Bioactivities of Centaurium erythraea (Gentianaceae) Decoctions: Antioxidant Activity, Enzyme Inhibition and Docking Studies


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
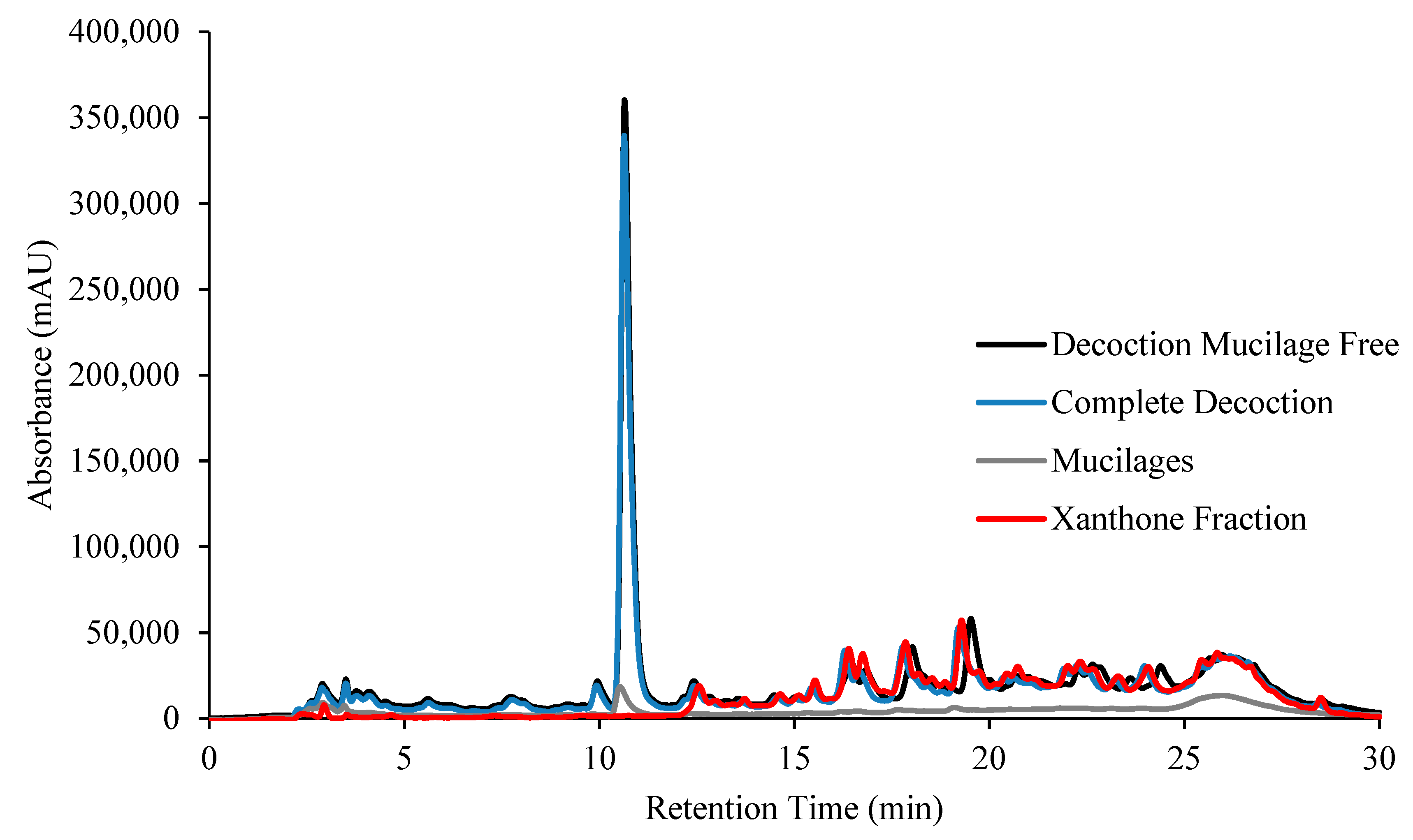
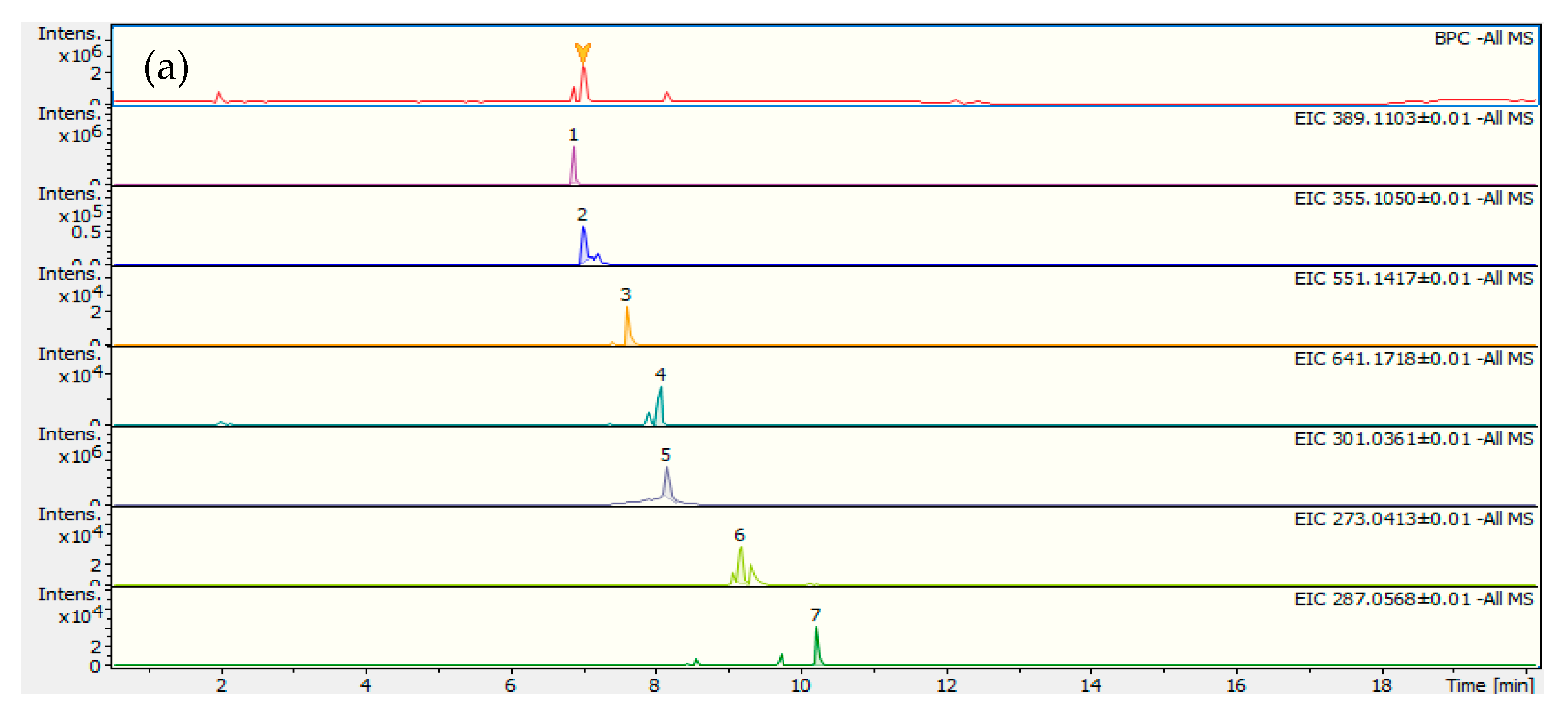
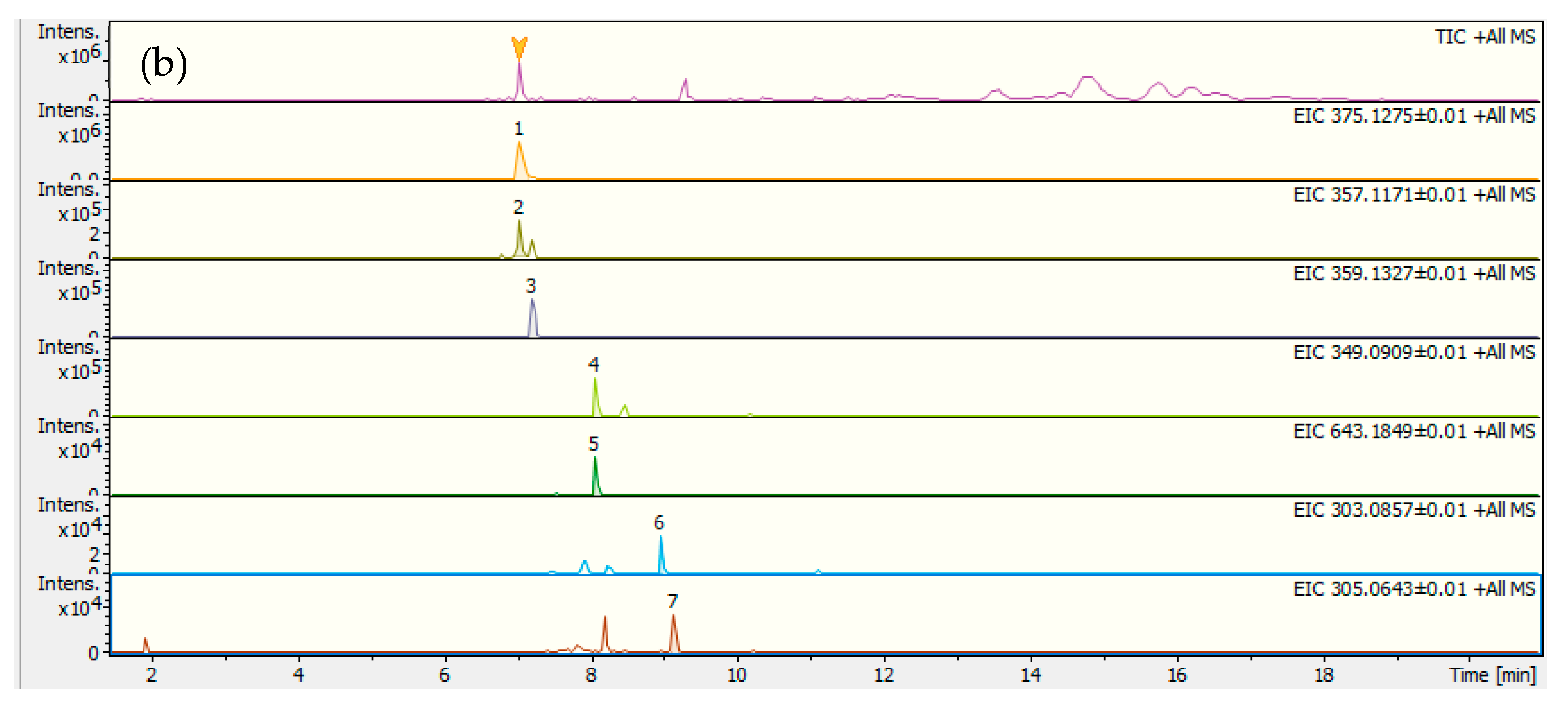
2.1. Decoction Phenolic Compounds
2.2. Total Phenols Quantfication
2.3. Antioxidant Activity
2.3.1. DPPH (2,2-Diphenyl-1-Picrylhydrazyl) Radical Scavenging Activity
2.3.2. Lipoperoxidation Inhibition
2.3.3. Nitric Oxide Radical Scavenging
2.4. Enzyme Inhibition
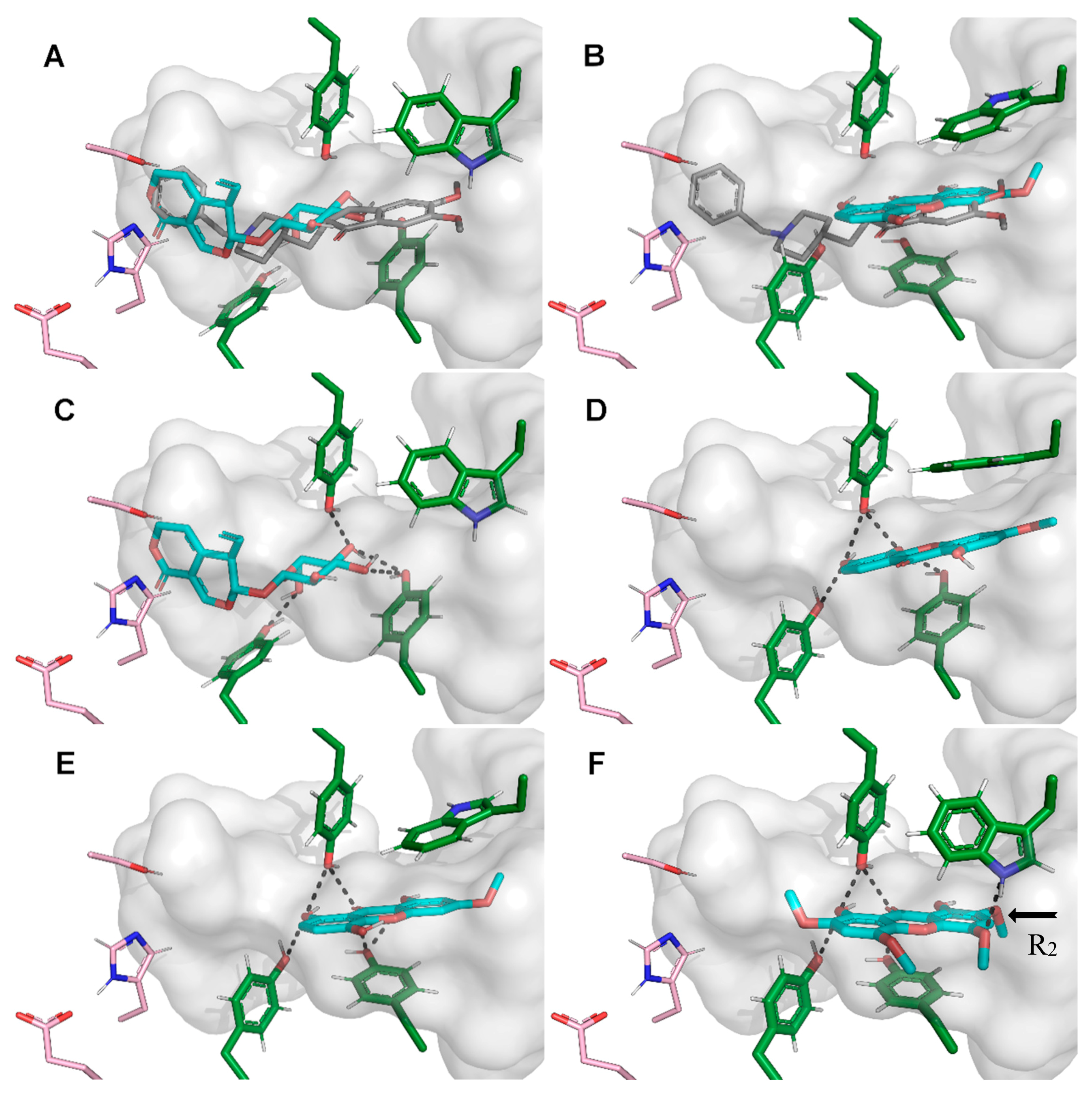
2.4.1. Acetylcholinesterase (AChE) Inhibitory Activities and Docking Studies
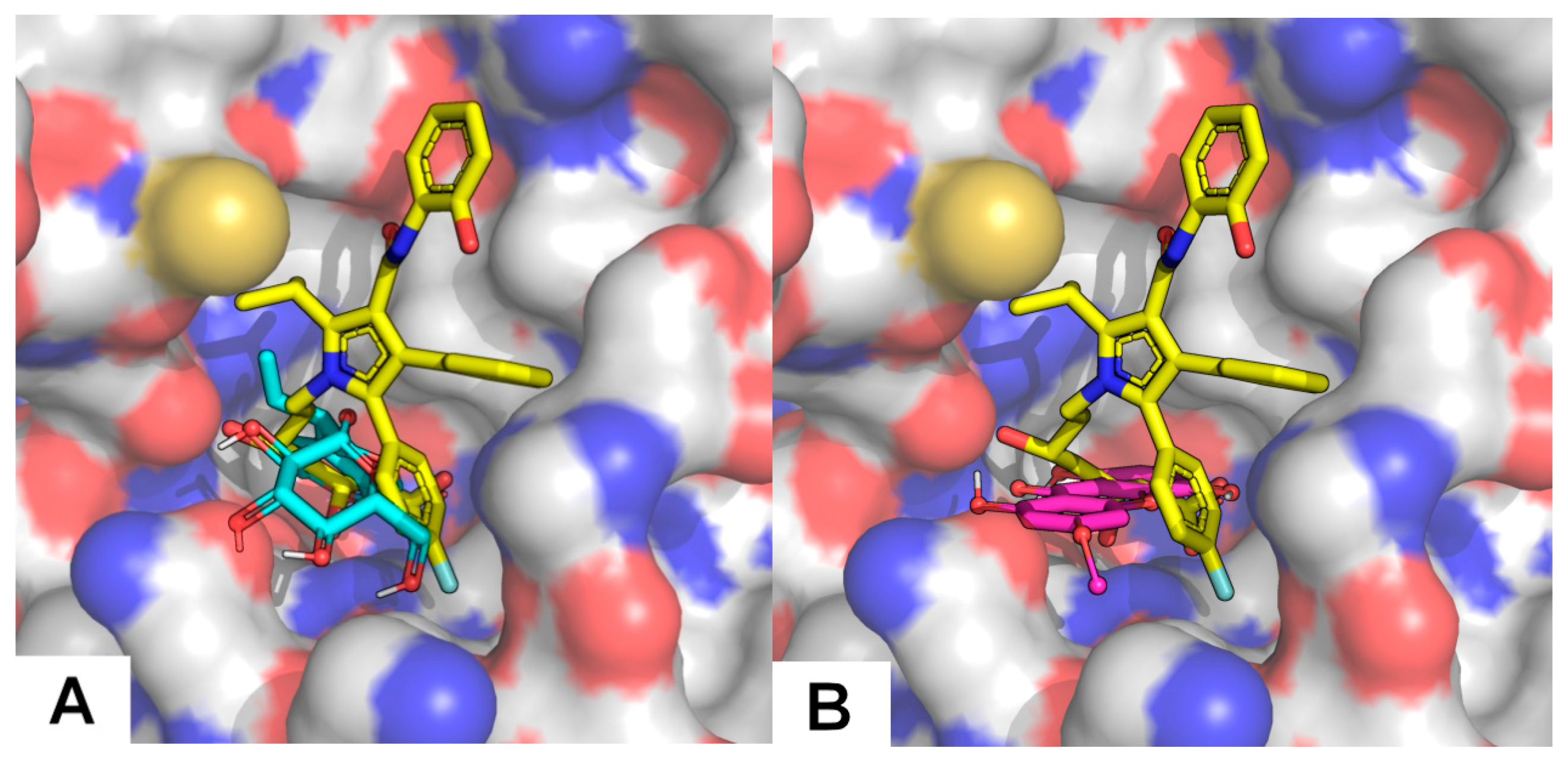
2.4.2. HMGR (3-hydroxy-3-methylglutaryl coenzyme A reductase) Inhibitory Activities and Docking Studies
3. Materials and Methods
3.1. Chemicals
3.2. Plant and Decoction Preparation
3.3. Purification of Phenolic Compounds
3.3.1. Mucilages’ Purification
3.3.2. Xanthones Fraction Collection
3.4. High-Performance Liquid Chromatography with Diode Array Detector (HPLC–DAD) Analysis
3.5. Compound Identification by Liquid Chromatography–High Resolution Tandem Mass Spectrometry (LC–MS/MS)
3.6. Total Phenol Content Quantification
3.7. Antioxidant Activity Determination
3.7.1. DPPH Radical Scavenging Activity
3.7.2. Lipoperoxidation Inhibition
3.7.3. Nitric Oxide Radical Scavenging Activity
3.8. Determination of Enzymatic Activity
3.8.1. Acetylcholinesterase Inhibitory Activity
3.8.2. HMGR Inhibitory Activity
3.9. Computational Methods
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cunha, A.P.; Silva, A.P.; Roque, O. Plantas e Produtos Vegetais em Fitoterapia; Fundação Calouste Gulbenkian: Lisbon, Portugal, 2003; pp. 216–334. ISBN 978-972-31-1435-5. [Google Scholar]
- Tuluce, Y.; Ozkol, H.; Koyuncu, I.; Ine, H. Gastroprotective effect of small centaury (Centaurium erythraea L.) on aspirin-induced gastric damage in rats. Toxicol. Ind. Health 2011, 27, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Benabdelkrim, I.; Rachid, A.; Zaid, A.; Cherrah, Y.; Alaoui, T.; Jean-Louis, G.; El Badaoui, K.K. Effect of aqueous extract from Erythrea Centaurium in Merione shawi. Int. J. Diabetol. Vasc. Dis. Res. 2014, 2, 71–75. [Google Scholar]
- EMA European Medicines Agency. Assessment Report on Centaurium Erythraea Rafn. s.l., Herba; EMA: London, UK, 2015. [Google Scholar]
- Deng, Y.; Wang, L.; Yang, Y.; Sun, W.; Xie, R.; Liu, X.; Wang, Q. In vitro inhibition and induction of human liver cytochrome P450 enzymes by gentiopicroside: Potent effect on CYP2A6. Drug Metab. Pharmacokinet. 2013, 28, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Mirzaee, F.; Hosseini, A.; Jouybari, H.B.; Davoodi, A.; Azadbakht, M. Medicinal, biological and phytochemical properties of Gentiana species. J. Tradit. Complement. Med. 2017, 7, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Lonkar, P.; Dedon, P.C. Reactive species and DNA damage in chronic inflammation: Reconciling chemical mechanisms and biological fates. Int. J. Cancer 2011, 128, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Ma, Q.; Guo, K.; Liu, J.; Li, W.; Wang, F.; Wu, E. Xanthones from mangosteen extracts as natural chemopreventive agents: Potential anticancer drugs. Curr. Mol. Med. 2011, 11, 666–677. [Google Scholar] [CrossRef]
- Roleira, F.M.F.; Tavares-Da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Guan, G.; Lan, S. Implications of antioxidant systems in inflammatory bowel disease. Biomed Res. Int. 2018, 2018, 1–7. [Google Scholar] [CrossRef]
- Kumarasamy, Y.; Nahar, L.; Cox, P.J.; Jaspars, M.; Sarker, S.D. Bioactivity of secoiridoid glycosides from Centaurium erythraea. Phytomedicine 2003, 10, 344–347. [Google Scholar] [CrossRef]
- Berkan, T.; Üstünes, L.; Lermioglu, F.; Özer, A. Antiinflammatory, Analgesic, and Antipyretic Effects of an Aqueous Extract of Erythraea centaurium. Planta Med. 1991, 57, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Tahraoui, A.; Israili, Z.H.; Lyoussi, B. Acute and sub-chronic toxicity of a lyophilised aqueous extract of Centaurium erythraea in rodents. J. Ethnopharmacol. 2010, 132, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Aberham, A.; Pieri, V.; Croom, E.M.; Ellmerer, E.; Stuppner, H. Analysis of iridoids, secoiridoids and xanthones in Centaurium erythraea, Frasera caroliniensis and Gentiana lutea using LC-MS and RP-HPLC. J. Pharm. Biomed Anal. 2011, 54, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Valentão, P.; Fernandes, E.; Carvalho, F.; Andrade, P.B.; Seabra, R.M.; Bastos, M.L. Antioxidant activity of Centaurium erythraea infusion evidenced by its superoxide radical scavenging and xanthine oxidase inhibitory activity. J. Agric. Food Chem. 2001, 49, 3476–3479. [Google Scholar] [CrossRef] [PubMed]
- Molina, M.A.R.; Bah, M.; Rojas, J.I.; Gutiérrez, D.M. Smooth muscle relaxing activity of gentiopicroside isolated from Gentiana spathacea. Planta Med. 2000, 66, 765–767. [Google Scholar]
- Chen, L.; Liu, J.; Zhang, X.; Guo, Y.; Xu, Z.; Cao, W.; Sun, X.; Sun, W.; Zhao, M.G. Down-regulation of NR2B receptors partially contributes to analgesic effects of gentiopicroside in persistent inflammatory pain. Neuropharmacology 2008, 54, 1175–1181. [Google Scholar] [CrossRef]
- Senol, F.S.; Yagci Tuzun, C.; Toker, G.; Orhan, I.E. An in vitro perspective to cholinesterase inhibitory and antioxidant activity of five Gentiana species and Gentianella caucasea. Int. J. Food Sci. Nutr. 2012, 63, 802–812. [Google Scholar] [CrossRef]
- Lian, L.H.; Wu, Y.L.; Wan, Y.; Li, X.; Xie, W.X.; Nan, J.X. Anti-apoptotic activity of gentiopicroside in d-galactosamine/lipopolysaccharide-induced murine fulminant hepatic failure. Chem. Biol. Interact. 2010, 188, 127–133. [Google Scholar] [CrossRef]
- Mustafayeva, K.; Di Giorgio, C.; Elias, R.; Kerimov, Y.; Ollivier, E.; De Méo, M. DNA-damaging, mutagenic, and clastogenic activities of gentiopicroside isolated from Cephalaria kotschyi roots. J. Nat. Prod. 2010, 73, 99–103. [Google Scholar] [CrossRef]
- Ruan, M.; Yu, B.; Xu, L.; Zhang, L.; Long, J.; Shen, X. Attenuation of stress-induced gastrointestinal motility disorder by gentiopicroside, from Gentiana macrophylla Pall. Fitoterapia 2015, 103, 265–276. [Google Scholar] [CrossRef]
- Stefkov, G.; Miova, B.; Dinevska-Kjovkarovska, S.; Stanoeva, J.P.; Stefova, M.; Petrusevska, G.; Kulevanova, S. Chemical characterization of Centaurium erythrea L. and its effects on carbohydrate and lipid metabolism in experimental diabetes. J. Ethnopharmacol. 2014, 152, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Bisht, V.K.; Negi, J.S.; Joshi, G.P.; Singh, P.; Rawat, M.S.M. Naturally occurring xanthones: Chemistry and biology. J. Appl. Chem. 2013, 2013, 1–9. [Google Scholar]
- Hassan, N.K.N.C.; Taher, M.; Susanti, D. Phytochemical constituents and pharmacological properties of Garcinia xanthochymus-A review. Biomed Pharmacother. 2018, 106, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, I.M.P.; Mohammed, A. Acetylcholinesterase mobility and stability at the neuromuscular junction of living mice. Mol. Biol. Cell 2007, 18, 2904–2911. [Google Scholar] [CrossRef] [PubMed]
- England, P.M. Bridging the gaps between synapses, circuits, and behavior. Chem. Biol. 2010, 17, 607–615. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Low, P.; Camilleri, M.; Veil, E.; Burton, D.; Kudva, Y.; Shah, P.; Gehrking, T.; Zinsmeister, A.R. A randomised controlled study of the effect of cholinesterase inhibition on colon function in patients with diabetes mellitus and constipation. Gut 2013, 62, 708–715. [Google Scholar] [CrossRef]
- Jarvie, E.M.; Cellek, S.; Sanger, G.J. Potentiation by cholinesterase inhibitors of cholinergic activity in rat isolated stomach and colon. Pharmacol. Res. 2008, 58, 297–301. [Google Scholar] [CrossRef]
- Nagahama, K.; Matsunaga, Y.; Kawachi, M.; Ito, K.; Tanaka, T.; Hori, Y.; Oka, H.; Takei, M. Acotiamide, a new orally active acetylcholinesterase inhibitor, stimulates gastrointestinal motor activity in conscious dogs. Neurogastroenterol. Motil. 2012, 24, 566–574. [Google Scholar] [CrossRef]
- Zhang, X.; Song, J.; Shi, X.; Miao, S.; Li, Y.; Wen, A. Absorption and metabolism characteristics of rutin in Caco-2 cells. Sci. World J. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- García-Ayllón, M.S.; Small, D.; Avila, J.; Sáez-Valero, J. Revisiting the role of acetylcholinesterase in Alzheimer’s disease: Cross-talk with P-tau and β-amyloid. Front. Mol. Neurosci. 2011, 4, 1–9. [Google Scholar] [CrossRef]
- Singh, N.; Tamariz, J.; Chamorro, G.; Medina-Franco, J.L. Inhibitors of HMG-CoA reductase: Current and future prospects. Mini-Rev. Med. Chem. 2009, 9, 1272–1283. [Google Scholar] [CrossRef] [PubMed]
- Stancu, C.; Sima, A. Statins: Mechanism of action and effects. J. Cell. Mol. Med. 2001, 5, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.R.; Schripsema, J. Chemotaxonomy and pharmacology of Gentianaceae. In Gentianaceae Systematics and Natural History; Cambridge University Press: Cambridge, UK, 2002; Volume 4, pp. 573–631. [Google Scholar]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Foti, M.C. Use and abuse of the DPPH radical. J. Agric. Food Chem. 2015, 63, 8765–8776. [Google Scholar] [CrossRef] [PubMed]
- Kiliç, I.; Yesiloglu, Y.; Bayrak, Y. Investigation on the antioxidant activity of roots and stem of Colchicum turcicum L. Asian J. Chem. 2014, 26, 5–9. [Google Scholar] [CrossRef]
- Attanayake, A.P.; Jayatilaka, K.A.P.W. Evaluation of antioxidant properties of 20 medicinal plant extracts traditionally used in Ayurvedic medicine in Sri Lanka. Indian J. Tradit. Knowl. 2016, 15, 50–56. [Google Scholar]
- Falé, P.L.; Ferreira, C.; Maruzzella, F.; Helena Florêncio, M.; Frazão, F.N.; Serralheiro, M.L.M. Evaluation of cholesterol absorption and biosynthesis by decoctions of Annona cherimola leaves. J. Ethnopharmacol. 2013, 150, 718–723. [Google Scholar] [CrossRef]
- Blanco-Ayala, T.; Lugo-Huitrón, R.; Serrano-López, E.M.; Reyes-Chilpa, R.; Rangel-López, E.; Pineda, B.; Medina-Campos, O.N.; Sánchez-Chapul, L.; Pinzón, E.; Cristina, T.S.; et al. Antioxidant properties of xanthones from Calophyllum brasiliense: Prevention of oxidative damage induced by FeSO4. BMC Complement. Altern. Med. 2013, 13, 1–15. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef]
- Henriques, J.; Ribeiro, M.J.; Falé, P.L.; Pacheco, R.; Ascensão, L.; Florêncio, M.H.; Serralheiro, M.L.M. Valorization of kiwifruit production: Leaves of the pruning branches of Actinidia deliciosa as a promising source of polyphenols. Eur. Food Res. Technol. 2017, 243, 1343–1353. [Google Scholar] [CrossRef]
- Urbain, A.; Marston, A.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. Xanthones from Gentiana campestris as new acetylcholinesterase inhibitors. Planta Med. 2004, 70, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.; Rodrigues, A.M.; Ciriani, M.; Falé, P.L.V.; Teixeira, V.; Madeira, P.; Machuqueiro, M.; Pacheco, R.; Florêncio, M.H.; Ascensão, L.; et al. Antiacetylcholinesterase activity and docking studies with chlorogenic acid, cynarin and arzanol from Helichrysum stoechas (Lamiaceae). Med. Chem. Res. 2017, 26, 2942–2950. [Google Scholar] [CrossRef]
- Valentão, P.; Fernandes, E.; Carvalho, F.; Andrade, P.B.; Seabra, R.M.; Bastos, M.L. Antioxidative properties of cardoon (Cynara cardunculus L.) infusion against superoxide radical, hydroxyl radical, and hypochlorous acid. J. Agric. Food Chem. 2002, 50, 4989–4993. [Google Scholar] [CrossRef] [PubMed]
- Valentão, P.; Areias, F.; Amaral, J.; Andrade, P.; Seabra, R. Tetraoxygenated xanthones from Centaurium erythraea. Nat. Prod. Lett. 2000, 14, 319–323. [Google Scholar] [CrossRef]
- Cheung, J.; Rudolph, M.J.; Burshteyn, F.; Cassidy, M.S.; Gary, E.N.; Love, J.; Franklin, M.C.; Height, J.J. Structures of human acetylcholinesterase in complex with pharmacologically important ligands. J. Med. Chem. 2012, 55, 10282–10286. [Google Scholar] [CrossRef]
- Nag, G.; Das, S.; Das, S.; Mandal, S.; De, B. Antioxidant, anti-acetylcholinesterase and anti-glycosidase properties of three species of Swertia, their xanthones and amarogentin: A comparative study. Pharmacogn. J. 2015, 7, 117–123. [Google Scholar] [CrossRef]
- Bhatt, J.; Vaidya, H.; Khanna, V.; Patel, N.; Goyal, R. In silico docking studies for designing potent anti-diabetic derivatives of swertiamarin with enzyme HMG COA reductase. Mol. Cytogenet. 2014, 7, 97. [Google Scholar] [CrossRef]
- Peffley, D.M.; Gayen, A.K. Plant-derived monoterpenes suppress hamster kidney cell 3-hydroxy-3-methylglutaryl coenzyme A reductase synthesis at the post- transcriptional level. J. Nutr. 2003, 133, 38–44. [Google Scholar] [CrossRef]
- Ressaissi, A.; Attia, N.; Falé, P.L.; Pacheco, R.; Victor, B.L.; Machuqueiro, M.; Serralheiro, M.L.M. Isorhamnetin derivatives and piscidic acid for hypercholesterolemia: Cholesterol permeability, HMG-CoA reductase inhibition, and docking studies. Arch. Pharm. Res. 2017, 40, 1278–1286. [Google Scholar] [CrossRef]
- Sarver, R.W.; Bills, E.; Bolton, G.; Bratton, L.D.; Caspers, N.L.; Dunbar, J.B.; Harris, M.S.; Hutchings, R.H.; Kennedy, R.M.; Larsen, S.D.; et al. Thermodynamic and structure guided design of statin based inhibitors of 3-hydroxy-3-methylglutaryl coenzyme A reductase. J. Med. Chem. 2008, 51, 3804–3813. [Google Scholar] [CrossRef]
- André, R.; Catarro, J.; Freitas, D.; Pacheco, R.; Oliveira, M.C.; Serralheiro, M.L.; Falé, P.L. Action of euptox A from Ageratina adenophora juice on human cell lines: A top-down study using FTIR spectroscopy and protein profiling. Toxicol. Vitr. 2019, 57, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Falé, P.L.; Ferreira, C.; Rodrigues, A.M.; Cleto, P.; Madeira, P.J.A.; Florêncio, M.H.; Frazão, F.N.; Serralheiro, M.L.M. Antioxidant and anti-acetylcholinesterase activity of commercially available medicinal infusions after in vitro gastrointestinal digestion. J. Med. Plants Res. 2013, 7, 1370–1378. [Google Scholar] [Green Version]
- Tokur, B.; Korkmaz, K. The effects of an iron-catalyzed oxidation system on lipids and proteins of dark muscle fish. Food Chem. 2007, 104, 754–760. [Google Scholar] [CrossRef]
- Sakat, S.S.; Juvekar, A.R.; Gambhire, M.N. In-vitro antioxidant and anti-inflammatory activity of methanol extract of Oxalis corniculata linn. Int. J. Pharm. Pharm. Sci. 2010, 2, 146–155. [Google Scholar]
- Mozzicafreddo, M.; Cuccioloni, M.; Eleuteri, A.M.; Angeletti, M. Rapid reverse phase-HPLC assay of HMG-CoA reductase activity. J. Lipid Res. 2010, 51, 2460–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision D.01, Gaussian. Inc.: Wallingford, CT, USA, 2009.
- Morris, G.; Huey, R.; Lindstrom, W.; Sanner, M.; Belew, R.; Goodsell, D.; Olson, A. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, R.; Olson, A.; Goodsell, D. Automated prediction of ligand-binding sites in proteins. Proteins Struct. Funct. Bioinforma. 2008, 70, 1506–1517. [Google Scholar] [CrossRef]
- Mehler, E.L.; Solmajer, T. Electrostatic effects in proteins: Comparison of dielectric and charge models. Protein Eng. 1991, 4, 903–910. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds and water extracts are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) | [M − H]−/[M + H]+ | Formula | Error (∆ ppm) | Main MS2 Fragment ions [m/z, Attribution, (Intensity %)] | Proposed Compound |
|---|---|---|---|---|---|---|
| 1− | 6.9 | 389.1102 | C16H22O11 | −3.1 | 345.1221 [C15H22O9]− (9); 209.0458 [C10H10O5]− (9); 183.0659 [C9H12O4]− (37); 165.0554 [C9H10O3]− (74); 139.0763 [C7H8O3]− (28); 121.0662 [C8H10O1]− (100) | Secologanoside |
| 3− | 7.7 | 551.1417 | C25H28O14 | −2.0 | 507.1517 [C24H28O12]− (66); 393.1187 [C19H22O9]− (80); 389.1081 [C16H22O11]− (51); 345.1195 [C15H22O9]− (44); 323.0793 [C15H16O8]− (51); 301.0352 [C14H22O7]− (40); 281.0676 [C13H14O7]− (68); 161.0236 [C9H6O3]− (100) | Caffeoyl-6′-secologa-noside |
| 4− | 8.1 | 641.1738 | C28H34O17 | −2.3 | 347.0776 [C17H16O8]− (59); 332.0543 [C16H13O8]− (100); 301.0367 [C15H11O7]− (23) | Di-hydroxy-tetrame-toxy -O-pentosyl-hexosylxanthone |
| 5− | 8.2 | 301.0362 | C15H10O7 | −2.7 | -- | Quercetin |
| 6− | 9.2 | 273.0413 | C14H10O6 | −3.1 | 258.0177 [C13H7O6]− (100); 257.0102 [C13H6O6]− (20); 230.0222 [C12H7O5]− (10); 229.0144 [C12H6O5]− (17) | Tri-hydroxy-monome-toxyxanthone |
| 7− | 10.3 | 287.0568 | C15H12O6 | −2.5 | 272.0335 [C14H9O6]− (22); 257.0096 [C13H6O6]− (100); 229.0143 [C12H6O5]− (22) | Di-hydroxy-dimetho-xyxanthone |
| 1+ | 7.1 | 375.1275 | C16H22O10 | 2.9 | 195.0645 [C10H10O4]+ (9); 177.0539 [C10H8O3]+ (100); 167.0696 [C9H10O3]+ (17) | Swertiamarin |
| 2+ | 7.1 | 357.1171 | C16H20O9 | 2.6 | 177.0538 [C10H8O3]+ (100); 163.0378 [C9H6O3]+ (70); 149.0601 [C9H8O2]+ (45) | Gentiopicroside |
| 3+ | 7.2 | 359.1327 | C16H22O9 | 2.8 | 197.0798 [C10H12O4]+ (100); 179.0693 [C10H10O3]+ (50); 177.0536 [C10H8O3]+ (63); 149.0597 [C9H8O2]+ (41); 127.0420 [C6H6O3]+ (84) | Sweroside |
| 4+ | 8.1 | 349.0909 | C17H16O8 | 2.6 | 334.0678 [C16H13O8]+ (7); 319.0443 [C15H10O8]+ (100); 301.0338 [C15H8O7]+ (12); 291.0496 [C14H10O7]+ (9) | Di-hydroxy-tetrame-thoxyxanthone |
| 6+ | 9.0 | 303.0857 | C16H14O6 | 1.9 | 288.0621 [C15H11O6]+ (100); 273.0389 [C14H8O6]+ (19); 245.0438 [C13H8O5]+ (37) | Monohydroxy-trime-toxyxanthone |
| 7+ | 9.1 | 305.0643 | C15H12O7 | 4.2 | 290.0416 [C14H9O7]+ (86) 275.0180 [C13H6O7]+ (100) 247.0238 [C12H6O6]+ (30) | Tri-hydroxy-dimeto-xyxanthone |
| Extracts/Standard | Total Phenols (μg GAE/mg of Extract) | Antioxidant Activity | Enzyme Inhibitory Activity | |||
|---|---|---|---|---|---|---|
| DPPH (50 μg/mL) (%) | TBARS (400 μg/mL) (%) | EC50 NO’s (μg/mL) | AChE (500 μg/mL) (%) | HMGR (10 μg/mL) (%) | ||
| D | 22.37 ± 0.36 | 25.25 ± 0.72 | 32.23 ± 0.93 | 774.9 ± 13.8 | 56.43 ± 0.84 | 47.99 ± 0.47 |
| DMf | 18.82 ± 0.25 | 21.95 ± 0.46 | - | - | 47.09 ± 0.95 | - |
| XF | 16.31 ± 0.25 | 17.63 ± 0.41 | - | - | 59.81 ± 0.7 | 26.86 ± 0.87 |
| Gentio-picroside | - | 1.31±0.53 | - | - | 6.51 ± 0.18 | 57.95 ± 0.51 |
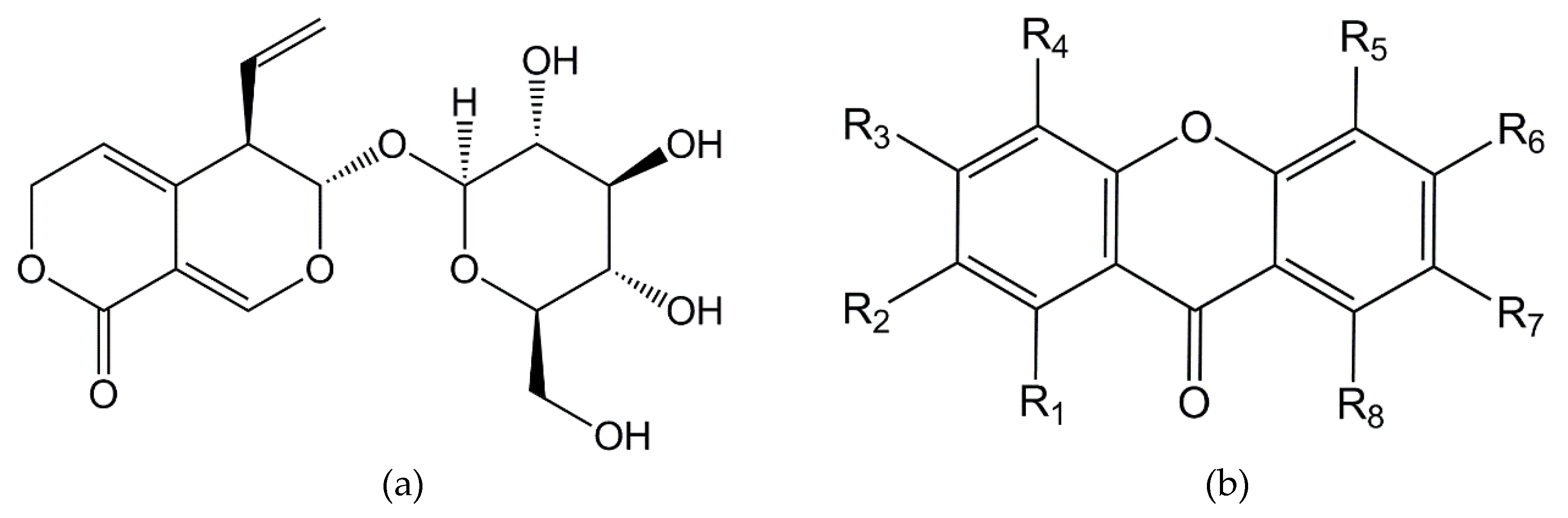
| Final Name | Peak | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | ΔGBIND (kcal/mol) | Ki (μM) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| xan01 * | 4+ | OH | OMe | OMe | OMe | OMe | OH | −6.0 | 36.9 | ||
| xan02 * | 4+ | OH | OMe | OMe | OH | OMe | OMe | −5.6 | 77.6 | ||
| xan03 * | 6+ | OH | OMe | OMe | OMe | −6.9 | 8.9 | ||||
| xan04 * | 6+ | OMe | OH | OMe | OMe | −6.6 | 15.3 | ||||
| xan05 * (decussatin) | 6+ | OH | OMe | OMe | OMe | −5.7 | 65.6 | ||||
| xan06 * | 6+ | OH | OMe | OMe | OMe | −5.9 | 50.9 | ||||
| xan07 * | 7+ | OH | OMe | OH | OMe | OH | −6.1 | 35.1 | |||
| xan08 * | 6− | OH | OMe | OH | OH | −7.5 | 2.9 | ||||
| xan09 * (bellidifolin) | 6− | OH | OMe | OH | OH | −7.5 | 2.9 | ||||
| xan10 * | 6− | OH | OMe | OH | OH | −6.5 | 17.0 | ||||
| xan11 * | 7− | OH | OMe | OMe | OH | −7.1 | 5.9 | ||||
| xan12 | 4+ | OH | OMe | OMe | OMe | OMe | OH | −7.5 | 3.4 | ||
| xan13 | 7+ | OH | OH | OH | OMe | OMe | −6.4 | 19.8 | |||
| xan14 | 6− | OH | OH | OMe | OH | −5.9 | 50.0 | ||||
| xan15 | 6− | OH | OH | OH | OMe | −7.5 | 3.0 | ||||
| xan08.1 | OH | OH | OH | OH | −7.3 | 4.2 | |||||
| xan08.2 | OH | OH | OH | −7.4 | 3.9 | ||||||
| xan08.3 | OH | OMe | OH | OH | OH | −7.1 | 6.0 | ||||
| xan09.1 | OH | OH | OH | OH | −7.4 | 3.5 | |||||
| xan09.2 | OH | OH | OH | −6.8 | 10.1 | ||||||
| xan12.1 | OH | OMe | OMe | OMe | OH | −7.9 | 1.6 | ||||
| xan12.2 | OH | OH | OMe | OMe | OMe | OH | −7.2 | 5.2 | |||
| xan12.3 | OH | OMe | OH | OMe | OMe | OH | −7.4 | 4.0 | |||
| xan12.4 | OH | OMe | OMe | OMe | OH | OH | −7.3 | 4.4 | |||
| xan15.1 | 7− | OMe | OH | OH | OMe | −6.7 | 12.3 |
| Name | ΔGBIND (kcal/mol) | Ki (μM) |
|---|---|---|
| xan01 | −5.7 | 70.6 |
| xan03 | −5.5 | 98.9 |
| xan04 (bellidifolin) | −5.3 | 129.6 |
| xan07 | −5.1 | 184.8 |
| xan13 | −5.6 | 78.1 |
| xan08 | −5.6 | 78.1 |
| xan05 | −5.5 | 94.1 |
| xan12 | −5.7 | 71.8 |
| xan10 | −5.2 | 150.9 |
| xan14 | −5.7 | 70.6 |
| xan15 | −5.3 | 129.6 |
| xan06 | −5.3 | 131.8 |
| xan02 | −5.8 | 54.8 |
| xan11 | −5.4 | 102.3 |
| xan09 | −5.6 | 74.3 |
| gentiopiocroside | −6.4 | 19.9 |
| statin | −11.5 | 3.5 × 10−3 a) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guedes, L.; Reis, P.B.P.S.; Machuqueiro, M.; Ressaissi, A.; Pacheco, R.; Serralheiro, M.L. Bioactivities of Centaurium erythraea (Gentianaceae) Decoctions: Antioxidant Activity, Enzyme Inhibition and Docking Studies. Molecules 2019, 24, 3795. https://doi.org/10.3390/molecules24203795
Guedes L, Reis PBPS, Machuqueiro M, Ressaissi A, Pacheco R, Serralheiro ML. Bioactivities of Centaurium erythraea (Gentianaceae) Decoctions: Antioxidant Activity, Enzyme Inhibition and Docking Studies. Molecules. 2019; 24(20):3795. https://doi.org/10.3390/molecules24203795
Chicago/Turabian StyleGuedes, Laura, Pedro B. P. S. Reis, Miguel Machuqueiro, Asma Ressaissi, Rita Pacheco, and Maria Luísa Serralheiro. 2019. "Bioactivities of Centaurium erythraea (Gentianaceae) Decoctions: Antioxidant Activity, Enzyme Inhibition and Docking Studies" Molecules 24, no. 20: 3795. https://doi.org/10.3390/molecules24203795