
Helicobacter pylori Inhibition, Gastritis Attenuation, and Gut Microbiota Protection in C57BL/6 Mice by Ligilactobacillus salivarius NCUH062003

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Strains and Culture Conditions
2.2. Animal Experimental Design
- Control group: 0.9% NaCl gavage (9 weeks)
- HP_NaCl group: HP (4 weeks) + 0.9% NaCl (4 weeks)
- HP_LP61 group: HP (4 weeks) + L. plantarum CICC 20261 (108 CFU mL−1, 4 weeks)
- HP_LS03 group: HP (4 weeks) + L. salivarius NCUH062003 (108 CFU mL−1, 4 weeks)
- HP_Ant group: HP (4 weeks) + mixed antibiotics (0.025 μg mL−1 omeprazole, 0.125 μg mL−1 amoxicillin and 0.5 μg mL−1 metronidazole, 14 days)
- Ant_LS03 group: HP (4 weeks) + mixed antibiotics (14 days) with L. salivarius NCUH062003 (108 CFU mL−1, 4 weeks).
- Control group: 0.9% NaCl gavage (9 weeks)
- NaCl_HP group: 0.9% NaCl (4 weeks) + HP (4 weeks)
- LP61_HP group: L. plantarum CICC 20261 (108 CFU mL−1, 4 weeks) + HP (4 weeks)
- LS03_HP group: L. salivarius NCUH062003 (108 CFU mL−1, 4 weeks) + HP (4 weeks)
2.3. Organ Coefficients Determination
2.4. H. pylori Colony Count
2.5. Urease Test
2.6. Pathological Examination
2.7. Immunological Factor Analysis
2.8. MPO and SOD Analyses
2.9. Immunohistochemistry (IHC) Analysis
2.10. Immunofluorescence (IF) Assay
2.11. Gut Microbe 16S rRNA Sequencing and Data Analysis
2.12. Fecal Short-Chain Fatty Acid Determination
2.13. Statistical Analysis
3. Results and Discussion
3.1. Body Weight and Organ Coefficient
3.2. Effects of L. salivarius NCUH062003 on H. pylori Activity in Gastric Antrum of HP-Infected Mice
3.3. Effects of L. salivarius NCUH062003 on Gastric Histopathology in HP-Infected Mice
3.4. Alleviation of L. salivarius NCUH062003 on Gastric Mucosal Inflammation in HP-Infected Mice
3.5. Restoration of L. salivarius NCUH062003 on Gastric Mucosa of HP-Infected Mice
3.6. Effect of Therapy Modalities on the Gut Microbiota of HP-Infected Mice
3.6.1. Diversity of Gut Microbiota
3.6.2. Composition of Gut Microbiota
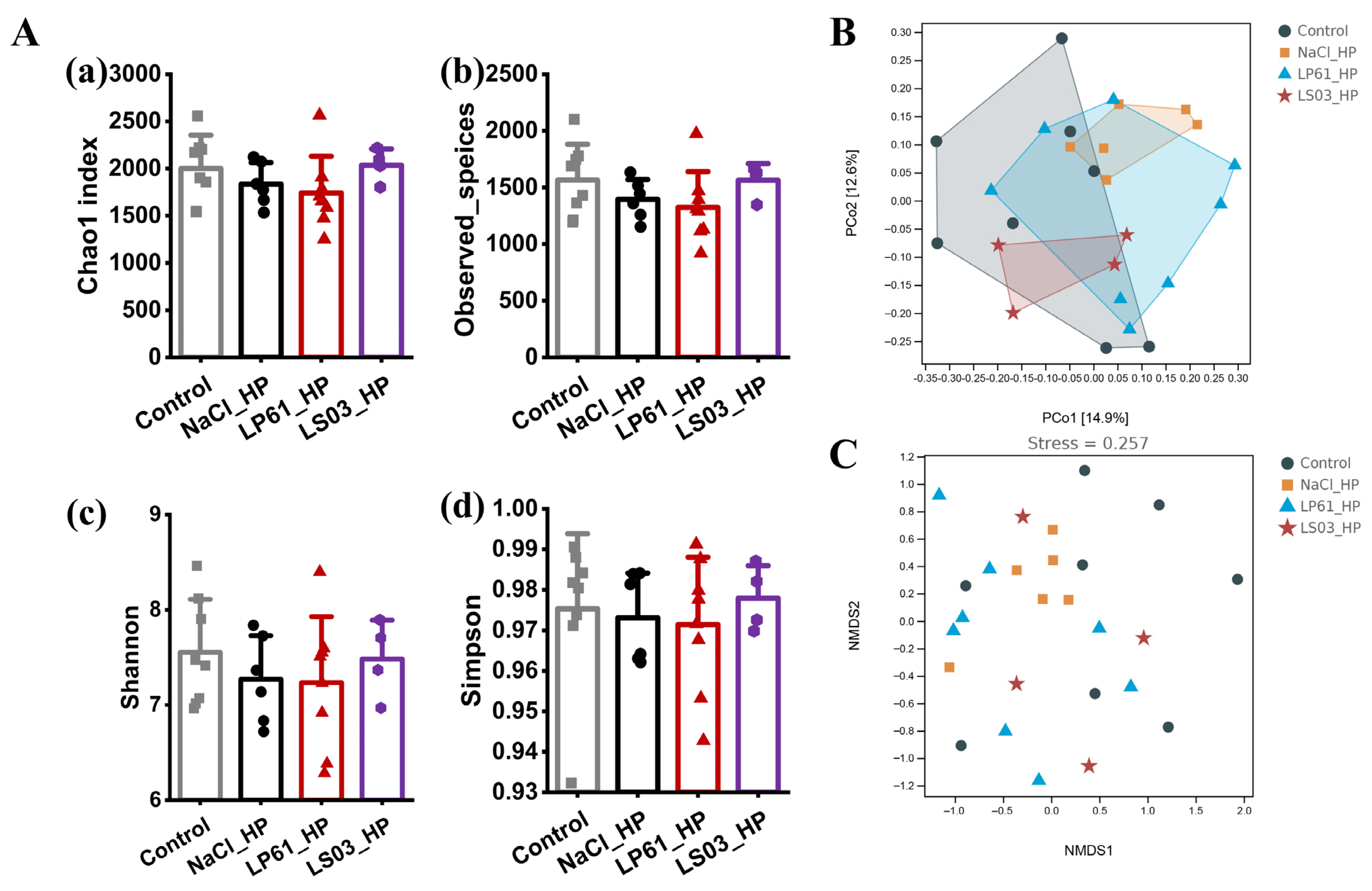
3.7. Effect of Prophylactic Modalities on the Gut Microbiota of HP-Infected Mice
3.7.1. Diversity of Gut Microbiota
3.7.2. Composition of Gut Microbiota
3.8. Effects of L. salivarius NCUH062003 on Intestinal Potential Function
3.9. Effects of L. salivarius NCUH062003 on Intestinal SCFAs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Marshall, B.J.; Warren, J.R. Unidentified Curved Bacilli in the Stomach of Patients with Gastritis and Peptic-Ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- National Toxicology Program. RoC Monograph Series. In Report on Carcinogens Monograph on Helicobacter pylori (Chronic Infection): RoC Monograph 14; National Toxicology Program; Research Triangle Park (NC): Durham, NC, USA, 2018. [Google Scholar]
- Katelaris, P.; Hunt, R.; Bazzoli, F.; Cohen, H.; Fock, K.M.; Gemilyan, M.; Malfertheiner, P.; Mégraud, F.; Piscoya, A.; Quach, D.; et al. Helicobacter pylori World Gastroenterology Organization Global Guideline. J. Clin. Gastroenterol. 2023, 57, 111–126. [Google Scholar] [CrossRef] [PubMed]
- van der Voort, P.H.; van der Hulst, R.W.; Zandstra, D.F.; van der Ende, A.; Kesecioglu, J.; Geraedts, A.A.; Tytgat, G.N. Gut decontamination of critically ill patients reduces Helicobacter pylori acquisition by intensive care nurses. J. Hosp. Infect. 2001, 47, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, Y.; Du, W.; Gong, L.; Chang, H.; Zou, Z. Tumor-associated macrophages: An accomplice in solid tumor progression. J. Biomed. Sci. 2019, 26, 78. [Google Scholar] [CrossRef]
- Sugano, K.; Tack, J.; Kuipers, E.J.; Graham, D.Y.; El-Omar, E.M.; Miura, S.; Haruma, K.; Asaka, M.; Uemura, N.; Malfertheiner, P. Kyoto global consensus report on Helicobacter pylori gastritis. Gut 2015, 64, 1353–1367. [Google Scholar] [CrossRef]
- Fallone, C.A.; Chiba, N.; van Zanten, S.V.; Fischbach, L.; Gisbert, J.P.; Hunt, R.H.; Jones, N.L.; Render, C.; Leontiadis, G.I.; Moayyedi, P.; et al. The Toronto Consensus for the Treatment of Helicobacter pylori Infection in Adults. Gastroenterology 2016, 151, 51–69.e14. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection-the Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef]
- Helicobacter pylori Study Group, Chinese Society of Gastroenterology, Chinese Medical Association. Sixth Chinese National Consensus Report on Management of Helicobacter pylori Infection (Treatment Excluded). Chin. J. Gastroenterol. 2022, 42, 15. [Google Scholar] [CrossRef]
- Ding, S.Z.; Du, Y.Q.; Lu, H.; Wang, W.H.; Cheng, H.; Chen, S.Y.; Chen, M.H.; Chen, W.C.; Chen, Y.; Fang, J.Y.; et al. Chinese Consensus Report on Family-Based Helicobacter pylori Infection Control and Management (2021 Edition). Gut 2022, 71, 238–253. [Google Scholar] [CrossRef]
- Wu, G.; Xu, T.; Zhao, N.; Lam, Y.Y.; Ding, X.; Wei, D.; Fan, J.; Shi, Y.; Li, X.; Li, M.; et al. A core microbiome signature as an indicator of health. Cell 2024, 187, 6550–6565.e11. [Google Scholar] [CrossRef]
- O’Connor, A.; Liou, J.M.; Gisbert, J.P.; O’Morain, C. Review: Treatment of Helicobacter pylori Infection 2019. Helicobacter 2019, 24, e12640. [Google Scholar] [CrossRef] [PubMed]
- Pellicano, R.; Fagoonee, S.; Palestro, G.; Rizzetto, M.; Figura, N.; Ponzetto, A. [The diagnosis of Helicobacter pylori infection: Guidelines from the Maastricht 2-2000 Consensus Report]. Minerva Gastroenterol. Dietol. 2004, 50, 125–133. [Google Scholar] [PubMed]
- Malfertheiner, P.; Bazzoli, F.; Delchier, J.C.; Celiñski, K.; Giguère, M.; Rivière, M.; Mégraud, F. Helicobacter pylori eradication with a capsule containing bismuth subcitrate potassium, metronidazole, and tetracycline given with omeprazole versus clarithromycin-based triple therapy: A randomised, open-label, non-inferiority, phase 3 trial. Lancet 2011, 377, 905–913. [Google Scholar] [CrossRef]
- Maier, L.; Goemans, C.V.; Wirbel, J.; Kuhn, M.; Eberl, C.; Pruteanu, M.; Müller, P.; Garcia-Santamarina, S.; Cacace, E.; Zhang, B.; et al. Unravelling the collateral damage of antibiotics on gut bacteria. Nature 2021, 599, 120–124. [Google Scholar] [CrossRef] [PubMed]
- GHelicobacter pylori Study Group, Chinese Society of Gastroenterology, Chinese Medical Association. Drug-resistant status of Helicobacter pylori in China and the impact of drug resistance on treatment: A national multicentre clinical study. Chin. J. Gastroenterol. 2007, 12, 6. [Google Scholar]
- Graham, D.Y.; Lu, H.; Yamaoka, Y. A report card to grade Helicobacter pylori therapy. Helicobacter 2007, 12, 275–278. [Google Scholar] [CrossRef]
- Engelsberger, V.; Gerhard, M.; Mejías-Luque, R. Effects of Helicobacter pylori infection on intestinal microbiota, immunity and colorectal cancer risk. Front. Cell Infect. Microbiol. 2024, 14, 1339750. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.-m. Holistic integrative medicine declaration. World J. Acupunct. Moxibustion 2024, 3, 271–273. [Google Scholar] [CrossRef]
- Kimura, K. Health Benefits of Probiotics: Probiotics for Helicobacter pylori Infection. Food Sci. Technol. Res. 2004, 10, 1–5. [Google Scholar] [CrossRef]
- Goderska, K.; Agudo Pena, S.; Alarcon, T. Helicobacter pylori treatment: Antibiotics or probiotics. Appl. Microbiol. Biotechnol. 2018, 102, 1–7. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Indian Council of Medical Research Task Force. ICMR-DBT guidelines for evaluation of probiotics in food. Indian. J. Med. Res. 2011, 134, 22–25. [Google Scholar]
- Sakamoto, I.; Igarashi, M.; Kimura, K.; Takagi, A.; Miwa, T.; Koga, Y. Suppressive effect of Lactobacillus gasseri OLL 2716 (LG21) on Helicobacter pylori infection in humans. J. Antimicrob. Chemother. 2001, 47, 709–710. [Google Scholar] [CrossRef] [PubMed]
- Aiba, Y.; Nakano, Y.; Koga, Y.; Takahashi, K.; Komatsu, Y. A highly acid-resistant novel strain of Lactobacillus johnsonii No. 1088 has antibacterial activity, including that against Helicobacter pylori, and inhibits gastrin-mediated acid production in mice. Microbiologyopen 2015, 4, 465–474. [Google Scholar] [CrossRef]
- Li, J.; Peng, F.; Huang, H.; Xu, X.; Guan, Q.; Xie, M.; Xiong, T. Characterization, mechanism and in vivo validation of Helicobacter pylori antagonism by probiotics screened from infants’ feces and oral cavity. Food Funct. 2024, 15, 1170–1190. [Google Scholar] [CrossRef]
- MacArthur Clark, J.A.; Sun, D. Guidelines for the ethical review of laboratory animal welfare People’s Republic of China National Standard GB/T 35892-2018 [Issued 6 February 2018 Effective from 1 September 2018]. Anim. Models Exp. Med. 2020, 3, 103–113. [Google Scholar] [CrossRef]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkilä, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef]
- Ran, X.; Xi, Y.; Lu, Y.; Wang, X.; Lu, Z. Comprehensive survey on hierarchical clustering algorithms and the recent developments. Artif. Intell. Rev. 2022, 56, 8219–8264. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Wei, X.; Koo, I.; Kim, S.; Zhang, X. Compound identification in GC-MS by simultaneously evaluating the mass spectrum and retention index. Analyst 2014, 139, 2507–2514. [Google Scholar] [CrossRef]
- Ueyanagi, Y.; Setoyama, D.; Kawakami, D.; Mushimoto, Y.; Matsumoto, S.; Hotta, T.; Kang, D. Fully Automated Quantitative Measurement of Serum Organic Acids via LC-MS/MS for the Diagnosis of Organic Acidemias: Establishment of an Automation System and a Proof-of-Concept Validation. Diagnostics 2021, 11, 2195. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Y.; Zhang, Z.; Huang, X.; Su, X.; Yang, S.; Xie, Y. Antibiotic-Induced Dysbiosis of the Gut Microbiota Impairs Gene Expression in Gut-Liver Axis of Mice. Genes 2023, 14, 1423. [Google Scholar] [CrossRef]
- Maftei, N.M.; Raileanu, C.R.; Balta, A.A.; Ambrose, L.; Boev, M.; Marin, D.B.; Lisa, E.L. The Potential Impact of Probiotics on Human Health: An Update on Their Health-Promoting Properties. Microorganisms 2024, 12, 234. [Google Scholar] [CrossRef]
- Hsu, M.; Safadi, A.O.; Lui, F. Physiology, Stomach; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Kong, Y.J.; Yi, H.G.; Dai, J.C.; Wei, M.X. Histological changes of gastric mucosa after Helicobacter pylori eradication: A systematic review and meta-analysis. World J. Gastroenterol. 2014, 20, 5903–5911. [Google Scholar] [CrossRef]
- Pennelli, G.; Grillo, F.; Galuppini, F.; Ingravallo, G.; Pilozzi, E.; Rugge, M.; Fiocca, R.; Fassan, M.; Mastracci, L. Gastritis: Update on etiological features and histological practical approach. Pathologica 2020, 112, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Michigami, Y.; Watari, J.; Ito, C.; Nakai, K.; Yamasaki, T.; Kondo, T.; Kono, T.; Tozawa, K.; Tomita, T.; Oshima, T.; et al. Long-term effects of H. pylori eradication on epigenetic alterations related to gastric carcinogenesis. Sci. Rep. 2018, 8, 14369. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Kumai, H.; Nakamichi, N.; Sugiyama, T.; Deguchi, R.; Takagi, A.; Koga, Y. Suppression of Helicobacter pylori-induced interleukin-8 production in vitro and within the gastric mucosa by a live Lactobacillus strain. J. Gastroenterol. Hepatol. 2006, 21, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Zeindler, J.; Angehrn, F.; Droeser, R.; Däster, S.; Piscuoglio, S.; Ng, C.K.Y.; Kilic, E.; Mechera, R.; Meili, S.; Isaak, A.; et al. Infiltration by myeloperoxidase-positive neutrophils is an independent prognostic factor in breast cancer. Breast Cancer Res. Treat. 2019, 177, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.W.; Lai, Y.C. The Role of Helicobacter pylori Neutrophil-Activating Protein in the Pathogenesis of H. pylori and Beyond: From a Virulence Factor to Therapeutic Targets and Therapeutic Agents. Int. J. Mol. Sci. 2022, 24, 91. [Google Scholar] [CrossRef]
- Nie, S.; Yuan, Y. The Role of Gastric Mucosal Immunity in Gastric Diseases. J. Immunol. Res. 2020, 2020, 7927054. [Google Scholar] [CrossRef]
- Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial Dysfunction, Oxidative Stress, and Neuroinflammation: Intertwined Roads to Neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef]
- Gómez-Crisóstomo, N.P.; López-Marure, R.; Zapata, E.; Zazueta, C.; Martínez-Abundis, E. Bax induces cytochrome c release by multiple mechanisms in mitochondria from MCF7 cells. J. Bioenerg. Biomembr. 2013, 45, 441–448. [Google Scholar] [CrossRef]
- Sun, X.; Kaufman, P.D. Ki-67: More than a proliferation marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Kuo, W.T.; Zuo, L.; Odenwald, M.A.; Madha, S.; Singh, G.; Gurniak, C.B.; Abraham, C.; Turner, J.R. The Tight Junction Protein ZO-1 Is Dispensable for Barrier Function but Critical for Effective Mucosal Repair. Gastroenterology 2021, 161, 1924–1939. [Google Scholar] [CrossRef] [PubMed]
- Cummins, P.M. Occludin: One protein, many forms. Mol. Cell Biol. 2012, 32, 242–250. [Google Scholar] [CrossRef]
- Musch, M.W.; Walsh-Reitz, M.M.; Chang, E.B. Roles of ZO-1, occludin, and actin in oxidant-induced barrier disruption. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G222–G231. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Švagelj, D.; Terzić, V.; Dovhanj, J.; Švagelj, M.; Cvrković, M.; Švagelj, I. Superoxide dismutases in chronic gastritis. Apmis 2016, 124, 252–256. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Roswell, M.; Dushoff, J.; Winfree, R. A conceptual guide to measuring species diversity. Oikos 2021, 130, 321–338. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Mori, A.S.; Isbell, F.; Seidl, R. β-Diversity, Community Assembly, and Ecosystem Functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Altman, N.; Krzywinski, M. Clustering. Nat. Methods 2017, 14, 545–546. [Google Scholar] [CrossRef]
- Sun, L.; Dong, S.; Ge, Y.; Fonseca, J.P.; Robinson, Z.T.; Mysore, K.S.; Mehta, P. DiVenn: An Interactive and Integrated Web-Based Visualization Tool for Comparing Gene Lists. Front. Genet. 2019, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Dubourg, G.; Lagier, J.C.; Armougom, F.; Robert, C.; Audoly, G.; Papazian, L.; Raoult, D. High-level colonisation of the human gut by Verrucomicrobia following broad-spectrum antibiotic treatment. Int. J. Antimicrob. Agents 2013, 41, 149–155. [Google Scholar] [CrossRef]
- Fugang, W.; Rixing, B.; Rixing, B.; Ming, Y.; Maomin, S. Current status and perspectives of intestinal flora in the study of metabolic surgical mechanisms. Chin. Electron. J. Obes. Metab. Dis. 2018, 4, 40–43. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. Biomed. Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef]
- Wang, Z.; Shao, S.L.; Xu, X.H.; Zhao, X.; Wang, M.Y.; Chen, A.; Cong, H.Y. Helicobacter pylori and gastric microbiota homeostasis: Progress and prospects. Future Microbiol. 2023, 18, 137–157. [Google Scholar] [CrossRef]
- Rostamian, M.; Rahmati, D.; Akya, A. Clinical manifestations, associated diseases, diagnosis, and treatment of human infections caused by Erysipelothrix rhusiopathiae: A systematic review. Germs 2022, 12, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Tillotson, G.; MacKenzie, T.N.; Warren, C.A.; Wexler, H.M.; Goldstein, E.J.C. Bacteroides and related species: The keystone taxa of the human gut microbiota. Anaerobe 2024, 85, 102819. [Google Scholar] [CrossRef] [PubMed]
- Wenjie, L.; Chenyang, L.; Xu, H.; Xuan, L.; Guoxin, Z. Progress in the study of Mucinophilic Ackermannia and prospects for its application. Chin. J. Gastroenterol. 2022, 42, 277–280. [Google Scholar] [CrossRef]
- Arato, V.; Raso, M.M.; Gasperini, G.; Berlanda Scorza, F.; Micoli, F. Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat. Int. J. Mol. Sci. 2021, 22, 4042. [Google Scholar] [CrossRef]
- Ranjan, K.P.; Ranjan, N. Citrobacter: An emerging health care associated urinary pathogen. Urol. Ann. 2013, 5, 313–314. [Google Scholar] [CrossRef]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile infection. Nat. Rev. Dis. Primers 2016, 2, 16020. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Lin, X.; Hu, T.; Wu, Z.; Li, L.; Wang, Y.; Wen, D.; Liu, X.; Li, W.; Liang, H.; Jin, X.; et al. Isolation of potentially novel species expands the genomic and functional diversity of Lachnospiraceae. Imeta 2024, 3, e174. [Google Scholar] [CrossRef]
- Mittal, R.; Sanchez-Luege, S.V.; Wagner, S.M.; Yan, D.; Liu, X.Z. Recent Perspectives on Gene-Microbe Interactions Determining Predisposition to Otitis Media. Front. Genet. 2019, 10, 1230. [Google Scholar] [CrossRef]
- Hin, L.S.; Kin, L.; Yu, Z.; Caiyuan, Y.; Yijie, W. Advances in the study of Alistipes and their relevance to disease. Chin. J. Microbiol. Immunol. 2023, 43, 642–646. [Google Scholar] [CrossRef]
- Yang, Y.; Li, M.; Liu, Q.; Zhao, Q.; Zeng, J.; Wang, Q.; Zhao, Y.; Du, F.; Chen, Y.; Shen, J.; et al. Starch from Pueraria lobata and the amylose fraction alleviates dextran sodium sulfate induced colitis in mice. Carbohydr. Polym. 2023, 302, 120329. [Google Scholar] [CrossRef] [PubMed]
- Salvetti, E.; Torriani, S.; Felis, G.E. The Genus Lactobacillus: A Taxonomic Update. Probiotics Antimicrob. Proteins 2012, 4, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, M.; You, X.; Zhao, J.; Chen, L.; Wang, L.; Luo, Y.; Chen, Y. Gut microbiota mediates the anti-obesity effect of calorie restriction in mice. Sci. Rep. 2018, 8, 13037. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, B.; Zhang, X.; Akbar, M.T.; Wu, T.; Zhang, Y.; Zhi, L.; Shen, Q. Exploration of the Muribaculaceae Family in the Gut Microbiota: Diversity, Metabolism, and Function. Nutrients 2024, 16, 2660. [Google Scholar] [CrossRef]
- Zhang, Y.; Xing, H.; Bolotnikov, G.; Krämer, M.; Gotzmann, N.; Knippschild, U.; Kissmann, A.K.; Rosenau, F. Enriched Aptamer Libraries in Fluorescence-Based Assays for Rikenella microfusus-Specific Gut Microbiome Analyses. Microorganisms 2023, 11, 2266. [Google Scholar] [CrossRef]
- Jia, B.; Baek, J.H.; Lee, J.K.; Sun, Y.; Kim, K.H.; Jung, J.Y.; Jeon, C.O. Expanding the β-Lactamase Family in the Human Microbiome. Adv. Sci. 2024, 2403563. [Google Scholar] [CrossRef]
- Jiaxuan, J. Screening of Pig-Derived Probiotics and Their Resistance to Salmonella Infection. Master’s Thesis, Southwest University, Chongqing, China, 2017. [Google Scholar]
- Pérez-Cobas, A.E.; Gosalbes, M.J.; Friedrichs, A.; Knecht, H.; Artacho, A.; Eismann, K.; Otto, W.; Rojo, D.; Bargiela, R.; von Bergen, M.; et al. Gut microbiota disturbance during antibiotic therapy: A multi-omic approach. Gut 2013, 62, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, Y.; Ouyang, K.; Chen, L.; Zhao, M.; Wang, W. Sulfated Cyclocarya paliurus polysaccharides improve immune function of immunosuppressed mice by modulating intestinal microbiota. Int. J. Biol. Macromol. 2022, 212, 31–42. [Google Scholar] [CrossRef]
- Lynch, J.B.; Gonzalez, E.L.; Choy, K.; Faull, K.F.; Jewell, T.; Arellano, A.; Liang, J.; Yu, K.B.; Paramo, J.; Hsiao, E.Y. Gut microbiota Turicibacter strains differentially modify bile acids and host lipids. Nat. Commun. 2023, 14, 3669. [Google Scholar] [CrossRef]
- Clavel, T.; Lepage, P.; Charrier, C. The Family Coriobacteriaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Legaria, M.C.; Nastro, M.; Camporro, J.; Heger, F.; Barberis, C.; Stecher, D.; Rodriguez, C.H.; Vay, C.A. Peptostreptococcus anaerobius: Pathogenicity, identification, and antimicrobial susceptibility. Review of monobacterial infections and addition of a case of urinary tract infection directly identified from a urine sample by MALDI-TOF MS. Anaerobe 2021, 72, 102461. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi Abdolmaleky, H.; Zhou, J.R. Gut Microbiota Dysbiosis, Oxidative Stress, Inflammation, and Epigenetic Alterations in Metabolic Diseases. Antioxidants 2024, 13, 985. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; He, L.H.; Xu, L.J.; Li, S.B. Short-chain fatty acids: Bridges between diet, gut microbiota, and health. J. Gastroenterol. Hepatol. 2024, 39, 1728–1736. [Google Scholar] [CrossRef]
- Tong, Y.; Lei, B. Research progress of physiological function of short-chain fatty acids in the intestine. Adv. Clin. Med. 2022, 12, 939–945. [Google Scholar] [CrossRef]
- Mann, E.R.; Lam, Y.K.; Uhlig, H.H. Short-chain fatty acids: Linking diet, the microbiome and immunity. Nat. Rev. Immunol. 2024, 24, 577–595. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Gu, C.; Shuo, W.; Huang, S.; Lihong, W.; Zhihua, Z. Study on the role and mechanism of short-chain fatty acids on antibiotic-associated diarrhoea. J. Clin. Med. Pract. 2023, 27, 61–66. [Google Scholar] [CrossRef]
- Ohara, T.; Yoshino, K.; Kitajima, M. Possibility of preventing colorectal carcinogenesis with probiotics. Hepatogastroenterology 2010, 57, 1411–1415. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (g) | Organ Coefficient (mg/g) | ||||
|---|---|---|---|---|---|---|
| Day 70 | Heart | Liver | Spleen | Lung | Kidney | |
| Control | 30.77 ± 1.67 a | 4.44 ± 0.60 a | 37.71 ± 3.95 a | 2.60 ± 0.70 a | 4.68 ± 0.56 a | 10.68 ± 0.68 a |
| HP_NaCl | 29.34 ± 1.45 a | 5.19 ± 1.51 ab | 33.64 ± 2.98 b | 2.54 ± 0.29 a | 4.87 ± 0.70 a | 9.92 ± 0.57 ab |
| HP_LP61 | 29.91 ± 1.41 a | 5.70 ± 1.05 ab | 34.27 ± 3.34 ab | 2.47 ± 0.21 a | 5.19 ± 0.77 a | 10.68 ± 1.11 a |
| HP_LS03 | 29.61 ± 2.41 a | 5.61 ± 1.14 ab | 35.97 ± 2.90 ab | 2.47 ± 0.51 a | 5.21 ± 1.08 a | 9.89 ± 0.48 ab |
| HP_Ant | 29.86 ± 1.30 a | 5.28 ± 0.91 ab | 33.32 ± 2.67 b | 2.41 ± 0.34 a | 5.03 ± 0.43 a | 10.51 ± 0.75 a |
| Ant_LS03 | 28.48 ± 1.53 ab | 5.92 ± 1.00 b | 35.85 ± 3.35 ab | 2.67 ± 0.60 a | 5.47 ± 0.71 ab | 10.85 ± 1.36 a |
| NaCl_HP | 29.77 ± 1.34 a | 6.06 ± 1.99 b | 32.09 ± 2.20 b | 2.05 ± 0.71 ab | 5.21 ± 1.17 a | 10.23 ± 1.66 a |
| LP61_HP | 30.36 ± 2.53 a | 5.18 ± 1.28 ab | 32.64 ± 2.41 b | 2.24 ± 0.28 ab | 5.32 ± 0.75 a | 10.80 ± 1.15 a |
| LS03_HP | 30.08 ± 1.15 a | 5.07 ± 0.60 ab | 34.48 ± 2.93 ab | 2.30 ± 0.29 ab | 4.75 ± 0.80 a | 9.81 ± 0.67 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xu, X.; Yang, S.; Liu, K.; Wu, M.; Xie, M.; Xiong, T. Helicobacter pylori Inhibition, Gastritis Attenuation, and Gut Microbiota Protection in C57BL/6 Mice by Ligilactobacillus salivarius NCUH062003. Microorganisms 2024, 12, 2521. https://doi.org/10.3390/microorganisms12122521
Li J, Xu X, Yang S, Liu K, Wu M, Xie M, Xiong T. Helicobacter pylori Inhibition, Gastritis Attenuation, and Gut Microbiota Protection in C57BL/6 Mice by Ligilactobacillus salivarius NCUH062003. Microorganisms. 2024; 12(12):2521. https://doi.org/10.3390/microorganisms12122521
Chicago/Turabian StyleLi, Junyi, Xiaoyan Xu, Shiyu Yang, Kui Liu, Min Wu, Mingyong Xie, and Tao Xiong. 2024. "Helicobacter pylori Inhibition, Gastritis Attenuation, and Gut Microbiota Protection in C57BL/6 Mice by Ligilactobacillus salivarius NCUH062003" Microorganisms 12, no. 12: 2521. https://doi.org/10.3390/microorganisms12122521
APA StyleLi, J., Xu, X., Yang, S., Liu, K., Wu, M., Xie, M., & Xiong, T. (2024). Helicobacter pylori Inhibition, Gastritis Attenuation, and Gut Microbiota Protection in C57BL/6 Mice by Ligilactobacillus salivarius NCUH062003. Microorganisms, 12(12), 2521. https://doi.org/10.3390/microorganisms12122521