
Mechanistic Basis for the Role of Phytochemicals in Inflammation-Associated Chronic Diseases


 , ,
, ,
Abstract
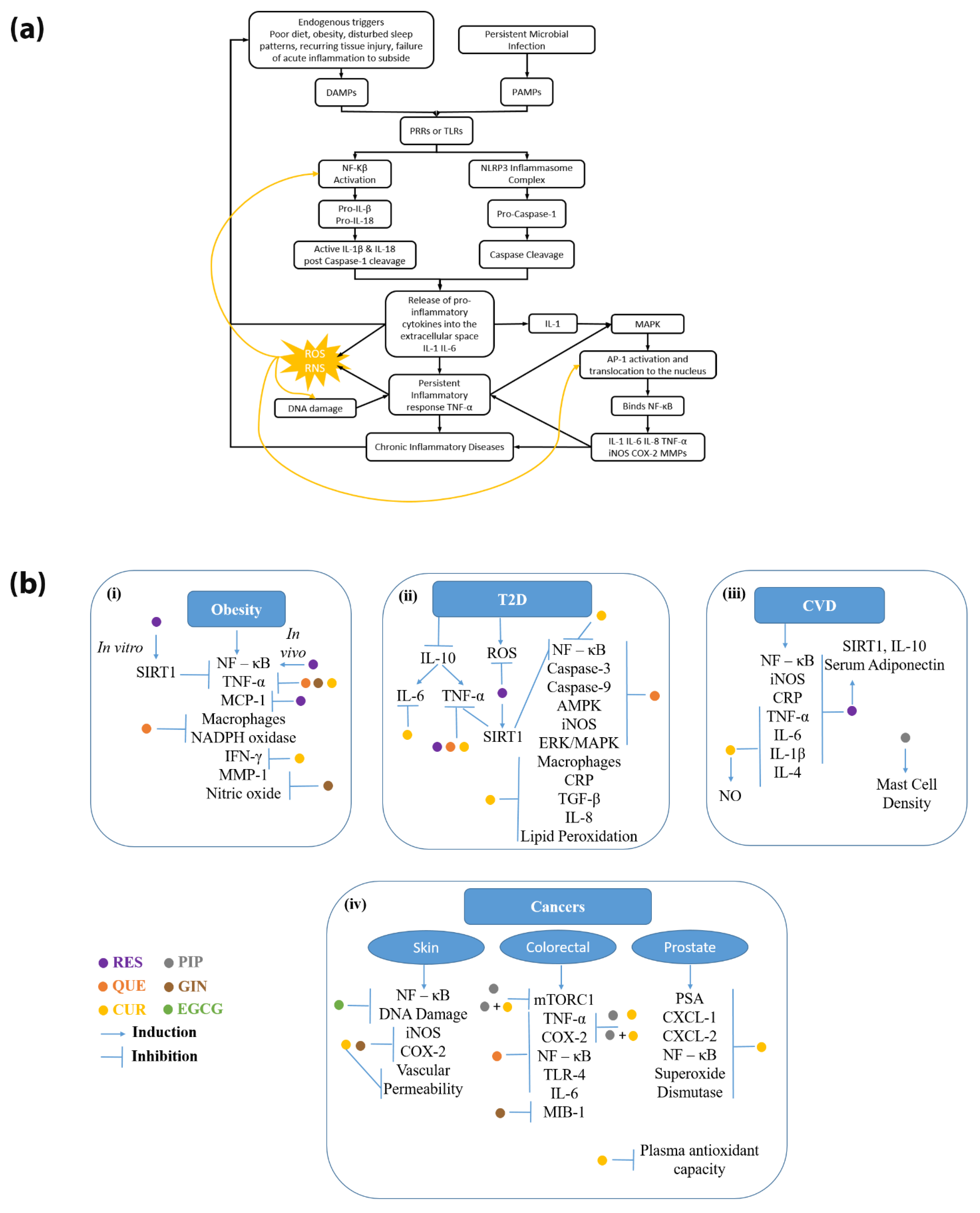
:1. Introduction
2. Current Research Supporting Phytochemical Use in Various Chronic Diseases
2.1. Diseases Related to Metabolic Syndrome
2.1.1. Obesity
2.1.2. Diabetes
2.1.3. Cardiovascular Diseases
2.2. Cancers
2.2.1. Skin Cancers
2.2.2. Colorectal Cancer
2.2.3. Prostate Cancer
3. Drug Interactions Associated with Phytochemical Use
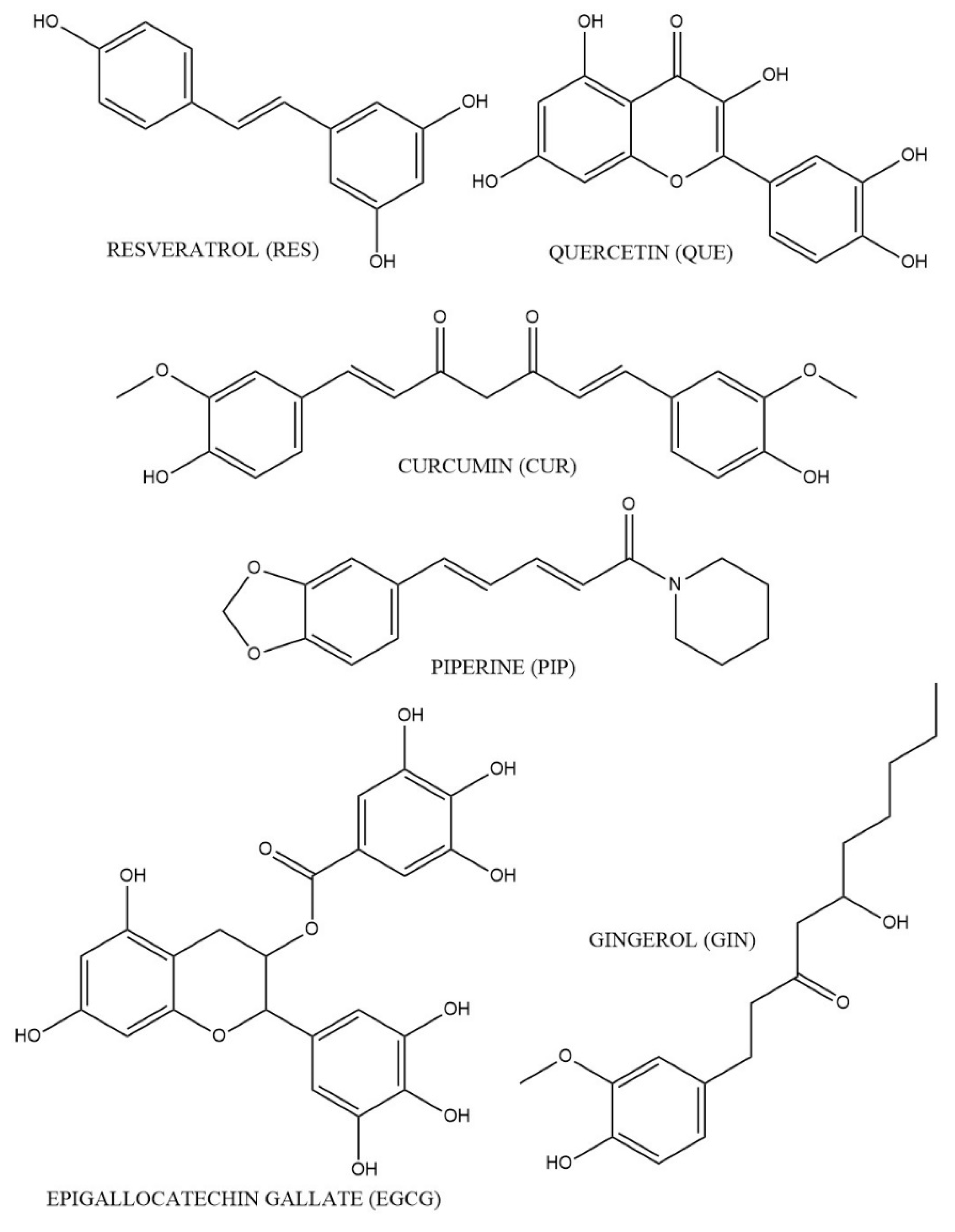
3.1. RES
3.2. QUE
3.3. GIN
3.4. CUR
3.5. PIP
3.6. EGCG

{kind=link}
{kind=link}
| Phytochemical | Model | Route and Dose | F | Tmax | Cmax | t1/2 | Vd | CL | References |
|---|---|---|---|---|---|---|---|---|---|
| curcumin | human | 4000 mg/day | 1.7 | 0.51 µM | [180] | ||||
| 6000 mg/day | 2.0 | 0.64 µM | |||||||
| 8000 mg/day | 1.8 | 1.78 µM | |||||||
| 10 g single oral dose | 3.3 | 2.3 ± 0.26 µg/mL | 6.8 h | [181] | |||||
| 12 g single oral dose | 3.3 | 1.73 ± 0.19 µg/mL | 6.8 h | ||||||
| diabetic rats | PO: 500 mg/kg | 47% | 15 min | 0.06 µg/mL | 32.7 min | Vd: 37.5 L/kg | 0.85 L/kg | [182] | |
| IV: 10 mg/kg | 100% | 5 min | 3.15 µg/mL | 8.64 min | Vd: 10.6 L/kg | 0.83 L/kg | |||
| resveratrol | human | 0.4 g single dose trans-resveratrol (fed) | <1% | 2 h | 42.2 ng/mL | 5.6 h | 3813 L/h | [183,184] | |
| 0.4 g single dose trans-resveratrol (fasting) | 0.5 h | 47.3 ng/mL | 5.9 h | 4249 L/h | |||||
| 0.5 g single dose | 0.83 h | 72.6 ng/mL | 2.8 h | 9198 L | CL/F: 2235 L/h | [183,185] | |||
| CLr/F: 1.177 L/h | |||||||||
| 1 g single dose | 0.76 h | 117.0 ng/mL | 8.87 h | 19,298 L | CL/F: 2593 L/h | ||||
| CLr/F: 0.696 L/h | |||||||||
| 2.5 g single dose | 1.4 h | 268.0 ng/mL | 4.2 h | 22,226 L | CL/F: 3471 L/h | ||||
| CLr/F: 0.656 L/h | |||||||||
| 5 g single dose | 1.5 h | 538.8 ng/mL | 8.5 h | 66,991 L | CL/F: 66,991 L/h | ||||
| CLr/F: 1.443 L/h | |||||||||
| piperine | rat | 170 mg/kg | 18.2 h | [186] | |||||
| 100 mg | Tmax1 6.3 h Tmax2 26.0 h | Cmax1 3.6 µg/mL Cmax2 1.8 µg/mL | 12.85 h | [187] | |||||
| PO: 20 mg/kg | 24% | 2 h | 0.983 µg/mL | 1.2 h | Vss/F: 4.7 L/kg | CL/F: 2.65 L/h/kg | [188] | ||
| IV: 10 mg/kg | 8.0 h | Vss: 7.0 L/kg | CL: 0.64 L/h/kg | ||||||
| quercetin | human | PO: 100 mg (in the form of an onion supplement) | 0.68 ± 0.22 h | 2.3 µg/mL | 11.0 h | Vss: 128 L | 13.3 L/h | [189] | |
| beagle dog | PO: 10 mg/kg | 4% | Tmax1:1.2 h Tmax1: 3.9 h | Cmax1: 0.23 µmol/L Cmax2: 0.23 µmol/L | [190] | ||||
| 6-gingerol | human | PO: 1000 mg | 55 min | 0.4 µg/mL | [165] | ||||
| PO: 1500 mg | 60 min | 1.69 µg/mL | |||||||
| PO: 2000 mg | 66 min | 0.85 µg/mL | 110 min | ||||||
| IV: 1.5 mg/kg | 1.6 h | Vss: 3 L/kg | 3.6 L/h/kg | [191] | |||||
| IV: 3 mg/kg | 1.9 h | Vss: 2.8 L/kg | 3.6 1.4 L/h/kg | ||||||
| IV: 6 mg//kg | 1.7 ± 0.8 h | Vss: 1.9 L/kg | 2.9 0.4 L/h/kg | ||||||
| EGCG | mouse | IV: 21.8 μmol/kg | unconjugated: 13.6 μmol/L | unconjugated: 237.5 min | unconjugated: 1.6 L/kg | unconjugated: 0.57 L/(min·kg) | [174] | ||
| total: 2.7 μmol/L | total: 211.5 min | total: 8.0 L/kg | total: 0.62 ± 0.17 L/(min·kg) | ||||||
| Intragastric: 163.8 μmol/kg | unconjugated: 12.4% | unconjugated: 158 min | unconjugated: 0.04 μmol/L | unconjugated: 465.0 L/kg | unconjugated: 0.45 L/(min·kg) | ||||
| total: 26.5% | total: 90 min | total: 0.28 μmol/L | total: 152.9 L/kg | total: 0.62 L/(min·kg) | |||||
| human | PO: 200 mg | 127.1 min | 73.7 ng/mL | 118.0 min | 2009 L | CL/F: 11.4 L/min | [192] | ||
| PO: 400 mg | 108.7 min | 111.8 ng/mL | 162.3 min | 4774 L | CL/F: 18.0 L/min | ||||
| PO: 600 mg | 180.0 min | 169.1 ng/mL | 183.7 min | 4368 L | CL/F: 12.8 L/min | ||||
| PO: 800 mg | 240.6 min | 438.5 ng/mL | 114.0 min | 1044 L | CL/F: 6.0 L/min |
4. Regulations Associated with Natural Product Use
4.1. Market Availability of Natural Products
4.2. Regulatory Issues with Currently Approved Natural Products
5. Natural Product Informational Databases
6. Discussion and Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, R.; Goyal, A.; Bansal, P.; Jialal, I. Chronic Inflammation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Von Ruesten, A.; Feller, S.; Bergmann, M.M.; Boeing, H. Diet and Risk of Chronic Diseases: Results from the First 8 Years of Follow-up in the EPIC-Potsdam Study. Eur. J. Clin. Nutr. 2013, 67, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.T.; Rimm, E.B.; Spiegelman, D.; Rifai, N.; Tofler, G.H.; Willett, W.C.; Hu, F.B. Association between Dietary Patterns and Plasma Biomarkers of Obesity and Cardiovascular Disease Risk. Am. J. Clin. Nutr. 2001, 73, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J. Fruit and Vegetables and Cancer Risk. Br. J. Cancer 2011, 104, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yang, Y.-Y.; Zhou, M.-W.; Liu, N.; Xing, H.-Y.; Liu, X.-X.; Li, F. Ketogenic Diet Attenuates Oxidative Stress and Inflammation after Spinal Cord Injury by Activating Nrf2 and Suppressing the NF-ΚB Signaling Pathways. Neurosci. Lett. 2018, 683, 13–18. [Google Scholar] [CrossRef]
- Deshmukh, A.A.; Shirvani, S.M.; Likhacheva, A.; Chhatwal, J.; Chiao, E.Y.; Sonawane, K. The Association Between Dietary Quality and Overall and Cancer-Specific Mortality Among Cancer Survivors, NHANES III. JNCI Cancer Spectr. 2018, 2, pky022. [Google Scholar] [CrossRef]
- Sawani, A.; Farhangi, M.; Aluganti, N.C.; Maul, T.M.; Parthasarathy, S.; Smallwood, J.; Wei, J.L. Limiting Dietary Sugar Improves Pediatric Sinonasal Symptoms and Reduces Inflammation. J. Med. Food 2018, 21, 527–534. [Google Scholar] [CrossRef]
- Chovatiya, R.; Medzhitov, R. Stress, Inflammation, and Defense of Homeostasis. Mol. Cell 2014, 54, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An Introduction to Immunology and Immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in Innate Immunity and Inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Gilroy, D.W. Chronic Inflammation: A Failure of Resolution? Int. J. Exp. Pathol. 2007, 88, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Henao-Mejia, J.; Elinav, E.; Thaiss, C.A.; Flavell, R.A. Chapter Three—The Intestinal Microbiota in Chronic Liver Disease. In Advances in Immunology; Alt, F.W., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 117, pp. 73–97. ISBN 0065-2776. [Google Scholar]
- Saïd-Sadier, N.; Ojcius, D.M. Alarmins, Inflammasomes and Immunity. Biomed. J. 2012, 35, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.-K.; Kim, J.K.; Shin, D.-M.; Sasakawa, C. Molecular Mechanisms Regulating NLRP3 Inflammasome Activation. Cell. Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and Regulation of the Inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-KappaB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Crockett-Torabi, E.; Ward, P.A. The Role of Leukocytes in Tissue Injury. Eur. J. Anaesthesiol. 1996, 13, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Boukhenouna, S.; Wilson, M.A.; Bahmed, K.; Kosmider, B. Reactive Oxygen Species in Chronic Obstructive Pulmonary Disease. Oxid. Med. Cell. Longev. 2018, 2018, 5730395. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, D.M.; Asaithamby, A.; Bailey, S.M.; Costes, S.V.; Doetsch, P.W.; Dynan, W.S.; Kronenberg, A.; Rithidech, K.N.; Saha, J.; Snijders, A.M.; et al. Understanding Cancer Development Processes after HZE-Particle Exposure: Roles of ROS, DNA Damage Repair and Inflammation. Radiat. Res. 2015, 183, 1–26. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z. Crosstalk of Reactive Oxygen Species and NF-ΚB Signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-KappaB and AP-1 Connection: Mechanism of NF-KappaB-Dependent Regulation of AP-1 Activity. Mol. Cell. Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [Green Version]
- Rameshwar, P.; Narayanan, R.; Qian, J.; Denny, T.N.; Colon, C.; Gascon, P. NF-Kappa B as a Central Mediator in the Induction of TGF-Beta in Monocytes from Patients with Idiopathic Myelofibrosis: An Inflammatory Response beyond the Realm of Homeostasis. J. Immunol. 2000, 165, 2271–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manicone, A.M.; McGuire, J.K. Matrix Metalloproteinases as Modulators of Inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirantes, C.; Passegué, E.; Pietras, E.M. Pro-Inflammatory Cytokines: Emerging Players Regulating HSC Function in Normal and Diseased Hematopoiesis. Exp. Cell Res. 2014, 329, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Kaptoge, S.; Seshasai, S.R.K.; Gao, P.; Freitag, D.F.; Butterworth, A.S.; Borglykke, A.; Di Angelantonio, E.; Gudnason, V.; Rumley, A.; Lowe, G.D.O.; et al. Inflammatory Cytokines and Risk of Coronary Heart Disease: New Prospective Study and Updated Meta-Analysis. Eur. Heart J. 2014, 35, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Febbraio, M.A. Role of Interleukins in Obesity: Implications for Metabolic Disease. Trends Endocrinol. Metab. TEM 2014, 25, 312–319. [Google Scholar] [CrossRef]
- Patwardhan, B.; Warude, D.; Pushpangadan, P.; Bhatt, N. Ayurveda and Traditional Chinese Medicine: A Comparative Overview. Evid.-Based Complement. Altern. Med. ECAM 2005, 2, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Cote, B.; Carlson, L.J.; Rao, D.A.; Alani, A.W.G. Combinatorial Resveratrol and Quercetin Polymeric Micelles Mitigate Doxorubicin Induced Cardiotoxicity In Vitro and In Vivo. J. Control. Release Off. J. Control. Release Soc. 2015, 213, 128–133. [Google Scholar] [CrossRef]
- Kocaadam, B.; Şanlier, N. Curcumin, an Active Component of Turmeric (Curcuma Longa), and Its Effects on Health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef]
- Meghwal, M.; Goswami, T.K. Piper Nigrum and Piperine: An Update. Phytother. Res. PTR 2013, 27, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Vázquez Cisneros, L.C.; López-Uriarte, P.; López-Espinoza, A.; Navarro Meza, M.; Espinoza-Gallardo, A.C.; Guzmán Aburto, M.B. Effects of green tea and its epigallocatechin (EGCG) content on body weight and fat mass in humans: A systematic review. Nutr. Hosp. 2017, 34, 731–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Yusof, Y.A. Gingerol and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 929, 177–207. [Google Scholar] [CrossRef] [PubMed]
- Metabolic Syndrome. Available online: https://www.nhlbi.nih.gov/health-topics/metabolic-syndrome (accessed on 17 December 2021).
- Pastor, R.F.; Repetto, M.G.; Lairion, F.; Lazarowski, A.; Merelli, A.; Manfredi Carabetti, Z.; Pastor, I.; Pastor, E.; Iermoli, L.V.; Bavasso, C.A.; et al. Supplementation with Resveratrol, Piperine and Alpha-Tocopherol Decreases Chronic Inflammation in a Cluster of Older Adults with Metabolic Syndrome. Nutrients 2020, 12, 3149. [Google Scholar] [CrossRef] [PubMed]
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 14 December 2021).
- Gustafson, B.; Hammarstedt, A.; Andersson, C.X.; Smith, U. Inflamed Adipose Tissue: A Culprit Underlying the Metabolic Syndrome and Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2276–2283. [Google Scholar] [CrossRef] [PubMed]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef]
- Zhu, J.; Yong, W.; Wu, X.; Yu, Y.; Lv, J.; Liu, C.; Mao, X.; Zhu, Y.; Xu, K.; Han, X.; et al. Anti-Inflammatory Effect of Resveratrol on TNF-Alpha-Induced MCP-1 Expression in Adipocytes. Biochem. Biophys. Res. Commun. 2008, 369, 471–477. [Google Scholar] [CrossRef]
- Fischer-Posovszky, P.; Kukulus, V.; Tews, D.; Unterkircher, T.; Debatin, K.-M.; Fulda, S.; Wabitsch, M. Resveratrol Regulates Human Adipocyte Number and Function in a Sirt1-Dependent Manner. Am. J. Clin. Nutr. 2010, 92, 5–15. [Google Scholar] [CrossRef]
- Yao, L.; Wan, J.; Li, H.; Ding, J.; Wang, Y.; Wang, X.; Li, M. Resveratrol Relieves Gestational Diabetes Mellitus in Mice through Activating AMPK. Reprod. Biol. Endocrinol. RBE 2015, 13, 118. [Google Scholar] [CrossRef] [Green Version]
- Konings, E.; Timmers, S.; Boekschoten, M.V.; Goossens, G.H.; Jocken, J.W.; Afman, L.A.; Müller, M.; Schrauwen, P.; Mariman, E.C.; Blaak, E.E. The Effects of 30 Days Resveratrol Supplementation on Adipose Tissue Morphology and Gene Expression Patterns in Obese Men. Int. J. Obes. 2014, 38, 470–473. [Google Scholar] [CrossRef]
- Yoshino, J.; Conte, C.; Fontana, L.; Mittendorfer, B.; Imai, S.; Schechtman, K.B.; Gu, C.; Kunz, I.; Rossi Fanelli, F.; Patterson, B.W.; et al. Resveratrol Supplementation Does Not Improve Metabolic Function in Nonobese Women with Normal Glucose Tolerance. Cell Metab. 2012, 16, 658–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.S.; Cantó, C. The Molecular Targets of Resveratrol. Resveratrol Chall. Transl. Pre-Clin. Find. Improv. Patient Outcomes 2015, 1852, 1114–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.-J.; Lee, Y.-J.; Hwang, J.-H.; Kim, K.-J.; Lee, B.-Y. The Inhibitory Effects of Quercetin on Obesity and Obesity-Induced Inflammation by Regulation of MAPK Signaling. J. Nutr. Biochem. 2015, 26, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Kobori, M.; Takahashi, Y.; Sakurai, M.; Akimoto, Y.; Tsushida, T.; Oike, H.; Ippoushi, K. Quercetin Suppresses Immune Cell Accumulation and Improves Mitochondrial Gene Expression in Adipose Tissue of Diet-Induced Obese Mice. Mol. Nutr. Food Res. 2016, 60, 300–312. [Google Scholar] [CrossRef] [Green Version]
- Egert, S.; Boesch-Saadatmandi, C.; Wolffram, S.; Rimbach, G.; Müller, M.J. Serum Lipid and Blood Pressure Responses to Quercetin Vary in Overweight Patients by Apolipoprotein E Genotype. J. Nutr. 2010, 140, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Kobori, M.; Takahashi, Y.; Takeda, H.; Takahashi, M.; Izumi, Y.; Akimoto, Y.; Sakurai, M.; Oike, H.; Nakagawa, T.; Itoh, M.; et al. Dietary Intake of Curcumin Improves EIF2 Signaling and Reduces Lipid Levels in the White Adipose Tissue of Obese Mice. Sci. Rep. 2018, 8, 9081. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Kim, K.-J.; Kim, B.-H.; Koh, E.-J.; Seo, M.-J.; Lee, B.-Y. 6-Gingerol Suppresses Adipocyte-Derived Mediators of Inflammation In Vitro and in High-Fat Diet-Induced Obese Zebra Fish. Planta Med. 2017, 83, 245–253. [Google Scholar] [CrossRef]
- Ferguson, J.J.A.; Stojanovski, E.; MacDonald-Wicks, L.; Garg, M.L. Curcumin Potentiates Cholesterol-Lowering Effects of Phytosterols in Hypercholesterolaemic Individuals. A Randomised Controlled Trial. Metabolism 2018, 82, 22–35. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Su, Y.-F.; Yang, H.-W.; Lee, Y.-H.; Chou, J.I.; Ueng, K.-C. Lipid-Lowering Effects of Curcumin in Patients with Metabolic Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Phytother. Res. PTR 2014, 28, 1770–1777. [Google Scholar] [CrossRef]
- Yuan, H.; Li, Y.; Ling, F.; Guan, Y.; Zhang, D.; Zhu, Q.; Liu, J.; Wu, Y.; Niu, Y. The Phytochemical Epigallocatechin Gallate Prolongs the Lifespan by Improving Lipid Metabolism, Reducing Inflammation and Oxidative Stress in High-Fat Diet-Fed Obese Rats. Aging Cell 2020, 19, e13199. [Google Scholar] [CrossRef]
- Bogdanski, P.; Suliburska, J.; Szulinska, M.; Stepien, M.; Pupek-Musialik, D.; Jablecka, A. Green Tea Extract Reduces Blood Pressure, Inflammatory Biomarkers, and Oxidative Stress and Improves Parameters Associated with Insulin Resistance in Obese, Hypertensive Patients. Nutr. Res. 2012, 32, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yuan, Y.; Zhou, J.; Hu, R.; Ji, L.; Jiang, G. Piperine Ameliorates Insulin Resistance via Inhibiting Metabolic Inflammation in Monosodium Glutamate-Treated Obese Mice. BMC Endocr. Disord. 2020, 20, 152. [Google Scholar] [CrossRef] [PubMed]
- National Diabetes Statistics Report, 2020; Centers for Disease Control and Prevention, U.S. Dept of Health and Human Services: Atlanta, GA, USA, 2020.
- Donath, M.Y.; Shoelson, S.E. Type 2 Diabetes as an Inflammatory Disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.; Karimzadeh Bardei, L.; Hojati, V.; Ghorbani, A.G.; Nabiuni, M. Anti-Inflammatory Effects of Curcumin on Insulin Resistance Index, Levels of Interleukin-6, C-Reactive Protein, and Liver Histology in Polycystic Ovary Syndrome-Induced Rats. Cell J. 2017, 19, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.-A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques-Vidal, P.; Schmid, R.; Bochud, M.; Bastardot, F.; von Känel, R.; Paccaud, F.; Glaus, J.; Preisig, M.; Waeber, G.; Vollenweider, P. Adipocytokines, Hepatic and Inflammatory Biomarkers and Incidence of Type 2 Diabetes. the CoLaus Study. PLoS ONE 2012, 7, e51768. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose Expression of Tumor Necrosis Factor-Alpha: Direct Role in Obesity-Linked Insulin Resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- Cruz, N.G.; Sousa, L.P.; Sousa, M.O.; Pietrani, N.T.; Fernandes, A.P.; Gomes, K.B. The Linkage between Inflammation and Type 2 Diabetes Mellitus. Diabetes Res. Clin. Pract. 2013, 99, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Côté, C.D.; Rasmussen, B.A.; Duca, F.A.; Zadeh-Tahmasebi, M.; Baur, J.A.; Daljeet, M.; Breen, D.M.; Filippi, B.M.; Lam, T.K.T. Resveratrol Activates Duodenal Sirt1 to Reverse Insulin Resistance in Rats through a Neuronal Network. Nat. Med. 2015, 21, 498–505. [Google Scholar] [CrossRef]
- González-Rodríguez, Á.; Santamaría, B.; Mas-Gutierrez, J.A.; Rada, P.; Fernández-Millán, E.; Pardo, V.; Álvarez, C.; Cuadrado, A.; Ros, M.; Serrano, M.; et al. Resveratrol Treatment Restores Peripheral Insulin Sensitivity in Diabetic Mice in a Sirt1-Independent Manner. Mol. Nutr. Food Res. 2015, 59, 1431–1442. [Google Scholar] [CrossRef]
- Kong, W.; Chen, L.; Zheng, J.; Zhang, H.; Hu, X.; Zeng, T.; Hu, D. Resveratrol Supplementation Restores High-Fat Diet-Induced Insulin Secretion Dysfunction by Increasing Mitochondrial Function in Islet. Exp. Biol. Med. 2015, 240, 220–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R.; et al. Resveratrol Improves Insulin Sensitivity, Reduces Oxidative Stress and Activates the Akt Pathway in Type 2 Diabetic Patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Gomez, Y.; Mattison, J.A.; Pearson, K.J.; Martin-Montalvo, A.; Palacios, H.H.; Sossong, A.M.; Ward, T.M.; Younts, C.M.; Lewis, K.; Allard, J.S.; et al. Resveratrol Improves Adipose Insulin Signaling and Reduces the Inflammatory Response in Adipose Tissue of Rhesus Monkeys on High-Fat, High-Sugar Diet. Cell Metab. 2013, 18, 533–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhao, X.; Ran, L.; Wan, J.; Wang, X.; Qin, Y.; Shu, F.; Gao, Y.; Yuan, L.; Zhang, Q.; et al. Resveratrol Improves Insulin Resistance, Glucose and Lipid Metabolism in Patients with Non-Alcoholic Fatty Liver Disease: A Randomized Controlled Trial. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2015, 47, 226–232. [Google Scholar] [CrossRef]
- Méndez-del Villar, M.; González-Ortiz, M.; Martínez-Abundis, E.; Pérez-Rubio, K.G.; Lizárraga-Valdez, R. Effect of Resveratrol Administration on Metabolic Syndrome, Insulin Sensitivity, and Insulin Secretion. Metab. Syndr. Relat. Disord. 2014, 12, 497–501. [Google Scholar] [CrossRef]
- Zare Javid, A.; Hormoznejad, R.; Yousefimanesh, H.A.; Zakerkish, M.; Haghighi-Zadeh, M.H.; Dehghan, P.; Ravanbakhsh, M. The Impact of Resveratrol Supplementation on Blood Glucose, Insulin, Insulin Resistance, Triglyceride, and Periodontal Markers in Type 2 Diabetic Patients with Chronic Periodontitis. Phytother. Res. PTR 2017, 31, 108–114. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, H.; Wang, M.; Zhang, J. Quercetin Isolated from Toona Sinensis Leaves Attenuates Hyperglycemia and Protects Hepatocytes in High-Carbohydrate/High-Fat Diet and Alloxan Induced Experimental Diabetic Mice. J. Diabetes Res. 2016, 2016, 8492780. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Ding, Y.; Zhang, Z.; Cai, X.; Bao, L.; Li, Y. Quercetin but Not Quercitrin Ameliorates Tumor Necrosis Factor-Alpha-Induced Insulin Resistance in C2C12 Skeletal Muscle Cells. Biol. Pharm. Bull. 2013, 36, 788–795. [Google Scholar] [CrossRef] [Green Version]
- Soetikno, V.; Sari, F.R.; Veeraveedu, P.T.; Thandavarayan, R.A.; Harima, M.; Sukumaran, V.; Lakshmanan, A.P.; Suzuki, K.; Kawachi, H.; Watanabe, K. Curcumin Ameliorates Macrophage Infiltration by Inhibiting NF-ΚB Activation and Proinflammatory Cytokines in Streptozotocin Induced-Diabetic Nephropathy. Nutr. Metab. 2011, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Na, L.X.; Yan, B.L.; Jiang, S.; Cui, H.L.; Li, Y.; Sun, C.H. Curcuminoids Target Decreasing Serum Adipocyte-Fatty Acid Binding Protein Levels in Their Glucose-Lowering Effect in Patients with Type 2 Diabetes. Biomed. Environ. Sci. BES 2014, 27, 902–906. [Google Scholar] [CrossRef]
- Khajehdehi, P.; Pakfetrat, M.; Javidnia, K.; Azad, F.; Malekmakan, L.; Nasab, M.H.; Dehghanzadeh, G. Oral Supplementation of Turmeric Attenuates Proteinuria, Transforming Growth Factor-β and Interleukin-8 Levels in Patients with Overt Type 2 Diabetic Nephropathy: A Randomized, Double-Blind and Placebo-Controlled Study. Scand. J. Urol. Nephrol. 2011, 45, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Franco-Robles, E.; Campos-Cervantes, A.; Murillo-Ortiz, B.O.; Segovia, J.; López-Briones, S.; Vergara, P.; Pérez-Vázquez, V.; Solís-Ortiz, M.S.; Ramírez-Emiliano, J. Effects of Curcumin on Brain-Derived Neurotrophic Factor Levels and Oxidative Damage in Obesity and Diabetes. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2014, 39, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Khalili, N.; Sahebi, E.; Namazi, S.; Simental-Mendía, L.; Majeed, M.; Sahebkar, A. Effects of Curcuminoids Plus Piperine on Glycemic, Hepatic and Inflammatory Biomarkers in Patients with Type 2 Diabetes Mellitus: A Randomized Double-Blind Placebo-Controlled Trial. Drug Res. 2018, 68, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; Alnuqaydan, A.M.; Babiker, A.Y.; Almogbel, M.A.; Khan, A.A.; Husain Rahmani, A. 6-Gingerol, a Bioactive Compound of Ginger Attenuates Renal Damage in Streptozotocin-Induced Diabetic Rats by Regulating the Oxidative Stress and Inflammation. Pharmaceutics 2021, 13, 317. [Google Scholar] [CrossRef]
- Makhdoomi Arzati, M.; Mohammadzadeh Honarvar, N.; Saedisomeolia, A.; Anvari, S.; Effatpanah, M.; Makhdoomi Arzati, R.; Yekaninejad, M.S.; Hashemi, R.; Djalali, M. The Effects of Ginger on Fasting Blood Sugar, Hemoglobin A1c, and Lipid Profiles in Patients with Type 2 Diabetes. Int. J. Endocrinol. Metab. 2017, 15, e57927. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.-M.; Cao, S.-Y.; Wei, X.-L.; Gan, R.-Y.; Wang, Y.-F.; Cai, S.-X.; Xu, X.-Y.; Zhang, P.-Z.; Li, H.-B. Effects and Mechanisms of Tea for the Prevention and Management of Diabetes Mellitus and Diabetic Complications: An Updated Review. Antioxidants 2019, 8, 170. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.-Y.; Li, Q.-S.; Lin, X.-M.; Qiao, R.-Y.; Yang, R.; Li, X.-M.; Dong, Z.-B.; Xiang, L.-P.; Zheng, X.-Q.; Lu, J.-L.; et al. Antidiabetic Effects of Tea. Molecules 2017, 22, 849. [Google Scholar] [CrossRef] [Green Version]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Aikawa, E.; Nahrendorf, M.; Figueiredo, J.-L.; Swirski, F.K.; Shtatland, T.; Kohler, R.H.; Jaffer, F.A.; Aikawa, M.; Weissleder, R. Osteogenesis Associates with Inflammation in Early-Stage Atherosclerosis Evaluated by Molecular Imaging in Vivo. Circulation 2007, 116, 2841–2850. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Seya, K.; Daitoku, K.; Motomura, S.; Fukuda, I.; Furukawa, K.-I. Tumor Necrosis Factor-α Accelerates the Calcification of Human Aortic Valve Interstitial Cells Obtained from Patients with Calcific Aortic Valve Stenosis via the BMP2-Dlx5 Pathway. J. Pharmacol. Exp. Ther. 2011, 337, 16–23. [Google Scholar] [CrossRef]
- Monaco, C.; Andreakos, E.; Kiriakidis, S.; Mauri, C.; Bicknell, C.; Foxwell, B.; Cheshire, N.; Paleolog, E.; Feldmann, M. Canonical Pathway of Nuclear Factor Kappa B Activation Selectively Regulates Proinflammatory and Prothrombotic Responses in Human Atherosclerosis. Proc. Natl. Acad. Sci. USA 2004, 101, 5634–5639. [Google Scholar] [CrossRef] [Green Version]
- Brisdelli, F.; D’Andrea, G.; Bozzi, A. Resveratrol: A Natural Polyphenol with Multiple Chemopreventive Properties. Curr. Drug Metab. 2009, 10, 530–546. [Google Scholar] [CrossRef] [PubMed]
- Cong, X.; Li, Y.; Lu, N.; Dai, Y.; Zhang, H.; Zhao, X.; Liu, Y. Resveratrol Attenuates the Inflammatory Reaction Induced by Ischemia/Reperfusion in the Rat Heart. Mol. Med. Rep. 2014, 9, 2528–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xie, C.; Zhuang, J.; Li, H.; Yao, Y.; Shao, C.; Wang, H. Resveratrol Attenuates Inflammation in the Rat Heart Subjected to Ischemia-Reperfusion: Role of the TLR4/NF-ΚB Signaling Pathway. Mol. Med. Rep. 2015, 11, 1120–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robich, M.P.; Osipov, R.M.; Chu, L.M.; Han, Y.; Feng, J.; Nezafat, R.; Clements, R.T.; Manning, W.J.; Sellke, F.W. Resveratrol Modifies Risk Factors for Coronary Artery Disease in Swine with Metabolic Syndrome and Myocardial Ischemia. Eur. J. Pharmacol. 2011, 664, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Palomer, X.; Capdevila-Busquets, E.; Álvarez-Guardia, D.; Barroso, E.; Pallàs, M.; Camins, A.; Davidson, M.M.; Planavila, A.; Villarroya, F.; Vázquez-Carrera, M. Resveratrol Induces Nuclear Factor-ΚB Activity in Human Cardiac Cells. Int. J. Cardiol. 2013, 167, 2507–2516. [Google Scholar] [CrossRef]
- Huang, F.-C.; Kuo, H.-C.; Huang, Y.-H.; Yu, H.-R.; Li, S.-C.; Kuo, H.-C. Anti-Inflammatory Effect of Resveratrol in Human Coronary Arterial Endothelial Cells via Induction of Autophagy: Implication for the Treatment of Kawasaki Disease. BMC Pharmacol. Toxicol. 2017, 18, 3. [Google Scholar] [CrossRef] [Green Version]
- Planavila, A.; Iglesias, R.; Giralt, M.; Villarroya, F. Sirt1 Acts in Association with PPARα to Protect the Heart from Hypertrophy, Metabolic Dysregulation, and Inflammation. Cardiovasc. Res. 2011, 90, 276–284. [Google Scholar] [CrossRef] [Green Version]
- An, R.; Zhao, L.; Xu, J.; Xi, C.; Li, H.; Shen, G.; Zhang, W.; Zhang, S.; Sun, L. Resveratrol Alleviates Sepsis-induced Myocardial Injury in Rats by Suppressing Neutrophil Accumulation, the Induction of TNF-α and Myocardial Apoptosis via Activation of Sirt1. Mol. Med. Rep. 2016, 14, 5297–5303. [Google Scholar] [CrossRef] [Green Version]
- Carter, S.; Miard, S.; Roy-Bellavance, C.; Boivin, L.; Li, Z.; Pibarot, P.; Mathieu, P.; Picard, F. Sirt1 Inhibits Resistin Expression in Aortic Stenosis. PLoS ONE 2012, 7, e35110. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Grape Resveratrol Increases Serum Adiponectin and Downregulates Inflammatory Genes in Peripheral Blood Mononuclear Cells: A Triple-Blind, Placebo-Controlled, One-Year Clinical Trial in Patients with Stable Coronary Artery Disease. Cardiovasc. Drugs Ther. 2013, 27, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. One-Year Consumption of a Grape Nutraceutical Containing Resveratrol Improves the Inflammatory and Fibrinolytic Status of Patients in Primary Prevention of Cardiovascular Disease. Am. J. Cardiol. 2012, 110, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.-S.; Sharma, A.R.; Nguyen, L.T.; Chakraborty, C.; Sharma, G.; Lee, S.-S. Application of Bioactive Quercetin in Oncotherapy: From Nutrition to Nanomedicine. Molecules 2016, 21, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaskar, S.; Sudhakaran, P.R.; Helen, A. Quercetin Attenuates Atherosclerotic Inflammation and Adhesion Molecule Expression by Modulating TLR-NF-ΚB Signaling Pathway. Cell. Immunol. 2016, 310, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Li, X.X.; Fang, Y.; Chen, X.; Xue, J. Therapeutic Potential of Quercetin as an Antiatherosclerotic Agent in Atherosclerotic Cardiovascular Disease: A Review. Evid. Based Complement. Alternat. Med. 2020, 2020, 5926381. [Google Scholar] [CrossRef]
- Cai, X.; Fang, Z.; Dou, J.; Yu, A.; Zhai, G. Bioavailability of Quercetin: Problems and Promises. Curr. Med. Chem. 2013, 20, 2572–2582. [Google Scholar] [CrossRef]
- Song, Y.; Ge, W.; Cai, H.; Zhang, H. Curcumin Protects Mice from Coxsackievirus B3-Induced Myocarditis by Inhibiting the Phosphatidylinositol 3 Kinase/Akt/Nuclear Factor-ΚB Pathway. J. Cardiovasc. Pharmacol. Ther. 2013, 18, 560–569. [Google Scholar] [CrossRef]
- Wang, N.-P.; Pang, X.-F.; Zhang, L.-H.; Tootle, S.; Harmouche, S.; Zhao, Z.-Q. Attenuation of Inflammatory Response and Reduction in Infarct Size by Postconditioning Are Associated with Downregulation of Early Growth Response 1 during Reperfusion in Rat Heart. Shock 2014, 41, 346–354. [Google Scholar] [CrossRef]
- Mito, S.; Watanabe, K.; Harima, M.; Thandavarayan, R.A.; Veeraveedu, P.T.; Sukumaran, V.; Suzuki, K.; Kodama, M.; Aizawa, Y. Curcumin Ameliorates Cardiac Inflammation in Rats with Autoimmune Myocarditis. Biol. Pharm. Bull. 2011, 34, 974–979. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Zhong, P.; Liang, D.; Xu, Z.; Skibba, M.; Zeng, C.; Li, X.; Wei, T.; Wu, L.; Liang, G. A Newly Designed Curcumin Analog Y20 Mitigates Cardiac Injury via Anti-Inflammatory and Anti-Oxidant Actions in Obese Rats. PLoS ONE 2015, 10, e0120215. [Google Scholar] [CrossRef] [Green Version]
- Ciftci, O.; Turkmen, N.B.; Taslıdere, A. Curcumin Protects Heart Tissue against Irinotecan-Induced Damage in Terms of Cytokine Level Alterations, Oxidative Stress, and Histological Damage in Rats. Naunyn. Schmiedebergs Arch. Pharmacol. 2018, 391, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Santos-Parker, J.R.; Strahler, T.R.; Bassett, C.J.; Bispham, N.Z.; Chonchol, M.B.; Seals, D.R. Curcumin Supplementation Improves Vascular Endothelial Function in Healthy Middle-Aged and Older Adults by Increasing Nitric Oxide Bioavailability and Reducing Oxidative Stress. Aging 2017, 9, 187–208. [Google Scholar] [CrossRef] [Green Version]
- Akazawa, N.; Choi, Y.; Miyaki, A.; Tanabe, Y.; Sugawara, J.; Ajisaka, R.; Maeda, S. Curcumin Ingestion and Exercise Training Improve Vascular Endothelial Function in Postmenopausal Women. Nutr. Res. 2012, 32, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Diwan, V.; Poudyal, H.; Brown, L. Piperine Attenuates Cardiovascular, Liver and Metabolic Changes in High Carbohydrate, High Fat-Fed Rats. Cell Biochem. Biophys. 2013, 67, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.-G.; Yuan, Y.-P.; Zhang, X.; Xu, S.-C.; Wang, S.-S.; Tang, Q.-Z. Piperine Attenuates Pathological Cardiac Fibrosis Via PPAR-γ/AKT Pathways. EBioMedicine 2017, 18, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taqvi, S.I.H.; Shah, A.J.; Gilani, A.H. Blood Pressure Lowering and Vasomodulator Effects of Piperine. J. Cardiovasc. Pharmacol. 2008, 52, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Tian, M.; Liu, P.; Wang, Z.; Guan, Y.; Liu, Y.; Wang, Y.; Shan, Z. Piperine Effectively Protects Primary Cultured Atrial Myocytes from Oxidative Damage in the Infant Rabbit Model. Mol. Med. Rep. 2014, 10, 2627–2632. [Google Scholar] [CrossRef]
- Stojanović-Radić, Z.; Pejčić, M.; Dimitrijević, M.; Aleksić, A.; Anil Kumar, N.V.; Salehi, B.; Cho, W.C.; Sharifi-Rad, J. Piperine-A Major Principle of Black Pepper: A Review of Its Bioactivity and Studies. Appl. Sci. 2019, 9, 4270. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, A.; Nabavi, S.F.; Sureda, A.; Moghaddam, A.H.; Khanjani, S.; Arcidiaco, P.; Nabavi, S.M.; Daglia, M. Antidepressive-like Effects and Antioxidant Activity of Green Tea and GABA Green Tea in a Mouse Model of Post-Stroke Depression. Mol. Nutr. Food Res. 2016, 60, 566–579. [Google Scholar] [CrossRef]
- Yang, G.-Z.; Wang, Z.-J.; Bai, F.; Qin, X.-J.; Cao, J.; Lv, J.-Y.; Zhang, M.-S. Epigallocatechin-3-Gallate Protects HUVECs from PM2.5-Induced Oxidative Stress Injury by Activating Critical Antioxidant Pathways. Molecules 2015, 20, 6626–6639. [Google Scholar] [CrossRef] [Green Version]
- Oyama, J.-I.; Shiraki, A.; Nishikido, T.; Maeda, T.; Komoda, H.; Shimizu, T.; Makino, N.; Node, K. EGCG, a Green Tea Catechin, Attenuates the Progression of Heart Failure Induced by the Heart/Muscle-Specific Deletion of MnSOD in Mice. J. Cardiol. 2017, 69, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.-Y.; Zhao, C.-N.; Gan, R.-Y.; Xu, X.-Y.; Wei, X.-L.; Corke, H.; Atanasov, A.G.; Li, H.-B. Effects and Mechanisms of Tea and Its Bioactive Compounds for the Prevention and Treatment of Cardiovascular Diseases: An Updated Review. Antioxidants 2019, 8, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, M.N.; van der Hooft, J.J.J.; Crozier, A. Human Studies on the Absorption, Distribution, Metabolism, and Excretion of Tea Polyphenols. Am. J. Clin. Nutr. 2013, 98, 1619S–1630S. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Horng, C.; Tsai, S.; Lee, Y.; Hsu, S.; Tsai, Y.; Tsai, F.; Chiang, J.; Kuo, D.; Yang, J. Relaxant and Vasoprotective Effects of Ginger Extracts on Porcine Coronary Arteries. Int. J. Mol. Med. 2018, 41, 2420–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roudsari, N.M.; Lashgari, N.-A.; Momtaz, S.; Roufogalis, B.; Abdolghaffari, A.H.; Sahebkar, A. Ginger: A Complementary Approach for Management of Cardiovascular Diseases. BioFactors 2021, 47, 933–951. [Google Scholar] [CrossRef]
- Cancer Statistics 2020. Available online: https://www.cancer.gov/about-cancer/understanding/statistics (accessed on 14 December 2021).
- Okada, F. Inflammation-Related Carcinogenesis: Current Findings in Epidemiological Trends, Causes and Mechanisms. Yonago Acta Med. 2014, 57, 65–72. [Google Scholar]
- Balamurugan, K. HIF-1 at the Crossroads of Hypoxia, Inflammation, and Cancer. Int. J. Cancer 2016, 138, 1058–1066. [Google Scholar] [CrossRef]
- Pan, M.-G.; Xiong, Y.; Chen, F. NFAT Gene Family in Inflammation and Cancer. Curr. Mol. Med. 2013, 13, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Montes de Oca, M.K.; Alkeswani, A.R.; McClees, S.F.; Das, T.; Elmets, C.A.; Afaq, F. Tea Polyphenols for the Prevention of UVB-Induced Skin Cancer. Photodermatol. Photoimmunol. Photomed. 2018, 34, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Park, K.-K.; Chun, K.-S.; Lee, J.-M.; Lee, S.S.; Surh, Y.-J. Inhibitory Effects of [6]-Gingerol, a Major Pungent Principle of Ginger, on Phorbol Ester-Induced Inflammation, Epidermal Ornithine Decarboxylase Activity and Skin Tumor Promotion in ICR Mice. Cancer Lett. 1998, 129, 139–144. [Google Scholar] [CrossRef]
- Maru, G.B.; Gandhi, K.; Ramchandani, A.; Kumar, G. The Role of Inflammation in Skin Cancer. In Inflammation and Cancer; Aggarwal, B., Sung, B., Gupta, S., Eds.; Advances in Experimental Medicine and Biology; Springer: Berlin, Germany, 2014; Volume 816, pp. 437–469. [Google Scholar]
- Surh, Y.J.; Park, K.K.; Chun, K.S.; Lee, L.J.; Lee, E.; Lee, S.S. Anti-Tumor-Promoting Activities of Selected Pungent Phenolic Substances Present in Ginger. J. Environ. Pathol. Toxicol. Oncol. Off. Organ Int. Soc. Environ. Toxicol. Cancer 1999, 18, 131–139. [Google Scholar]
- Wu, H.; Hsieh, M.-C.; Lo, C.-Y.; Liu, C.B.; Sang, S.; Ho, C.-T.; Pan, M.-H. 6-Shogaol Is More Effective than 6-Gingerol and Curcumin in Inhibiting 12-O-Tetradecanoylphorbol 13-Acetate-Induced Tumor Promotion in Mice. Mol. Nutr. Food Res. 2010, 54, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-P.; Lou, Y.-R.; Xie, J.-G.; Peng, Q.-Y.; Liao, J.; Yang, C.S.; Huang, M.-T.; Conney, A.H. Topical Applications of Caffeine or (-)-Epigallocatechin Gallate (EGCG) Inhibit Carcinogenesis and Selectively Increase Apoptosis in UVB-Induced Skin Tumors in Mice. Proc. Natl. Acad. Sci. USA 2002, 99, 12455–12460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, M.; Ma, W.Y.; Huang, C.; Yang, C.S.; Bowden, G.T.; Miyamoto, K.; Dong, Z. Inhibition of Ultraviolet B-Induced AP-1 Activation by Theaflavins from Black Tea. Mol. Carcinog. 2000, 28, 148–155. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Perez, A.; Mukhtar, H. Green Tea Polyphenol Treatment to Human Skin Prevents Formation of Ultraviolet Light B-Induced Pyrimidine Dimers in DNA. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 3864–3869. [Google Scholar]
- Tuomisto, A.E.; Mäkinen, M.J.; Väyrynen, J.P. Systemic Inflammation in Colorectal Cancer: Underlying Factors, Effects, and Prognostic Significance. World J. Gastroenterol. 2019, 25, 4383–4404. [Google Scholar] [CrossRef]
- Han, M.; Song, Y.; Zhang, X. Quercetin Suppresses the Migration and Invasion in Human Colon Cancer Caco-2 Cells Through Regulating Toll-like Receptor 4/Nuclear Factor-Kappa B Pathway. Pharmacogn. Mag. 2016, 12, S237–S244. [Google Scholar] [CrossRef] [Green Version]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and Colon Cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef]
- Marjaneh, R.M.; Rahmani, F.; Hassanian, S.M.; Rezaei, N.; Hashemzehi, M.; Bahrami, A.; Ariakia, F.; Fiuji, H.; Sahebkar, A.; Avan, A.; et al. Phytosomal Curcumin Inhibits Tumor Growth in Colitis-Associated Colorectal Cancer. J. Cell. Physiol. 2018, 233, 6785–6798. [Google Scholar] [CrossRef]
- Kaur, H.; Kaur, G. A Critical Appraisal of Solubility Enhancement Techniques of Polyphenols. J. Pharm. 2014, 2014, 180845. [Google Scholar] [CrossRef] [Green Version]
- Carroll, R.E.; Benya, R.V.; Turgeon, D.K.; Vareed, S.; Neuman, M.; Rodriguez, L.; Kakarala, M.; Carpenter, P.M.; McLaren, C.; Meyskens, F.L.; et al. Phase IIa Clinical Trial of Curcumin for the Prevention of Colorectal Neoplasia. Cancer Prev. Res. 2011, 4, 354–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citronberg, J.; Bostick, R.; Ahearn, T.; Turgeon, D.K.; Ruffin, M.T.; Djuric, Z.; Sen, A.; Brenner, D.E.; Zick, S.M. Effects of Ginger Supplementation on Cell-Cycle Biomarkers in the Normal-Appearing Colonic Mucosa of Patients at Increased Risk for Colorectal Cancer: Results from a Pilot, Randomized, and Controlled Trial. Cancer Prev. Res. 2013, 6, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhowmik, A.C.; Oksala, N.K.I.; Auriko, J.P.; Paavonen, T.; Mustonen, H.; Paimela, H. Structural Signaling Regulates Inflammation-Induced Enhanced Restitution and Increased Mib-1 and Bax-Indexes after Superficial Injury in Isolated Guinea Pig Gastric Mucosa. Dig. Dis. Sci. 2004, 49, 1526–1530. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.A.; Paulhill, K.J.; Davidson, L.A.; Lupton, J.R.; Taddeo, S.S.; Hong, M.Y.; Carroll, R.J.; Chapkin, R.S.; Turner, N.D. Quercetin May Suppress Rat Aberrant Crypt Foci Formation by Suppressing Inflammatory Mediators That Influence Proliferation and Apoptosis. J. Nutr. 2009, 139, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-A.; Zhang, S.; Yin, Q.; Zhang, J. Quercetin Induces Human Colon Cancer Cells Apoptosis by Inhibiting the Nuclear Factor-Kappa B Pathway. Pharmacogn. Mag. 2015, 11, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Key Statistics for Prostate Cancer. Available online: https://www.cancer.org/cancer/prostate-cancer/about/key-statistics.html (accessed on 20 December 2021).
- Ide, H.; Tokiwa, S.; Sakamaki, K.; Nishio, K.; Isotani, S.; Muto, S.; Hama, T.; Masuda, H.; Horie, S. Combined Inhibitory Effects of Soy Isoflavones and Curcumin on the Production of Prostate-Specific Antigen. Prostate 2010, 70, 1127–1133. [Google Scholar] [CrossRef]
- Stark, T.; Livas, L.; Kyprianou, N. Inflammation in Prostate Cancer Progression and Therapeutic Targeting. Transl. Androl. Urol. 2015, 4, 455–463. [Google Scholar] [CrossRef]
- Killian, P.H.; Kronski, E.; Michalik, K.M.; Barbieri, O.; Astigiano, S.; Sommerhoff, C.P.; Pfeffer, U.; Nerlich, A.G.; Bachmeier, B.E. Curcumin Inhibits Prostate Cancer Metastasis in Vivo by Targeting the Inflammatory Cytokines CXCL1 and -2. Carcinogenesis 2012, 33, 2507–2519. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Williams, M.; Sharma, H.; Chaudry, A.; Bellamy, P. A Double-Blind, Placebo-Controlled Randomised Trial Evaluating the Effect of a Polyphenol-Rich Whole Food Supplement on PSA Progression in Men with Prostate Cancer—the U.K. NCRN Pomi-T Study. Prostate Cancer Prostatic Dis. 2014, 17, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Hejazi, J.; Rastmanesh, R.; Taleban, F.-A.; Molana, S.-H.; Hejazi, E.; Ehtejab, G.; Hara, N. Effect of Curcumin Supplementation During Radiotherapy on Oxidative Status of Patients with Prostate Cancer: A Double Blinded, Randomized, Placebo-Controlled Study. Nutr. Cancer 2016, 68, 77–85. [Google Scholar] [CrossRef]
- Hines, R.N.; McCarver, D.G. The Ontogeny of Human Drug-Metabolizing Enzymes: Phase I Oxidative Enzymes. J. Pharmacol. Exp. Ther. 2002, 300, 355–360. [Google Scholar] [CrossRef] [PubMed]
- McCarver, D.G.; Hines, R.N. The Ontogeny of Human Drug-Metabolizing Enzymes: Phase II Conjugation Enzymes and Regulatory Mechanisms. J. Pharmacol. Exp. Ther. 2002, 300, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, H.-H.S.; Garland, L.L.; Hsu, C.-H.; Vining, D.R.; Chew, W.M.; Miller, J.A.; Perloff, M.; Crowell, J.A.; Alberts, D.S. Resveratrol Modulates Drug- and Carcinogen-Metabolizing Enzymes in a Healthy Volunteer Study. Cancer Prev. Res. 2010, 3, 1168–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, W.K.; Delucchi, A.B. Resveratrol, a Red Wine Constituent, Is a Mechanism-Based Inactivator of Cytochrome P450 3A4. Life Sci. 2000, 67, 3103–3112. [Google Scholar] [CrossRef]
- Piver, B.; Berthou, F.; Dreano, Y.; Lucas, D. Inhibition of CYP3A, CYP1A and CYP2E1 Activities by Resveratrol and Other Non Volatile Red Wine Components. Toxicol. Lett. 2001, 125, 83–91. [Google Scholar] [CrossRef]
- Teo, Y.L.; Ho, H.K.; Chan, A. Metabolism-Related Pharmacokinetic Drug-Drug Interactions with Tyrosine Kinase Inhibitors: Current Understanding, Challenges and Recommendations. Br. J. Clin. Pharmacol. 2015, 79, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Bedada, S.K.; Yellu, N.R.; Neerati, P. Effect of Resveratrol on the Pharmacokinetics of Fexofenadine in Rats: Involvement of P-Glycoprotein Inhibition. Pharmacol. Rep. PR 2016, 68, 338–343. [Google Scholar] [CrossRef]
- Keon Wook Kang, K.W.K.; Jun-Shik Choi, J.-S.C.; Byung-Chul Choi, B.-C.C. Effect of Resveratrol on the Pharmacokinetics of Oral and Intravenous Nicardipine in Rats: Possible Role of P-Glycoprotein Inhibition by Resveratrol. Pharmazie 2009, 64, 49–52. [Google Scholar] [CrossRef]
- Hong, S.-P.; Choi, D.-H.; Choi, J.-S. Effects of Resveratrol on the Pharmacokinetics of Diltiazem and Its Major Metabolite, Desacetyldiltiazem, in Rats. Cardiovasc. Ther. 2008, 26, 269–275. [Google Scholar] [CrossRef]
- Elbarbry, F.; Ung, A.; Abdelkawy, K. Studying the Inhibitory Effect of Quercetin and Thymoquinone on Human Cytochrome P450 Enzyme Activities. Pharmacogn. Mag. 2018, 13, S895–S899. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, H.; Sun, S.; Wang, Y.; Chai, Y.; Yuan, Y. Effect of Ginkgo Leaf Tablets on the Pharmacokinetics of Amlodipine in Rats. Eur. J. Drug Metab. Pharmacokinet. 2016, 41, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Hong, M.; Lee, M.Y.; Jee, J.-G.; Lee, Y.M.; Bae, J.-S.; Jeong, T.C.; Lee, S. Selective Inhibition of the Cytochrome P450 Isoform by Hyperoside and Its Potent Inhibition of CYP2D6. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 59, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Bedada, S.K.; Neerati, P. The Effect of Quercetin on the Pharmacokinetics of Chlorzoxazone, a CYP2E1 Substrate, in Healthy Subjects. Eur. J. Clin. Pharmacol. 2018, 74, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Bedada, S.K.; Neerati, P. Evaluation of the Effect of Quercetin Treatment on CYP2C9 Enzyme Activity of Diclofenac in Healthy Human Volunteers. Phytother. Res. PTR 2018, 32, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, P.; Yue, Q.; Li, J.; Chu, R.; Zhang, W.; Wang, H. Pungent Ginger Components Modulates Human Cytochrome P450 Enzymes in Vitro. Acta Pharmacol. Sin. 2013, 34, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Mukkavilli, R.; Gundala, S.R.; Yang, C.; Donthamsetty, S.; Cantuaria, G.; Jadhav, G.R.; Vangala, S.; Reid, M.D.; Aneja, R. Modulation of Cytochrome P450 Metabolism and Transport across Intestinal Epithelial Barrier by Ginger Biophenolics. PLoS ONE 2014, 9, e108386. [Google Scholar] [CrossRef]
- Zick, S.M.; Djuric, Z.; Ruffin, M.T.; Litzinger, A.J.; Normolle, D.P.; Alrawi, S.; Feng, M.R.; Brenner, D.E. Pharmacokinetics of 6-Gingerol, 8-Gingerol, 10-Gingerol, and 6-Shogaol and Conjugate Metabolites in Healthy Human Subjects. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2008, 17, 1930–1936. [Google Scholar] [CrossRef] [Green Version]
- Volak, L.P.; Ghirmai, S.; Cashman, J.R.; Court, M.H. Curcuminoids Inhibit Multiple Human Cytochromes P450, UDP-Glucuronosyltransferase, and Sulfotransferase Enzymes, Whereas Piperine Is a Relatively Selective CYP3A4 Inhibitor. Drug Metab. Dispos. Biol. Fate Chem. 2008, 36, 1594–1605. [Google Scholar] [CrossRef] [Green Version]
- Basu, N.K.; Kole, L.; Kubota, S.; Owens, I.S. Human UDP-Glucuronosyltransferases Show Atypical Metabolism of Mycophenolic Acid and Inhibition by Curcumin. Drug Metab. Dispos. Biol. Fate Chem. 2004, 32, 768–773. [Google Scholar] [CrossRef] [Green Version]
- Vietri, M.; Pietrabissa, A.; Mosca, F.; Spisni, R.; Pacifici, G.M. Curcumin Is a Potent Inhibitor of Phenol Sulfotransferase (SULT1A1) in Human Liver and Extrahepatic Tissues. Xenobiotica Fate Foreign Compd. Biol. Syst. 2003, 33, 357–363. [Google Scholar] [CrossRef]
- Zhang, W.; Tan, T.M.C.; Lim, L.-Y. Impact of Curcumin-Induced Changes in P-Glycoprotein and CYP3A Expression on the Pharmacokinetics of Peroral Celiprolol and Midazolam in Rats. Drug Metab. Dispos. Biol. Fate Chem. 2007, 35, 110–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, G.; Raina, R.K.; Zutshi, U.; Bedi, K.L.; Johri, R.K.; Sharma, S.C. Effect of Piperine on Bioavailability and Pharmacokinetics of Propranolol and Theophylline in Healthy Volunteers. Eur. J. Clin. Pharmacol. 1991, 41, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Velpandian, T.; Jasuja, R.; Bhardwaj, R.K.; Jaiswal, J.; Gupta, S.K. Piperine in Food: Interference in the Pharmacokinetics of Phenytoin. Eur. J. Drug Metab. Pharmacokinet. 2001, 26, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Atal, C.K.; Dubey, R.K.; Singh, J. Biochemical Basis of Enhanced Drug Bioavailability by Piperine: Evidence That Piperine Is a Potent Inhibitor of Drug Metabolism. J. Pharmacol. Exp. Ther. 1985, 232, 258–262. [Google Scholar] [PubMed]
- Bhardwaj, R.K.; Glaeser, H.; Becquemont, L.; Klotz, U.; Gupta, S.K.; Fromm, M.F. Piperine, a Major Constituent of Black Pepper, Inhibits Human P-Glycoprotein and CYP3A4. J. Pharmacol. Exp. Ther. 2002, 302, 645–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, J.D.; Lee, M.-J.; Lu, H.; Meng, X.; Hong, J.J.J.; Seril, D.N.; Sturgill, M.G.; Yang, C.S. Epigallocatechin-3-Gallate Is Absorbed but Extensively Glucuronidated Following Oral Administration to Mice. J. Nutr. 2003, 133, 4172–4177. [Google Scholar] [CrossRef]
- Selvendiran, K.; Banu, S.M.; Sakthisekaran, D. Oral Supplementation of Piperine Leads to Altered Phase II Enzymes and Reduced DNA Damage and DNA-Protein Cross Links in Benzo(a)Pyrene Induced Experimental Lung Carcinogenesis. Mol. Cell. Biochem. 2005, 268, 141–147. [Google Scholar] [CrossRef]
- Chow, H.-H.S.; Hakim, I.A.; Vining, D.R.; Crowell, J.A.; Cordova, C.A.; Chew, W.M.; Xu, M.-J.; Hsu, C.-H.; Ranger-Moore, J.; Alberts, D.S. Effects of Repeated Green Tea Catechin Administration on Human Cytochrome P450 Activity. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2006, 15, 2473–2476. [Google Scholar] [CrossRef] [Green Version]
- Qian, F.; Wei, D.; Zhang, Q.; Yang, S. Modulation of P-Glycoprotein Function and Reversal of Multidrug Resistance by (-)-Epigallocatechin Gallate in Human Cancer Cells. Biomed. Pharmacother. Biomedecine Pharmacother. 2005, 59, 64–69. [Google Scholar] [CrossRef]
- Knop, J.; Misaka, S.; Singer, K.; Hoier, E.; Müller, F.; Glaeser, H.; König, J.; Fromm, M.F. Inhibitory Effects of Green Tea and (-)-Epigallocatechin Gallate on Transport by OATP1B1, OATP1B3, OCT1, OCT2, MATE1, MATE2-K and P-Glycoprotein. PLoS ONE 2015, 10, e0139370. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zhang, Q.; Yang, Y.; Xu, J.; Fan, A.; Yang, C.S.; Li, N.; Lu, Y.; Chen, J.; Zhao, D.; et al. Epigallocatechin-3-Gallate Decreases the Transport and Metabolism of Simvastatin in Rats. Xenobiotica Fate Foreign Compd. Biol. Syst. 2017, 47, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.L.; Hsu, C.H.; Lin, J.K.; Hsu, M.M.; Ho, Y.F.; Shen, T.S.; Ko, J.Y.; Lin, J.T.; Lin, B.R.; Ming-Shiang, W.; et al. Phase I Clinical Trial of Curcumin, a Chemopreventive Agent, in Patients with High-Risk or Pre-Malignant Lesions. Anticancer Res. 2001, 21, 2895–2900. [Google Scholar] [PubMed]
- Vareed, S.K.; Kakarala, M.; Ruffin, M.T.; Crowell, J.A.; Normolle, D.P.; Djuric, Z.; Brenner, D.E. Pharmacokinetics of Curcumin Conjugate Metabolites in Healthy Human Subjects. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2008, 17, 1411–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierres, V.O.; Campos, M.L.; Arcaro, C.A.; Assis, R.P.; Baldan-Cimatti, H.M.; Peccinini, R.G.; Paula-Gomes, S.; Kettelhut, I.C.; Baviera, A.M.; Brunetti, I.L. Curcumin Pharmacokinetic and Pharmacodynamic Evidences in Streptozotocin-Diabetic Rats Support the Antidiabetic Activity to Be via Metabolite(s). Evid.-Based Complement. Altern. Med. ECAM 2015, 2015, 678218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergides, C.; Chirilă, M.; Silvestro, L.; Pitta, D.; Pittas, A. Bioavailability and Safety Study of Resveratrol 500 mg Tablets in Healthy Male and Female Volunteers. Exp. Ther. Med. 2016, 11, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaz-da-Silva, M.; Loureiro, A.I.; Falcao, A.; Nunes, T.; Rocha, J.-F.; Fernandes-Lopes, C.; Soares, E.; Wright, L.; Almeida, L.; Soares-da-Silva, P. Effect of Food on the Pharmacokinetic Profile of Trans-Resveratrol. Int. J. Clin. Pharmacol. Ther. 2008, 46, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Boocock, D.J.; Faust, G.E.S.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J.; et al. Phase I Dose Escalation Pharmacokinetic Study in Healthy Volunteers of Resveratrol, a Potential Cancer Chemopreventive Agent. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, D.; Srinivasan, K. Tissue Distribution & Elimination of Capsaicin, Piperine & Curcumin Following Oral Intake in Rats. Indian J. Med. Res. 2010, 131, 682–691. [Google Scholar] [PubMed]
- Shao, B.; Cui, C.; Ji, H.; Tang, J.; Wang, Z.; Liu, H.; Qin, M.; Li, X.; Wu, L. Enhanced Oral Bioavailability of Piperine by Self-Emulsifying Drug Delivery Systems: In Vitro, in Vivo and in Situ Intestinal Permeability Studies. Drug Deliv. 2015, 22, 740–747. [Google Scholar] [CrossRef] [Green Version]
- Sahu, P.K.; Sharma, A.; Rayees, S.; Kour, G.; Singh, A.; Khullar, M.; Magotra, A.; Paswan, S.K.; Gupta, M.; Ahmad, I.; et al. Pharmacokinetic Study of Piperine in Wistar Rats after Oral and Intravenous Administration. Int. J. Drug Deliv. 2014, 6, 82–87. [Google Scholar]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and Bioavailability of Quercetin Glycosides in Humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Reinboth, M.; Wolffram, S.; Abraham, G.; Ungemach, F.R.; Cermak, R. Oral Bioavailability of Quercetin from Different Quercetin Glycosides in Dogs. Br. J. Nutr. 2010, 104, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.G.; Shin, B.S.; Choi, Y.; Ryu, J.K.; Shin, S.W.; Choo, H.W.; Yoo, S.D. Determination and Pharmacokinetics of [6]-Gingerol in Mouse Plasma by Liquid Chromatography-Tandem Mass Spectrometry. Biomed. Chromatogr. BMC 2012, 26, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.H.; Cai, Y.; Alberts, D.S.; Hakim, I.; Dorr, R.; Shahi, F.; Crowell, J.A.; Yang, C.S.; Hara, Y. Phase I Pharmacokinetic Study of Tea Polyphenols Following Single-Dose Administration of Epigallocatechin Gallate and Polyphenon E. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2001, 10, 53–58. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- CFR—Code of Federal Regulations. Title 21, Chapter I—Food and Drug Administration, Department of Health and Human Services, Subchapter B—Food for Human Consumption, Part 111 Current Good Manufacturing Practice in Manufacturing, Packaging, Labeling, or Holding Operations for Dietary Supplements. 2021; 21 CFR Part 111. Available online: https://www.ecfr.gov/current/title-21/chapter-I/subchapter-B/part-111?toc=1 (accessed on 17 December 2021).
- Complementary and Alternative Medicine Products and Their Regulation by the Food and Drug Administration—Draft Gudance for Industry. 2007; FDA Guidance Document Docket Number FDA-2206-D-0102. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/complementary-and-alternative-medicine-products-and-their-regulation-food-and-drug-administration (accessed on 23 November 2021).
- Agbabiaka, T.B.; Spencer, N.H.; Khanom, S.; Goodman, C. Prevalence of Drug-Herb and Drug-Supplement Interactions in Older Adults: A Cross-Sectional Survey. Br. J. Gen. Pract. J. R. Coll. Gen. Pract. 2018, 68, e711–e717. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Dunkel, M.; Fullbeck, M.; Neumann, S.; Preissner, R. SuperNatural: A Searchable Database of Available Natural Compounds. Nucleic Acids Res. 2006, 34, D678–D683. [Google Scholar] [CrossRef] [Green Version]
- PubChem Compound Summary for CID 445154, Resveratrol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Resveratrol (accessed on 23 November 2021).
- PubChem Compound Summary for CID 5280343, Quercetin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Quercetin (accessed on 23 November 2021).
- PubChem Compound Summary for CID 969516, Curcumin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Curcumin (accessed on 23 November 2021).
- PubChem Compound Summary for CID 638024, Piperine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Piperine (accessed on 23 November 2021).
- PubChem Compound Summary for CID 442793, Gingerol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Gingerol (accessed on 23 November 2021).
- PubChem Compound Summary for CID 65064, Epigallocatechin Gallate. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Epigallocatechin-gallate (accessed on 23 November 2021).
- Pandit, A.P.; Joshi, S.R.; Dalal, P.S.; Patole, V.C. Curcumin as a Permeability Enhancer Enhanced the Antihyperlipidemic Activity of Dietary Green Tea Extract. BMC Complement. Altern. Med. 2019, 19, 129. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, J.; Ankola, D.D.; Beniwal, V.; Singh, D.; Kumar, M.N.V.R. Nanoparticle Encapsulation Improves Oral Bioavailability of Curcumin by at Least 9-Fold When Compared to Curcumin Administered with Piperine as Absorption Enhancer. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2009, 37, 223–230. [Google Scholar] [CrossRef]
- Scannell, J.W.; Blanckley, A.; Boldon, H.; Warrington, B. Diagnosing the Decline in Pharmaceutical R&D Efficiency. Nat. Rev. Drug Discov. 2012, 11, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; Brayden, D.J.; Casettari, L.; Illum, L. Application of Permeation Enhancers in Oral Delivery of Macromolecules: An Update. Pharmaceutics 2019, 11, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
| Phytochemical | MW | Log P | Intrinsic Aqueous Solubility | Oral Bioavailability |
|---|---|---|---|---|
| RES [199] | 228.24 | 3.1 | 100 µg/mL | <<1% |
| QUE [200] | 302.23 | 1.5 | 60 µg/mL | 4% |
| CUR [182,201] | 368.40 | 3.2 | Insoluble in water | 0.47% |
| PIP [188,202] | 285.34 | 3.5 | 40 µg/mL | 24% |
| GIN [165,203] | 294.4 | 2.5 | 80 µg/mL | low |
| EGCG [204,205] | 458.4 | 1.2 | 32.8 µg/mL | 0.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cote, B.; Elbarbry, F.; Bui, F.; Su, J.W.; Seo, K.; Nguyen, A.; Lee, M.; Rao, D.A. Mechanistic Basis for the Role of Phytochemicals in Inflammation-Associated Chronic Diseases. Molecules 2022, 27, 781. https://doi.org/10.3390/molecules27030781
Cote B, Elbarbry F, Bui F, Su JW, Seo K, Nguyen A, Lee M, Rao DA. Mechanistic Basis for the Role of Phytochemicals in Inflammation-Associated Chronic Diseases. Molecules. 2022; 27(3):781. https://doi.org/10.3390/molecules27030781
Chicago/Turabian StyleCote, Brianna, Fawzy Elbarbry, Fiona Bui, Joe W. Su, Karen Seo, Arthur Nguyen, Max Lee, and Deepa A. Rao. 2022. "Mechanistic Basis for the Role of Phytochemicals in Inflammation-Associated Chronic Diseases" Molecules 27, no. 3: 781. https://doi.org/10.3390/molecules27030781
APA StyleCote, B., Elbarbry, F., Bui, F., Su, J. W., Seo, K., Nguyen, A., Lee, M., & Rao, D. A. (2022). Mechanistic Basis for the Role of Phytochemicals in Inflammation-Associated Chronic Diseases. Molecules, 27(3), 781. https://doi.org/10.3390/molecules27030781