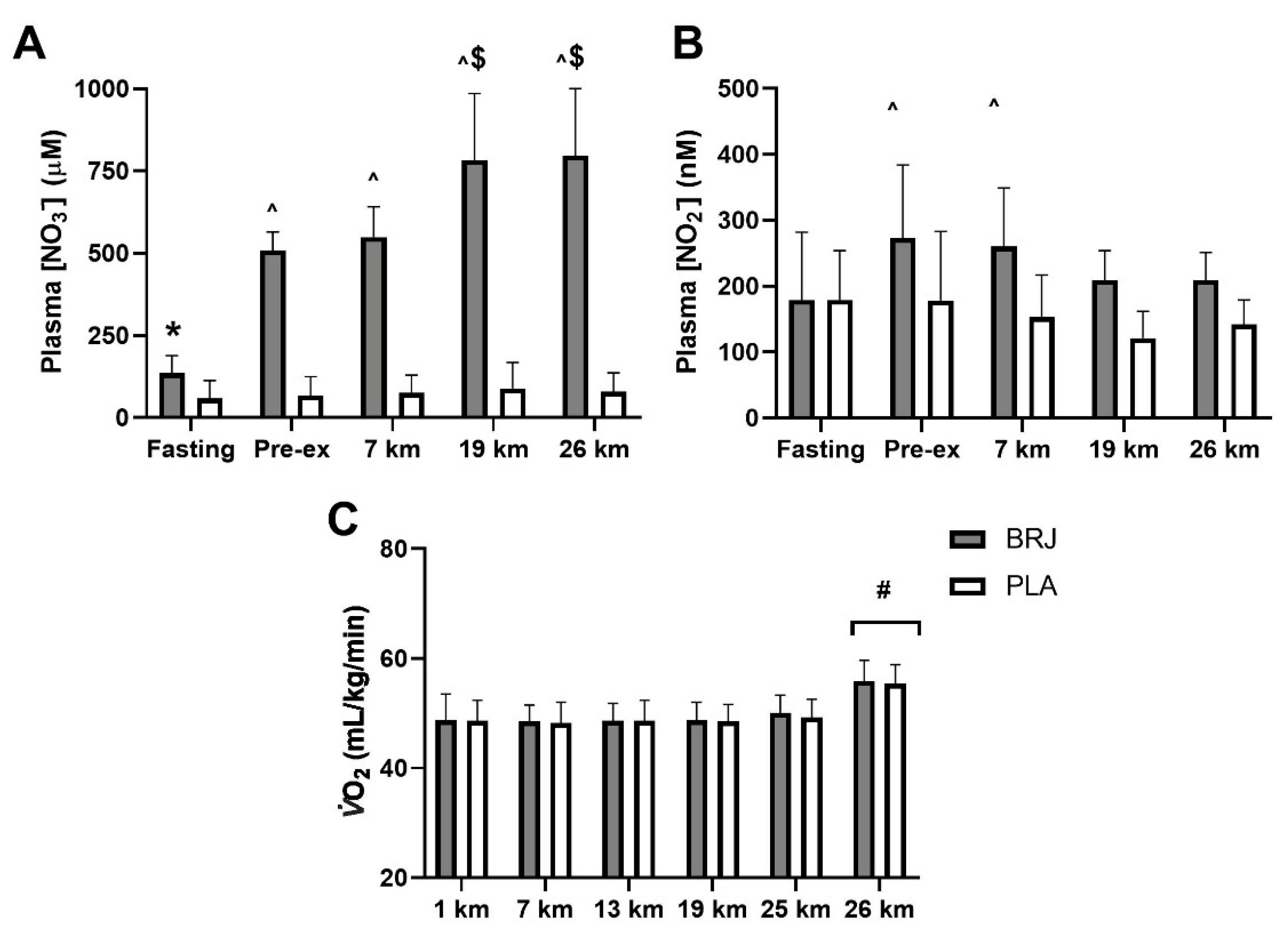
3.2. Results of Study One (Beetroot Juice)
3.3. Results of Study Two (Carb Max)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joyner, M.J.; Hunter, S.K.; Lucia, A.; Jones, A.M. Physiology and fast marathons. J. Appl. Physiol. 2020, 128, 1065–1068. [Google Scholar] [CrossRef]
- Jones, A.M.; Kirby, B.S.; Clark, I.E.; Rice, H.M.; Fulkerson, E.; Wylie, L.J.; Wilkerson, D.P.; Vanhatalo, A.; Wilkins, B.W. Physiological demands of running at 2-hour marathon race pace. J. Appl. Physiol. 2021, 130, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Jeukendrup, A.E.; Jones, A.M.; Mooses, M. Contemporary Nutrition Strategies to Optimize Performance in Distance Runners and Race Walkers. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stellingwerff, T.; Bovim, I.M.; Whitfield, J. Contemporary Nutrition Interventions to Optimize Performance in Middle-Distance Runners. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 106–116. [Google Scholar] [CrossRef]
- Perrey, S. Commentaries on Viewpoint: Physiology and fast marathons. J. Appl. Physiol. 2020, 128, 1069–1085. [Google Scholar] [CrossRef]
- Burke, L.M.; Hawley, J.; Jeukendrup, A.; Morton, J.P.; Stellingwerff, T.; Maughan, R.J. Toward a Common Understanding of Diet–Exercise Strategies to Manipulate Fuel Availability for Training and Competition Preparation in Endurance Sport. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Daniels, J.; Daniels, N. Running economy of elite male and elite female runners. Med. Sci. Sports Exerc. 1992, 24, 483–489. [Google Scholar] [CrossRef]
- Morgan, D.W.; Baldini, F.D.; Martin, P.E.; Kohrt, W.M. Ten kilometer performance and predicted velocity at V02max among well-trained male runners. Med. Sci. Sports Exerc. 1989, 21, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P.U.; Pyne, D.B.; Telford, R.D.; Hawley, J.A. Factors Affecting Running Economy in Trained Distance Runners. Sports Med. 2004, 34, 465–485. [Google Scholar] [CrossRef]
- Kipp, S.; Kram, R.; Hoogkamer, W. Extrapolating Metabolic Savings in Running: Implications for Performance Predictions. Front. Physiol. 2019, 10, 79. [Google Scholar] [CrossRef]
- Burgess, M. The Incredible Science behind Eliud Kipchoge’s 1:59 Marathon. Wired. Available online: https://www.wired.co.uk/article/eliud-kipchoge-ineos-159-marathon (accessed on 1 August 2021).
- Hoogkamer, W.; Kipp, S.; Frank, J.H.; Farina, E.M.; Luo, G.; Kram, R. A Comparison of the Energetic Cost of Running in Marathon Racing Shoes. Sports Med. 2018, 48, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Hoogkamer, W.; Snyder, K.L.; Arellano, C.J. Modeling the Benefits of Cooperative Drafting: Is There an Optimal Strategy to Facilitate a Sub-2-Hour Marathon Performance? Sports Med. 2018, 48, 2859–2867. [Google Scholar] [CrossRef]
- Hoogkamer, W.; Snyder, K.L.; Arellano, C.J. Reflecting on Eliud Kipchoge’s Marathon World Record: An Update to Our Model of Cooperative Drafting and Its Potential for a Sub-2-Hour Performance. Sports Med. 2019, 49, 167–170. [Google Scholar] [CrossRef]
- Larsen, F.J.; Weitzberg, E.; Lundberg, J.O.; Ekblom, B. Effects of dietary nitrate on oxygen cost during exercise. Acta Physiol. 2007, 191, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.; Winyard, P.; Vanhatalo, A.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Tarr, J.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation reduces the O2cost of low-intensity exercise and enhances tolerance to high-intensity exercise in humans. J. Appl. Physiol. 2009, 107, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Lansley, K.E.; Winyard, P.; Fulford, J.; Vanhatalo, A.; Bailey, S.; Blackwell, J.R.; DiMenna, F.J.; Gilchrist, M.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation reduces the O2cost of walking and running: A placebo-controlled study. J. Appl. Physiol. 2011, 110, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Thompson, C.; Wylie, L.; Vanhatalo, A. Dietary Nitrate and Physical Performance. Annu. Rev. Nutr. 2018, 38, 303–328. [Google Scholar] [CrossRef]
- Whitfield, J.; Ludzki, A.; Heigenhauser, G.J.F.; Senden, J.M.G.; Verdijk, L.; van Loon, L.J.; Spriet, L.L.; Holloway, G.P. Beetroot juice supplementation reduces whole body oxygen consumption but does not improve indices of mitochondrial efficiency in human skeletal muscle. J. Physiol. 2016, 594, 421–435. [Google Scholar] [CrossRef] [Green Version]
- McIlvenna, L.C.; Muggeridge, D.J.; Whitfield, J. Exploring the mechanisms by which nitrate supplementation improves skeletal muscle contractile function: One fibre at a time. J. Physiol. 2019, 598, 25–27. [Google Scholar] [CrossRef]
- Tan, R.; Wylie, L.; Thompson, C.; Blackwell, J.R.; Bailey, S.; Vanhatalo, A.; Jones, A.M. Beetroot juice ingestion during prolonged moderate-intensity exercise attenuates progressive rise in O2 uptake. J. Appl. Physiol. 2018, 124, 1254–1263. [Google Scholar] [CrossRef] [Green Version]
- Breese, B.C.; McNarry, M.; Marwood, S.; Blackwell, J.R.; Bailey, S.; Jones, A.M. Beetroot juice supplementation speeds O2uptake kinetics and improves exercise tolerance during severe-intensity exercise initiated from an elevated metabolic rate. Am. J. Physiol. Integr. Comp. Physiol. 2013, 305, R1441–R1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.M.; Sharma, A.P.; Heikura, I.; Forbes, S.F.; Holloway, M.; McKay, A.K.A.; Bone, J.L.; Leckey, J.J.; Welvaert, M.; Ross, M.L. Crisis of confidence averted: Impairment of exercise economy and performance in elite race walkers by ketogenic low carbohydrate, high fat (LCHF) diet is reproducible. PLoS ONE 2020, 15, e0234027. [Google Scholar] [CrossRef]
- Burke, L.M.; Whitfield, J.; Heikura, I.A.; Ross, M.L.R.; Tee, N.; Forbes, S.F.; Hall, R.; McKay, A.K.A.; Wallett, A.M.; Sharma, A.P. Adaptation to a low carbohydrate high fat diet is rapid but impairs endurance exercise metabolism and performance despite enhanced glycogen availability. J. Physiol. 2021, 599, 771–790. [Google Scholar] [CrossRef]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.; Burke, L.M.; Mckay, A.K.A.; Heikura, I.A.; Hall, R.; Fensham, N.; Sharma, A.P. Acute Ketogenic Diet and Ketone Ester Supplementation Impairs Race Walk Performance. Med. Sci. Sports Exerc. 2021, 53, 776–784. [Google Scholar] [CrossRef]
- Shaw, D.M.; Merien, F.; Braakhuis, A.; Maunder, E.; Dulson, D.K. Effect of a Ketogenic Diet on Submaximal Exercise Capacity and Efficiency in Runners. Med. Sci. Sports Exerc. 2019, 51, 2135–2146. [Google Scholar] [CrossRef]
- Burke, L.M. Ketogenic low-CHO, high-fat diet: The future of elite endurance sport? J. Physiol. 2021, 599, 819–843. [Google Scholar] [CrossRef] [PubMed]
- Zuntz, N.; Schumberg, W. Studien zu Einer Physiologie des Marsches; Hirschwald: Berlin, Germany, 1901; p. 361. [Google Scholar]
- Krogh, A.; Lindhard, J. The Relative Value of Fat and Carbohydrate as Sources of Muscular Energy. Biochem. J. 1920, 14, 290–363. [Google Scholar] [CrossRef]
- Leverve, X.; Batandier, C.; Fontaine, E.; Batandier, C. Choosing the Right Substrate. Novartis Found. Symp. 2008, 280, 108–127. [Google Scholar] [CrossRef]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. Position of the Academy of Nutrition and Dietetics, Dietitians of Canada, and the American College of Sports Medicine: Nutrition and Athletic Performance. J. Acad. Nutr. Diet. 2016, 116, 501–528. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A. Modulation of carbohydrate and fat utilization by diet, exercise and environment. Biochem. Soc. Trans. 2003, 31, 1270–1273. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, M. Exercise, muscle, and CHO metabolism. Scand. J. Med. Sci. Sports 2015, 25, 29–33. [Google Scholar] [CrossRef]
- Burke, L.M.; van Loon, L.J.C.; Hawley, J.A. Postexercise muscle glycogen resynthesis in humans. J. Appl. Physiol. 2017, 122, 1055–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stellingwerff, T.; Cox, G. Systematic review: Carbohydrate supplementation on exercise performance or capacity of varying durations. Appl. Physiol. Nutr. Metab. 2014, 39, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.J.; Miall, A.; Khoo, A.; Rauch, C.; Snipe, R.; Camões-Costa, V.; Gibson, P. Gut-training: The impact of two weeks repetitive gut-challenge during exercise on gastrointestinal status, glucose availability, fuel kinetics, and running performance. Appl. Physiol. Nutr. Metab. 2017, 42, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeukendrup, A.E. Training the Gut for Athletes. Sports Med. 2017, 47, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Margolskee, R.F.; Dyer, J.; Kokrashvili, Z.; Salmon, K.S.H.; Ilegems, E.; Daly, K.; Maillet, E.; Ninomiya, Y.; Mosinger, B.; Shirazi-Beechey, S.P. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc. Natl. Acad. Sci. USA 2007, 104, 15075–15080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, L.J.; Kelly, J.; Bailey, S.J.; Blackwell, J.R.; Skiba, P.F.; Winyard, P.G.; Jeukendrup, A.E.; Vanhatalo, A.; Jones, A.M. Beetroot juice and exercise: Pharmacodynamic and dose-response relationships. J. Appl. Physiol. 2013, 115, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American college of sports medicine joint position statement. nutrition and athletic performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar] [CrossRef]
- Heikura, I.A.; Stellingwerff, T.; Burke, L.M. Self-Reported Periodization of Nutrition in Elite Female and Male Runners and Race Walkers. Front. Physiol. 2018, 9, 1732. [Google Scholar] [CrossRef]
- Burke, L.M.; Cox, G.R.; Cummings, N.K.; Desbrow, B. Guidelines for daily CHO intake: Do athletes achieve them? Sports Med. 2001, 31, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Mirtschin, J.G.; Forbes, S.; Cato, L.E.; Heikura, I.; Strobel, N.; Hall, R.; Burke, L.M. Organization of Dietary Control for Nutrition-Training Intervention Involving Periodized Carbohydrate Availability and Ketogenic Low-Carbohydrate High-Fat Diet. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Nana, A.; Slater, G.; Stewart, A.; Burke, L.M. Methodology Review: Using Dual-Energy X-Ray Absorptiometry (DXA) for the Assessment of Body Composition in Athletes and Active People. Int. J. Sport Nutr. Exerc. Metab. 2015, 25, 198–215. [Google Scholar] [CrossRef]
- Bone, J.L.; Burke, L.M. No Difference in Young Adult Athletes’ Resting Energy Expenditure When Measured Under Inpatient or Outpatient Conditions. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 464–467. [Google Scholar] [CrossRef]
- Saunders, P.U.; Telford, R.; Pyne, D.; Cunningham, R.B.; Gore, C.J.; Hahn, A.G.; Hawley, J. Improved running economy in elite runners after 20 days of simulated moderate-altitude exposure. J. Appl. Physiol. 2004, 96, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Hanley, B. An Analysis of Pacing Profiles of World-Class Racewalkers. Int. J. Sports Physiol. Perform. 2013, 8, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Péronnet, F.; Massicotte, D. Table of nonprotein respiratory quotient: An update. Can. J. Sport Sci. 1991, 16, 23–29. [Google Scholar]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. Metab. 1993, 265, E380–E391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peronnet, F.; Massicotte, D.; Brisson, G.; Hillaire-Marcel, C. Use of 13C substrates for metabolic studies in exercise: Methodological considerations. J. Appl. Physiol. 1990, 69, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.J.; Koziet, J.; Chauvet, D.; Darmaun, D.; Desjeux, J.F.; Young, V.R. Use of 13C-labeled glucose for estimating glucose oxidation: Some design considerations. J. Appl. Physiol. 1987, 63, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Pallikarakis, N.; Sphiris, N.; Lefebvre, P. Influence of the bicarbonate pool and on the occurrence of 13CO2 in exhaled air. Eur. J. Appl. Physiol. Occup. Physiol. 1991, 63, 179–183. [Google Scholar] [CrossRef]
- Maughan, R.J.; Burke, L.M.; Dvorak, J.; Larson-Meyer, D.E.; Peeling, P.; Phillips, S.; Rawson, E.S.; Walsh, N.P.; Garthe, I.; Geyer, H.; et al. IOC Consensus Statement: Dietary Supplements and the High-Performance Athlete. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 104–125. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Vanhatalo, A.; Seals, D.R.; Rossman, M.J.; Piknova, B.; Jonvik, K.L. Dietary Nitrate and Nitric Oxide Metabolism: Mouth, Circulation, Skeletal Muscle, and Exercise Performance. Med. Sci. Sports Exerc. 2021, 53, 280–294. [Google Scholar] [CrossRef]
- Pawlak-Chaouch, M.; Boissière, J.; Gamelin, F.X.; Cuvelier, G.; Berthoin, S.; Aucouturier, J. Effect of dietary nitrate supplementation on metabolic rate during rest and exercise in human: A systematic review and a meta-analysis. Nitric Oxide 2016, 53, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.L.; Alorda-Capo, F.; Pareja-Galeano, H.; Jiménez, S.L. Influence of Nitrate Supplementation on Endurance Cyclic Sports Performance: A Systematic Review. Nutrients 2020, 12, 1796. [Google Scholar] [CrossRef]
- Senefeld, J.W.; Wiggins, C.C.; Regimbal, R.J.; Dominelli, P.B.; Baker, S.E.; Joyner, M.J. Ergogenic Effect of Nitrate Supplementation: A Systematic Review and Meta-analysis. Med. Sci. Sports Exerc. 2020, 52, 2250–2261. [Google Scholar] [CrossRef] [PubMed]
- Hlinský, T.; Kumstát, M.; Vajda, P. Effects of Dietary Nitrates on Time Trial Performance in Athletes with Different Training Status: Systematic Review. Nutrients 2020, 12, 2734. [Google Scholar] [CrossRef] [PubMed]
- Jonvik, K.L.; Nyakayiru, J.; van Loon, L.J.; Verdijk, L.B. Can elite athletes benefit from dietary nitrate supplementation? J. Appl. Physiol. 2015, 119, 759–761. [Google Scholar] [CrossRef] [Green Version]
- Jonvik, K.L.; Nyakayiru, J.; van Loon, L.J.; Verdijk, L.B. Last Word on Viewpoint: Can elite athletes benefit from dietary nitrate supplementation? J. Appl. Physiol. 2015, 119, 770. [Google Scholar] [CrossRef] [Green Version]
- Hultström, M.; De Paula, C.A.; Fontes, M.A.P.; Porcelli, S.; Bellistri, G.; Pugliese, L.; Rasica, L.; Marzorati, M.; Pavei, G.; Ferguson, S.K.; et al. Commentaries on Viewpoint: Can elite athletes benefit from dietary nitrate supplementation? J. Appl. Physiol. 2015, 119, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Nyakayiru, J.; van Loon, L.J.; Verdijk, L.B. Could intramuscular storage of dietary nitrate contribute to its ergogenic effect? A mini-review. Free. Radic. Biol. Med. 2020, 152, 295–300. [Google Scholar] [CrossRef]
- Senkus, K.E.; Crowe-White, K.M. Influence of mouth rinse use on the enterosalivary pathway and blood pressure regulation: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 60, 2874–2886. [Google Scholar] [CrossRef]
- Coyle, E.F.; Jeukendrup, A.E.; Oseto, M.C.; Hodgkinson, B.J.; Zderic, T.W. Low-fat diet alters intramuscular substrates and reduces lipolysis and fat oxidation during exercise. Am. J. Physiol. Metab. 2001, 280, E391–E398. [Google Scholar] [CrossRef]
- Simonsen, J.C.; Sherman, W.M.; Lamb, D.R.; Dernbach, A.R.; Doyle, J.A.; Strauss, R. Dietary carbohydrate, muscle glycogen, and power output during rowing training. J. Appl. Physiol. 1991, 70, 1500–1505. [Google Scholar] [CrossRef] [Green Version]
- Baur, D.A.; Saunders, M.J. Carbohydrate supplementation: A critical review of recent innovations. Eur. J. Appl. Physiol. 2021, 121, 23–66. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E. Carbohydrate and exercise performance: The role of multiple transportable carbohydrates. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, T. Chris Froome: Team Sky’s Unprecedented Release of Data Reveals How British Rider Won Giro d’Italia. 2018. Available online: https://www.bbc.com/sport/cycling/44694122 (accessed on 1 August 2021).
- Pfeiffer, B.; Stellingwerff, T.; Hodgson, A.B.; Randell, R.; Pöttgen, K.; Res, P.; Jeukendrup, A.E. Nutritional Intake and Gastrointestinal Problems during Competitive Endurance Events. Med. Sci. Sports Exerc. 2012, 44, 344–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, G.; Clark, S.A.; Cox, A.J.; Halson, S.; Hargreaves, M.; Hawley, J.; Jeacocke, N.; Snow, R.J.; Yeo, W.K.; Burke, L.M. Daily training with high carbohydrate availability increases exogenous carbohydrate oxidation during endurance cycling. J. Appl. Physiol. 2010, 109, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Stellingwerff, T.; Godin, J.-P.; Beaumont, M.; Tavenard, A.; Grathwohl, D.; Van Bladeren, P.J.; Kapp, A.-F.; Le Coutre, J.; Damak, S. Effects of pre-exercise sucralose ingestion on carbohydrate oxidation during exercise. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 584–592. [Google Scholar] [CrossRef]
- Jeukendrup, A.E.; Jentjens, R. Oxidation of carbohydrate feedings during prolonged exercise: Current thoughts, guidelines and directions for future research. Sports Med. 2000, 29, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Barnes, K.R.; Kilding, A.E. Running economy: Measurement, norms, and determining factors. Sports Med. Open 2015, 1, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Crouter, S.E.; Antczak, A.; Hudak, J.R.; Dellavalle, D.M.; Haas, J.D. Accuracy and reliability of the ParvoMedics TrueOne 2400 and MedGraphics VO2000 metabolic systems. Eur. J. Appl. Physiol. 2006, 98, 139–151. [Google Scholar] [CrossRef]
- Hopkins, W.G.; Hawley, J.; Burke, L.M. Design and analysis of research on sport performance enhancement. Med. Sci. Sports Exerc. 1999, 31, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P.U.; Pyne, D.; Telford, R.; Hawley, J. Reliability and Variability of Running Economy in Elite Distance Runners. Med. Sci. Sports Exerc. 2004, 36, 1972–1976. [Google Scholar] [CrossRef] [PubMed]
- Hoogkamer, W.; Kipp, S.; Spiering, B.A.; Kram, R. Altered Running Economy Directly Translates to Altered Distance-Running Performance. Med. Sci. Sports Exerc. 2016, 48, 2175–2180. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study One: BRJ | Study Two: Carb Max | ||
|---|---|---|---|
| Characteristics | (n = 14) | CON (n = 10) | MAX (n = 9) |
| Age (y) | 30.7 (4.2) | 29.4 (4.6) | 29.7 (4.2) |
| Body Mass (kg) | 67.9 (4.7) | 68.4 (9.4) | 68.7 (5.0) |
| O2peak (mL/kg/min) | 63.9 (5.5) | 60.9 (5.3) | 63.1 (4.6) |
| 10 km personal best min:sec.00) | 40:45.32 (1:02.50) | 41:11.21 (1:33.31) | 40:55.00 (1:03.36) |
| 20 km personal best (hr:min.sec) | 1:22.53 (0:02.03) | 1:24.29 (0:04.45) | 1:23.04 (0:01.59) |
| Baseline Testing | Post Testing | |||
|---|---|---|---|---|
| CON (n = 10) | MAX (n = 9) | CON (n = 10) | MAX (n = 9) | |
| O2peak (mL/kg/min) | 60.9 (5.3) | 63.1 (4.6) | 63.1 (4.1) * | 64.1 (4.6) * |
| Body Mass (kg) | 68.4 (9.4) | 68.7 (5.0) | 68.2 (9.9) * | 67.3 (4.6) * |
| Fat Mass (kg) | 8.7 (1.9) | 9.3 (2.8) | 7.8 (2.2) | 7.9 (2.3) * |
| Fat Free Mass (kg) | 60.3 (8.8) | 60.0 (5.4) | 61.0 (8.8) | 60.1 (5.0) |
| RMR (kJ/d) | 7992 (1017) | 7597 (427) | 7614 (1168) * | 7391 (573) * |
| RMR (kcal/kg/FFM) | 133 (13) | 127 (9) | 126 (9) * | 123 (10) * |
| Pre | 1 km | 7 km | 13 km | 19 km | 25 km | 26 km | ||
|---|---|---|---|---|---|---|---|---|
| Body Mass (kg) | BRJ | 69.8 (6.4) | 68.2 (6.3) | |||||
| PLA | 68.7 (5.4) | 67.6 (6.1) | ||||||
| CHO Ox (g/min) | BRJ | 3.98 (0.75) | 3.42 (0.64) * | 3.38 (0.52) * | 3.14 (0.61) * | 2.79 (0.58) * | 4.53 (0.83) ^ | |
| PLA | 4.10 (0.70) | 3.39 (0.67) * | 3.19 (0.68) * | 2.95 (0.48) * | 2.75 (0.53) * | 4.45 (0.68) ^ | ||
| Fat Ox (g/min) | BRJ | 0.21 (0.15) | 0.41 (0.14) * | 0.41 (0.10) * | 0.50 (0.15) * | 0.67 (0.14) * | 0.25 (0.18) | |
| PLA | 0.15 (0.13) | 0.39 (0.16) * | 0.47 (0.21) * | 0.55 (0.10) * | 0.64 (0.12) * | 0.21 (0.15) | ||
| RER | BRJ | 0.96 (0.03) | 0.93 (0.03) * | 0.93 (0.02) * | 0.91 (0.03) * | 0.88 (0.03) * | 0.96 (0.04) | |
| PLA | 0.97 (0.02) | 0.93 (0.03) * | 0.91 (0.03) * | 0.90 (0.02) * | 0.89 (0.02) * | 0.96 (0.02) | ||
| RPE | BRJ | 11.1 (1.6) | 11.7 (1.2) | 12.4 (0.9) * | 13.2 (1.1) * | 15.8 (1.3) ^ | ||
| PLA | 11.0 (1.8) | 11.6 (1.3) | 12.5 (1.0) * | 13.1 (1.1) * | 15.6 (1.4) ^ | |||
| Heart Rate (bpm) | BRJ | 141 (8) | 154 (8) * | 154 (7) * | 156 (8) * | 162 (9) * | 172 (8) ^ | |
| PLA | 143 (7) | 154 (9) * | 154 (10) * | 156 (9) * | 158 (9) * | 167 (8) ^ | ||
| Lactate (mmol/L) | BRJ | 2.4 (0.9) * | 3.2 (0.8) | 2.0 (0.9) * | 2.1 (1.3) * | 1.5 (0.4) * | 3.7 (1.7) | |
| PLA | 2.2 (0.7) * | 2.9 (0.9) | 1.8 (0.4) * | 1.8 (0.8) * | 1.6 (0.6) * | 3.1 (0.8) |
| Pre Trial A | Pre Trial B | 14 d Intervention | Pre Trial C | |||||
|---|---|---|---|---|---|---|---|---|
| MAX (n = 9) | CON (n = 10) | MAX (n = 9) | CON (n = 10) | MAX (n = 9) | CON (n = 10) | MAX (n = 9) | ||
| Energy | kJ | 16,262 (2234) | 15,254 (2421) | 15,150 (766) | 15,183 (1786) | 16,018 (950) | 16,065 (2688) | 16,474 (1254) |
| kJ/kg | 238 (26) | 222 (28) | 223 (10) | 222 (7) | 234 (7) * | 232 (18) | 242 (13) | |
| CHO | g | 782 (130) #$^ | 526 (74) | 534 (30) | 454 (56) | 694 (49) * | 554 (82) | 778 (104) #$^ |
| g/kg | 11.4 (1.6) #$^ | 7.7 (0.9) | 7.9 (0.3) | 6.6 (0.2) | 10.1 (0.3) * | 8.0 (0.4) | 11.4 (1.3) #$^ | |
| Protein | g | 101 (12) #$^ | 133 (19) | 133 (9) | 147 (17) | 135 (8) | 140 (20) | 108 (11) #$^ |
| g/kg | 1.5 (0.1) #$^ | 1.8 (0.3) | 2.0 (0.1) | 2.1 (0.1) | 2.0 (0.0) | 2.0 (0.3) | 1.6 (0.2) #$^ | |
| Fat | g | 37 (5) #$^ | 102 (23) | 96 (6) | 127 (15) | 49 (6) * | 108 (26) | 42 (15) #$^ |
| g/kg | 0.5 (0.1) #$^ | 1.5 (0.3) | 1.4 (0.1) | 1.9 (0.1) | 0.7 (0.1) * | 1.6 (0.3) | 0.6 (0.2) #$^ | |
| Fibre | g | 25.9 (5.4) #$^ | 43.7 (7.0) | 42.5 (3.3) | 45.0 (4.9) | 47.9 (2.4) | 45.7 (9.0) | 25.8 (5.8) #$^ |
| CON (n = 10) | MAX (n = 9) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Km | Pre | 1 | 7 | 13 | 19 | 25 | 26 | Pre | 1 | 7 | 13 | 19 | 25 | 26 | |
| O2 mL/kg/ min | Trial A | 48.8 $ (4.1) | 48.5 $ (3.9) | 48.7 $ (3.8) | 49.4 $ (3.3) | 49.8 $ (3.4) | 56.6 (3.5) | ||||||||
| B | 46.4 $ (3.7) | 46.0 $ (4.0) | 46.5 $ (3.8) | 46.5 $ (3.7) | 47.9 $ (3.3) | 53.3 (3.6) | 48.7 $ (4.2) | 48.1 $ (3.7) | 48.5 $ (4.0) | 48.5 $ (2.8) | 48.7 $ (3.6) | 54.9 (3.8) | |||
| C | 46.6 $ (3.7) | 45.7 $ (3.2) | 46.1 $ (3.4) | 46.7 $ (3.5) | 47.5 (3.6) | 53.8 (3.6) | 49.3 $ (3.6) | 48.2 $ (2.8) | 48.5 $ (3.7) | 48.1 $ (3.2) | 48.7 $ (3.4) | 56.5 (4.1) | |||
| RER | A | 0.98 (0.03) | 0.96 $ (0.02) | 0.95 $ (0.02) | 0.94 $ (0.03) | 0.93 $ (0.02) | 1.00 (0.02) | ||||||||
| B | 0.97 (0.05) | 0.93 $ (0.03) | 0.92 $ (0.03) | 0.90 $ (0.02) | 0.88 $ (0.04) | 0.97 (0.03) | 0.97 (0.03) | 0.92 $ (0.03) | 0.92 $ (0.03) | 0.90 $ (0.01) | 0.88 $ (0.01) | 0.97 (0.02) | |||
| C | 0.96 (0.04) | 0.92 $ (0.03) | 0.91 $ (0.03) | 0.90 $ (0.02) | 0.87 $ (0.04) | 0.95 (0.05) | 0.96 (0.04) | 0.94 $ (0.03) | 0.94 $ (0.03) | 0.94 $^ (0.03) | 0.93 $^ (0.03) | 0.98 (0.02) | |||
| Heart Rate (bpm) | A | 146 $ (12) | 152 $ (10) | 155 $ (7) | 155 $ (8) | 159 $ (9) | 171 (9) | ||||||||
| B | 144 $ (12) | 153 $ (11) | 155 $ (10) | 157 $ (10) | 163 $ (9) | 174 (8) | 144 $ (9) | 154 $ (8) | 153 $ (9) | 155 $ (8) | 157 $ (8) | 169 (8) | |||
| C #^ | 151 $ (18) | 148 $ (11) | 148 $ (10) | 150 $ (10) | 155 $ (10) | 166 (9) | 138 $ (10) | 145 $ (6) | 144 $ (5) | 145 $ (6) | 149 $ (8) | 163 (5) | |||
| RPE | A | 10.6 $ (1.5) | 11.4 $ (1.0) | 12.4 $ (1.7) | 13.2 $ (1.8) | 14.7 (1.8) | |||||||||
| B | 11.8 $ (1.5) | 12.1 $ (1.1) | 12.9 $ (1.2) | 13.4 $ (1.5) | 16.7 (2.1) | 10.9 $ (2.0) | 11.2 $ (1.5) | 12.2 $ (1.2) | 12.9 $ (0.9) | 15.2 (1.3) | |||||
| C | 11.9 $ (1.1) | 12.2 $ (0.9) | 12.7 $ (1.2) | 13.7 $ (1.3) | 15.9 (1.1) | 11.4 $ (1.0) | 11.9 $ (0.9) | 12.6 $ (0.9) | 13.1 $ (0.9) | 15.4 (0.7) | |||||
| Serum glucose mmol/L | A | 4.4 (0.8) | 5.5 $ (0.5) | 6.9 * (0.5) | 6.8 * (1.0) | 7.2 * (0.8) | 6.6 * (1.2) | ||||||||
| B | 4.5 (0.7) | 4.5 $ (0.8) | 6.4 * (0.9) | 5.7 * (0.7) | 5.5 * (0.7) | 5.8 * (0.8) | 4.4 (0.6) | 4.3 $ (0.6) | 5.7 * (0.6) | 5.3 * (0.7) | 5.5 * (0.6) | 5.8 * (0.6) | |||
| C# | 3.6 (0.4) | 4.2 $ (0.7) | 5.8 * (1.1) | 5.4 * (1.0) | 5.0 * (0.5) | 5.9 * (1.8) | 3.7 (0.5) | 4.4 $ (0.6) | 5.7 * (1.2) | 5.9 * (1.6) | 6.2 * (1.3) | 6.2 * (0.9) | |||
| Blood lactate mmol/L | A | 2.5 (0.8) | 3.0 (0.9) | 1.9 $ (0.6) | 2.4 $ (0.8) | 1.7 $ (0.4) | 4.0 * (1.5) | ||||||||
| B | 2.3 (0.6) | 3.3 (1.1) | 1.9 $ (0.7) | 1.8 $ (0.8) | 1.6 $ (0.4) | 3.6 (0.9) | 2.1 (0.8) | 2.8 (1.2) | 1.6 $ (0.5) | 1.7 $ (0.6) | 1.9 $ (1.5) | 2.7 * (0.7) | |||
| C | 2.5 (0.7) | 3.0 (0.8) | 2.0 $ (0.6) | 1.9 $ (0.8) | 2.0 $ (0.9) | 2.9 (1.0) | 2.9 (0.8) | 3.5 (0.9) | 2.4 $ (1.1) | 1.8 $ (0.5) | 2.6 $ (1.3) | 3.9 * (1.8) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burke, L.M.; Hall, R.; Heikura, I.A.; Ross, M.L.; Tee, N.; Kent, G.L.; Whitfield, J.; Forbes, S.F.; Sharma, A.P.; Jones, A.M.; et al. Neither Beetroot Juice Supplementation nor Increased Carbohydrate Oxidation Enhance Economy of Prolonged Exercise in Elite Race Walkers. Nutrients 2021, 13, 2767. https://doi.org/10.3390/nu13082767
Burke LM, Hall R, Heikura IA, Ross ML, Tee N, Kent GL, Whitfield J, Forbes SF, Sharma AP, Jones AM, et al. Neither Beetroot Juice Supplementation nor Increased Carbohydrate Oxidation Enhance Economy of Prolonged Exercise in Elite Race Walkers. Nutrients. 2021; 13(8):2767. https://doi.org/10.3390/nu13082767
Chicago/Turabian StyleBurke, Louise M., Rebecca Hall, Ida A. Heikura, Megan L. Ross, Nicolin Tee, Georgina L. Kent, Jamie Whitfield, Sara F. Forbes, Avish P. Sharma, Andrew M. Jones, and et al. 2021. "Neither Beetroot Juice Supplementation nor Increased Carbohydrate Oxidation Enhance Economy of Prolonged Exercise in Elite Race Walkers" Nutrients 13, no. 8: 2767. https://doi.org/10.3390/nu13082767