
The Human Mycobiome: Colonization, Composition and the Role in Health and Disease


Abstract
1. Introduction
2. Gut Mycobiome
2.1. Composition of Gut Mycobiome
2.2. Gut Colonization and Shaping of Gut Mycobiome
2.3. Gut Mycobiome Dysbiosis in Disease
3. Oral Mycobiome
3.1. Composition of Oral Mycobiome
3.2. Oral Cavity Colonization and Shaping of Oral Mycobiome
3.3. Oral Mycobiome Dysbiosis in Disease
4. Skin Mycobiome
4.1. Composition of Skin Mycobiome
4.2. Skin Colonization and Shaping of Skin Mycobiome
4.3. Skin Mycobiome Dysbiosis in Disease
5. Genitourinary Tract Mycobiome
5.1. Composition of Genitourinary Mycobiome
5.2. Genitourinary Tract Colonization and Shaping of Genitourinary Mycobiome
5.3. Genitourinary Mycobiome Dysbiosis in Disease
6. Respiratory Tract Mycobiome
6.1. Composition of Respiratory Mycobiome
6.2. Respiratory Tract Colonization and Shaping of Respiratory Mycobiome
6.3. Respiratory Tract Mycobiome Dysbiosis in Disease
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matijasic, M.; Mestrovic, T.; Paljetak, H.C.; Peric, M.; Baresic, A.; Verbanac, D. Gut Microbiota beyond Bacteria-Mycobiome, Virome, Archaeome, and Eukaryotic Parasites in IBD. Int. J. Mol. Sci. 2020, 21, 2668. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Seed, P.C. The Human Mycobiome. Cold Spring Harb. Perspect. Med. 2015, 5, a019810. [Google Scholar] [CrossRef] [PubMed]
- Gouba, N.; Drancourt, M. Digestive Tract Mycobiota: A Source of Infection. Médecine Et Mal. Infect. 2015, 45, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Iliev, I.D. The Mycobiota: Interactions between Commensal Fungi and the Host Immune System. Nat. Rev. Immunol. 2014, 14, 405–416. [Google Scholar] [CrossRef]
- Santus, W.; Devlin, J.R.; Behnsen, J. Crossing Kingdoms: How the Mycobiota and Fungal-Bacterial Interactions Impact Host Health and Disease. Infect. Immun. 2021, 89, e00648–20. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the Oral Fungal Microbiome (Mycobiome) in Healthy Individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef]
- Drell, T.; Lillsaar, T.; Tummeleht, L.; Simm, J.; Aaspõllu, A.; Väin, E.; Saarma, I.; Salumets, A.; Donders, G.G.G.; Metsis, M. Characterization of the Vaginal Micro- and Mycobiome in Asymptomatic Reproductive-Age Estonian Women. PLoS ONE 2013, 8, e54379. [Google Scholar] [CrossRef]
- Oh, J.; Freeman, A.F.; NISC Comparative Sequencing Program; Park, M.; Sokolic, R.; Candotti, F.; Holland, S.M.; Segre, J.A.; Kong, H.H. The Altered Landscape of the Human Skin Microbiome in Patients with Primary Immunodeficiencies. Genome Res. 2013, 23, 2103–2114. [Google Scholar] [CrossRef]
- van Woerden, H.C.; Gregory, C.; Brown, R.; Marchesi, J.R.; Hoogendoorn, B.; Matthews, I.P. Differences in Fungi Present in Induced Sputum Samples from Asthma Patients and Non-Atopic Controls: A Community Based Case Control Study. BMC Infect. Dis. 2013, 13, 69. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Frau, A.; Kenny, J.G.; Lenzi, L.; Campbell, B.J.; Ijaz, U.Z.; Duckworth, C.A.; Burkitt, M.D.; Hall, N.; Anson, J.; Darby, A.C.; et al. DNA Extraction and Amplicon Production Strategies Deeply Inf Luence the Outcome of Gut Mycobiome Studies. Sci. Rep. 2019, 9, 9328. [Google Scholar] [CrossRef] [PubMed]
- Huseyin, C.E.; Rubio, R.C.; O’Sullivan, O.; Cotter, P.D.; Scanlan, P.D. The Fungal Frontier: A Comparative Analysis of Methods Used in the Study of the Human Gut Mycobiome. Front. Microbiol. 2017, 8, 1432. [Google Scholar] [CrossRef]
- De Filippis, F.; Laiola, M.; Blaiotta, G.; Ercolini, D. Different Amplicon Targets for Sequencing-Based Studies of Fungal Diversity. Appl. Environ. Microbiol. 2017, 83, e00905-17. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Huseyin, C.E.; O’Toole, P.W.; Cotter, P.D.; Scanlan, P.D. Forgotten Fungi—the Gut Mycobiome in Human Health and Disease. Fems Microbiol. Rev. 2017, 41, 479–511. [Google Scholar] [CrossRef]
- Tiew, P.Y.; Mac Aogain, M.; Ali, N.A.B.M.; Thng, K.X.; Goh, K.; Lau, K.J.X.; Chotirmall, S.H. The Mycobiome in Health and Disease: Emerging Concepts, Methodologies and Challenges. Mycopathologia 2020, 185, 207–231. [Google Scholar] [CrossRef]
- Cui, L.; Morris, A.; Ghedin, E. The Human Mycobiome in Health and Disease. Genome Med. 2013, 5, 63. [Google Scholar] [CrossRef]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; NIH Intramural Sequencing Center Comparative Sequencing Program; et al. Topographic Diversity of Fungal and Bacterial Communities in Human Skin. Nature 2013, 498, 367. [Google Scholar] [CrossRef]
- Cui, L.; Lucht, L.; Tipton, L.; Rogers, M.B.; Fitch, A.; Kessinger, C.; Camp, D.; Kingsley, L.; Leo, N.; Greenblatt, R.M.; et al. Topographic Diversity of the Respiratory Tract Mycobiome and Alteration in HIV and Lung Disease. Am. J. Respir. Crit. Care Med. 2015, 191, 932–942. [Google Scholar] [CrossRef]
- Chin, V.K.; Yong, V.C.; Chong, P.P.; Amin Nordin, S.; Basir, R.; Abdullah, M. Mycobiome in the Gut: A Multiperspective Review. Mediat. Inflamm. 2020, 2020, 9560684. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Vandenborght, L.-E.; Coron, N.; Bazin, T.; Prevel, R.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Lamireau, T.; Delhaes, L. The Mycobiome: A Neglected Component in the Microbiota-Gut-Brain Axis. Microorganisms 2018, 6, UNSP 22. [Google Scholar] [CrossRef]
- Hallen-Adams, H.E.; Suhr, M.J. Fungi in the Healthy Human Gastrointestinal Tract. Virulence 2016, 8, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Lai, G.C.; Tan, T.G.; Pavelka, N. The Mammalian Mycobiome: A Complex System in a Dynamic Relationship with the Host. Wiley Interdiscip. Rev. Syst. Biol. Med. 2018, 11, e1438. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D.; Bernstein, C.N.; Tremlett, H.; Van Domselaar, G.; Knox, N.C. A Fungal World: Could the Gut Mycobiome Be Involved in Neurological Disease? Front. Microbiol. 2019, 9, 3249. [Google Scholar] [CrossRef]
- Schei, K.; Avershina, E.; Øien, T.; Rudi, K.; Follestad, T.; Salamati, S.; Ødegård, R.A. Early Gut Mycobiota and Mother-Offspring Transfer. Microbiome 2017, 5, 107. [Google Scholar] [CrossRef]
- Wampach, L.; Heintz-Buschart, A.; Hogan, A.; Muller, E.E.L.; Narayanasamy, S.; Laczny, C.C.; Hugerth, L.W.; Bindl, L.; Bottu, J.; Andersson, A.F.; et al. Colonization and Succession within the Human Gut Microbiome by Archaea, Bacteria, and Microeukaryotes during the First Year of Life. Front. Microbiol. 2017, 8, 738. [Google Scholar] [CrossRef]
- Ward, T.L.; Knights, D.; Gale, C.A. Infant Fungal Communities: Current Knowledge and Research Opportunities. BMC Med. 2017, 15, 30. [Google Scholar] [CrossRef]
- Suhr, M.J.; Banjara, N.; Hallen-Adams, H.E. Sequence-based methods for detecting and evaluating the human gut mycobiome. Lett. Appl. Microbiol. 2016, 62, 209–215. [Google Scholar] [CrossRef]
- Hallen-Adams, H.E.; Kachman, S.D.; Kim, J.; Legge, R.M.; Martínez, I. Fungi Inhabiting the Healthy Human Gastrointestinal Tract: A Diverse and Dynamic Community. Fungal Ecol. 2015, 15, 9–17. [Google Scholar] [CrossRef]
- Borges, F.M.; de Paula, T.O.; Sarmiento, M.R.A.; de Oliveira, M.G.; Pereira, M.L.M.; Toledo, I.V.; Nascimento, T.C.; Ferreira-Machado, A.B.; Silva, V.L.; Diniz, C.G. Fungal Diversity of Human Gut Microbiota Among Eutrophic, Overweight, and Obese Individuals Based on Aerobic Culture-Dependent Approach. Curr. Microbiol. 2018, 75, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Gouba, N.; Raoult, D.; Drancourt, M. Gut Microeukaryotes during Anorexia Nervosa: A Case Report. BMC Res. Notes 2014, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Mar Rodriguez, M.; Perez, D.; Javier Chaves, F.; Esteve, E.; Marin-Garcia, P.; Xifra, G.; Vendrell, J.; Jove, M.; Pamplona, R.; Ricart, W.; et al. Obesity Changes the Human Gut Mycobiome. Sci. Rep. 2015, 5, 14600. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Dollive, S.; Grunberg, S.; Chen, J.; Li, H.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Archaea and Fungi of the Human Gut Microbiome: Correlations with Diet and Bacterial Residents. PLoS ONE 2013, 8, e66019. [Google Scholar] [CrossRef]
- Strati, F.; Di Paola, M.; Stefanini, I.; Albanese, D.; Rizzetto, L.; Lionetti, P.; Calabrò, A.; Jousson, O.; Donati, C.; Cavalieri, D.; et al. Age and Gender Affect the Composition of Fungal Population of the Human Gastrointestinal Tract. Front. Microbiol. 2016, 7, 1227. [Google Scholar] [CrossRef]
- Jo, J.-H.; Deming, C.; Kennedy, E.A.; Conlan, S.; Polley, E.C.; Ng, W.-I.; Segre, J.A.; Kong, H.H. Diverse Human Skin Fungal Communities in Children Converge in Adulthood. J. Invest. Dermatol. 2016, 136, 2356–2363. [Google Scholar] [CrossRef]
- Ezeonu, I.M.; Ntun, N.W.; Ugwu, K.O. Intestinal Candidiasis and Antibiotic Usage in Children: Case Study of Nsukka, South Eastern Nigeria. Afr. Health Sci. 2017, 17, 1178–1184. [Google Scholar] [CrossRef][Green Version]
- Tao, R.; Wang, R.; Wan, Z.; Song, Y.; Wu, Y.; Li, R. Ketoconazole 2% Cream Alters the Skin Fungal Microbiome in Seborrhoeic Dermatitis: A Cohort Study. Clin. Exp. Dermatol. 2022, 47, 1088–1096. [Google Scholar] [CrossRef]
- Leung, M.H.Y.; Chan, K.C.K.; Lee, P.K.H. Skin Fungal Community and Its Correlation with Bacterial Community of Urban Chinese Individuals. Microbiome 2016, 4, 46. [Google Scholar] [CrossRef]
- Fechney, J.M.; Browne, G.V.; Prabhu, N.; Irinyi, L.; Meyer, W.; Hughes, T.; Bockmann, M.; Townsend, G.; Salehi, H.; Adler, C.J. Preliminary Study of the Oral Mycobiome of Children with and without Dental Caries. J. Oral Microbiology 2019, 11, 1536182. [Google Scholar] [CrossRef]
- Mishra, K.; Bukavina, L.; Ghannoum, M. Symbiosis and Dysbiosis of the Human Mycobiome. Front. Microbiol. 2021, 12, 636131. [Google Scholar] [CrossRef] [PubMed]
- Sam, Q.H.; Chang, M.W.; Chai, L.Y.A. The Fungal Mycobiome and Its Interaction with Gut Bacteria in the Host. Int. J. Mol. Sci. 2017, 18, 330. [Google Scholar] [CrossRef] [PubMed]
- Mims, T.S.; Abdallah, Q.A.; Stewart, J.D.; Watts, S.P.; White, C.T.; Rousselle, T.V.; Gosain, A.; Bajwa, A.; Han, J.C.; Willis, K.A.; et al. The Gut Mycobiome of Healthy Mice Is Shaped by the Environment and Correlates with Metabolic Outcomes in Response to Diet. Commun. Biol. 2021, 4, 281. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, I.; Gao, I.H.; Lin, W.-Y.; Allen, M.; Li, X.V.; Fiers, W.D.; De Celie, M.B.; Putzel, G.G.; Yantiss, R.K.; Johncilla, M.; et al. Mucosal Fungi Promote Gut Barrier Function and Social Behavior via Type 17 Immunity. Cell 2022, 185, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, S.; Amaretti, A.; Gozzoli, C.; Simone, M.; Righini, L.; Candeliere, F.; Brun, P.; Ardizzoni, A.; Colombari, B.; Paulone, S.; et al. Longitudinal Survey of Fungi in the Human Gut: ITS Profiling, Phenotyping, and Colonization. Front. Microbiol. 2019, 10, 1575. [Google Scholar] [CrossRef]
- Schulze, J.; Sonnenborn, U. Yeasts in the Gut: From Commensals to Infectious Agents. Dtsch. Arztebl. Int. 2009, 106, 837–842. [Google Scholar]
- Li, Q.; Wang, C.; Tang, C.; He, Q.; Li, N.; Li, J. Dysbiosis of Gut Fungal Microbiota Is Associated with Mucosal Inflammation in Crohn’s Disease. J. Clin. Gastroenterol. 2014, 48, 513–523. [Google Scholar] [CrossRef]
- Huët, M.A.L.; Wong, L.W.; Goh, C.B.S.; Hussain, M.H.; Muzahid, N.H.; Dwiyanto, J.; Lee, S.W.H.; Ayub, Q.; Reidpath, D.; Lee, S.M.; et al. Investigation of Culturable Human Gut Mycobiota from the Segamat Community in Johor, Malaysia. World. J. Microbiol. Biotechnol. 2021, 37, 113. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Guo, R.; Chen, N.; Lu, H.; Huang, S.; Wang, J.; Li, L. Correlation between Gastrointestinal Fungi and Varying Degrees of Chronic Hepatitis B Virus Infection. Diagn. Microbiol. Infect. Dis. 2011, 70, 492–498. [Google Scholar] [CrossRef]
- Khatib, R.; Riederer, K.M.; Ramanathan, J.; Baran, J. Faecal Fungal Flora in Healthy Volunteers and Inpatients. Mycoses 2001, 44, 151–156. [Google Scholar] [CrossRef]
- Hamad, I.; Sokhna, C.; Raoult, D.; Bittar, F. Molecular Detection of Eukaryotes in a Single Human Stool Sample from Senegal. PLoS ONE 2012, 7, e40888. [Google Scholar] [CrossRef] [PubMed]
- Motooka, D.; Fujimoto, K.; Tanaka, R.; Yaguchi, T.; Gotoh, K.; Maeda, Y.; Furuta, Y.; Kurakawa, T.; Goto, N.; Yasunaga, T.; et al. Fungal ITS1 Deep-Sequencing Strategies to Reconstruct the Composition of a 26-Species Community and Evaluation of the Gut Mycobiota of Healthy Japanese Individuals. Front. Microbiol. 2017, 8, 238. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.K.; Siddharth, J.; Verma, P.; Bavdekar, A.; Patole, M.S.; Shouche, Y.S. Molecular Typing of Fecal Eukaryotic Microbiota of Human Infants and Their Respective Mothers. J. Biosci. 2012, 37, 221–226. [Google Scholar] [CrossRef]
- Kabwe, M.H.; Vikram, S.; Mulaudzi, K.; Jansson, J.K.; Makhalanyane, T.P. The Gut Mycobiota of Rural and Urban Individuals Is Shaped by Geography. BMC Microbiol. 2020, 20, 257. [Google Scholar] [CrossRef]
- Botschuijver, S.; Roeselers, G.; Levin, E.; Jonkers, D.M.; Welting, O.; Heinsbroek, S.E.M.; de Weerd, H.H.; Boekhout, T.; Fornai, M.; Masclee, A.A.; et al. Intestinal Fungal Dysbiosis Is Associated with Visceral Hypersensitivity in Patients with Irritable Bowel Syndrome and Rats. Gastroenterology 2017, 153, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Ott, S.J.; Kühbacher, T.; Musfeldt, M.; Rosenstiel, P.; Hellmig, S.; Rehman, A.; Drews, O.; Weichert, W.; Timmis, K.N.; Schreiber, S. Fungi and Inflammatory Bowel Diseases: Alterations of Composition and Diversity. Scand. J. Gastroenterol. 2008, 43, 831–841. [Google Scholar] [CrossRef]
- Coker, O.O.; Nakatsu, G.; Dai, R.Z.; Wu, W.K.K.; Wong, S.H.; Ng, S.C.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. Enteric Fungal Microbiota Dysbiosis and Ecological Alterations in Colorectal Cancer. Gut 2019, 68, 654–662. [Google Scholar] [CrossRef]
- Gouba, N.; Raoult, D.; Drancourt, M. Plant and Fungal Diversity in Gut Microbiota as Revealed by Molecular and Culture Investigations. PLoS ONE 2013, 8, e59474. [Google Scholar] [CrossRef]
- Agırbaslı, H.; Özcan, S.A.K.; Gedikoğlu, G. Fecal Fungal Flora of Pediatric Healthy Volunteers and Immunosuppressed Patients. Mycopathologia 2005, 159, 515–520. [Google Scholar] [CrossRef]
- Robert, V.; Stegehuis, G.; Stalpers, J. The MycoBank Engine and Related Databases. 2005. Available online: www.mycobank.org (accessed on 15 August 2022).
- Auchtung, T.A.; Fofanova, T.Y.; Stewart, C.J.; Nash, A.K.; Wong, M.C.; Gesell, J.R.; Auchtung, J.M.; Ajami, N.J.; Petrosino, J.F. Investigating Colonization of the Healthy Adult Gastrointestinal Tract by Fungi. mSphere 2018, 3, e00092. [Google Scholar] [CrossRef]
- Richard, M.L.; Lamas, B.; Liguori, G.; Hoffmann, T.W.; Sokol, H. Gut Fungal Microbiota: The Yin and Yang of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2015, 21, 656–665. [Google Scholar] [CrossRef]
- Nagata, R.; Nagano, H.; Ogishima, D.; Nakamura, Y.; Hiruma, M.; Sugita, T. Transmission of the Major Skin Microbiota, Malassezia, from Mother to Neonate. Pediatr. Int. 2012, 54, 350–355. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zuo, T.; Cheung, C.P.; Gu, W.; Wan, Y.; Zhang, F.; Chen, N.; Zhan, H.; Yeoh, Y.K.; Niu, J.; et al. Population-Level Configurations of Gut Mycobiome Across 6 Ethnicities in Urban and Rural China. Gastroenterology 2021, 160, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Moubasher, A.-A.H.; Abdel–Sater, M.A.; Soliman, Z. Biodiversity of Filamentous and Yeast Fungi in Citrus and Grape Fruits and Juices in Assiut Area, Egypt. JMBFS 2018, 7, 353–365. [Google Scholar] [CrossRef]
- Vadkertiová, R.; Molnárová, J.; Vránová, D.; Sláviková, E. Yeasts and Yeast-like Organisms Associated with Fruits and Blossoms of Different Fruit Trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Tournas, V.H.; Heeres, J.; Burgess, L. Moulds and Yeasts in Fruit Salads and Fruit Juices. Food Microbiol. 2006, 23, 684–688. [Google Scholar] [CrossRef]
- Tournas, V.; Niazi, N.; Kohn, J. Fungal Presence in Selected Tree Nuts and Dried Fruits. Microbiol. Insights 2015, 8, 1–6. [Google Scholar] [CrossRef]
- Tournas, V.H. Moulds and Yeasts in Fresh and Minimally Processed Vegetables, and Sprouts. Int. J. Food Microbiol. 2005, 99, 71–77. [Google Scholar] [CrossRef]
- Reed, G.; Nagodawithana, T.W. Use of Yeasts in the Dairy Industry. In Yeast Technology; Springer: Dordrecht, The Netherlands, 1990; pp. 441–445. [Google Scholar]
- Griffin, S.; Falzon, O.; Camilleri, K.; Valdramidis, V.P. Bacterial and Fungal Contaminants in Caprine and Ovine Cheese: A Meta-Analysis Assessment. Food Res. Int. 2020, 137, 109445. [Google Scholar] [CrossRef]
- Venturini Copetti, M. Yeasts and Molds in Fermented Food Production: An Ancient Bioprocess. Curr. Opin. Food Sci. 2019, 25, 57–61. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Irisawa, T.; Dicks, L.; Tanasupawat, S. FERMENTED FOODS|Fermentations of East and Southeast Asia. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 846–851. [Google Scholar]
- Suo, B.; Nie, W.; Wang, Y.; Ma, J.; Xing, X.; Huang, Z.; Xu, C.; Li, Z.; Ai, Z. Microbial Diversity of Fermented Dough and Volatile Compounds in Steamed Bread Prepared with Traditional Chinese Starters. LWT 2020, 126, 109350. [Google Scholar] [CrossRef]
- Li, Z.; Li, H.; Song, K.; Cui, M. Performance of Non-Saccharomyces Yeasts Isolated from Jiaozi in Dough Fermentation and Steamed Bread Making. LWT 2019, 111, 46–54. [Google Scholar] [CrossRef]
- Shah, S.; Locca, A.; Dorsett, Y.; Cantoni, C.; Ghezzi, L.; Lin, Q.; Bokoliya, S.; Panier, H.; Suther, C.; Gormley, M.; et al. Alterations of the Gut Mycobiome in Patients with MS. EBioMedicine 2021, 71, 103557. [Google Scholar] [CrossRef]
- Gosiewski, T.; Salamon, D.; Szopa, M.; Sroka, A.; Malecki, M.T.; Bulanda, M. Quantitative Evaluation of Fungi of the Genus Candida in the Feces of Adult Patients with Type 1 and 2 Diabetes—a Pilot Study. Gut. Pathog. 2014, 6, 43. [Google Scholar] [CrossRef]
- Chiaro, T.R.; Soto, R.; Stephens, W.Z.; Kubinak, J.L.; Petersen, C.; Gogokhia, L.; Bell, R.; Delgado, J.C.; Cox, J.; Voth, W.; et al. A Member of the Gut Mycobiota Modulates Host Purine Metabolism Exacerbating Colitis in Mice. Sci. Transl. Med. 2017, 9, eaaf9044. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.-P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal Microbiota Dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef]
- Hong, G.; Li, Y.; Yang, M.; Li, G.; Qian, W.; Xiong, H.; Bai, T.; Song, J.; Zhang, L.; Hou, X. Gut Fungal Dysbiosis and Altered Bacterial-Fungal Interaction in Patients with Diarrhea-Predominant Irritable Bowel Syndrome: An Explorative Study. Neurogastroenterol. Motil. 2020, 32, e13891. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Duan, Y.; Liu, J.; Torralba, M.G.; Kuelbs, C.; Ventura-Cots, M.; Abraldes, J.G.; Bosques-Padilla, F.; Verna, E.C.; Brown, R.S.; et al. Intestinal Fungal Dysbiosis and Systemic Immune Response to Fungi in Patients With Alcoholic Hepatitis. Hepatology 2020, 71, 522–538. [Google Scholar] [CrossRef]
- Jayasudha, R.; Das, T.; Kalyana Chakravarthy, S.; Sai Prashanthi, G.; Bhargava, A.; Tyagi, M.; Rani, P.K.; Pappuru, R.R.; Shivaji, S. Gut Mycobiomes Are Altered in People with Type 2 Diabetes Mellitus and Diabetic Retinopathy. PLoS ONE 2020, 15, e0243077. [Google Scholar] [CrossRef] [PubMed]
- Bhute, S.S.; Suryavanshi, M.V.; Joshi, S.M.; Yajnik, C.S.; Shouche, Y.S.; Ghaskadbi, S.S. Gut Microbial Diversity Assessment of Indian Type-2-Diabetics Reveals Alterations in Eubacteria, Archaea, and Eukaryotes. Front. Microbiol. 2017, 8, 214. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, J.; Vuorela, A.; Muthas, D.; Orivuori, L.; Luopajärvi, K.; Tejesvi, M.V.G.; Lavrinienko, A.; Pirttilä, A.M.; Fogarty, C.L.; Härkönen, T.; et al. Fungal Dysbiosis and Intestinal Inflammation in Children with Beta-Cell Autoimmunity. Front. Immunol. 2020, 11, 468. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Ali, S.; Shrode, R.L.; Shahi, S.K.; Jensen, S.N.; Hoang, J.; Cassidy, S.; Olalde, H.; Guseva, N.; Paullus, M.; et al. Multiple Sclerosis Patients Have an Altered Gut Mycobiome and Increased Fungal to Bacterial Richness. PLoS ONE 2022, 17, e0264556. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wei, S.; Gu, Y.; Wang, Y.; Feng, Y.; Sheng, J.; Hu, L.; Gu, C.; Jiang, P.; Tian, Y.; et al. Gut Mycobiome in Patients with Chronic Kidney Disease Was Altered and Associated with Immunological Profiles. Front. Immunol. 2022, 13, 843695. [Google Scholar] [CrossRef]
- Mok, K.; Suratanon, N.; Roytrakul, S.; Charoenlappanit, S.; Patumcharoenpol, P.; Chatchatee, P.; Vongsangnak, W.; Nakphaichit, M. ITS2 Sequencing and Targeted Meta-Proteomics of Infant Gut Mycobiome Reveal the Functional Role of Rhodotorula Sp. during Atopic Dermatitis Manifestation. J. Fungi 2021, 7, 748. [Google Scholar] [CrossRef]
- Pablo-Fernandez, E.D.; Gebeyehu, G.G.; Flain, L.; Slater, R.; Frau, A.; Ijaz, U.Z.; Warner, T.; Probert, C. The Faecal Metabolome and Mycobiome in Parkinson’s Disease. Parkinsonism Relat. Disord. 2022, 95, 65–69. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, L.; Zhang, Z.; Zhou, Y.; Jiang, H.; Ruan, B. Analysis of Gut Mycobiota in First-Episode, Drug-Naïve Chinese Patients with Schizophrenia: A Pilot Study. Behav. Brain Res. 2020, 379, 112374. [Google Scholar] [CrossRef]
- Gosalbes, M.J.; Jimenéz-Hernandéz, N.; Moreno, E.; Artacho, A.; Pons, X.; Ruíz-Pérez, S.; Navia, B.; Estrada, V.; Manzano, M.; Talavera-Rodriguez, A.; et al. Interactions among the Mycobiome, Bacteriome, Inflammation, and Diet in People Living with HIV. Gut Microbes 2022, 14, 2089002. [Google Scholar] [CrossRef]
- Chehoud, C.; Albenberg, L.G.; Judge, C.; Hoffmann, C.; Grunberg, S.; Bittinger, K.; Baldassano, R.N.; Lewis, J.D.; Bushman, F.D.; Wu, G.D. Fungal Signature in the Gut Microbiota of Pediatric Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2015, 21, 1948–1956. [Google Scholar] [CrossRef]
- Hoarau, G.; Mukherjee, P.K.; Gower-Rousseau, C.; Hager, C.; Chandra, J.; Retuerto, M.A.; Neut, C.; Vermeire, S.; Clemente, J.; Colombel, J.F.; et al. Bacteriome and Mycobiome Interactions Underscore Microbial Dysbiosis in Familial Crohn’s Disease. mBio 2016, 7, e01250-16. [Google Scholar] [CrossRef] [PubMed]
- Liguori, G.; Lamas, B.; Richard, M.L.; Brandi, G.; da Costa, G.; Hoffmann, T.W.; Di Simone, M.P.; Calabrese, C.; Poggioli, G.; Langella, P.; et al. Fungal Dysbiosis in Mucosa-Associated Microbiota of Crohn’s Disease Patients. J. Crohns Colitis 2016, 10, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Chen, E.Z.; Baldassano, R.N.; Otley, A.R.; Griffiths, A.M.; Lee, D.; Bittinger, K.; Bailey, A.; Friedman, E.S.; Hoffmann, C.; et al. Inflammation, Antibiotics, and Diet as Environmental Stressors of the Gut Microbiome in Pediatric Crohn’s Disease. Cell Host Microbe 2015, 18, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Prochazkova, P.; Roubalova, R.; Dvorak, J.; Kreisinger, J.; Hill, M.; Tlaskalova-Hogenova, H.; Tomasova, P.; Pelantova, H.; Cermakova, M.; Kuzma, M.; et al. The Intestinal Microbiota and Metabolites in Patients with Anorexia Nervosa. Gut Microbes 2021, 13, 1902771. [Google Scholar] [CrossRef] [PubMed]
- Li, B.-Y.; Xu, X.-Y.; Gan, R.-Y.; Sun, Q.-C.; Meng, J.-M.; Shang, A.; Mao, Q.-Q.; Li, H.-B. Targeting Gut Microbiota for the Prevention and Management of Diabetes Mellitus by Dietary Natural Products. Foods 2019, 8, 440. [Google Scholar] [CrossRef] [PubMed]
- Soyucen, E.; Gulcan, A.; Aktuglu-Zeybek, A.C.; Onal, H.; Kiykim, E.; Aydin, A. Differences in the Gut Microbiota of Healthy Children and Those with Type 1 Diabetes. Pediatr. Int. 2014, 56, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Monteiro-da-Silva, F.; Araujo, R.; Sampaio-Maia, B. Interindividual Variability and Intraindividual Stability of Oral Fungal Microbiota over Time. Med. Mycol. 2014, 52, 498–505. [Google Scholar] [CrossRef]
- Li, Y.; Wang, K.; Zhang, B.; Tu, Q.; Yao, Y.; Cui, B.; Ren, B.; He, J.; Shen, X.; Van Nostrand, J.D.; et al. Salivary Mycobiome Dysbiosis and Its Potential Impact on Bacteriome Shifts and Host Immunity in Oral Lichen Planus. Int. J. Oral Sci. 2019, 11, 1–10. [Google Scholar] [CrossRef]
- Oba, P.M.; Holscher, H.D.; Mathai, R.A.; Kim, J.; Swanson, K.S. Diet Influences the Oral Microbiota of Infants during the First Six Months of Life. Nutrients 2020, 12, 3400. [Google Scholar] [CrossRef]
- Azevedo, M.J.; Pereira, M.d.L.; Araujo, R.; Ramalho, C.; Zaura, E.; Sampaio-Maia, B. Influence of Delivery and Feeding Mode in Oral Fungi Colonization—A Systematic Review. Microb. Cell 2020, 7, 36–45. [Google Scholar] [CrossRef]
- Diaz, P.I.; Dongari-Bagtzoglou, A. Critically Appraising the Significance of the Oral Mycobiome. J. Dent. Res. 2021, 100, 133–140. [Google Scholar] [CrossRef]
- Peters, B.A.; Wu, J.; Hayes, R.B.; Ahn, J. The Oral Fungal Mycobiome: Characteristics and Relation to Periodontitis in a Pilot Study. BMC Microbiol. 2017, 17, 157. [Google Scholar] [CrossRef]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the Human Oral Mycobiome with Improved Practices in Amplicon-Based Taxonomy: Discovery of Malassezia as a Prominent Commensal. PLoS ONE 2014, 9, e90899. [Google Scholar] [CrossRef]
- Khadija, B.; Imran, M.; Faryal, R. Keystone Salivary Mycobiome in Postpartum Period in Health and Disease Conditions. J. Mycol. Med. 2021, 31, 101101. [Google Scholar] [CrossRef]
- Stehlikova, Z.; Tlaskal, V.; Galanova, N.; Roubalova, R.; Kreisinger, J.; Dvorak, J.; Prochazkova, P.; Kostovcikova, K.; Bartova, J.; Libanska, M.; et al. Oral Microbiota Composition and Antimicrobial Antibody Response in Patients with Recurrent Aphthous Stomatitis. Microorganisms 2019, 7, 636. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.I.; Hong, B.-Y.; Dupuy, A.K.; Strausbaugh, L.D. Mining the Oral Mycobiome: Methods, Components, and Meaning. Virulence 2017, 8, 313–323. [Google Scholar] [CrossRef]
- Charlson, E.S.; Diamond, J.M.; Bittinger, K.; Fitzgerald, A.S.; Yadav, A.; Haas, A.R.; Bushman, F.D.; Collman, R.G. Lung-Enriched Organisms and Aberrant Bacterial and Fungal Respiratory Microbiota after Lung Transplant. Am. J. Respir. Crit. Care Med. 2012, 186, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Bandara, H.M.H.N.; Panduwawala, C.P.; Samaranayake, L.P. Biodiversity of the Human Oral Mycobiome in Health and Disease. Oral Dis. 2019, 25, 363–371. [Google Scholar] [CrossRef]
- Ikebe, K.; Morii, K.; Matsuda, K.; Hata, K.; Nokubi, T. Association of Candidal Activity with Denture Use and Salivary Flow in Symptom-Free Adults over 60 Years1. J. Oral Rehab. 2006, 33, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.L.; Dominguez-Bello, M.G.; Heisel, T.; Al-Ghalith, G.; Knights, D.; Gale, C.A. Development of the Human Mycobiome over the First Month of Life and across Body Sites. mSystems 2018, 3, e00140–17. [Google Scholar] [CrossRef] [PubMed]
- Darwazeh, A.M.; al-Bashir, A. Oral Candidal Flora in Healthy Infants. J. Oral Pathol. Med. 1995, 24, 361–364. [Google Scholar] [CrossRef]
- Burcham, Z.M.; Garneau, N.L.; Comstock, S.S.; Tucker, R.M.; Knight, R.; Metcalf, J.L. Genetics of Taste Lab Citizen Scientists Patterns of Oral Microbiota Diversity in Adults and Children: A Crowdsourced Population Study. Sci. Rep. 2020, 10, 2133. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, M.N.; Furuta, M.; Takeshita, T.; Shibata, Y.; Sundari, R.; Eshima, N.; Ninomiya, T.; Yamashita, Y. Oral Mycobiome in Community-Dwelling Elderly and Its Relation to Oral and General Health Conditions. Oral Dis. 2017, 23, 973–982. [Google Scholar] [CrossRef]
- O’Connell, L.M.; Santos, R.; Springer, G.; Burne, R.A.; Nascimento, M.M.; Richards, V.P. Site-Specific Profiling of the Dental Mycobiome Reveals Strong Taxonomic Shifts during Progression of Early-Childhood Caries. Appl. Environ. Microbiol. 2020, 86, e02825–19. [Google Scholar] [CrossRef]
- Tati, S.; Davidow, P.; McCall, A.; Hwang-Wong, E.; Rojas, I.G.; Cormack, B.; Edgerton, M. Candida Glabrata Binding to Candida Albicans Hyphae Enables Its Development in Oropharyngeal Candidiasis. PLoS Pathog. 2016, 12, e1005522. [Google Scholar] [CrossRef]
- Sajid, M.; Sharma, P.; Srivastava, S.; Hariprasad, R.; Singh, H.; Bharadwaj, M. Smokeless Tobacco Consumption Induces Dysbiosis of Oral Mycobiome: A Pilot Study. Appl. Microbiol. Biotechnol. 2022, 106, 5643–5657. [Google Scholar] [CrossRef]
- Zhu, T.; Duan, Y.-Y.; Kong, F.-Q.; Galzote, C.; Quan, Z.-X. Dynamics of Skin Mycobiome in Infants. Front. Microbiol. 2020, 11, 1790. [Google Scholar] [CrossRef] [PubMed]
- Ratanapokasatit, Y.; Laisuan, W.; Rattananukrom, T.; Petchlorlian, A.; Thaipisuttikul, I.; Sompornrattanaphan, M. How Microbiomes Affect Skin Aging: The Updated Evidence and Current Perspectives. Life 2022, 12, 936. [Google Scholar] [CrossRef] [PubMed]
- Boxberger, M.; Cenizo, V.; Cassir, N.; La Scola, B. Challenges in Exploring and Manipulating the Human Skin Microbiome. Microbiome 2021, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.-H.; Kennedy, E.A.; Kong, H.H. Topographical and Physiological Differences of the Skin Mycobiome in Health and Disease. Virulence 2016, 8, 324–333. [Google Scholar] [CrossRef]
- Li, H.; Goh, B.N.; Teh, W.K.; Jiang, Z.; Goh, J.P.Z.; Goh, A.; Wu, G.; Hoon, S.S.; Raida, M.; Camattari, A.; et al. Skin Commensal Malassezia Globosa Secreted Protease Attenuates Staphylococcus Aureus Biofilm Formation. J. Investig. Dermatol. 2018, 138, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Leong, C.; Schmid, B.; Toi, M.J.; Wang, J.; Irudayaswamy, A.S.; Goh, J.P.Z.; Bosshard, P.P.; Glatz, M.; Dawson, T.L. Geographical and Ethnic Differences Influence Culturable Commensal Yeast Diversity on Healthy Skin. Front. Microbiol. 2019, 10, 1891. [Google Scholar] [CrossRef] [PubMed]
- Huffnagle, G.B.; Noverr, M.C. The Emerging World of the Fungal Microbiome. Trends Microbiol. 2013, 21, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Keum, H.L.; Kim, H.; Kim, H.-J.; Park, T.; Kim, S.; An, S.; Sul, W.J. Structures of the Skin Microbiome and Mycobiome Depending on Skin Sensitivity. Microorganisms 2020, 8, E1032. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The Skin Microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Cho, Y.-J.; Kim, T.; Croll, D.; Park, M.; Kim, D.; Keum, H.L.; Sul, W.J.; Jung, W.H. Genome of Malassezia Arunalokei and Its Distribution on Facial Skin. Microbiol. Spectr. 2022, 10, e00506-22. [Google Scholar] [CrossRef]
- Gupta, A.K.; Kohli, Y.; Summerbell, R.C.; Faergemann, J. Quantitative Culture of Malassezia Species from Different Body Sites of Individuals with or without Dermatoses. Med. Mycol. 2001, 39, 243–251. [Google Scholar] [CrossRef]
- Tong, X.; Leung, M.H.Y.; Wilkins, D.; Cheung, H.H.L.; Lee, P.K.H. Neutral Processes Drive Seasonal Assembly of the Skin Mycobiome. mSystems 2019, 4, e00004-19. [Google Scholar] [CrossRef]
- Koike, Y.; Kuwatsuka, S.; Nishimoto, K.; Motooka, D.; Murota, H. Skin Mycobiome of Psoriasis Patients Is Retained during Treatment with TNF and IL-17 Inhibitors. Int. J. Mol. Sci. 2020, 21, 3892. [Google Scholar] [CrossRef]
- Crespo-Erchiga, V.; Florencio, V.D. Malassezia Yeasts and Pityriasis Versicolor. Curr. Opin. Infect. Dis. 2006, 19, 139–147. [Google Scholar] [CrossRef]
- Moosbrugger-Martinz, V.; Hackl, H.; Gruber, R.; Pilecky, M.; Knabl, L.; Orth-Höller, D.; Dubrac, S. Initial Evidence of Distinguishable Bacterial and Fungal Dysbiosis in the Skin of Patients with Atopic Dermatitis or Netherton Syndrome. J. Investig. Dermatol. 2021, 141, 114–123. [Google Scholar] [CrossRef]
- Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J.; Bohac, P.; Pinto, Y.; Uzan, A.; et al. Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-Occurrence of Fungal and Bacterial Communities. Front. Microbiol. 2019, 10, 438. [Google Scholar] [CrossRef]
- Bjerre, R.D.; Holm, J.B.; Palleja, A.; Sølberg, J.; Skov, L.; Johansen, J.D. Skin Dysbiosis in the Microbiome in Atopic Dermatitis Is Site-Specific and Involves Bacteria, Fungus and Virus. BMC Microbiol. 2021, 21, 256. [Google Scholar] [CrossRef] [PubMed]
- Dylag, M.; Leniak, E.; Gnat, S.; Szepietowski, J.C.; Kozubowski, L. A Case of Anti-Pityriasis Versicolortherapy That Preserves Healthy Mycobiome. BMC Dermatol. 2020, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Honnavar, P.; Prasad, G.S.; Ghosh, A.; Dogra, S.; Handa, S.; Rudramurthy, S.M. Malassezia Arunalokei Sp. Nov., a Novel Yeast Species Isolated from Seborrheic Dermatitis Patients and Healthy Individuals from India. J. Clin. Microbiol. 2016, 54, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Sahni, K.; Singh, S.; Dogra, S. Newer Topical Treatments in Skin and Nail Dermatophyte Infections. Indian Dermatol. Online J. 2018, 9, 149–158. [Google Scholar] [PubMed]
- Chng, K.R.; Tay, A.S.L.; Li, C.; Ng, A.H.Q.; Wang, J.; Suri, B.K.; Matta, S.A.; McGovern, N.; Janela, B.; Wong, X.F.C.C.; et al. Whole Metagenome Profiling Reveals Skin Microbiome-Dependent Susceptibility to Atopic Dermatitis Flare. Nat. Microbiol. 2016, 1, 1–10. [Google Scholar] [CrossRef]
- Crespo Erchiga, V.; Ojeda Martos, A.; Vera Casaño, A.; Crespo Erchiga, A.; Sanchez Fajardo, F. Malassezia Globosa as the Causative Agent of Pityriasis Versicolor. Br. J. Dermatol. 2000, 143, 799–803. [Google Scholar] [CrossRef]
- Shah, A.; Koticha, A.; Ubale, M.; Wanjare, S.; Mehta, P.; Khopkar, U. Identification and Speciation of Malassezia in Patients Clinically Suspected of Having Pityriasis Versicolor. Indian. J. Dermatol. 2013, 58, 239. [Google Scholar] [CrossRef]
- Carter, B.; Jones, C.P.; Creadick, R.N.; Parker, R.T.; Turner, V. The Vaginal Fungi. Ann. N.Y. Acad. Sci. 1959, 83, 265–279. [Google Scholar] [CrossRef]
- Bradford, L.L.; Ravel, J. The Vaginal Mycobiome: A Contemporary Perspective on Fungi in Women’s Health and Diseases. Virulence 2017, 8, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, A.L.; Underhill, D.M. The Mycobiome of the Human Urinary Tract: Potential Roles for Fungi in Urology. Ann. Transl. Med. 2017, 5, 31. [Google Scholar] [CrossRef]
- Guo, R.; Zheng, N.; Lu, H.; Yin, H.; Yao, J.; Chen, Y. Increased Diversity of Fungal Flora in the Vagina of Patients with Recurrent Vaginal Candidiasis and Allergic Rhinitis. Microb. Ecol. 2012, 64, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.M.; Hilt, E.E.; Rosenfeld, A.B.; Zilliox, M.J.; Thomas-White, K.; Fok, C.; Kliethermes, S.; Schreckenberger, P.C.; Brubaker, L.; Gai, X.; et al. The Female Urinary Microbiome: A Comparison of Women with and without Urgency Urinary Incontinence. mBio 2014, 5, e01283-01214. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.; Young, M.; Lee, O.; Chen, C.-A. Vulvovaginal Carriage of Yeasts Other than Candida Albicans. Sex. Transm. Infect. 2003, 79, 249–250. [Google Scholar] [CrossRef]
- Papaemmanouil, V.; Georgogiannis, N.; Plega, M.; Lalaki, J.; Lydakis, D.; Dimitriou, M.; Papadimitriou, A. Prevalence and Susceptibility of Saccharomyces Cerevisiae Causing Vaginitis in Greek Women. Anaerobe 2011, 17, 298–299. [Google Scholar] [CrossRef]
- Kalia, N.; Singh, J.; Kaur, M. Microbiota in Vaginal Health and Pathogenesis of Recurrent Vulvovaginal Infections: A Critical Review. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 5. [Google Scholar] [CrossRef]
- Pytka, M.; Kordowska-Wiater, M.; Jarocki, P. Mikrobiome of the Women’s Genital System. Adv. Microbiol. 2019, 58, 227–236. [Google Scholar]
- Noverr, M.C.; Huffnagle, G.B. Does the Microbiota Regulate Immune Responses Outside the Gut? Trends Microbiol. 2004, 12, 562–568. [Google Scholar] [CrossRef]
- Sabbatini, S.; Visconti, S.; Gentili, M.; Lusenti, E.; Nunzi, E.; Ronchetti, S.; Perito, S.; Gaziano, R.; Monari, C. Lactobacillus Iners Cell-Free Supernatant Enhances Biofilm Formation and Hyphal/Pseudohyphal Growth by Candida Albicans Vaginal Isolates. Microorganisms 2021, 9, 2577. [Google Scholar] [CrossRef]
- Liu, N.-N.; Zhao, X.; Tan, J.-C.; Liu, S.; Li, B.-W.; Xu, W.-X.; Peng, L.; Gu, P.; Li, W.; Shapiro, R.; et al. Mycobiome Dysbiosis in Women with Intrauterine Adhesions. Microbiol. Spectr. 2022, 10, e01324-22. [Google Scholar] [CrossRef]
- Kovachev, S.M. Cervical Cancer and Vaginal Microbiota Changes. Arch. Microbiol. 2020, 202, 323–327. [Google Scholar] [CrossRef]
- Tipton, L.; Ghedin, E.; Morris, A. The Lung Mycobiome in the Next-Generation Sequencing Era. Virulence 2017, 8, 334–341. [Google Scholar] [CrossRef]
- Krause, R.; Moissl-Eichinger, C.; Halwachs, B.; Gorkiewicz, G.; Berg, G.; Valentin, T.; Prattes, J.; Högenauer, C.; Zollner-Schwetz, I. Mycobiome in the Lower Respiratory Tract—A Clinical Perspective. Front. Microbiol. 2017, 7, 2169. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.D.N.; Viscogliosi, E.; Delhaes, L. The Lung Mycobiome: An Emerging Field of the Human Respiratory Microbiome. Front. Microbiol. 2015, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Carpagnano, G.E.; Susca, A.; Scioscia, G.; Lacedonia, D.; Cotugno, G.; Soccio, P.; Santamaria, S.; Resta, O.; Logrieco, G.; Barbaro, M.P.F. A Survey of Fungal Microbiota in Airways of Healthy Volunteer Subjects from Puglia (Apulia), Italy. BMC Infect. Dis. 2019, 19, 78. [Google Scholar] [CrossRef] [PubMed]
- Gago, S.; Overton, N.L.D.; Ben-Ghazzi, N.; Novak-Frazer, L.; Read, N.D.; Denning, D.W.; Bowyer, P. Lung Colonization by Aspergillus Fumigatus Is Controlled by ZNF77. Nat. Commun. 2018, 9, 3835. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, E.M.H.; Eagan, T.M.L.; Leiten, E.O.; Haaland, I.; Husebø, G.R.; Knudsen, K.S.; Drengenes, C.; Sanseverino, W.; Paytuví-Gallart, A.; Nielsen, R. The Pulmonary Mycobiome—A Study of Subjects with and without Chronic Obstructive Pulmonary Disease. PLoS ONE 2021, 16, e0248967. [Google Scholar] [CrossRef]
- Ali, N.A.B.M.; Mac Aogain, M.; Morales, R.F.; Tiew, P.Y.; Chotirmall, S.H. Optimisation and Benchmarking of Targeted Amplicon Sequencing for Mycobiome Analysis of Respiratory Specimens. Int. J. Mol. Sci. 2019, 20, 4991. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Clark, S.T.; Surendra, A.; Copeland, J.K.; Wang, P.W.; Ammar, R.; Collins, C.; Tullis, D.E.; Nislow, C.; Hwang, D.M.; et al. Global Analysis of the Fungal Microbiome in Cystic Fibrosis Patients Reveals Loss of Function of the Transcriptional Repressor Nrg1 as a Mechanism of Pathogen Adaptation. PLOS Pathog. 2015, 11, e1005308. [Google Scholar] [CrossRef]
- Kramer, R.; Sauer-Heilborn, A.; Welte, T.; Guzman, C.A.; Abraham, W.-R.; Hoefle, M.G. Cohort Study of Airway Mycobiome in Adult Cystic Fibrosis Patients: Differences in Community Structure between Fungi and Bacteria Reveal Predominance of Transient Fungal Elements. J. Clin. Microbiol. 2015, 53, 2900–2907. [Google Scholar] [CrossRef] [PubMed]
- Soret, P.; Vandenborght, L.-E.; Francis, F.; Coron, N.; Enaud, R.; Avalos, M.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Thiebaut, R.; et al. Respiratory Mycobiome and Suggestion of Inter-Kingdom Network during Acute Pulmonary Exacerbation in Cystic Fibrosis. Sci. Rep. 2020, 10, 3589. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.; Gago, S.; Bromley, M.; Bowyer, P. The Human Lung Mycobiome in Chronic Respiratory Disease: Limitations of Methods and Our Current Understanding. Curr. Fungal Infect. Rep. 2019, 13, 109–119. [Google Scholar] [CrossRef]
- Huffnagle, G.B. The Microbiota and Allergies/Asthma. PLOS Pathog. 2010, 6, e1000549. [Google Scholar] [CrossRef]



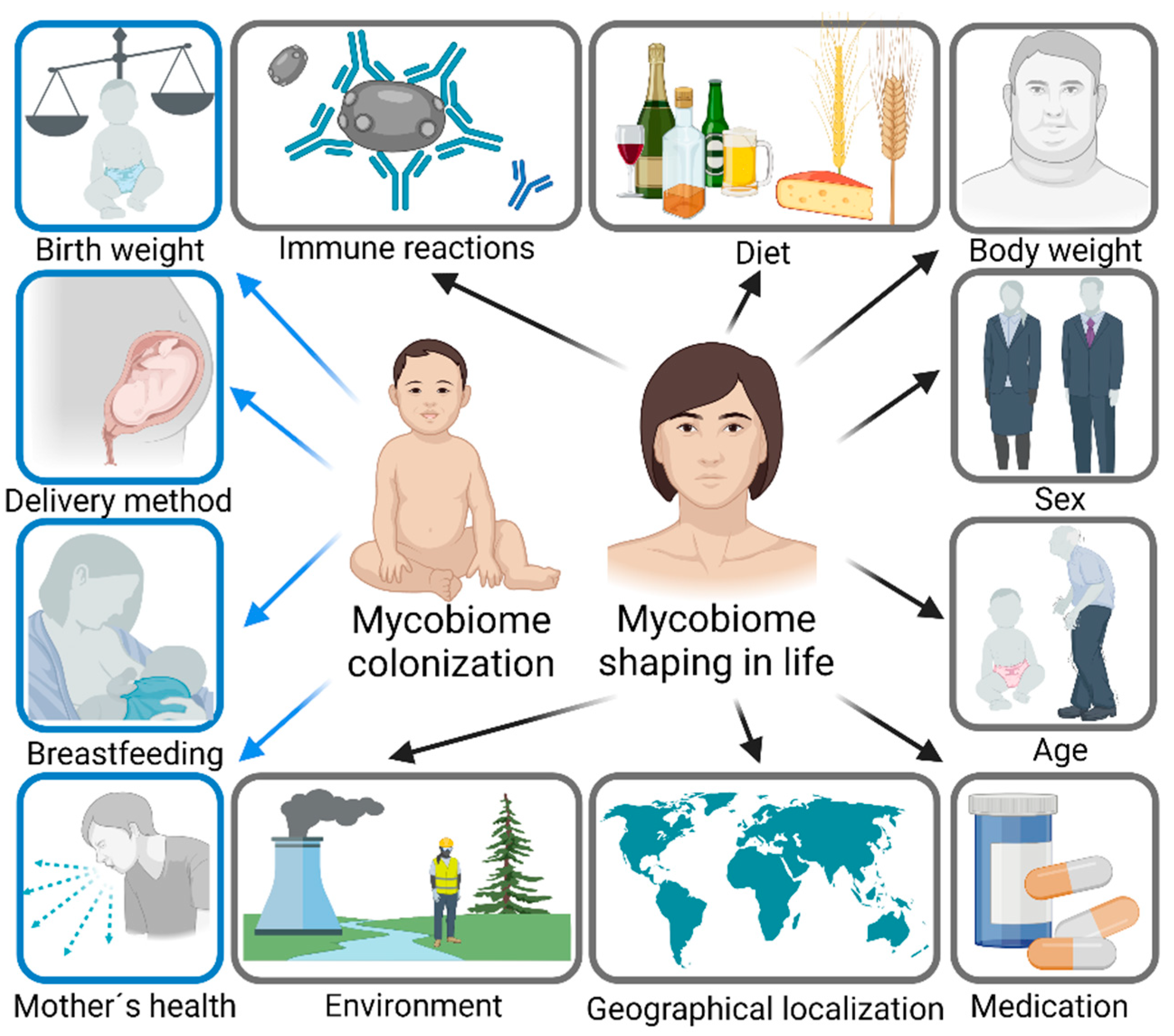{kind=link}
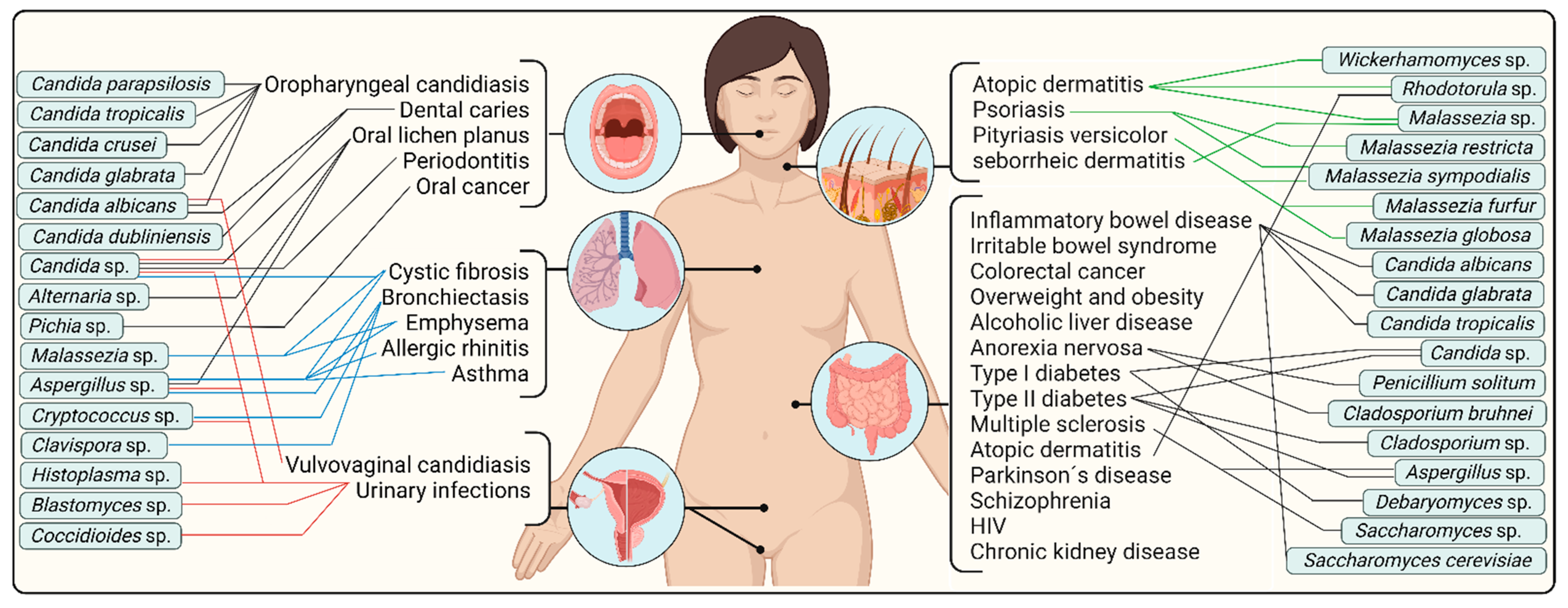{kind=link}
| Genus | The Most Common Species | Abundance/ Frequency | Number of Species | List of all Species | References |
|---|---|---|---|---|---|
| Saccharomyces | S. cerevisiae | ~100% [47]; 96.8%[11]; 23% [48]; 12.80% [49]; 6.94% [31]; 5.1% [50]; 1.14% [35] | 5 | S. cerevisiae, S. paradoxus, S. caariocanus, S. bayanus, S. castellii | [11,31,35,47,48,49,50,51,52,53,54,55,56,57] |
| Candida | Candida sp. | 59.72% [31]; 2.48% [49] | 21 | C. albicans, C. tropicalis, C. dubliniensis, C. glabrata, C. krissi, C. rugosa, C. austromarina, C. parapsilosis, C. lusitaniae, C. pararugosa, C. deformans, C. fermentati, C. intermedia, C. metapsilosis, C. zeylanoides, C. sake, C. krusei, C. kefyr, C. guilliermondii, C. lambica, C. humilis | [11,29,31,32,35,47,48,49,50,51,52,53,54,55,56,58,59] |
| C. albicans | 80.8%[11]; 62.7% [50]; 39.8% [35]; ~30% [47]; 28% [48]; 16.67% [31]; 11.16% [49]; 5.81% [56] | ||||
| Malassezia | M. restricta | 88.30% [11] | 5 | M. slooffiae, M. globosa, M. pachydermatis, M. restricta, M. sympodialis | [29,32,47,49,51,54,58] |
| Penicillium | Penicillium sp. | 65.28% [31]; 7% [11] | 14 | P. turbatum, P. chrysogenum, P. freii, P. camemberti, P. brevicompactum, P. allii, P. dipodomyicola, P. sacculum, P. italicum, P. glabrum, P. verrucosum, P. crutosum, P. paneum, P. roqueforti | [11,29,31,32,35,49,52,54,55,58] |
| P. brevicompactum | 3.72% [35] | ||||
| Aspergillus | Aspergillus sp. | 58.33% [31] | 16 | A. cristasus, A. flavipes, A. glaucus, A. pseudoglaucus, A. oryzeae, A. flavus, A. fumigatus, A. cf. niger, A. cf. carbonarius, A. cf. foetidus, A. cf. tubingensis, A. restrictus, A. versicolor, A. clavatus, A. ochraceoroseus, A. rambelli | [11,29,31,32,35,47,49,54,55,57,58] |
| A. versicolor | 4.95% [49] | ||||
| Galactomyces | G. candidum | 46.1%[11]; 6.98% [56]; 0.83% [49] | 2 | G. geotrichum, G. candidum | [11,32,49,51,56,58] |
| Trichosporon | Trichosporon sp. | 30.56% [31] | 4 | T. asahii, T. faecale, T. caseorum, T. cutaneum | [31,32,35,51,52,59] |
| T. asahii | 0.57% [35] | ||||
| Rhodotorula | Rhodotorula sp. | 8.50% [50] | 4 | R. mucilaginosa, R. rubra, R. graminis, R. glutinis | [31,32,35,50,54,55,56] |
| R. mucilaginosa | 15.28% [31]; 12.6% [35]; 6.98% [56] | ||||
| Geotrichum | G. silvicola | 9.72% [31] | 1 | G. silvicola | [31,32,50,51,59] |
| Cryptococcus | C. carnescens | 4.65% [56] | 6 | C. amylolyticus, C. carnescens, C. neoformans, C. saitoi, C. albidus, C. luteolus | [29,35,47,50,56] |
| Pichia | P. manshurica | 3.43% [35]; 1.39% [31] | 5 | P. kudriavzevii, P. fermentans, P. manshurica, P. kluyveri, P. caribica | [11,29,31,35,54] |
| Exophiala | E. dermatitidis | 1.39% [31] | 4 | E. mesophila, E. heteromorpha, E. equina, E. dermatitidis | [29,31,32,57] |
| Cladosporium | Cladosporium sp. | 1.16% [56] | 1 | C. cladosporioides | [11,54,55,56,58] |
| C. cladosporioides | not found | ||||
| Mucor | M. circinelloides | 0.57% [35] | 2 | M. circinelloides, M. racemosus | [31,35,53,54] |
| Alternaria | A. alternata | not found | 7 | A. alternata, A. cf. arborescens, A cf. brassicola, A. cf. citri, A.cf. mali, A. cf. tenuissima, A. metachromatica | [4,11,29,32,48,55] |
| Debaryomyces | D.fabryi | not found | 3 | D. hansenii, D. fabryi, D. prosopidis | [11,32,52,54,55,57] |
| Food Product | Microscopic Fungi | References |
|---|---|---|
| Fruit and vegetables | ||
| Fresh citrus and grape | Candida prunicola, Pichia fermentans | [66] |
| Fresh apple, plum and pear | Saccharomyces cerevisiae, Pichia kluyveri, Pichia kudriavzevii, Galactomyces candidus, Hanseniaspora uvarum, Hanseniaspora guilliermondii | [67] |
| Peeled fruit salads | Candida sp., Debaryomyces sp., Rhodotorula sp., Penicillium sp., Cladosporium sp. | [68] |
| Dried fruit | Cladosporium sp., Aspergillus niger, Aspergillus tubingiensis, Penicillium palitans | [68,69] |
| Various fresh vegetables (salad, tomato, cucumber, green inion, lettuce, spinach, etc.) | Geotrichum sp., Alternaria sp., Cladosporium sp., Penicillium sp. | [68,70] |
| Dairy | ||
| Various cheeses (Blue cheese, Camembert, Cheddar) | Penicillium sp., Candida sp., Scopulariopsis sp. | [64] |
| Acidophilus milk | Saccharomyces fragilis, Candida pseudotropicalis | [71,72] |
| Meat | ||
| Various meats (fermented sausage, dried meat, salami, ham) | Debaryomyces sp., Penicillium sp. | [64] |
| Beverages | ||
| Wine | Hanseniaspora sp., Saccharomyces sp. | [73,74] |
| Beer | Brettanomyces sp., Saccharomyces sp. | [73,74] |
| Sake | Aspergillus sp., Saccharomyces sp. | [73] |
| Other | ||
| Various nuts (pecan, almond, walnut, pine nut) | Aspergillus sp. Penicillium sp., Alternaria sp., Cladosporium sp., Rhizopus sp., Fusarium sp. | [69] |
| Koji | Aspergillus sp., Rhizopus sp. | [75] |
| Soy sauce | Aspergillus sp., Hansenula sp., Zygosaccharomyces sp. | [73] |
| Steamed pastry | Wickerhamomyces anomalus | [76,77] |
| Lipid Metabolism | |||||
|---|---|---|---|---|---|
| BMI Index and Fat Mass | Serum Total Cholesterol Fasting Triglycerides | LDL 1 Cholesterol | HDL 2 Cholesterol | References | |
| Negative correlations | Agaricomycetes | Eurotiomycetes | Mucoraceae | Sacharomycetes | [33,79] |
| Nectriaceae | Hypocraceae | Tremellomycetes | |||
| Mucoraceae | Mucoraceae | Mucor sp. | Cystobasidiomycetes | ||
| Mucor sp. | Mucor sp. | Candida sp. | Erythrobasidiaceae | ||
| Penicillium sp. | Candida sp. | ||||
| Positive correlations | Sacharomycetes | Dipodascaceae | _ | Aspergillaceae | [33,65,78]. |
| Tremellomycetes | Penicillium sp. | ||||
| Cystobasidiomycetes | Eurotiomycetes | ||||
| Erythrobasidiaceae | Candida dubliniensis | ||||
| Dipodascaceae | |||||
| Aspergillus sp. | |||||
| Hannaella sp. | |||||
| Carbohydrate Metabolism | |||||
|---|---|---|---|---|---|
| Glucose | Insulin | Glycated Haemoglobin | References | ||
| Negative correlations | Candida sp. C. dubliniensis | _ | Agaricomycetes | Mucor sp. | [33,80] |
| Ceratocystidaceae | Monilliela sp. | ||||
| Debariomycetaceae | Eupenicillium sp. | ||||
| Mucoraceae | |||||
| Corticiaceae | Ceratocystis sp. | ||||
| Positive correlations | Eurotium sp. | Ascomycota | Eurotium sp. | - | [33] |
| Genus | The Most Common Species | Abundance/ Frequency | Number of Species | List of All Species | References |
|---|---|---|---|---|---|
| Candida | Candida sp. | 100% [106]; 100% [105]; 85% [101]; 75% [7]; 67% [100] | 6 | C. albicans, C. khmerensis, C. metapsilosis, C. parapsilosis, C. tropicalis, C. glabrata | [7,61,100,101,105,106,107,108] |
| Aspergillus | Aspergillus sp. | 100% [106]; 100% [105]; 75% [100]; 45% [101]; 35% [7] | 7 | A. amstelodami, A. caesiellus, A. flavus, A. oryzae, A. penicillioides, A. ruber, A. niger | [7,61,100,101,105,106,107,108] |
| Penicillium | Penicillium sp. | 97% [105]; 85% [100]; 70% [101]; 50% [106] | 4 | P. brevicompactum, P. glabrum, P. spinulosum, P. citrinum | [7,61,100,101,105,107] |
| Cladosporium | Cladosporium sp. | 100% [106]; 72.5% [100]; 65% [7] | 4 | C. cladosporioides, C. herbarum, C. sphaerospermu, C. teniussimum | [7,61,100,106,107,108] |
| Phoma | Phoma sp. | 100% [106]; 65% [101] | 3 | P. foveata, P. plurivora, P. herbarum | [7,61,101,106] |
| Malassezia | Malassezia sp. | 100% [106]; 40% [101] | 3 | M. restricta, M. globosa, M. sympodialis | [106,107,108] |
| Cryptococcus | Cryptococcus sp. | 100% [106]; 20% [7] | 2 | C. cellulolyticus, C. diffluens | [7,106] |
| Alternaria | Alternaria sp. | 100% [106]; 5% [100] | 4 | A. tenuissima, A. triticina, A. alternata, A. armoraciae | [7,100,101,106,107,108] |
| Fusarium | Fusarium sp. | 83% [106]; 30% [7] | 3 | F. oxysporum, F. culmorum, F. poae | [7,107,108] |
| Rhodotorula | Rhodotorula sp. | 75% [100] | 1 | R. mucilaginosa | [100,107,108] |
| Aureobasidium | Aureobasidium sp. | 67% [106]; 50% [7] | 0 | not classified | [7,100,108] |
| Saccharomyces | Saccharomyces sp. | 50% [7]; 50% [106]; 45% [101] | 3 | S. bayanus, S. cerevisiae, S. ellipsoideus | [7,61,101,107] |
| Trichoderma | Trichoderma sp. | 50% [106]; 45% [101]; 10% [100] | 1 | T. aureoviride | [100,101,107] |
| Site | Genus | The Most Common Species | Abundance/ Frequency | Number of Species | List of All Species | References |
|---|---|---|---|---|---|---|
| Scalp | Malassezia | M sympodialis | 23.8–57.1% [125] | 7 | M. sympodialis, M. furfur, M. globosa, M. restricta, M. sympodialis, M. dermatis, M. slooffiae | [125,126] |
| Candida | C. albicans | 50–65% [126] | 2 | C. albicans, C. metapsilosis | [125,126] | |
| Aspergillus | Aspergillus sp. | not found | 0 | not classified | [125] | |
| Cryptococcus | Cryptococcus sp. | not found | 0 | not classified | [126] | |
| Aureobasidium | Aureobasidium sp. | not found | 0 | not classified | [125] | |
| Acremonium | Acremonium sp. | not found | 0 | not classified | [126] | |
| Cladosporium | Cladosporium sp. | not found | 0 | not classified | [125] | |
| Hortaea | Hortaea sp. | not found | 0 | not classified | [125] | |
| Trichosporon | Trichosporon sp. | not found | 0 | not classified | [125] | |
| Pallidocercospora | Pallidocercospora sp. | not found | 0 | not classified | [125] | |
| Didymella | Didymella sp. | not found | 0 | not classified | [126] | |
| Areas with a high density of sebaceous glands | Malassezia | M. globosa | 91% [127] 13.5–56.8% [125] | 6 | M. globosa, M. furfur, M. restricta, M. sympodialis, M. dermatis, M. slooffiae | [125,127] |
| Candida | C. parapsilosis | 6.25–35.1% [125] | 2 | C. parapsilosis, C. orthopsilosis | [125] | |
| Alternaria | Alternaria sp. | not found | 0 | not classified | [120] | |
| Cladosporium | Cladosporium sp. | not found | 0 | not classified | [120] | |
| Cryptococcus | Cryptococcus sp. | not found | 0 | not classified | [120] | |
| Schizophillum | Schizophillum sp. | not found | 0 | not classified | [120] | |
| Rhodotorula | R. mucilaginosa | 13.5% [120]; 0.62% [125] | 1 | R. mucilaginosa | [120,125] | |
| Penicillium | Penicillium sp. | not found | 0 | not classified | [125] | |
| Lodderomyces | L. elongisporus | not found | 1 | L. elongisporus | [125] | |
| Rhodosporium | R. toloroides | not found | 1 | R. toloroides | [125] | |
| Naganisha | N. albidosimilis | not found | 1 | N. albidosimilis | [125] | |
| Areas with a low density of sebaceous glands | Malassezia | Malassezia sp. | 24.48–43.43% [120] | 6 | M. globosa M. restricta, M. furfur, M. obtuse, M. sympodialis, M. japonica | [120] |
| Mucor | Mucor sp. | not found | 0 | not classified | [127] | |
| Neurospora | Neurospora sp. | not found | 0 | not classified | [127] | |
| Aspergillus | Aspergillus sp. | not found | 0 | not classified | [127] | |
| Preussia | Preussia sp. | not found | 0 | not classified | [127] | |
| Saccharomyces | Saccharomyces sp. | not found | 0 | not classified | [127] | |
| Rasamsonia | Rasamsonia sp. | not found | 0 | not classified | [120] | |
| Alternaria | Alternaria sp. | not found | 0 | not classified | [120] | |
| Cladosporium | Cladosporium sp. | 5.71–9.85% [120] | 0 | not classified | [120] | |
| Candida | Candida sp. | 4.42–7.05% [120] | 0 | not classified | [120] | |
| Cryptococcus | Cryptococcus sp. | not found | 0 | not classified | [120] | |
| Rhodotorula | Rhodotorula sp. | not found | 0 | not classified | [120] | |
| Penicillium | Penicillium sp. | not found | 0 | not classified | [120] |
| Site | Genus | The Most Common Species | Abundance/ Frequency | Number of Species | List of all Species | References |
|---|---|---|---|---|---|---|
| Urinary tract | Candida | Candida sp. | not found | 1 | not classified | [145,147] |
| Saccharomyces | Saccharomyces sp. | not found | 1 | not classified | [145] | |
| Malassezia | Malassezia sp. | not found | 1 | not classified | [145] | |
| Vagina | Candida | Candida sp. | 1–69% [148]; 36.9% [8] | 9 | C. albicans, C. glabrata, C. krusei, C. tropicalis, C. parapsilosis, C. dubliniensis, C. guillermondi, C. kefyr, C. humicolus, | [8,148] |
| C. albicans | 34.1 [8] | |||||
| Saccharomyces | S. cerevisiae | 5% [148] | 1 | S. cerevisiae | [148,149] | |
| Cladosporium | C. perangustum | >1% [8] | 1 | C. perangustum | [8] | |
| Alternaria | A. alternata | >1% [8] | 1 | A. alternata | [8] | |
| Rhodotorula | Rhodotorula sp. | >1% [8] | 0 | not classified | [8] |
| Site | Genus | The Most Common Species | Number of Species | List of Species | References |
|---|---|---|---|---|---|
| Lungs | Cladosporium | Cladosporium sp. | 0 | not classified | [156] |
| Penicillium | Penicillium sp. | 0 | not classified | [156] | |
| Aspergillus | Aspergillus sp. | 0 | not classified | [156] | |
| Candida | Candida sp. | 0 | not classified | [156] | |
| Malassezia | Malassezia sp. | 0 | not classified | [156] | |
| Kluyveromyces | Kluyveromyces sp. | 0 | not classified | [156] | |
| Pneumocystis | Pneumocystis sp. | 0 | not classified | [126,156] | |
| Bronchoalveolar lavage | Cladosporium | Cladosporium sp. | 0 | not classified | [145] |
| Candida | C. guilliermondii | 2 | C. guilliermondii, C. albicans | [20,156] | |
| Aspergillus | Aspergillus sp. | 2 | A. fumigatus, A. flavus | [145,156] | |
| Penicillium | Penicillium sp. | 0 | not classified | [145] | |
| Saccharomyces | S. cerevisiae | 0 | not classified | [20] | |
| Pichia | Pichia sp. | 2 | P. Jadini, P. brevicompactum | [20] | |
| Exhaled breath Condensate (EBC) | Cladosporium | Cladosporium sp. | 1 | C. herbarum | [159] |
| Aspergillus | A. sydowii | 1 | A. sydowii | [159] | |
| Penicillium | P. brevicompactum | 5 | P. brevicompactum, P. Expansum, P. glabrum, P. olsonii, P. bilaiae | [159] | |
| Alternaria | A. alternata | 2 | A. alternata, A. infectoria | [159] | |
| Sputum | Saccharomyces | Saccharomyces sp. | 0 | not classified | [162] |
| Candida | Candida sp. | 2 | C. albicans, C. dubliniensis | [162] | |
| Ganoderma | Ganoderma sp. | 0 | not classified | [20] | |
| Malassezia | Malassezia sp. | 0 | not classified | [162] | |
| Aspergillus | A. fumigatus | 1 | A. fumigatus | [20] | |
| Cryptococcus | C. magnus | 1 | C. magnus | [20] | |
| Peniophora | P. incarnata | 2 | P. Incarnata, P. Cinerea | [20] | |
| Daedaleopsis | D. confragosa | 1 | D. confragosa | [20] | |
| Sistotrema | S. brinkmannii | 1 | S. brinkmannii | [20] | |
| Stereum | S. hirsutum | 1 | S. hirsutum | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belvoncikova, P.; Splichalova, P.; Videnska, P.; Gardlik, R. The Human Mycobiome: Colonization, Composition and the Role in Health and Disease. J. Fungi 2022, 8, 1046. https://doi.org/10.3390/jof8101046
Belvoncikova P, Splichalova P, Videnska P, Gardlik R. The Human Mycobiome: Colonization, Composition and the Role in Health and Disease. Journal of Fungi. 2022; 8(10):1046. https://doi.org/10.3390/jof8101046
Chicago/Turabian StyleBelvoncikova, Paulina, Petra Splichalova, Petra Videnska, and Roman Gardlik. 2022. "The Human Mycobiome: Colonization, Composition and the Role in Health and Disease" Journal of Fungi 8, no. 10: 1046. https://doi.org/10.3390/jof8101046
APA StyleBelvoncikova, P., Splichalova, P., Videnska, P., & Gardlik, R. (2022). The Human Mycobiome: Colonization, Composition and the Role in Health and Disease. Journal of Fungi, 8(10), 1046. https://doi.org/10.3390/jof8101046