A Review on Sources and Pharmacological Aspects of Sakuranetin


Abstract
1. Introduction
2. Sources of Sakuranetin
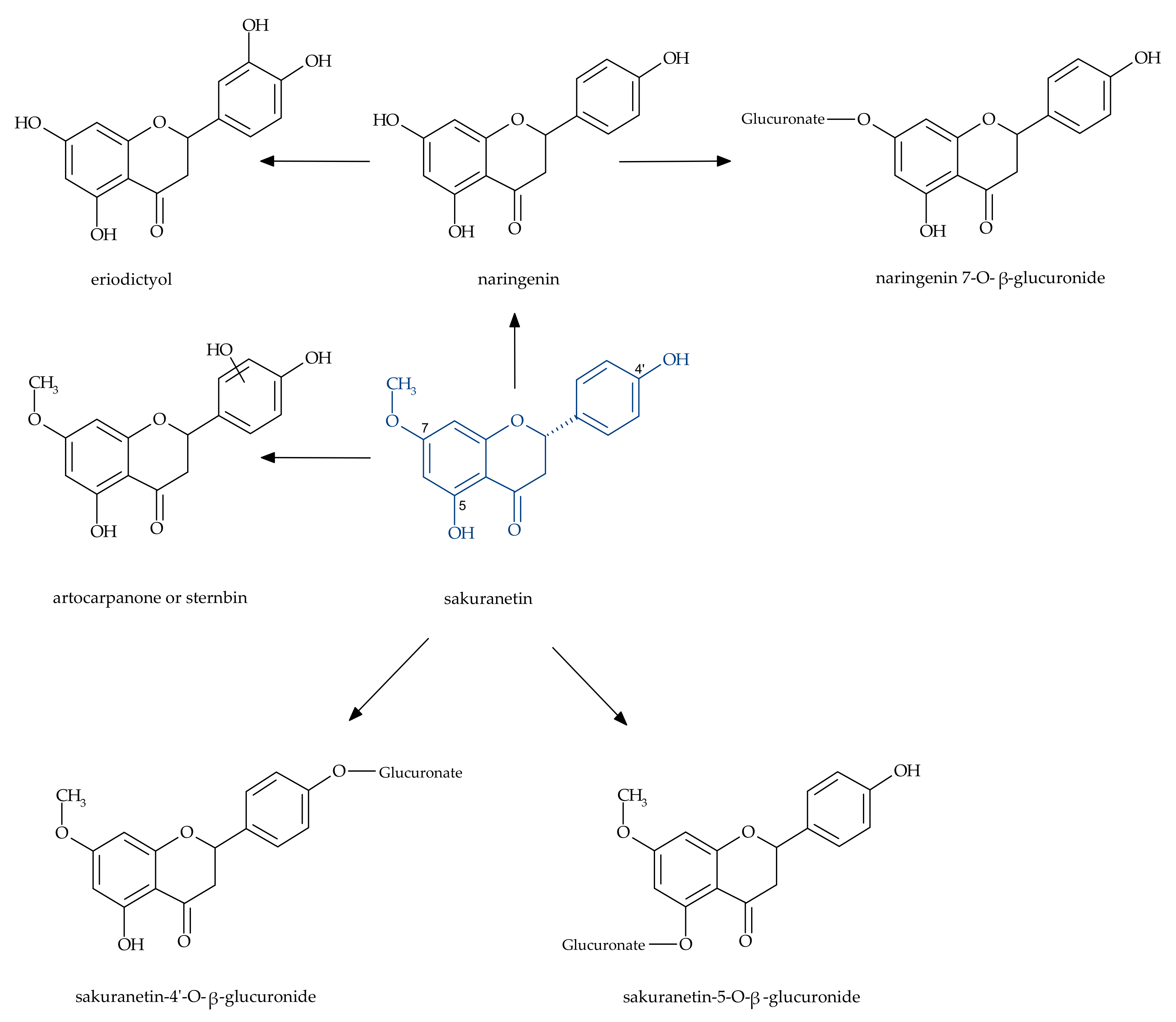
3. Metabolism of Sakuranetin in the Human Body
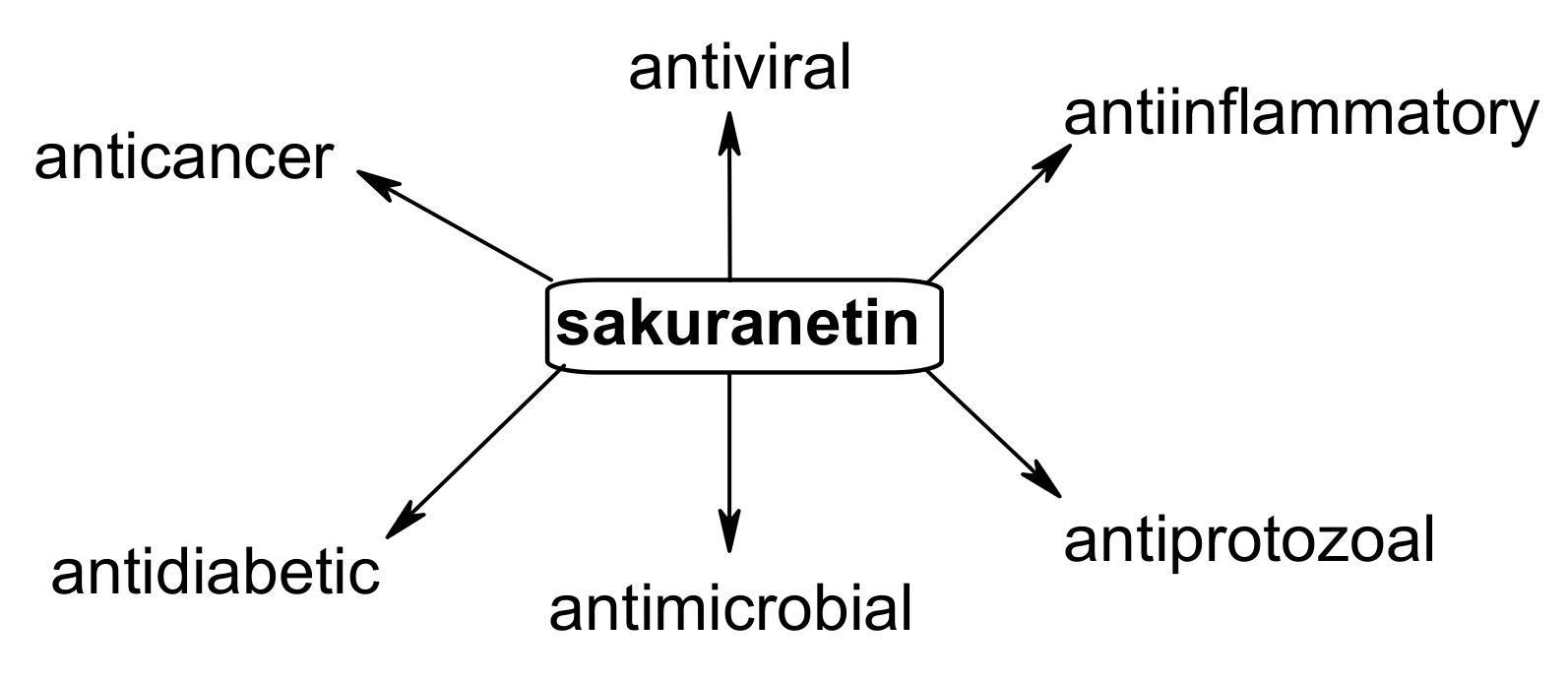
4. Biological Potential of Sakuranetin
4.1. Anticancer Effects
4.2. Antimicrobial Activity
4.3. Antiprotozoal Properties
4.4. Antiviral Activity
4.5. Antiinflammatory Activity
4.6. Beneficial Role of Sakuranetin in Alzheimer’s Disease (AD)
4.7. Other Effects of Sakuranetin
5. Conclusions
Funding
Conflicts of Interest
References
- Benkherouf, A.Y.; Soini, S.L.; Stompor, M.; Uusi-Oukari, M. Possitive allosteric modulation of native and recombinant GABAA receptors by hops prenylflavonoids. Eur. J. Pharmacol. 2019, 852, 34–41. [Google Scholar] [CrossRef]
- Stompor, M.; Świtalska, M.; Wietrzyk, J. The influence of a single and double biotinylation of xanthohumol on its anticancer activity. Acta Biochim. Pol. 2019, 66, 559–565. [Google Scholar] [CrossRef]
- Li, F.; Yao, Y.; Huang, H.; Hao, H.; Ying, M. Xanthohumol attenuates cisplatin-induced nephrotoxicity through inhibiting NF-ĸB and activating Nrf2 signaling pathways. Int. Immunopharmacol. 2018, 61, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Stompor, M.; Żarowska, B. Antimicrobial activity of xanthohumol and its selected structural analogues. Molecules 2016, 21, 608. [Google Scholar] [CrossRef]
- Berim, A.; Gang, D.R. Methoxylated flavones: Occurrence, importance, biosynthesis. Phytochem. Rev. 2016, 15, 363–390. [Google Scholar] [CrossRef]
- Miyazawa, M.; Kinoshita, H.; Okuno, Y. Antimutagenic activity of sakuranetin from Prunus Jamasakura. J. Food Sci. 2003, 68, 52–56. [Google Scholar] [CrossRef]
- Kokubun, T.; Harborne, J.B. Phytoalexin induction in the sapwood of plants of the Maloideae (Rosaceae) biphenyls or dibenzofurans. Phytochemistry 1995, 40, 1649–1654. [Google Scholar] [CrossRef]
- Kodama, O.; Miyakawa, J.; Akatsuka, T.; Kiyosawa, S. Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves. Phytochemistry 1992, 31, 3807–3809. [Google Scholar] [CrossRef]
- Dillon, V.M.; Overton, J.; Grayer, R.J.; Harborne, J.B. Differences in phytoalexin response among rice cultivars of different resistance to blast. Phytochemistry 1997, 44, 599–603. [Google Scholar] [CrossRef]
- Rakwal, R.; Hasegawa, M.; Kodama, O. A methyltransferase for synthesis of the flavanone phytoalexin sakuranetin in rice leaves. Biochem. Biophys. Res. Commun. 1996, 222, 732–735. [Google Scholar] [CrossRef]
- Duan, L.; Liu, H.; Li, X.; Xiao, J.; Wang, S. Multiple phytohormones and phytoalexins are involved in disease resistance to Magnaporthe oryzae invaded from roots in rice. Physiol. Plant. 2014, 152, 486–500. [Google Scholar] [CrossRef]
- Tomogami, S.; Rakwal, R.; Kodama, O. Phytoalexin production by amino acid conjugates of jasmonic acid through induction of naringenin-7-O-methyltransferase, a key enzyme on phytoalexin biosynthesis in rice (Oryza sativa L.). FEBS Lett. 1997, 401, 239–242. [Google Scholar] [CrossRef]
- Aft, H. Chemistry of dihydroquercetin. I. Acetate derivatives. J. Org. Chem. 1961, 26, 1958–1963. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, B.G.; Ahn, J.H. Biosynthesis of bioactive O-methylated flavonoids in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 195–204. [Google Scholar] [CrossRef]
- Asahina, Y.; Shinoda, J.; Inubuse, M. Uber sakuranin. J. Pharm. Soc. Jpn. 1927, 550, 133–139. [Google Scholar]
- Narasimhachabi, N.; Seshadbi, T.R. A note on the components of the bark of Prunus puddum. Proc. Indian Acad. Sci. A 1949, 30, 274–276. [Google Scholar]
- Asahina, Y. Ueber das sakuranin, ein neues glykosid der rinde von Prunus pseudo-Cerasus lindl. var. sieboldi Maxim. Arch. Pharm. 1908, 246, 59–72. [Google Scholar] [CrossRef]
- Grecco, D.S.; Dorigueto, A.C.; Landre, I.M.; Soares, M.G.; Martho, K.; Lima, R.; Pascon, R.C.; Vallim, M.A.; Capello, T.M.; Romoff, P.; et al. Structural crystalline characterization of sakuranetin—An antimicrobial flavanone from twigs of Baccharis retusa (Asteraceae). Molecules 2014, 19, 7528–7542. [Google Scholar] [CrossRef]
- Atkinson, P.; Blakeman, J.P. Seasonal occurrence of an antimicrobial flavanone, sakuranetin, associated with glands on leaves of Ribes nigrum. New Phytol. 1982, 92, 63–74. [Google Scholar] [CrossRef]
- Ghisalberti, E.L.; Jefferies, P.R.; Lanteri, R.; Matisons, J. Constituents of propolis. Experientia 1978, 34, 157–158. [Google Scholar] [CrossRef]
- Agrwal, V.K.; Thappa, R.K.; Agrwal, S.G.; Ghar, K.L. Phenolic constituents of Iris milessi rhizomes. Phytochemistry 1984, 23, 1342–1343. [Google Scholar] [CrossRef]
- Liu, Y.L.; Ho, D.K.; Cassady, J.M.; Cook, V.M.; Baird, W.M. Isolation of potential cancer chemopreventive agents from Eriodictyon californicum. J. Nat. Prod. 1992, 55, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Messana, I.; Ferrari, F.; de Moraes e Souza, M.A.; Gács-Baitz, E. (-)-Salzol, an isopimarane diterpene, and a chalcone from Hyptis salzmanii. Phytochemistry 1990, 29, 329–332. [Google Scholar] [CrossRef]
- Bashir, A.K. Methylated flavones of Teucrium stocksianum. J. Herbs Spices Med. Plants 1995, 3, 17–24. [Google Scholar] [CrossRef]
- Melo, M.; Oliveira, A.P.; Wiecikowski, A.F.; Castro, J.L.; De Oliveira, A.A.G.; Pereira, H.M.G.; Da Veiga, V.; Capella, M.M.A.; Rocha, L.; Holandino, C. Phenolic compounds from Viscum album tinctures enhanced antitumor activity in melanoma murine cancer cells. Saudi Pharm. J. 2018, 26, 311–322. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, W.G.; Mesquita, A.A.L.; Kubitzki, K.; Gottlieb, O.R. Xanthones from Bonnetia dinizii. Phytochemistry 1990, 29, 1893–1894. [Google Scholar] [CrossRef]
- Greenaway, W.; English, S.; May, J.; Whatley, F.R. Analysis of phenolic of bud exudate of Populus sieboldii by GC-MS. Phytochemistry 1991, 30, 3005–3008. [Google Scholar] [CrossRef]
- Rojas, A.; Cruz, S.; Ponce-Monter, H.; Mata, R. Smooth muscle relaxing compounds from Dodonaea viscosa. Planta Med. 1996, 62, 154–159. [Google Scholar] [CrossRef]
- Aires, A.; Dias, C.; Carvalho, R.; Saavedra, M.J. Analysis of glycosylated flavonoids extracted from sweet-cherry stems, as antibacterial agents against pathogenic Escherichia coli isolates. Acta Biochim. Pol. 2017, 64, 265–271. [Google Scholar] [CrossRef]
- Zhang, L.B.; Ji, J.; Lei, C.; Wang, H.Y.; Zhao, Q.S.; Hou, A.J. Isoprenylated flavonoid and adipogenesis-promoting constituents of Dodonaea viscosa. J. Nat. Prod. 2012, 75, 699–706. [Google Scholar] [CrossRef]
- Ogawa, Y.; Oku, H.; Iwaoka, E.; Inuma, M.; Ishiguro, K. Allergy-preventive flavonoids from Xanthorrhoea hastilis. Chem. Pharm. Bull. 2007, 55, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, X.A.; De La Fuente, E.R. Sakuranetin and pulcherryl acetate from Eupatorium havanense. Phytochemistry 1973, 12, 2060. [Google Scholar] [CrossRef]
- De Pascual, T.; González, M.S.; Muriel, M.R.; Bellido, I.S. Phenolic derivatives from Artemisia campestris subsp. glutinosa. Phytochemistry 1984, 23, 1819–1821. [Google Scholar] [CrossRef]
- Liang, S.; Tian, J.M.; Feng, Y.; Liu, X.H.; Xiong, Z.; Zhang, W.D. Flavonoids from Daphne aurantiaca and their inhibitory activities against nitric oxide production. Chem. Pharm. Bull. 2011, 59, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Ticha, L.A.; Klaasen, J.A.; Green, I.R.; Naidoo, S.; Baker, B.; Pietersen, R.D. Phytochemical and antimicrobial screening of flavanones and chalcones from Galenia africana and Dicerothamnus rhinocerotis. Nat. Prod. Commun. 2015, 10, 1185–1190. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Saleri, F.; Guo, M. Analysis of flavonoids in Rhamnus davurica and its antiproliferative activities. Molecules 2016, 21, 1275. [Google Scholar] [CrossRef]
- Hong, L.; Ying, S.H. Ethanol extract and isolated constituents from Artemisia dracunculus inhibit esophageal squamous cell carcinoma and induce apoptotic cell death. Drug Res. 2015, 65, 101–106. [Google Scholar] [CrossRef]
- Shen, S.; Wang, J.; Chen, X.; Liu, T.; Zhuo, Q.; Zhang, S.Q. Evaluation of cellular antioxidant components of honeys using UPLC-MS/MS and HPLC-FLD based on the quantitative composition-activity relationship. Food Chem. 2019, 293, 169–177. [Google Scholar] [CrossRef]
- Najmanová, I.; Vopršalova, M.; Saso, L.; Mladĕnka, P. The pharmacokinetics of flavanones. Crit. Rev. Food Sci. Nutr. 2019, 25, 1–17. [Google Scholar] [CrossRef]
- Jeong, H.; Lee, J.; Kim, S.; Yeo, Y.Y.; So, H.; Wu, H.; Song, Y.S.; Jang, C.Y.; Kim, H.D.; Kim, H.D.; et al. Hepatic metabolism of sakuranetin and its modulating effects on cytochrome P450s and UDP-glucuronosyl transferases. Molecules 2018, 23, 1542. [Google Scholar] [CrossRef]
- Benković, G.; Rimac, H.; Maleš, Ž.; Tomić, S.; Lončar, Z.; Bojić, M. Characetrization of O-demethylations and aromatic hydroxylations mediated by cytochromes P450 in the metabolism of flawonoid agycons. Croat. Chem. Acta 2019, 92. [Google Scholar] [CrossRef]
- Ibrahim, A.R.S.; Galal, A.M.; Ahmed, M.S.; Mossa, G.S. O-demethylation and sulfation of 7-methoxylated flavanones by Cunninghamella elegans. Chem. Pharm. Bull. 2003, 51, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Katsumata, S.; Hamana, K.; Toshima, H.; Hasegawa, M. Identification of sternbin and naringenin as detoxified metabolites from the rice flavanone phytoalexin sakuranetin by Pyricularia oryzae. Chem. Biodivers. 2017, 14, 1600240. [Google Scholar] [CrossRef]
- Lee, E.R.; Kim, J.H.; Choi, H.Y.; Jeon, K.; Cho, S.G. Cytoprotective effect of eriodictyol in UV-irradiated keratinocytes via phosphatase-dependent modulation of both the p38 MAPK and Akt signaling pathways. Cell. Physiol. Biochem. 2011, 27, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Stompor, M.; Uram, Ł.; Podgórski, R. In vitro effect of 8-prenylnaringenin on fibroblasts and glioblastoma cells-cellular accumulation and cytotoxicity. Molecules 2017, 22, 1092. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sarifi-Rad, J. The therapeutic potential of naringenin: A review of clinical trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Lan, Y.; Chen, M.; Wen, C.; Hu, Y.; Liu, Z.; Ye, L. Eriodictyol, not its glucuronide metabolites, attenuates acetaminophen-induced hepatotoxicity. Mol. Pharm. 2017, 14, 2937–2951. [Google Scholar] [CrossRef]
- Le Bail, J.C.; Varnat, F.; Nicolas, J.C.; Habrioux, G. Estrogenic and antiproliferative activities on MCF-7 human breast cancer cells by flavonoids. Cancer Lett. 1998, 130, 209–216. [Google Scholar] [CrossRef]
- Abd El-Hafeez, A.A.; Fujimura, T.; Kamei, R.; Hirakawa, N.; Baba, K.; Ono, K.; Kawamoto, S. A methoxyflavanone derivative from the Asian medicinal herb (Perilla frutescens) induces p53-mediated G2/M cell cycle arrest and apoptosis in A549 human lung adenocarcinoma. Cytotechnology 2018, 70, 899–912. [Google Scholar] [CrossRef]
- Woo, Y.; Shin, S.Y.; Hyun, J.; Lee, S.D.; Lee, Y.H.; Lim, Y. Flavanone inhibit the clonogenicity of HCT116 cololectal cancer cells. Int. J. Mol. Med. 2012, 29, 403–408. [Google Scholar]
- Štuliková, K.; Karabin, M.; Nešpor, J.; Dostálek, P. Therapeutic perspectives of 8-prenylnaringenin, a potent phytoestrogen from hops. Molecules 2018, 23, 660. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.; Maier, C. Phytoestrogens and breast cancer: In vitro anticancer activities of isoflavones, lignans. Coumestans, stilbenes and their analogs and derivatives. Biomed. Pharmacother. 2018, 107, 1648–1666. [Google Scholar] [CrossRef] [PubMed]
- Aufderklamm, S.; Miller, F.; Galasso, A.; Stenzl, A.; Gakis, G. Chemoprevention of prostate cancer by isoflavonoids. Recent Results Cancer Res. 2014, 202, 101–108. [Google Scholar] [PubMed]
- Venturelli, S.; Burkard, M.; Biendl, M.; Lauer, U.M.; Frank, J.; Busch, C. Prenylated chalcones and flavonoids for the prevention and treatment of cancer. Nutrition 2016, 32, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, I.M.C.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef]
- Roleira, F.M.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef]
- Singh, J.P.; Selvendiran, K.; Banu, S.M.; Padmavathi, R.; Sakthisekaran, D. Protective role of apigenin on the status of lipid peroxidation and antioxidant defense against hepatocarcinogenesis in Wistar albino rats. Phytomedicine 2015, 11, 309–314. [Google Scholar] [CrossRef]
- Ugocsai, K.; Varga, A.; Molnar, P.; Antus, S.; Molnar, J. Effects of selected flavonoids and carotenoids on drug accumulation and apoptosis induction in multidrug-resistant colon cancer cells expressing MDR1/LRP. In Vivo 2005, 19, 433–438. [Google Scholar]
- Park, J.H.; Fu, Y.Y.; Chung, I.S.; Hahn, T.R.; Cho, M.H. Cytotoxic property of ultraviolet-induced rice phytoalexins to human colon carcinoma HCT-116 cell. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 237–241. [Google Scholar] [CrossRef]
- Drira, R.; Sakamoto, K. Sakuranetin induces melanogenesis in B16BL6 melanoma cells through inhibition of ERK and PI3K/AKT signaling pathways. Phytother. Res. 2016, 30, 997–1002. [Google Scholar] [CrossRef]
- Anders, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schafer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety aspects of the use of quercetin as a dietary suplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef] [PubMed]
- Habbu, P.; Hiremath, M.; Madagundi, S.; Vankudri, R.; Patil, B.; Savant, C. Phytotherapeutics of polyphenolic-loaded drug delivery systems: A review. Phcog. Rev. 2018, 12, 7–19. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure–activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Davison, E.K.; Brimble, M.A. Natural product derived privileged scaffolds in drug discovery. Curr. Opin. Chem. Biol. 2019, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Danelutte, A.P.; Lago, J.H.G.; Young, M.C.M.; Kato, M. Antifungal flavanones and prenylated hydroquinones from Piper crassinervium Kunth. Phytochemistry 2003, 64, 555–559. [Google Scholar] [CrossRef]
- Pacciaroni, A.V.; Gette, M.A.; Derita, M.; Ariza-Espinar, L.; Gil, R.R.; Zacchino, S.A.; Silva, G.L. Antifungal activity of Heterothalamus alienus metabolites. Phytother. Res. 2008, 22, 524–528. [Google Scholar] [CrossRef]
- Aida, Y.; Tomogami, S.; Kodama, O.; Tsukiboshi, T. Synthesis of 7-methoxyapigeninidin and its fungicidal activity against Gloeocercospora sorghi. Biosci. Biotechnol. Biochem. 1996, 60, 1495–1496. [Google Scholar] [CrossRef]
- Zhang, L.; Kong, Y.; Wu, D.; Zhang, H.; Wu, J.; Chen, J.; Ding, J.; Hu, L.; Jiang, H.; Shen, X. Three flavonoids targeting the beta-hydroxyacyl carrier protein dehydratase from Helicobacetr pylori: Crystal structure characetrization with enzymatic inhibition assay. Protein Sci. 2008, 17, 1971–1978. [Google Scholar] [CrossRef]
- Grecco, S.S.; Reimāo, J.Q.; Tempone, A.G.; Sartorelli, P.; Cunha, R.L.O.R.; Romoff, P.; Ferreira, M.J.P.; Fávero, O.A.; Lago, J.H.G. In vitro antileishmanial and antitrypanosomal activities of flavanones from Baccharis retusa DS. (Asteraceae). Exp. Parasitol. 2012, 130, 141–145. [Google Scholar] [CrossRef]
- Kwon, D.H.; Ji, J.H.; Yim, S.H.; Kim, B.S.; Choi, H.J. Suppression of influenza B virus replication by sakuranetin and mode of its action. Phytother. Res. 2018, 32, 2475–2479. [Google Scholar] [CrossRef]
- Choi, H.J. In vitro antiviral activity of sakuranetin against human rhinovirus 3. Osong Public Health Res. Perspect. 2017, 8, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt-Mernak, M.I.; Pinheiro, N.M.; Santana, F.P.; Guerreiro, M.P.; Saraiv Romanholo, B.M.; Grecco, S.S. Prophylactic and therapeutic treatment with the flavonone sakuranetin ameliorates LPS-induced acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Mernak, M.; Santana, F.; Pinheiro, N.; Saraiva-Ramanholo, B.; Grecco, S.; Tiberio, I.; Martins, M.; Lago, J.; Prado, C. Lung inflammation was attenuated by sakuranetin treatment in a model of acute lung injury. Eur. Res. J. 2014, 44, 3932. [Google Scholar]
- Kim, K.Y.; Kang, H. Sakuranetin inhibits inflammatory enzyme, cytokine, and costimulatory molecule expression in macrophages through modulation of JNK, p38, and STAT1. Evid. Based Complement. Alternat. Med. 2016, 2016, 9824203. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, C.P.P.; de Toledo, A.C.; Perini, A.; Pinheiro, N.M.; Hiyane, M.I.; Grecco, S.D.S.; de Fátima Lopes Calvo Tibério, I.; Câmara, N.O.S.; de Arruda Martins, M.; Lago, J.H.G.; et al. Sakuranetin reverses vascular peribronchial and lung parenchyma remodeling in a murine model of chronic allergic pulmonary inflammation. Acta Histochem. 2016, 118, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Santana, F.P.R.; da Silva, R.C.; Grecco, S.S.; Pinheiro, A.J.M.C.R.; Caperuto, L.C.; Arantes-Costa, F.M.; Claudio, S.R.; Yoshizaki, K.; Macchione, M.; Ribeiro, D.A.; et al. Inhibition of MAPK and STAT3-SOCS3 by sakuranetin attenuated chronic allergic airway inflammation in mice. Mediat. Inflamm. 2019, 2019, 1356356. [Google Scholar] [CrossRef]
- Taguchi, L.; Pinheiro, N.M.; Choqueta-Toledo, A.; Grecco, S.S.; Lopes, F.D.; Caperuto, L.C.; Martins, L.C.; Tiberio, I.F.; Câmara, N.O.; Lago, J.H.; et al. A flavanone from Baccharis retusa (Asteraceae) prevents elastase-induced emphysema in mice by regulating NF-κB, oxidative stress and metalloproteinases. Respir. Res. 2015, 16, 79. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Okuyama, T.; Ishii, T.; Okumura, T.; Ikeya, Y.; Nishizawa, M. Sakuranetin downregulated inducible nitric oxide synthase expression by affecting interleukin-1 receptor and CCAAT/enhancer-binding protein β. J. Nat. Med. 2019, 73, 353–368. [Google Scholar] [CrossRef]
- Toledo, A.C.; Sakoda, C.P.P.; Perini, A.; Pinheiro, N.M.; Magalhães, R.M.; Grecco, S.; Tibério, I.F.L.C.; Cãmara, N.O.; Martins, M.A.; Lago, J.H.G.; et al. Flavanone treatment reverses airway inflammation and remodeling in an asthma murine model. Br. J. Pharmacol. 2013, 168, 1736–1749. [Google Scholar] [CrossRef]
- Zhang, X.; Hung, T.M.; Phuong, P.T.; Ngoc, T.M.; Min, B.S.; Song, K.S.; Seong, Y.H.; Bae, K. Anti-inflammatory activity of flavonoids from Populus davidiana. Arch. Pharm. Res. 2006, 29, 1102–1108. [Google Scholar] [CrossRef]
- Li, C.; Hu, C.; Wang, R.; Wang, H.; Ma, Q.; Chen, S.; He, Y. Protective effect of sakuranetin in brain cells of dementia model rats. Cell. Mol. Biol. 2019, 65, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Abe, D.; Sekiya, K. Sakuranetin induces adipogenesis of 3T3-L1 cells through enhanced expression of PPARγ2. Biochem. Biophys. Res. Commun. 2008, 372, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Hernández, V.; Recio, M.C.; Máñez, S.; Giner, R.M.; Rios, J.L. Effects of naturally occuring dihydroflavonols from Inula viscosa on inflammation and enzymes involved in the arachidonic acid metabolism. Life Sci. 2007, 81, 480–488. [Google Scholar] [CrossRef]
- Charles, C.; Nachtergael, A.; Ouedraogo, M.; Belayew, A.; Duez, P. Effects of chemopreventive natural products on non-homologous end-joining DNA double-strand break repair. Mut. Res. Genet. Toxicol Environm. Mutag. 2014, 768, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, E.H.; Lee, T.H.; Cho, M.H. The methoxyflavonoid isosakuranetin suppresses UV-B-induced matrix metalloproteinase-1 expression and collagen degradation relevant for skin photoaging. Int. J. Mol. Sci. 2016, 17, 1449. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.N.; Kinghorn, A.D.; Slack, J.P.; McCluskey, T.S.; Odley, A.; Jia, Z. In vitro evaluation of flavonoids from Eriodictyon californicum for antagonist activity against the bitterness receptor hTAS2R31. J. Agric. Food Chem. 2011, 59, 13117–13121. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plant Name | Part of the Plant | Ref. |
|---|---|---|
| Prunus puddum (Rosaceae) | bark | [16] |
| Prunus spp. (Rosaceae) | cherry tree bark | [17] |
| Baccharis retusa (Asteraceae) | dried and powdered twigs | [18] |
| Ribes nigrum L. (Grossulariaceae) | leaves | [19] |
| Iris milesii (Iridaceae) | rhizomes | [21] |
| Eriodictyon californicum (Boraginaceae) | leaves | [22] |
| Hyptis salzmanii (Lamiaceae) | leaves | [23] |
| Teucrium stocksianum (Labiatae) | aerial parts | [24] |
| Viscum album L. (Santalaceae) | tinctures | [25] |
| Bonnetia dinizii (Guttiferae) | wood | [26] |
| Primula sieboldii (Primulaceae) | bud exudate | [27] |
| Dodonaea viscosa (L.) Jacq. (Sapindaceae) | - | [28] |
| Prunus avium L. (Rosaceae) | sweet-cherry | [29] |
| Dodonaea viscosa (Sapindaceae) | aerial parts | [30] |
| Xanthorrhoea hastilis (Xanthorrhoeaceae) | dried resin | [31] |
| Eupatorium havanense (Asteraceae) | whole plant | [32] |
| Artemisia campestris subsp. glutinosa (Asteraceae) | - | [33] |
| Daphne aurantiaca (Thymelaeaceae) | stem bark | [34] |
| Dicerothamnus rhinoceroses’ (Asteraceae) | dried leaves | [35] |
| Honey | Acacia | Jujube | Vitex | Linden | Buckwheat | Fennel | Manuka |
|---|---|---|---|---|---|---|---|
| Sakuranetin [ng/g] | 6.49 ± 2.62 | 7.14 ± 4.55 | 6.56 ± 4.98 | 62.2 ± 63.8 | 17.0 ± 15.0 | 4.09 ± 2.71 | 31.6 ± 25.8 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stompor, M. A Review on Sources and Pharmacological Aspects of Sakuranetin. Nutrients 2020, 12, 513. https://doi.org/10.3390/nu12020513
Stompor M. A Review on Sources and Pharmacological Aspects of Sakuranetin. Nutrients. 2020; 12(2):513. https://doi.org/10.3390/nu12020513
Chicago/Turabian StyleStompor, Monika. 2020. "A Review on Sources and Pharmacological Aspects of Sakuranetin" Nutrients 12, no. 2: 513. https://doi.org/10.3390/nu12020513
APA StyleStompor, M. (2020). A Review on Sources and Pharmacological Aspects of Sakuranetin. Nutrients, 12(2), 513. https://doi.org/10.3390/nu12020513