
Effects of Randomized Controlled Infancy-Onset Dietary Intervention on Leukocyte Telomere Length—The Special Turku Coronary Risk Factor Intervention Project (STRIP)

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. The Special Turku Coronary Risk Factor Intervention Project Cohort
2.2. Intervention and Assessment of Diet, Smoking and Physical Activity
2.3. Physical Examination
2.4. Laboratory Measurements
2.5. Telomere Length Measurement
2.6. Genetic Risk Score Calculation
2.7. Statistical Analyses
3. Results
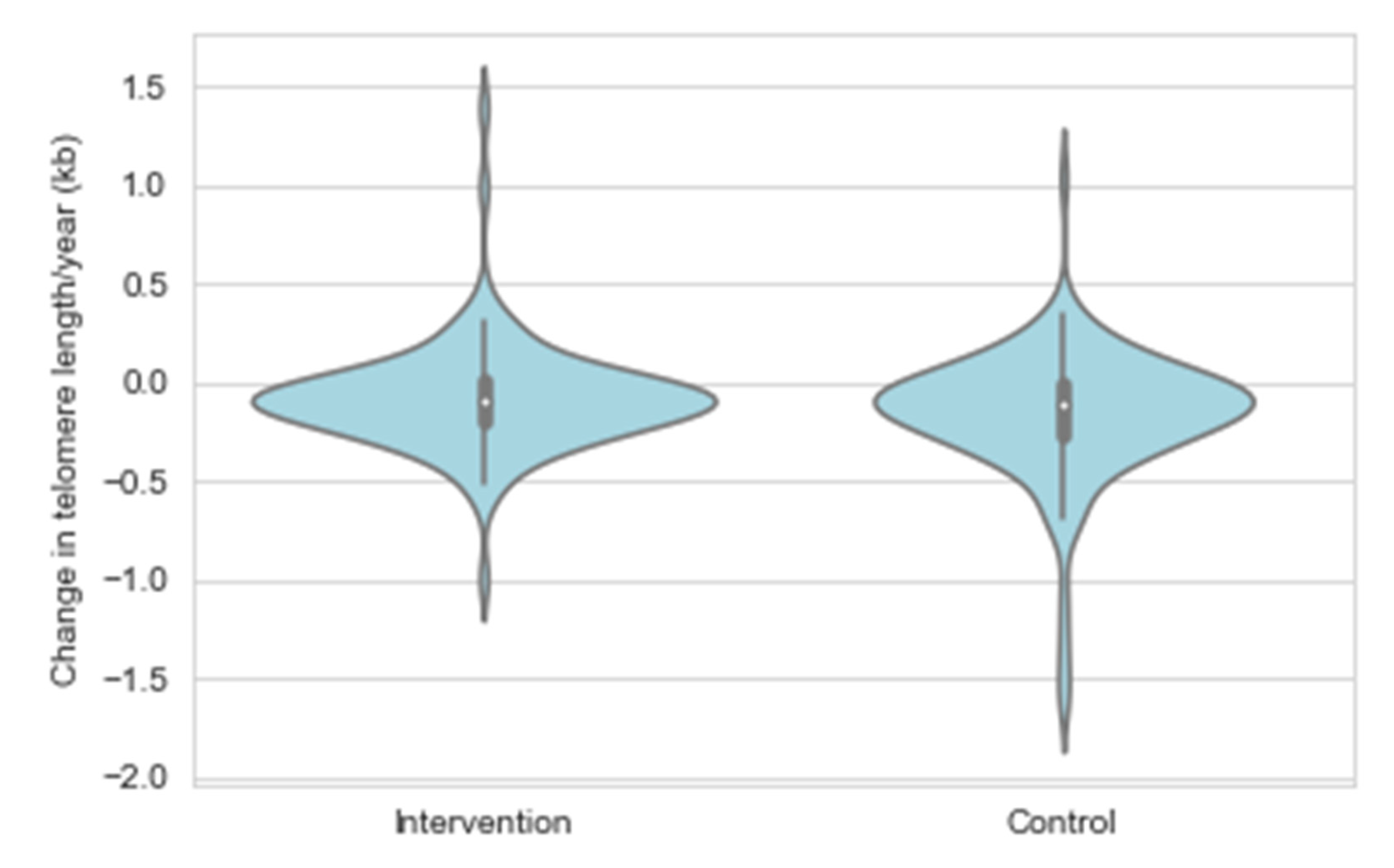
3.1. Characteristics of the Study Population, Telomere Length Attrition Rate and Yearly Change in the Short Telomere Proportion
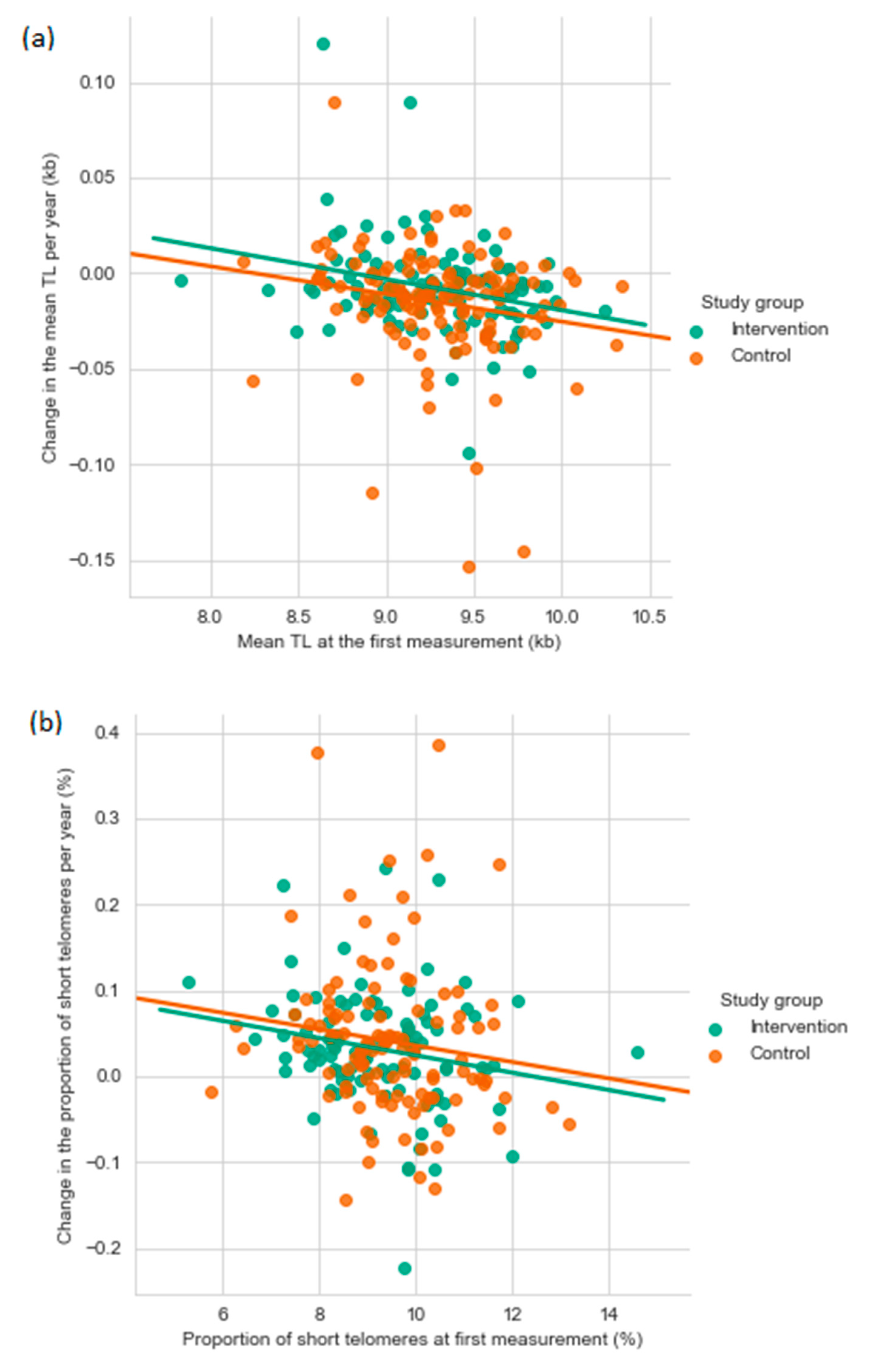
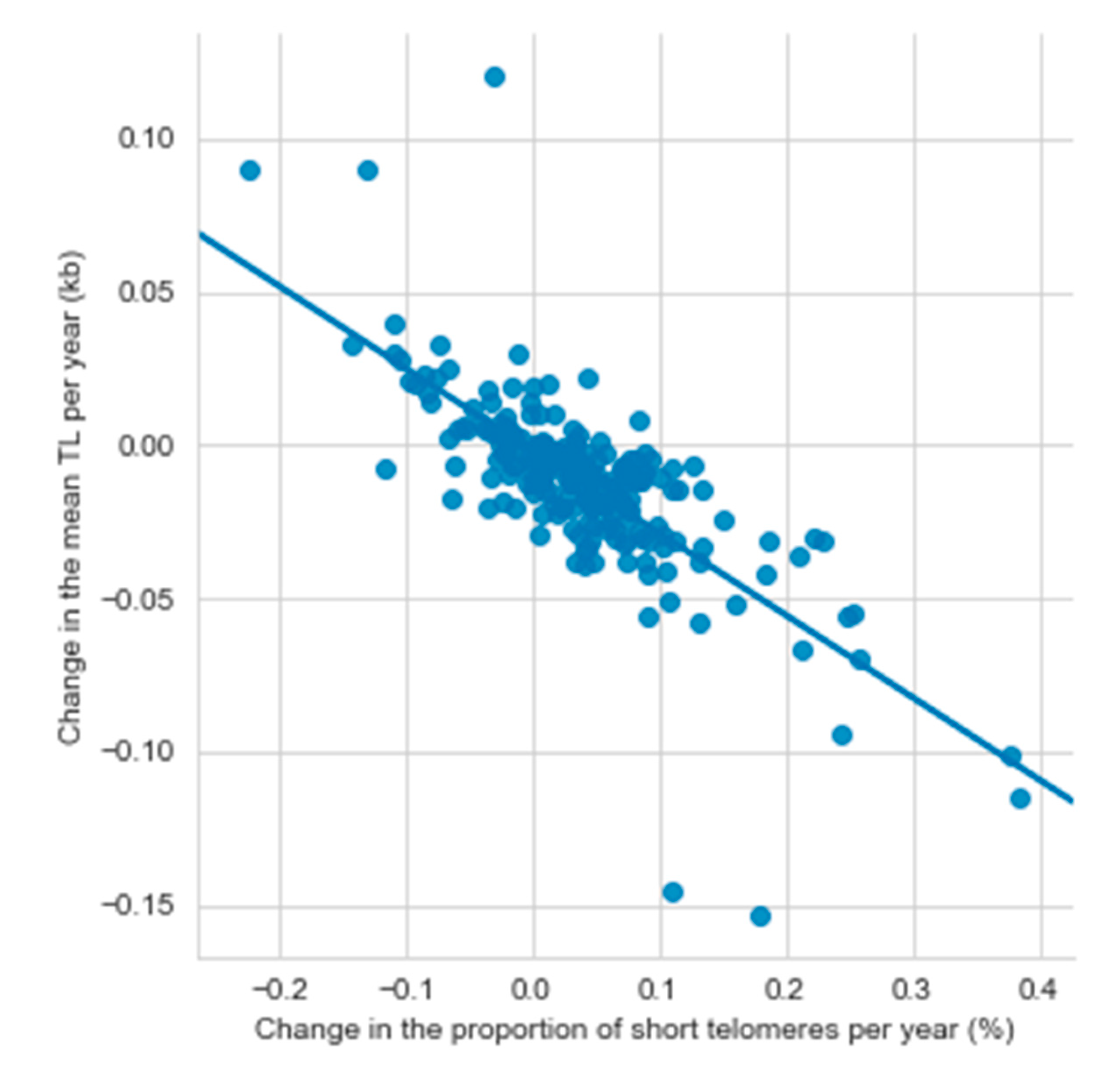
3.2. Established Telomere Length Determinants
3.3. Effect of STRIP Intervention on the Yearly TL Attrition Rate and the Yearly Change in the Short Telomere Proportion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, X.; Gutin, B.; Davis, C.L.; Keeton, D.; Thomas, J.; Stallmann-Jorgensen, I.; Mooken, G.; Bundy, V.; Snieder, H.; et al. Leukocyte telomere length in healthy Caucasian and African-American adolescents: Relationships with race, sex, adiposity, adipokines, and physical activity. J. Pediatr. 2011, 158, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Bardeguez, A.; Gardner, J.P.; Rodriguez, P.; Ganesh, V.; Kimura, M.; Skurnick, J.; Awad, G.; Aviv, A. Telomere length in the newborn. Pediatr. Res. 2002, 52, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Andrew, T.; Gardner, J.P.; Kimura, M.; Oelsner, E.; Cherkas, L.F.; Aviv, A.; Spector, T.D. Obesity, cigarette smoking, and telomere length in women. Lancet 2005, 366, 662–664. [Google Scholar] [CrossRef]
- Aviv, A.; Chen, W.; Gardner, J.P.; Kimura, M.; Brimacombe, M.; Cao, X.; Srinivasan, S.R.; Berenson, G.S. Leukocyte telomere dynamics: Longitudinal findings among young adults in the Bogalusa Heart Study. Am. J. Epidemiol. 2009, 169, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of seven loci affecting mean telomere length and their association with disease. Nat. Genet. 2013, 45, 427. [Google Scholar] [CrossRef]
- Njajou, O.T.; Cawthon, R.M.; Damcott, C.M.; Wu, S.H.; Ott, S.; Garant, M.J.; Blackburn, E.H.; Mitchell, B.D.; Shuldiner, A.R.; Hsueh, W.C. Telomere length is paternally inherited and is associated with parental lifespan. Proc. Natl. Acad. Sci. USA 2007, 104, 12135–12139. [Google Scholar] [CrossRef]
- Broer, L.; Codd, V.; Nyholt, D.R.; Deelen, J.; Mangino, M.; Willemsen, G.; Albrecht, E.; Amin, N.; Beekman, M.; De Geus, E.J.; et al. Meta-analysis of telomere length in 19,713 subjects reveals high heritability, stronger maternal inheritance and a paternal age effect. Eur. J. Hum. Genet. 2013, 21, 1163–1168. [Google Scholar] [CrossRef]
- Sun, Q.; Shi, L.; Prescott, J.; Chiuve, S.E.; Hu, F.B.; De Vivo, I.; Stampfer, M.J.; Franks, P.W.; Manson, J.E.; Rexrode, K.M. Healthy lifestyle and leukocyte telomere length in U.S. women. PLoS ONE 2012, 7, e38374. [Google Scholar] [CrossRef]
- D’Mello, M.J.; Ross, S.A.; Briel, M.; Anand, S.S.; Gerstein, H.; Pare, G. Association between shortened leukocyte telomere length and cardiometabolic outcomes: Systematic review and meta-analysis. Circ. Cardiovasc. Genet. 2015, 8, 82–90. [Google Scholar] [CrossRef]
- Barrett, J.H.; Iles, M.M.; Dunning, A.M.; Pooley, K.A. Telomere length and common disease: Study design and analytical challenges. Hum. Genet. 2015, 134, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, A.L.; Kronmal, R.A.; Gardner, J.P.; Psaty, B.M.; Jenny, N.S.; Tracy, R.P.; Walston, J.; Kimura, M.; Aviv, A. Leukocyte telomere length and cardiovascular disease in the cardiovascular health study. Am. J. Epidemiol. 2007, 165, 14–21. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- De Meyer, T.; Rietzschel, E.R.; De Buyzere, M.L.; Van Criekinge, W.; Bekaert, S. Telomere length and cardiovascular aging: The means to the ends? Ageing Res. Rev. 2011, 10, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; De Vivo, I.; Liu, Y.; Han, J.; Prescott, J.; Hunter, D.J.; Rimm, E.B. Associations between diet, lifestyle factors, and telomere length in women. Am. J. Clin. Nutr. 2010, 91, 1273–1280. [Google Scholar] [CrossRef]
- Nettleton, J.A.; Diez-Roux, A.; Jenny, N.S.; Fitzpatrick, A.L.; Jacobs, D.R., Jr. Dietary patterns, food groups, and telomere length in the Multi-Ethnic Study of Atherosclerosis (MESA). Am. J. Clin. Nutr. 2008, 88, 1405–1412. [Google Scholar] [CrossRef]
- De Meyer, T.; Bekaert, S.; De Buyzere, M.L.; De Bacquer, D.D.; Langlois, M.R.; Shivappa, N.; Hébert, J.R.; Gillebert, T.C.; Rietzschel, E.R.; Huybrechts, I. Leukocyte telomere length and diet in the apparently healthy, middle-aged Asklepios population. Sci. Rep. 2018, 8, 6540. [Google Scholar] [CrossRef]
- Song, Y.; You, N.C.; Song, Y.; Kang, M.K.; Hou, L.; Wallace, R.; Eaton, C.B.; Tinker, L.F.; Liu, S. Intake of small-to-medium-chain saturated fatty acids is associated with peripheral leukocyte telomere length in postmenopausal women. J. Nutr. 2013, 143, 907–914. [Google Scholar] [CrossRef]
- Garcia-Calzon, S.; Moleres, A.; Martinez-Gonzalez, M.A.; Martinez, J.A.; Zalba, G.; Marti, A. Dietary total antioxidant capacity is associated with leukocyte telomere length in a children and adolescent population. Clin. Nutr. 2015, 34, 694–699. [Google Scholar] [CrossRef]
- Crous-Bou, M.; Fung, T.T.; Prescott, J.; Julin, B.; Du, M.; Sun, Q.; Rexrode, K.M.; Hu, F.B.; De Vivo, I. Mediterranean diet and telomere length in Nurses’ Health Study: Population based cohort study. BMJ 2014, 349, g6674. [Google Scholar] [CrossRef]
- García-Calzón, S.; Martínez-González, M.A.; Razquin, C.; Arós, F.; Lapetra, J.; Martínez, J.A.; Zalba, G.; Marti, A. Mediterranean diet and telomere length in high cardiovascular risk subjects from the PREDIMED-NAVARRA study. Clin. Nutr. 2016, 35, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Tiainen, A.M.; Männistö, S.; Blomstedt, P.A.; Moltchanova, E.; Perälä, M.M.; Kaartinen, N.E.; Kajantie, E.; Kananen, L.; Hovatta, I.; Eriksson, J.G. Leukocyte telomere length and its relation to food and nutrient intake in an elderly population. Eur. J. Clin. Nutr. 2012, 66, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Simell, O.; Niinikoski, H.; Rönnemaa, T.; Raitakari, O.T.; Lagström, H.; Laurinen, M.; Aromaa, M.; Hakala, P.; Jula, A.; Jokinen, E.; et al. Cohort Profile: The STRIP Study (Special Turku Coronary Risk Factor Intervention Project), an Infancy-onset Dietary and Life-style Intervention Trial. Int. J. Epidemiol. 2009, 38, 650–655. [Google Scholar] [CrossRef]
- Oranta, O.; Pahkala, K.; Ruottinen, S.; Niinikoski, H.; Lagström, H.; Viikari, J.S.; Jula, A.; Loo, B.M.; Simell, O.; Rönnemaa, T.; et al. Infancy-onset dietary counseling of low-saturated-fat diet improves insulin sensitivity in healthy adolescents 15–20 years of age: The Special Turku Coronary Risk Factor Intervention Project (STRIP) study. Diabetes Care 2013, 36, 2952–2959. [Google Scholar] [CrossRef] [PubMed]
- Niinikoski, H.; Pahkala, K.; Ala-Korpela, M.; Viikari, J.; Rönnemaa, T.; Lagström, H.; Jokinen, E.; Jula, A.; Savolainen, M.J.; Simell, O.; et al. Effect of repeated dietary counseling on serum lipoproteins from infancy to adulthood. Pediatrics 2012, 129, e704–e713. [Google Scholar] [CrossRef]
- Niinikoski, H.; Jula, A.; Viikari, J.; Rönnemaa, T.; Heino, P.; Lagström, H.; Jokinen, E.; Simell, O. Blood pressure is lower in children and adolescents with a low-saturated-fat diet since infancy: The special turku coronary risk factor intervention project. Hypertension 2009, 53, 918–924. [Google Scholar] [CrossRef]
- Raitakari, O.T.; Rönnemaa, T.; Jarvisalo, M.J.; Kaitosaari, T.; Volanen, I.; Kallio, K.; Lagström, H.; Jokinen, E.; Niinikoski, H.; Viikari, J.S.A.; et al. Endothelial function in healthy 11-year-old children after dietary intervention with onset in infancy: The Special Turku Coronary Risk Factor Intervention Project for children (STRIP). Circulation 2005, 112, 3786–3794. [Google Scholar] [CrossRef] [PubMed]
- Nupponen, M.; Pahkala, K.; Juonala, M.; Magnussen, C.G.; Niinikoski, H.; Rönnemaa, T.; Viikari, J.S.; Saarinen, M.; Lagström, H.; Jula, A.; et al. Metabolic syndrome from adolescence to early adulthood: Effect of infancy-onset dietary counseling of low saturated fat: The Special Turku Coronary Risk Factor Intervention Project (STRIP). Circulation 2015, 131, 605–613. [Google Scholar] [CrossRef]
- Nordic Nutrition Recommendations. Integrating Nutrition and Physical Activity, 4th ed.; Nordic Council of Ministers: Copenhagen, Denmark, 2004. [Google Scholar]
- Cole, T.J.; Bellizzi, M.C.; Flegal, K.M.; Dietz, W.H. Establishing a standard definition for child overweight and obesity worldwide: International survey. BMJ 2000, 320, 1240–1243. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Niinikoski, H.; Lagström, H.; Jokinen, E.; Siltala, M.; Rönnemaa, T.; Viikari, J.; Raitakari, O.T.; Jula, A.; Marniemi, J.; Näntö-Salonen, K.; et al. Impact of repeated dietary counseling between infancy and 14 years of age on dietary intakes and serum lipids and lipoproteins: The STRIP study. Circulation 2007, 116, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Kostner, G.M. Letter: Enzymatic determination of cholesterol in high-density lipoprotein fractions prepared by polyanion precipitation. Clin. Chem. 1976, 22, 695. [Google Scholar] [CrossRef] [PubMed]
- Fyhrquist, F.; Tiitu, A.; Saijonmaa, O.; Forsblom, C.; Groop, P.H.; FinnDiane Study Group. Telomere length and progression of diabetic nephropathy in patients with type 1 diabetes. J. Intern. Med. 2010, 267, 278–286. [Google Scholar] [CrossRef]
- Voight, B.F.; Kang, H.M.; Ding, J.; Palmer, C.D.; Sidore, C.; Chines, P.S.; Burtt, N.P.; Fuchsberger, C.; Li, Y.; Erdmann, J.; et al. The metabochip, a custom genotyping array for genetic studies of metabolic, cardiovascular, and anthropometric traits. PLoS Genet. 2012, 8, e1002793. [Google Scholar] [CrossRef] [PubMed]
- Delaneau, O.; Marchini, J.; Zagury, J.F. A linear complexity phasing method for thousands of genomes. Nat. Methods 2011, 9, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Howie, B.N.; Donnelly, P.; Marchini, J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009, 5, e1000529. [Google Scholar] [CrossRef]
- Durbin, R.M.; Abecasis, G.R.; Altshuler, D.L.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Gibbs, R.A.; Hurles, M.E.; McVean, G.A. A map of human genome variation from population-scale sequencing. Nature 2010, 467, 1061–1073. [Google Scholar]
- Demanelis, K.; Jasmine, F.; Chen, L.S.; Chernoff, M.; Tong, L.; Delgado, D.; Zhang, C.; Shinkle, J.; Sabarinathan, M.; Lin, H.; et al. Determinants of telomere length across human tissues. Science 2020, 369, eaaz6876. [Google Scholar] [CrossRef]
- Garcia-Calzon, S.; Moleres, A.; Marcos, A.; Campoy, C.; Moreno, L.A.; Azcona-Sanjulián, M.C.; Martínez-González, M.A.; Martínez, J.A.; Zalba, G.; Marti, A.; et al. Telomere length as a biomarker for adiposity changes after a multidisciplinary intervention in overweight/obese adolescents: The EVASYON study. PLoS ONE 2014, 9, e89828. [Google Scholar] [CrossRef]
- Hovatta, I.; de Mello, V.D.; Kananen, L.; Lindström, J.; Eriksson, J.G.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Peltonen, M.; Tuomilehto, J.; Uusitupa, M. Leukocyte telomere length in the Finnish Diabetes Prevention Study. PLoS ONE 2012, 7, e34948. [Google Scholar] [CrossRef]
- Kark, J.D.; Goldberger, N.; Kimura, M.; Sinnreich, R.; Aviv, A. Energy intake and leukocyte telomere length in young adults. Am. J. Clin. Nutr. 2012, 95, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Meinilä, J.; Perälä, M.M.; Kautiainen, H.; Männistö, S.; Kanerva, N.; Shivappa, N.; Hébert, J.R.; Iozzo, P.; Guzzardi, M.A.; Eriksson, J.G. Healthy diets and telomere length and attrition during a 10-year follow-up. Eur. J. Clin. Nutr. 2019, 73, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Balan, E.; Decottignies, A.; Deldicque, L. Physical Activity and Nutrition: Two Promising Strategies for Telomere Maintenance? Nutrients 2018, 10, 1942. [Google Scholar] [CrossRef]
- Masi, S.; Nightingale, C.M.; Day, I.N.; Guthrie, P.; Rumley, A.; Lowe, G.D.; von Zglinicki, T.; D’Aiuto, F.; Taddei, S.; Klein, N.; et al. Inflammation and not cardiovascular risk factors is associated with short leukocyte telomere length in 13- to 16-year-old adolescents. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Buxton, J.L.; Walters, R.G.; Visvikis-Siest, S.; Meyre, D.; Froguel, P.; Blakemore, A.I. Childhood obesity is associated with shorter leukocyte telomere length. J. Clin. Endocrinol. Metab. 2011, 96, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Price, L.H.; Kao, H.T.; Burgers, D.E.; Carpenter, L.L.; Tyrka, A.R. Telomeres and early-life stress: An overview. Biol. Psychiatry 2013, 73, 15–23. [Google Scholar] [CrossRef]
- Frenck, R.W., Jr.; Blackburn, E.H.; Shannon, K.M. The rate of telomere sequence loss in human leukocytes varies with age. Proc. Natl. Acad. Sci. USA 1998, 95, 5607–5610. [Google Scholar] [CrossRef]
- Hemann, M.T.; Strong, M.A.; Hao, L.Y.; Greider, C.W. The shortest telomere, not average telomere length, is critical for cell viability and chromosome stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Measurement | Second Measurement | |||
|---|---|---|---|---|
| Intervention | Control | Intervention | Control | |
| Age, years | 7.50 ± 2.00 | 7.48 ± 2.00 | 19.8 ± 0.46 | 19.8 ± 0.44 |
| BMI (kg/m2) | 16.3 ± 2.00 | 16.3 ± 3.21 | 23.0 ± 3.50 | 23.0 ± 4.34 |
| Overweight/obese (%) | 16.7 | 14.5 | 18.5 | 21.8 |
| Underweight (%) | 6.5 | 15.3 | 4.6 | 8.1 |
| Waist circumference (cm) | 56.1 ± 5.45 | 56.4 ± 7.86 | 78.3 ± 9.00 | 77.7 ± 10.6 |
| Systolic blood pressure (mmHg) | 102 ± 9.56 | 101 ± 10.6 | 123 ± 12.0 | 120 ± 14.4 |
| Diastolic blood pressure (mmHg) | 59.0 ± 6.38 | 58.5 ± 6.55 | 66.8 ± 7.46 | 65.7 ± 8.16 |
| Total cholesterol (mmol/L) | 4.40 ± 0.65 | 4.56 ± 0.67 | 4.30 ± 0.76 | 4.36 ± 0.78 |
| HDL-C (mmol/L) | 1.28 ± 0.23 | 1.33 ± 0.25 | 1.31 ± 0.33 | 1.34 ± 0.33 |
| LDL-C (mmol/L) | 2.79 ± 0.53 | 2.90 ± 0.60 | 2.49 ± 0.58 | 2.54 ± 0.67 |
| Triglycerides (mmol/L | 0.69 ± 0.30 | 0.68 ± 0.26 | 1.11 ± 0.53 | 1.05 ± 0.52 |
| Fasting plasma glucose (mmol/L) | 4.47 ± 0.52 | 4.48 ± 0.46 | 4.96 ± 1.10 | 4.85 ± 0.63 |
| Fasting serum insulin (mU/L) | 4.61 ± 1.29 | 5.22 ± 3.60 | 11.4 ± 37.6 | 8.38 ± 9.73 |
| First Measurement | Second Measurement | ||||||
|---|---|---|---|---|---|---|---|
| Intervention | Control | p-Value | Intervention | Control | p-Value | ||
| Mean telomere length (mean, SD) | Model 1 | 9.27 (0.41) | 9.29 (0.39) | 0.81 | 9.16 (0.42) | 9.12 (0.44) | 0.42 |
| Model 2 | 9.30 (0.38) | 9.29 (0.37) | 0.49 | 9.19 (0.42) | 9.11 (0.44) | 0.080 | |
| Short telomere proportion (%) | Model 1 | 9.25 (1.38) | 9.44 (1.27) | 0.27 | 9.66 (1.44) | 9.91 (1.46) | 0.21 |
| Model 2 | 9.17 (1.36) | 9.37 (1.19) | 0.064 | 9.54 (1.29) | 9.89 (1.49) | 0.019 | |
| Mean TL | Short Telomere Proportion | ||||||
|---|---|---|---|---|---|---|---|
| β Estimate (SE) | p-Value | N (Intervention/ Control) | β Estimate (SE) | p-Value | N (Intervention/ Control) | ||
| Model 1 | Study group, age and sex | 0.007 (0.004) | 0.040 | 108/124 | −0.011 (0.011) | 0.310 | 99/114 |
| Model 2 | Model 1 + (P + M)/S-ratio + fiber intake | 0.011 (0.005) | 0.023 | 73/97 | −0.022 (0.015) | 0.146 | 67/88 |
| Model 3 | Model 1 + total cholesterol + apoB + LDL cholesterol | 0.006 (0.003) | 0.042 | 108/124 | −0.011 (0.011) | 0.332 | 99/114 |
| Model 4 | Model 1 + insulin | 0.007 (0.040) | 0.039 | 108/124 | −0.011 (0.011) | 0.308 | 99/114 |
| Model 5 | Model 1 + systolic blood pressure | 0.007 (0.004) | 0.044 | 108/124 | −0.011 (0.011) | 0.327 | 99/114 |
| Model 6 | Model 1 + physical activity + smoking | 0.008 (0.004) | 0.027 | 105/119 | −0.014 (0.011) | 0.221 | 96/111 |
| Model 7 | All models combined | 0.013 (0.005) | 0.009 | 71/93 | −0.029 (0.015) | 0.065 | 65/85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pitkänen, N.; Pahkala, K.; Rovio, S.P.; Saijonmaa, O.J.; Nyman, A.E.; Jula, A.; Lagström, H.; Viikari, J.S.A.; Rönnemaa, T.; Niinikoski, H.; et al. Effects of Randomized Controlled Infancy-Onset Dietary Intervention on Leukocyte Telomere Length—The Special Turku Coronary Risk Factor Intervention Project (STRIP). Nutrients 2021, 13, 318. https://doi.org/10.3390/nu13020318
Pitkänen N, Pahkala K, Rovio SP, Saijonmaa OJ, Nyman AE, Jula A, Lagström H, Viikari JSA, Rönnemaa T, Niinikoski H, et al. Effects of Randomized Controlled Infancy-Onset Dietary Intervention on Leukocyte Telomere Length—The Special Turku Coronary Risk Factor Intervention Project (STRIP). Nutrients. 2021; 13(2):318. https://doi.org/10.3390/nu13020318
Chicago/Turabian StylePitkänen, Niina, Katja Pahkala, Suvi P. Rovio, Outi J. Saijonmaa, Anna E. Nyman, Antti Jula, Hanna Lagström, Jorma S. A. Viikari, Tapani Rönnemaa, Harri Niinikoski, and et al. 2021. "Effects of Randomized Controlled Infancy-Onset Dietary Intervention on Leukocyte Telomere Length—The Special Turku Coronary Risk Factor Intervention Project (STRIP)" Nutrients 13, no. 2: 318. https://doi.org/10.3390/nu13020318
APA StylePitkänen, N., Pahkala, K., Rovio, S. P., Saijonmaa, O. J., Nyman, A. E., Jula, A., Lagström, H., Viikari, J. S. A., Rönnemaa, T., Niinikoski, H., Simell, O., Fyhrquist, F., & Raitakari, O. T. (2021). Effects of Randomized Controlled Infancy-Onset Dietary Intervention on Leukocyte Telomere Length—The Special Turku Coronary Risk Factor Intervention Project (STRIP). Nutrients, 13(2), 318. https://doi.org/10.3390/nu13020318