
A Chimeric IL-15/IL-15Rα Molecule Expressed on NFκB-Activated Dendritic Cells Supports Their Capability to Activate Natural Killer Cells †

 , ,
, ,  and
and {kind=link}
{kind=link}
Abstract
:1. Introduction
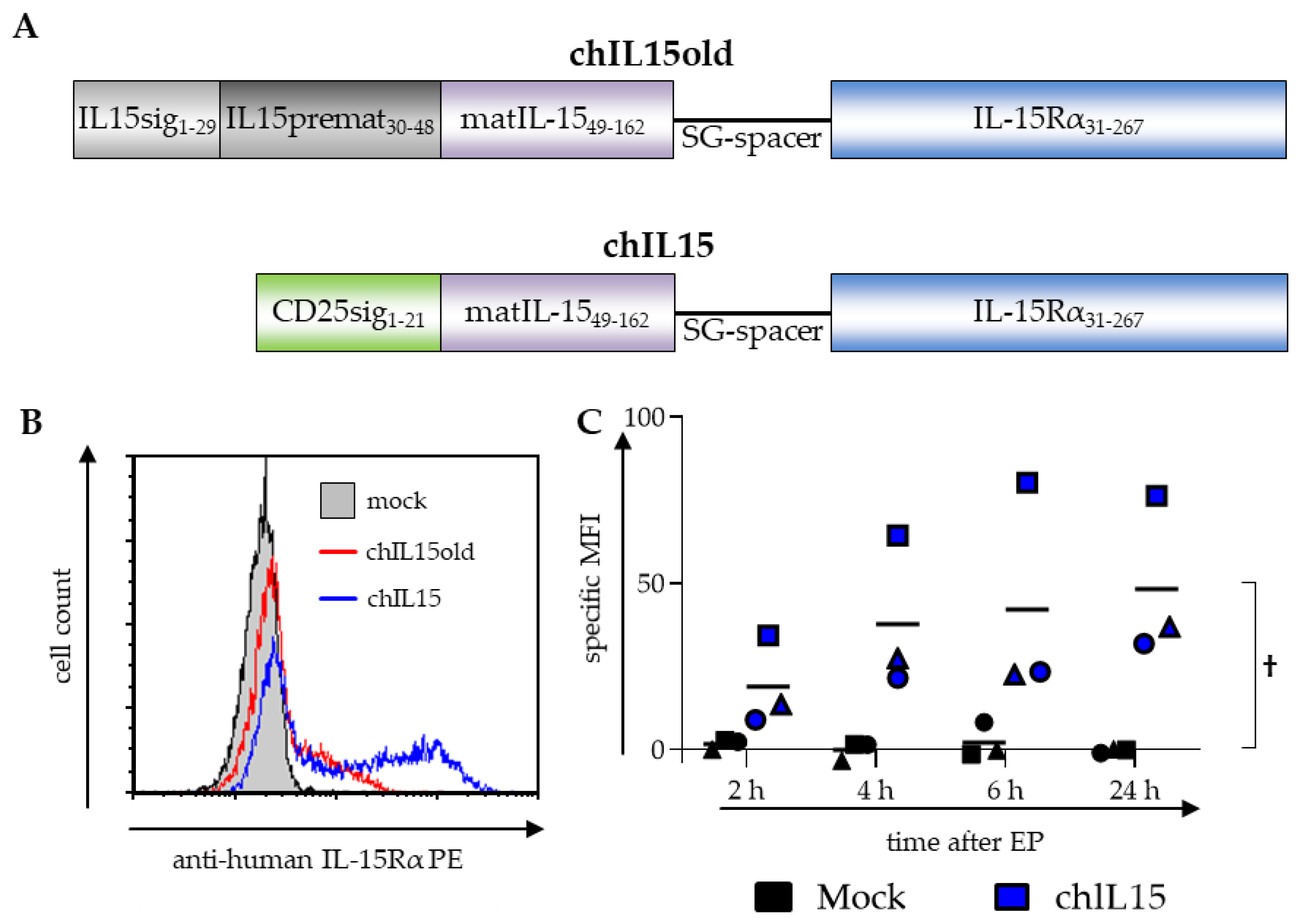
2. Results
2.1. chIL15 Is Expressed on chIL15-mRNA-electroporated DCs
2.2. Stimulation with DCs Transfected with a Combination of caIKKβ and chIL15 Leads to an Enhanced Activation of NK Cells
3. Discussion
4. Materials and Methods
4.1. Acquisition of Primary Cells
4.2. RNA Constructs and Electroporation of DCs
4.3. moDCs and PBMCs Co-Cultures
4.4. Analysis of Marker Expression on the Cell Surface
4.5. Measurement of Cytokine Secretion
4.6. Cytotoxicity Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schuler, G. Dendritic cells in cancer immunotherapy. Eur. J. Immunol. 2010, 40, 2123–2130. [Google Scholar] [CrossRef]
- Terme, M.; Ullrich, E.; Delahaye, N.F.; Chaput, N.; Zitvogel, L. Natural killer cell-directed therapies: Moving from unexpected results to successful strategies. Nat. Immunol. 2008, 9, 486–494. [Google Scholar] [CrossRef]
- Moretta, A. Natural killer cells and dendritic cells: Rendezvous in abused tissues. Nat. Rev. Immunol. 2002, 2, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Lion, E.; Smits, E.L.; Berneman, Z.N.; Van Tendeloo, V.F. NK cells: Key to success of DC-based cancer vaccines? Oncologist 2012, 17, 1256–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romani, N.; Gruner, S.; Brang, D.; Kampgen, E.; Lenz, A.; Trockenbacher, B.; Konwalinka, G.; Fritsch, P.O.; Steinman, R.M.; Schuler, G. Proliferating dendritic cell progenitors in human blood. J. Exp. Med. 1994, 180, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonuleit, H.; Kuhn, U.; Muller, G.; Steinbrink, K.; Paragnik, L.; Schmitt, E.; Knop, J.; Enk, A.H. Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. Eur. J. Immunol. 1997, 27, 3135–3142. [Google Scholar] [CrossRef]
- Mailliard, R.B.; Wankowicz-Kalinska, A.; Cai, Q.; Wesa, A.; Hilkens, C.M.; Kapsenberg, M.L.; Kirkwood, J.M.; Storkus, W.J.; Kalinski, P. alpha-type-1 polarized dendritic cells: A novel immunization tool with optimized CTL-inducing activity. Cancer Res. 2004, 64, 5934–5937. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.D.; Coulie, P.G.; Van den Eynde, B.J.; Agostinis, P. Integrating Next-Generation Dendritic Cell Vaccines into the Current Cancer Immunotherapy Landscape. Trends Immunol. 2017, 38, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, I.A.; Hoyer, S.; Gerer, K.F.; Voll, R.E.; Knippertz, I.; Guckel, E.; Schuler, G.; Schaft, N.; Dorrie, J. Triggering of NF-kappaB in cytokine-matured human DCs generates superior DCs for T-cell priming in cancer immunotherapy. Eur. J. Immunol. 2014, 44, 3413–3428. [Google Scholar] [CrossRef]
- Gerer, K.F.; Erdmann, M.; Hadrup, S.R.; Lyngaa, R.; Martin, L.M.; Voll, R.E.; Schuler-Thurner, B.; Schuler, G.; Schaft, N.; Hoyer, S.; et al. Preclinical evaluation of NF-kappaB-triggered dendritic cells expressing the viral oncogenic driver of Merkel cell carcinoma for therapeutic vaccination. Ther. Adv. Med. Oncol. 2017, 9, 451–464. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.P.; Trinchieri, G. Interleukin-12 in anti-tumor immunity and immunotherapy. Cytokine Growth Factor Rev. 2002, 13, 155–168. [Google Scholar] [CrossRef]
- Yu, Y.; Hagihara, M.; Ando, K.; Gansuvd, B.; Matsuzawa, H.; Tsuchiya, T.; Ueda, Y.; Inoue, H.; Hotta, T.; Kato, S. Enhancement of human cord blood CD34+ cell-derived NK cell cytotoxicity by dendritic cells. J. Immunol. 2001, 166, 1590–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, N.C.; Voll, R.E.; Voskens, C.J.; Gross, S.; Seliger, B.; Schuler, G.; Schaft, N.; Dörrie, J. NF-kappaB activation triggers NK-cell stimulation by monocyte-derived dendritic cells. Ther. Adv. Med Oncol. 2019, 11, 1758835919891622. [Google Scholar] [CrossRef]
- Kennedy, M.K.; Glaccum, M.; Brown, S.N.; Butz, E.A.; Viney, J.L.; Embers, M.; Matsuki, N.; Charrier, K.; Sedger, L.; Willis, C.R.; et al. Reversible defects in natural killer and memory CD8 T cell lineages in interleukin 15-deficient mice. J. Exp. Med 2000, 191, 771–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stonier, S.W.; Schluns, K.S. Trans-presentation: A novel mechanism regulating IL-15 delivery and responses. Immunol. Lett. 2010, 127, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Bergamaschi, C.; Jalah, R.; Kulkarni, V.; Rosati, M.; Zhang, G.M.; Alicea, C.; Zolotukhin, A.S.; Felber, B.K.; Pavlakis, G.N. Secretion and biological activity of short signal peptide IL-15 is chaperoned by IL-15 receptor alpha in vivo. J. Immunol. 2009, 183, 3064–3072. [Google Scholar] [CrossRef]
- Duitman, E.H.; Orinska, Z.; Bulanova, E.; Paus, R.; Bulfone-Paus, S. How a cytokine is chaperoned through the secretory pathway by complexing with its own receptor: Lessons from interleukin-15 (IL-15)/IL-15 receptor alpha. Mol. Cell. Biol. 2008, 28, 4851–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bergh, J.; Willemen, Y.; Lion, E.; Van, A.H.; De, R.H.; Anguille, S.; Goossens, H.; Berneman, Z.; Van, T.V.; Smits, E. Transpresentation of interleukin-15 by IL-15/IL-15Ralpha mRNA-engineered human dendritic cells boosts antitumoral natural killer cell activity. Oncotarget 2015, 6, 44123–44133. [Google Scholar] [CrossRef] [Green Version]
- Hasan, A.N.; Selvakumar, A.; Shabrova, E.; Liu, X.R.; Afridi, F.; Heller, G.; Riviere, I.; Sadelain, M.; Dupont, B.; O’Reilly, R.J. Soluble and membrane bound IL-15 Ralpha/IL-15 complexes mediate proliferation of high avidity central memory CD8+ T-cells for adoptive immunotherapy of cancer and infections. Clin. Exp. Immunol. 2016, 186, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Stoklasek, T.A.; Schluns, K.S.; Lefrancois, L. Combined IL-15/IL-15Ralpha immunotherapy maximizes IL-15 activity in vivo. J. Immunol. 2006, 177, 6072–6080. [Google Scholar] [CrossRef] [Green Version]
- Berger, C.; Berger, M.; Hackman, R.C.; Gough, M.; Elliott, C.; Jensen, M.C.; Riddell, S.R. Safety and immunologic effects of IL-15 administration in nonhuman primates. Blood 2009, 114, 2417–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortier, E.; Advincula, R.; Kim, L.; Chmura, S.; Barrera, J.; Reizis, B.; Malynn, B.A.; Ma, A. Macrophage- and dendritic-cell-derived interleukin-15 receptor alpha supports homeostasis of distinct CD8+ T cell subsets. Immunity 2009, 31, 811–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldmann, T.A.; Tagaya, Y. The multifaceted regulation of interleukin-15 expression and the role of this cytokine in NK cell differentiation and host response to intracellular pathogens. Annu. Rev. Immunol. 1999, 17, 19–49. [Google Scholar] [CrossRef] [Green Version]
- Clausen, J.; Vergeiner, B.; Enk, M.; Petzer, A.L.; Gastl, G.; Gunsilius, E. Functional significance of the activation-associated receptors CD25 and CD69 on human NK-cells and NK-like T-cells. Immunobiology 2003, 207, 85–93. [Google Scholar] [CrossRef]
- Robertson, M.J.; Caligiuri, M.A.; Manley, T.J.; Levine, H.; Ritz, J. Human natural killer cell adhesion molecules. Differential expression after activation and participation in cytolysis. J. Immunol. 1990, 145, 3194–3201. [Google Scholar]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Sun, S.; Hwang, I.; Tough, D.F.; Sprent, J. Potent and selective stimulation of memory-phenotype CD8+ T cells in vivo by IL-15. Immunity 1998, 8, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Goldrath, A.W.; Sivakumar, P.V.; Glaccum, M.; Kennedy, M.K.; Bevan, M.J.; Benoist, C.; Mathis, D.; Butz, E.A. Cytokine requirements for acute and Basal homeostatic proliferation of naive and memory CD8+ T cells. J. Exp. Med. 2002, 195, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Cheever, M.A. Twelve immunotherapy drugs that could cure cancers. Immunol. Rev. 2008, 222, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.; Usiskin, I.M.; Bergamaschi, C.; Hanlon, D.J.; Edelson, R.L.; Justesen, S.; Pavlakis, G.N.; Flavell, R.A.; Fahmy, T.M. Configuration-dependent Presentation of Multivalent IL-15:IL-15Ralpha Enhances the Antigen-specific T Cell Response and Anti-tumor Immunity. J. Biol. Chem. 2016, 291, 8931–8950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehniger, T.A.; Caligiuri, M.A. Interleukin 15: Biology and relevance to human disease. Blood 2001, 97, 14–32. [Google Scholar] [CrossRef]
- Sato, N.; Sabzevari, H.; Fu, S.; Ju, W.; Petrus, M.N.; Bamford, R.N.; Waldmann, T.A.; Tagaya, Y. Development of an IL-15-autocrine CD8 T-cell leukemia in IL-15-transgenic mice requires the cis expression of IL-15Ralpha. Blood 2011, 117, 4032–4040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dörrie, J.; Schaft, N.; Schuler, G.; Schuler-Thurner, B. Therapeutic Cancer Vaccination with Ex Vivo RNA-Transfected Dendritic Cells-An Update. Pharmaceutics 2020, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souri, Z.; Wierenga, A.P.A.; Mulder, A.; Jochemsen, A.G.; Jager, M.J. HLA Expression in Uveal Melanoma: An Indicator of Malignancy and a Modifiable Immunological Target. Cancers (Basel) 2019, 11, 1132. [Google Scholar] [CrossRef] [Green Version]
- Hazini, A.; Fisher, K.; Seymour, L. Deregulation of HLA-I in cancer and its central importance for immunotherapy. J. Immunother. Cancer 2021, 9, e002899. [Google Scholar] [CrossRef]
- Schaft, N.; Dorrie, J.; Thumann, P.; Beck, V.E.; Muller, I.; Schultz, E.S.; Kampgen, E.; Dieckmann, D.; Schuler, G. Generation of an optimized polyvalent monocyte-derived dendritic cell vaccine by transfecting defined RNAs after rather than before maturation. J. Immunol. 2005, 174, 3087–3097. [Google Scholar] [CrossRef] [Green Version]
- Gerer, K.F.; Hoyer, S.; Dorrie, J.; Schaft, N. Electroporation of mRNA as Universal Technology Platform to Transfect a Variety of Primary Cells with Antigens and Functional Proteins. Methods Mol. Biol. 2017, 1499, 165–178. [Google Scholar] [CrossRef]
- Schaft, N.; Dorrie, J.; Muller, I.; Beck, V.; Baumann, S.; Schunder, T.; Kampgen, E.; Schuler, G. A new way to generate cytolytic tumor-specific T cells: Electroporation of RNA coding for a T cell receptor into T lymphocytes. Cancer Immunol. Immunother. 2006, 55, 1132–1141. [Google Scholar] [CrossRef]
- BD FACSDiva, version 8.0.1; Becton, Dickinson and Company: San Jose, CA, USA, 2013.
- FCS Express 5 Flow Cytometry, version 5.01.0082; De Novo Software: Glendale, CA, USA, 2014.
- Hofflin, S.; Prommersberger, S.; Uslu, U.; Schuler, G.; Schmidt, C.W.; Lennerz, V.; Dorrie, J.; Schaft, N. Generation of CD8(+) T cells expressing two additional T-cell receptors (TETARs) for personalised melanoma therapy. Cancer Biol. Ther. 2015, 16, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Muzyka, O.; Tarkanii, O.; Varchenko, K.; Balantsev, A.; Shved, O. Prism, version 8.3.0 (538); GraphPad Software, LLC: San Diego, CA, USA, 2019. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosch, N.C.; Martin, L.-M.; Voskens, C.J.; Berking, C.; Seliger, B.; Schuler, G.; Schaft, N.; Dörrie, J. A Chimeric IL-15/IL-15Rα Molecule Expressed on NFκB-Activated Dendritic Cells Supports Their Capability to Activate Natural Killer Cells. Int. J. Mol. Sci. 2021, 22, 10227. https://doi.org/10.3390/ijms221910227
Bosch NC, Martin L-M, Voskens CJ, Berking C, Seliger B, Schuler G, Schaft N, Dörrie J. A Chimeric IL-15/IL-15Rα Molecule Expressed on NFκB-Activated Dendritic Cells Supports Their Capability to Activate Natural Killer Cells. International Journal of Molecular Sciences. 2021; 22(19):10227. https://doi.org/10.3390/ijms221910227
Chicago/Turabian StyleBosch, Naomi C., Lena-Marie Martin, Caroline J. Voskens, Carola Berking, Barbara Seliger, Gerold Schuler, Niels Schaft, and Jan Dörrie. 2021. "A Chimeric IL-15/IL-15Rα Molecule Expressed on NFκB-Activated Dendritic Cells Supports Their Capability to Activate Natural Killer Cells" International Journal of Molecular Sciences 22, no. 19: 10227. https://doi.org/10.3390/ijms221910227
APA StyleBosch, N. C., Martin, L.-M., Voskens, C. J., Berking, C., Seliger, B., Schuler, G., Schaft, N., & Dörrie, J. (2021). A Chimeric IL-15/IL-15Rα Molecule Expressed on NFκB-Activated Dendritic Cells Supports Their Capability to Activate Natural Killer Cells. International Journal of Molecular Sciences, 22(19), 10227. https://doi.org/10.3390/ijms221910227