
N-Acetylcysteine Antagonizes NGF Activation of TrkA through Disulfide Bridge Interaction, an Effect Which May Contribute to Its Analgesic Activity

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
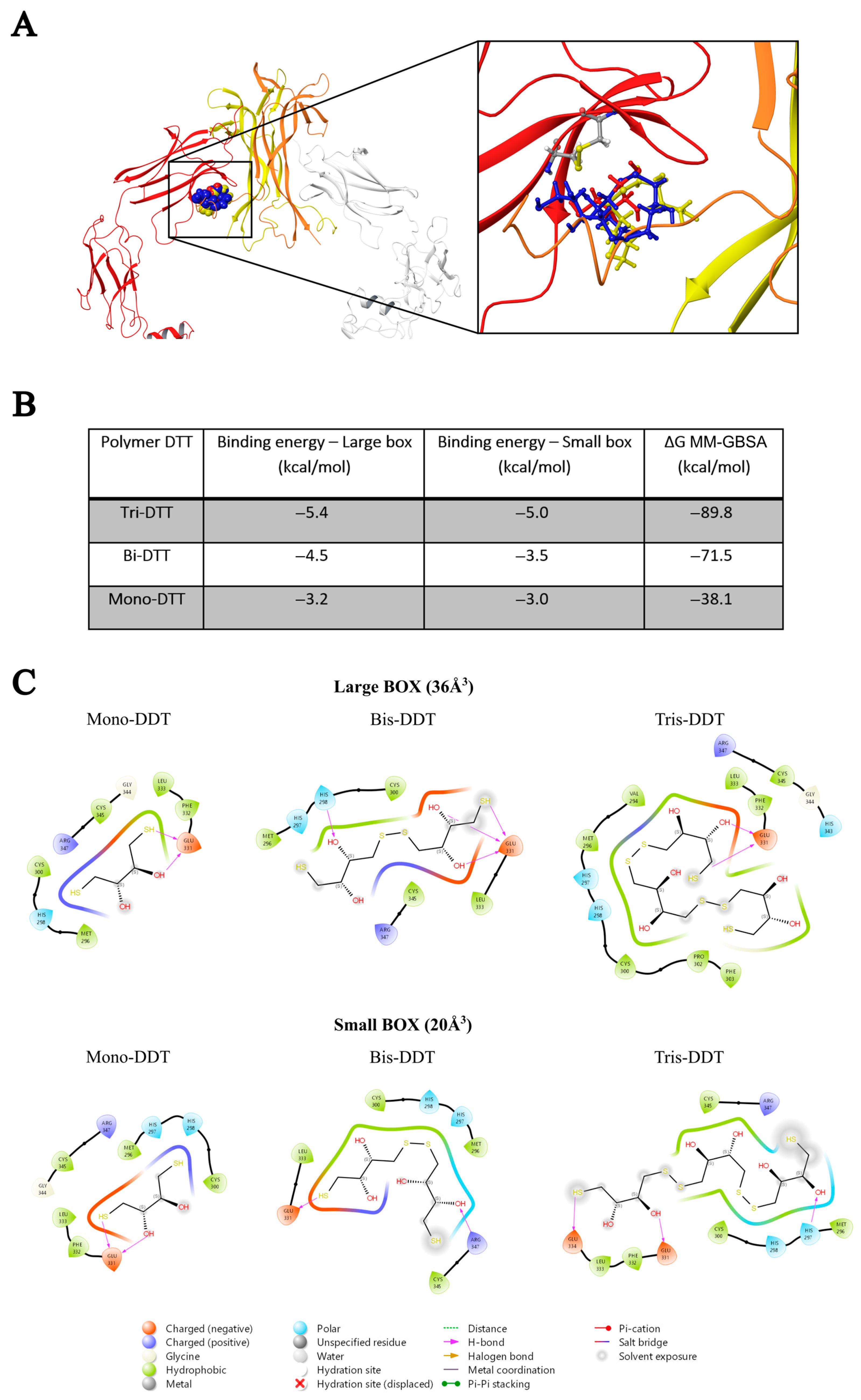
2.1. In Silico Studies
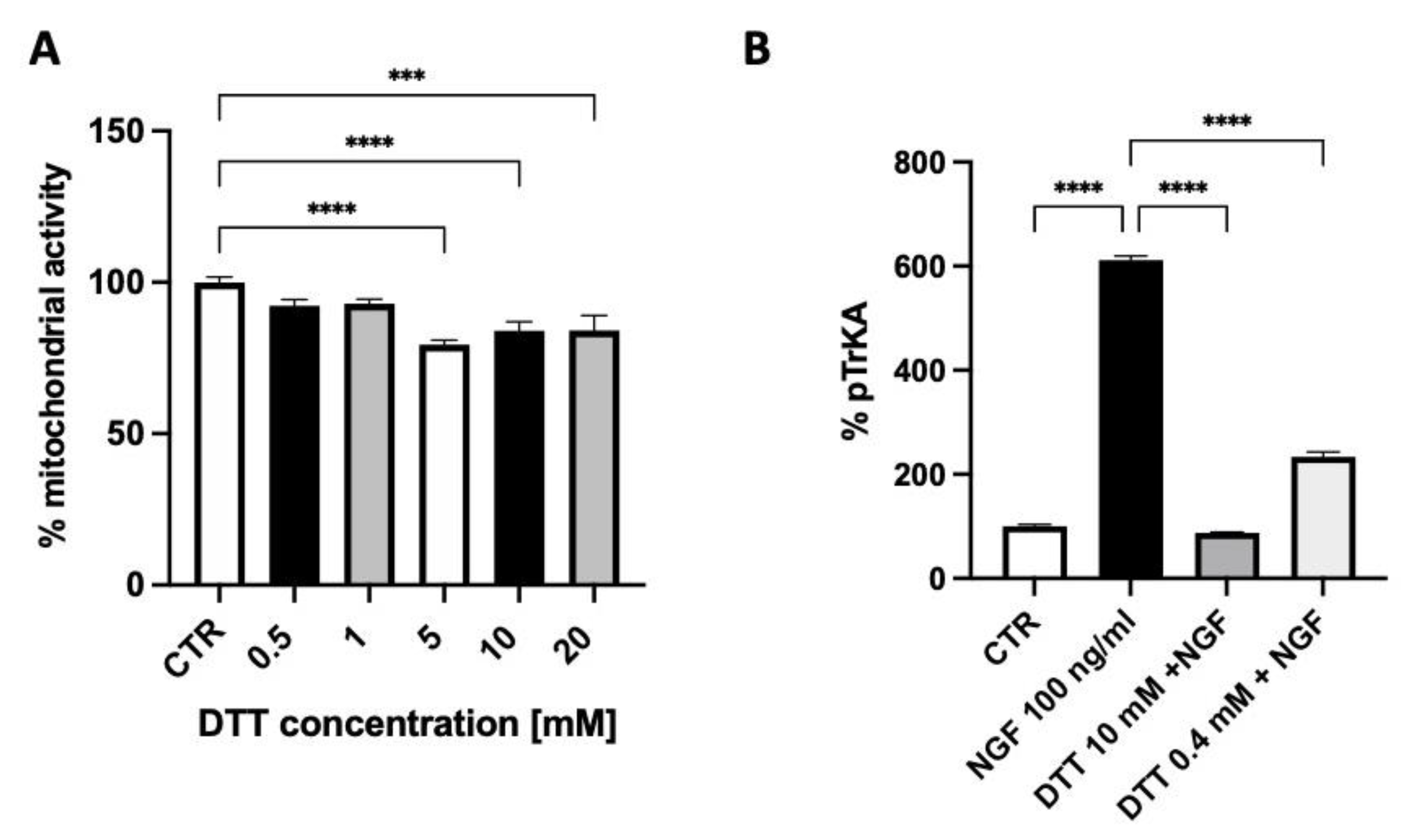
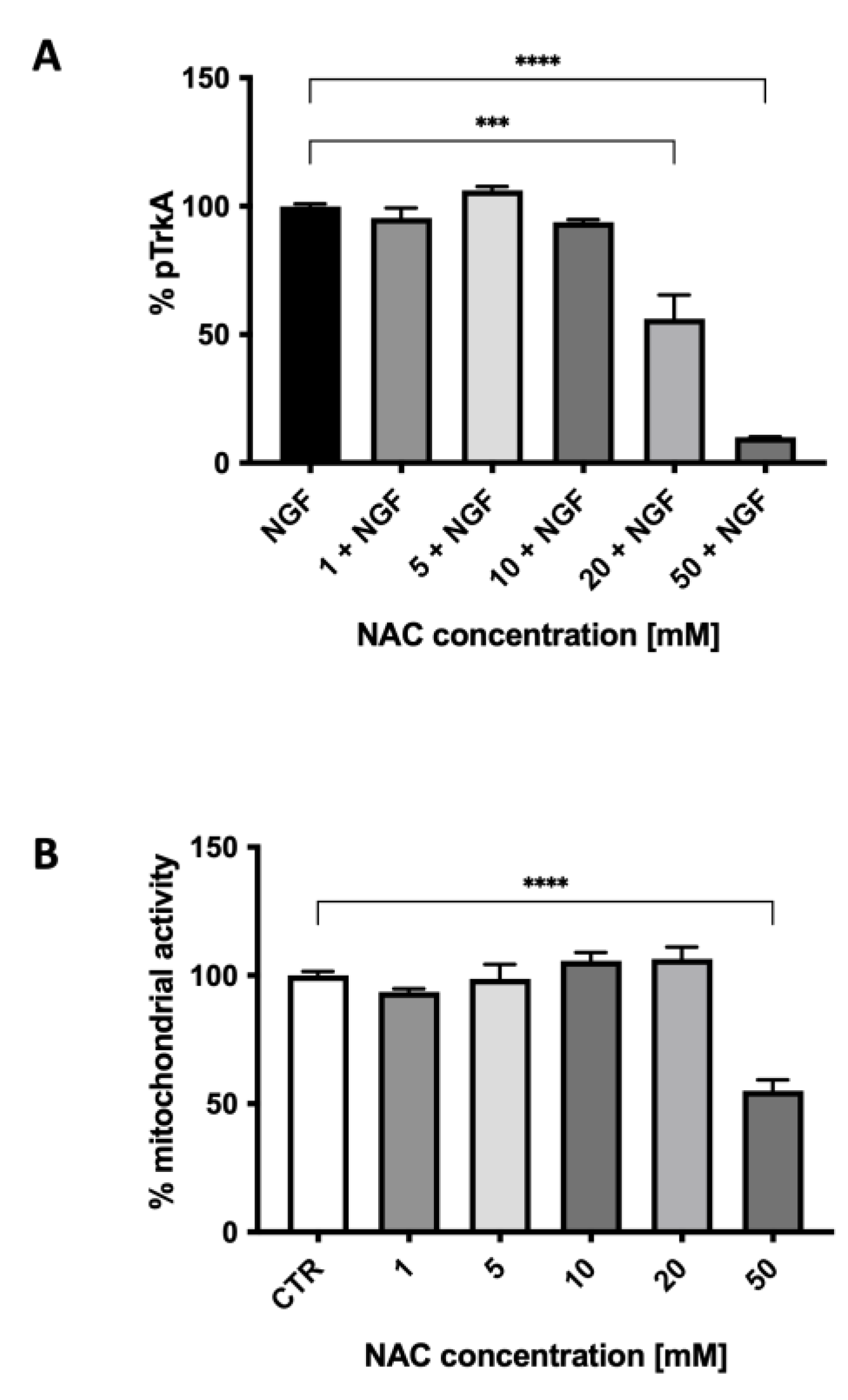
2.2. In Vitro Studies in Cultured Neuroblastoma Cells
3. Discussion
4. Materials and Methods
4.1. In Silico Studies
4.2. In Vitro Studies
4.3. MTT Assay
4.4. ELISA Assay
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shang, X.; Wang, Z.; Tao, H. Mechanism and Therapeutic Effectiveness of Nerve Growth Factor in Osteoarthritis Pain. Ther. Clin. Risk Manag. 2017, 13, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Denk, F.; Bennett, D.L.; McMahon, S.B. Nerve Growth Factor and Pain Mechanisms. Annu. Rev. Neurosci. 2017, 40, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Smith, M.T. Neurotrophins and Neuropathic Pain: Role in Pathobiology. Molecules 2015, 20, 10657–10688. [Google Scholar] [CrossRef] [PubMed]
- Mizumura, K.; Murase, S. Role of Nerve Growth Factor in Pain. Handb. Exp. Pharmacol. 2015, 227, 57–77. [Google Scholar] [CrossRef] [PubMed]
- Testa, G.; Cattaneo, A.; Capsoni, S. Understanding Pain Perception through Genetic Painlessness Diseases: The Role of NGF and ProNGF. Pharmacol. Res. 2021, 169, 105662. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Sung, K.; Xu, W.; Rodriguez, M.J.; Wu, A.C.; Santos, S.A.; Fang, S.; Uber, R.K.; Dong, S.X.; Guillory, B.C.; et al. A Missense Point Mutation in Nerve Growth Factor (NGFR100W) Results in Selective Peripheral Sensory Neuropathy. Prog. Neurobiol. 2020, 194, 101886. [Google Scholar] [CrossRef]
- Masuoka, T.; Yamashita, Y.; Yoshida, J.; Nakano, K.; Tawa, M.; Nishio, M.; Ishibashi, T. Sensitization of Glutamate Receptor-Mediated Pain Behaviour via Nerve Growth Factor-Dependent Phosphorylation of Transient Receptor Potential V1 under Inflammatory Conditions. Br. J. Pharmacol. 2020, 177, 4223–4241. [Google Scholar] [CrossRef]
- Sousa-Valente, J.; Calvo, L.; Vacca, V.; Simeoli, R.; Arévalo, J.C.; Malcangio, M. Role of TrkA Signalling and Mast Cells in the Initiation of Osteoarthritis Pain in the Monoiodoacetate Model. Osteoarthr. Cartil. 2018, 26, 84–94. [Google Scholar] [CrossRef]
- Enomoto, M.; Mantyh, P.W.; Murrell, J.; Innes, J.F.; Lascelles, B.D.X. Anti-Nerve Growth Factor Monoclonal Antibodies for the Control of Pain in Dogs and Cats. Vet. Rec. 2019, 184, 23. [Google Scholar] [CrossRef]
- Webb, M.P.; Helander, E.M.; Menard, B.L.; Urman, R.D.; Kaye, A.D. Tanezumab: A Selective Humanized MAb for Chronic Lower Back Pain. Ther. Clin. Risk Manag. 2018, 14, 361–367. [Google Scholar] [CrossRef]
- Norman, B.H.; McDermott, J.S. Targeting the Nerve Growth Factor (NGF) Pathway in Drug Discovery. Potential Applications to New Therapies for Chronic Pain. J. Med. Chem. 2017, 60, 66–88. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.J.; Frigotto, L.; Smith, M.E.; Baar, C.; Ivanova-Berndt, G.; Jaulent, A.; Stace, C.; Ullman, C.G.; Hine, A.V. A C-Terminal Cysteine Residue Is Required for Peptide-Based Inhibition of the NGF/TrkA Interaction at NM Concentrations: Implications for Peptide-Based Analgesics. Sci. Rep. 2019, 9, 930. [Google Scholar] [CrossRef] [PubMed]
- Fantucci, P.; Govoni, S.; Vanoli, E. Peptides Having TrkA-Receptor-Agonistic Activity or Having NGF-Antagonistic Activity. WO2016146730A1, 22 September 2016. [Google Scholar]
- Schwalfenberg, G.K. N-Acetylcysteine: A Review of Clinical Usefulness (an Old Drug with New Tricks). J. Nutr. Metab. 2021, 9, 9949453. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N- cetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, N.; Govoni, S.; Allegri, M. Non-drug Pain Relievers Active on Non-opioid Pain Mechanisms. Pain. Pract. 2022, 22, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Borrelli, K.W.; Greenwood, J.R.; Day, T.; Abel, R.; Farid, R.S.; Harder, E. Docking covalent inhibitors: A parameter free approach to pose prediction and scoring. J. Chem. Inf. Model. 2014, 54, 1932–1940. [Google Scholar] [CrossRef] [PubMed]
- Notartomaso, S.; Scarselli, P.; Mascio, G.; Liberatore, F.; Mazzon, E.; Mammana, S.; Gugliandolo, A.; Cruccu, G.; Bruno, V.; Nicoletti, F.; et al. N-Acetylcysteine causes analgesia in a mouse model of painful diabetic neuropathy. Mol. Pain. 2020, 16, 1744806920904292. [Google Scholar] [CrossRef]
- Wilson, S.H.; Sirianni, J.M.; Bridges, K.H.; Wolf, B.J.; Valente, I.E.; Scofield, M.D. The impact of intraoperative N-acetylcysteine on opioid consumption following spine surgery: A randomized pilot trial. Pain Manag. 2023, 13, 593–602. [Google Scholar] [CrossRef]
- Mohiuddin, M.; Pivetta, B.; Gilron, I.; Khan, J.S. Efficacy and Safety of N-Acetylcysteine for the Management of Chronic Pain in Adults: A Systematic Review and Meta-Analysis. Pain. Med. 2021, 22, 2896–2907. [Google Scholar] [CrossRef]
- Anastasi, E.; Scaramuzzino, S.; Viscardi, M.F.; Viggiani, V.; Piccioni, M.G.; Cacciamani, L.; Merlino, L.; Angeloni, A.; Muzii, L.; Porpora, M.G. Efficacy of N-Acetylcysteine on Endometriosis-Related Pain, Size Reduction of Ovarian Endometriomas, and Fertility Outcomes. Int. J. Environ. Res. Public. Health 2023, 20, 4686. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhang, J.; Liu, Y.; Ma, H.; Cao, F.; Xu, J.; Hou, Y.; Xu, L. The combination effects of acetaminophen and N-acetylcysteine on cytokines production and NF-κB activation of lipopolysaccharide-challenged piglet mononuclear phagocytes in vitro and in vivo. Vet. Immunol. Immunopathol. 2013, 152, 381–388. [Google Scholar] [CrossRef]
- Parisien, M.; Lima, L.V.; Dagostino, C.; El-Hachem, N.; Drury, G.L.; Grant, A.V.; Huising, J.; Verma, V.; Meloto, C.B.; Silva, J.R.; et al. Acute inflammatory response via neutrophil activation protects against the development of chronic pain. Sci. Transl. Med. 2022, 11, eabj9954. [Google Scholar] [CrossRef] [PubMed]
- Fantucci, P.; Govoni, S.; Pascale, A.; Marchesi, N.; Allegri, M.; Vanoli, E.; Calvillo, L.; Reggiani, P.; Pace, V. N-Acetyl Cysteine for Neuraxial Use as a TrkA Tyrosine Kinase Receptor Inhibitor for the Treatment of Acute and Chronic Pain. WO2022223590A1, 22 July 2022. [Google Scholar]
- Hochberg, M.C.; Tive, L.A.; Abramson, S.B.; Vignon, E.; Verburg, K.M.; West, C.R.; Smith, M.D.; Hungerford, D.S. When Is Osteonecrosis Not Osteonecrosis?: Adjudication of Reported Serious Adverse Joint Events in the Tanezumab Clinical Development Program. Arthritis Rheumatol. 2016, 68, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein-Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef]
- Tubert-Brohman, I.; Sherman, W.; Repasky, M.; Beuming, T. Improved docking of polypeptides with Glide. J. Chem. Inf. Model. 2013, 53, 1689–1699. [Google Scholar] [CrossRef]
- Banks, J.L.; Beard, H.S.; Cao, Y.; Cho, A.E.; Damm, W.; Farid, R.; Felts, A.K.; Halgren, T.A.; Mainz, D.T.; Maple, J.R.; et al. Integrated Modeling Program, Applied Chemical Theory (IMPACT). J. Comp. Chem. 2005, 26, 1752. [Google Scholar] [CrossRef]
- Lyne, P.D.; Lamb, M.L.; Saeh, J.C. Accurate prediction of the relative potencies of members of a series of kinase inhibitors using molecular docking and MM-GBSA scoring. J. Med. Chem. 2006, 49, 4805–4808. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Notes and Reference |
|---|---|
| Diabetic neuropathy | In a mouse model [18] |
| Postoperative opioid consumption | Clinical trial in patients (Clinical Trial Registration: NCT04562597) [19] |
| Chronic pain | NAC in chronic pain management [20] |
| Endometriosis | Efficacy of N-Acetylcysteine on endometriosis [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govoni, S.; Fantucci, P.; Marchesi, N.; Vertemara, J.; Pascale, A.; Allegri, M.; Calvillo, L.; Vanoli, E. N-Acetylcysteine Antagonizes NGF Activation of TrkA through Disulfide Bridge Interaction, an Effect Which May Contribute to Its Analgesic Activity. Int. J. Mol. Sci. 2024, 25, 206. https://doi.org/10.3390/ijms25010206
Govoni S, Fantucci P, Marchesi N, Vertemara J, Pascale A, Allegri M, Calvillo L, Vanoli E. N-Acetylcysteine Antagonizes NGF Activation of TrkA through Disulfide Bridge Interaction, an Effect Which May Contribute to Its Analgesic Activity. International Journal of Molecular Sciences. 2024; 25(1):206. https://doi.org/10.3390/ijms25010206
Chicago/Turabian StyleGovoni, Stefano, Piercarlo Fantucci, Nicoletta Marchesi, Jacopo Vertemara, Alessia Pascale, Massimo Allegri, Laura Calvillo, and Emilio Vanoli. 2024. "N-Acetylcysteine Antagonizes NGF Activation of TrkA through Disulfide Bridge Interaction, an Effect Which May Contribute to Its Analgesic Activity" International Journal of Molecular Sciences 25, no. 1: 206. https://doi.org/10.3390/ijms25010206
APA StyleGovoni, S., Fantucci, P., Marchesi, N., Vertemara, J., Pascale, A., Allegri, M., Calvillo, L., & Vanoli, E. (2024). N-Acetylcysteine Antagonizes NGF Activation of TrkA through Disulfide Bridge Interaction, an Effect Which May Contribute to Its Analgesic Activity. International Journal of Molecular Sciences, 25(1), 206. https://doi.org/10.3390/ijms25010206