
Plasticity-Enhancing Effects of Levodopa Treatment after Stroke


Abstract
:
1. Introduction
2. Results
2.1. Increased Dopaminergic Innervation in the Ischemic Territory Following Experimental Stroke
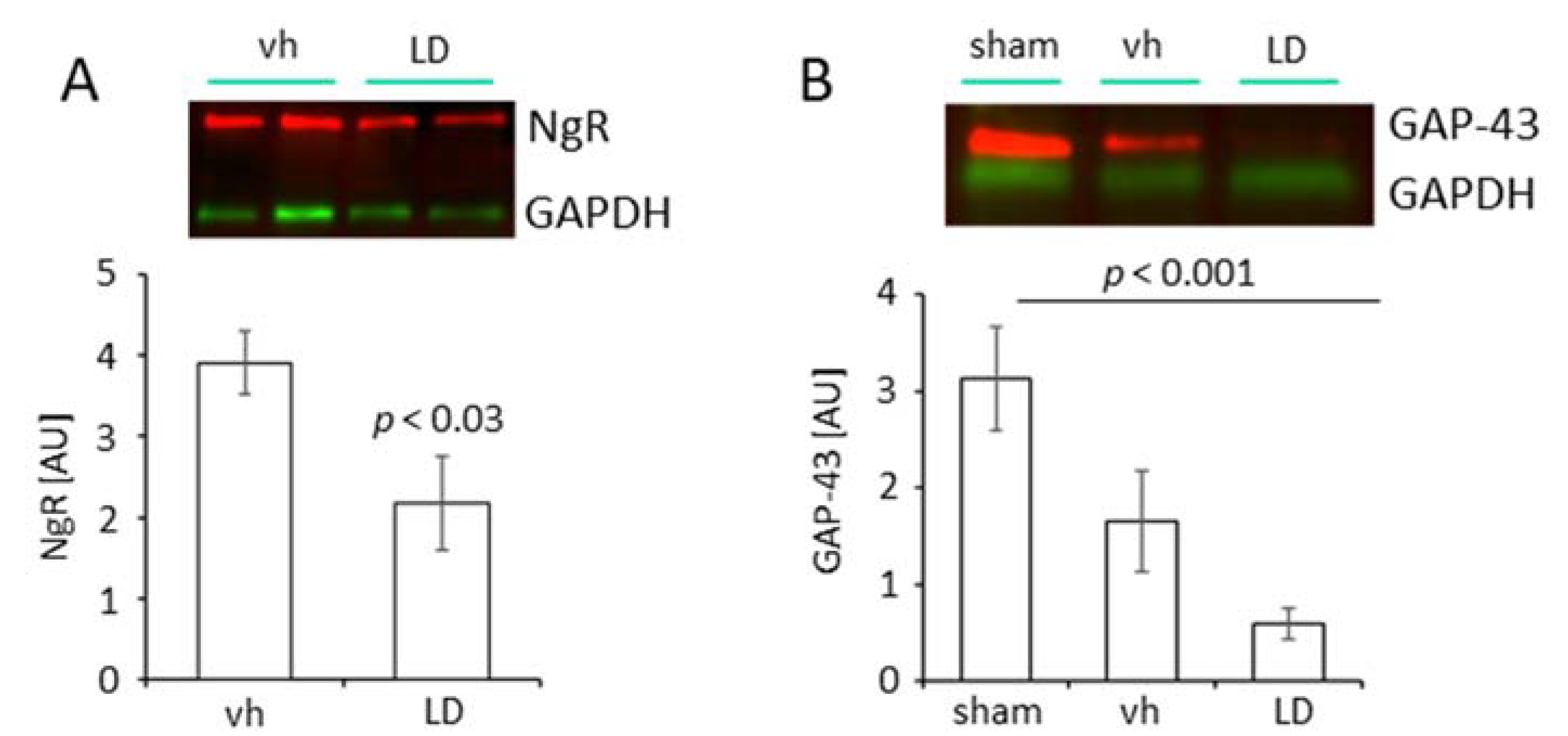
2.2. Treatment with Levodopa Reduces the Number of Nogo A+ Mature Oligodendrocytes in the Peri-Infarct Area
2.3. Increased Number of Adult Oligodendrocyte Precursor Cells in the Peri-Infarct Area of Rats Treated with Levodopa Following tMCAO
3. Discussion
3.1. Increased Dopaminergic Innervation in the Ischemic Territory
3.2. Effects of Levodopa Treatment on Nogo A Pathway and GAP 43
3.3. Levodopa Treatment and Oligodendrogenesis
4. Materials and Methods
4.1. Experimental Design
4.2. Transient/Permanent Occlusion of the Middle Cerebral Artery (tMCAO/pMCAO) and Photothrombosis (PT)
4.3. Western Blots
4.4. Immunofluorescence/Immunohistochemistry
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hermann, D.M.; Chopp, M. Promoting brain remodelling and plasticity for stroke recovery: Therapeutic promise and potential pitfalls of clinical translation. Lancet. Neurol. 2012, 11, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Cramer, S.C. Repairing the human brain after stroke: I. Mechanisms of spontaneous recovery. Ann. Neurol. 2008, 63, 272–287. [Google Scholar] [CrossRef]
- Quattromani, M.J.; Hakon, J.; Rauch, U.; Bauer, A.Q.; Wieloch, T. Changes in resting-state functional connectivity after stroke in a mouse brain lacking extracellular matrix components. Neurobiol. Dis. 2018, 112, 91–105. [Google Scholar] [CrossRef]
- Ruscher, K.; Shamloo, M.; Rickhag, M.; Ladunga, I.; Soriano, L.; Gisselsson, L.; Toresson, H.; Ruslim-Litrus, L.; Oksenberg, D.; Urfer, R.; et al. The sigma-1 receptor enhances brain plasticity and functional recovery after experimental stroke. Brain 2011, 134, 732–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, M.; Ninomiya, I.; Kanazawa, M. Angiogenesis and neuronal remodeling after ischemic stroke. Neural Regen. Res. 2020, 15, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.E.; Strittmatter, S.M. Nogo limits neural plasticity and recovery from injury. Curr. Opin. Neurobiol. 2014, 27, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Nyatia, E.; Lang, D.M.M. Localisation and expression of a myelin associated neurite inhibitor, Nogo-A and its receptor Nogo-receptor by mammalian CNS cells. Res. Vet. Sci. 2007, 83, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Pernet, V.; Schwab, M.E. The role of Nogo-A in axonal plasticity, regrowth and repair. Cell Tissue Res. 2012, 349, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.B.; Weinmann, O.; Brösamle, C.; Oertle, T.; Schwab, M.E. Patterns of Nogo mRNA and Protein Expression in the Developing and Adult Rat and after CNS Lesions. J. Neurosci. 2002, 22, 3553–3567. [Google Scholar] [CrossRef] [Green Version]
- Sandvig, A.; Berry, M.; Barrett, L.B.; Butt, A.; Logan, A. Myelin-, reactive glia-, and scar-derived CNS axon growth inhibitors: Expression, receptor signaling, and correlation with axon regeneration. Glia 2004, 46, 225–251. [Google Scholar] [CrossRef]
- Overman, J.J.; Carmichael, S.T. Plasticity in the injured brain: More than molecules matter. Neuroscientist 2014, 20, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Rust, R.; Grönnert, L.; Gantner, C.; Enzler, A.; Mulders, G.; Weber, R.Z.; Siewert, A.; Limasale, Y.D.P.; Meinhardt, A.; Maurer, M.A.; et al. Nogo-A targeted therapy promotes vascular repair and functional recovery following stroke. Proc. Natl. Acad. Sci. USA 2019, 116, 14270–14279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruscher, K.; Kuric, E.; Wieloch, T. Levodopa treatment improves functional recovery after experimental stroke. Stroke 2012, 43, 507–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häggman Henrikson, J.; Pombo Antunes, A.R.; Wieloch, T.; Ruscher, K. Enhanced functional recovery by levodopa is associated with decreased levels of synaptogyrin following stroke in aged mice. Brain Res. Bull. 2020, 155, 61–66. [Google Scholar] [CrossRef]
- Hosp, J.A.; Pekanovic, A.; Rioult-Pedotti, M.S.; Luft, A.R. Dopaminergic projections from midbrain to primary motor cortex mediate motor skill learning. J. Neurosci. 2011, 31, 2481–2487. [Google Scholar] [CrossRef] [Green Version]
- Kuric, E.; Wieloch, T.; Ruscher, K. Dopamine receptor activation increases glial cell line-derived neurotrophic factor in experimental stroke. Exp. Neurol. 2013, 247, 202–208. [Google Scholar] [CrossRef]
- Kuric, E.; Ruscher, K. Reduction of rat brain CD8+ T-cells by levodopa/benserazide treatment after experimental stroke. Eur. J. Neurosci. 2014, 40, 2463–2470. [Google Scholar] [CrossRef]
- Talhada, D.; Rabenstein, M.; Ruscher, K. The role of dopaminergic immune cell signalling in poststroke inflammation. Ther. Adv. Neurol. Disord. 2018, 11, 1756286418774225. [Google Scholar] [CrossRef] [Green Version]
- Acler, M.; Fiaschi, A.; Manganotti, P. Long-term levodopa admnistration in chronic stroke patients. A clinical and neurophysiologic single-blind placebo-controlled cross-over pilot study. Restor. Neurol. Neurosci. 2009, 27, 277–283. [Google Scholar] [CrossRef]
- Ford, G.A.; Bhakta, B.B.; Cozens, A.; Hartley, S.; Holloway, I.; Meads, D.; Pearn, J.; Ruddock, S.; Sackley, C.M.; Saloniki, E.C.; et al. Safety and efficacy of co-careldopa as an add-on therapy to occupational and physical therapy in patients after stroke (DARS): A randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2019, 18, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Rosin, C.; Colombo, S.; Calver, A.A.; Bates, T.E.; Skaper, S.D. Dopamine D2 and D3 receptor agonists limit oligodendrocyte injury caused by glutamate oxidative stress and oxygen/glucose deprivation. Glia 2005, 52, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Ruscher, K.; Johannesson, E.; Brugiere, E.; Erickson, A.; Rickhag, M.; Wieloch, T. Enriched environment reduces apolipoprotein E (ApoE) in reactive astrocytes and attenuates inflammation of the peri-infarct tissue after experimental stroke. J. Cereb. Blood Flow Metab. 2009, 29, 1796–1805. [Google Scholar] [CrossRef] [Green Version]
- Schmandke, A.; Mosberger, A.C.; Schmandke, A.; Celen, Z.; Schwab, M.E. The neurite growth inhibitory protein Nogo-A has diverse roles in adhesion and migration. Cell Adhes. Migr. 2013, 7, 451–454. [Google Scholar] [CrossRef] [Green Version]
- Wills, Z.P.; Mandel-Brehm, C.; Mardinly, A.R.; McCord, A.E.; Giger, R.J.; Greenberg, M.E. The nogo receptor family restricts synapse number in the developing hippocampus. Neuron 2012, 73, 466–481. [Google Scholar] [CrossRef] [Green Version]
- Puderbaugh, M.; Emmady, P.D. Neuroplasticity. In StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Burda, J.E.; Sofroniew, M.V. Reactive gliosis and the multicellular response to CNS damage and disease. Neuron 2014, 81, 229–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berretta, A.; Tzeng, Y.-C.; Clarkson, A.N. Post-stroke recovery: The role of activity-dependent release of brain-derived neurotrophic factor. Expert Rev. Neurother. 2014, 14, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Schäbitz, W.R.; Berger, C.; Kollmar, R.; Seitz, M.; Tanay, E.; Kiessling, M.; Schwab, S.; Sommer, C. Effect of Brain-Derived Neurotrophic Factor Treatment and Forced Arm Use on Functional Motor Recovery after Small Cortical Ischemia. Stroke 2004, 35, 992–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoonkitiwongsa, P.S.; Jackson-Friedman, C.; McMillan, P.J.; Schultz, R.L.; Lyden, P.D. Angiogenesis after stroke is correlated with increased numbers of macrophages: The clean-up hypothesis. J. Cereb. Blood Flow Metab. 2001, 21, 1223–1231. [Google Scholar] [CrossRef]
- Piltonen, M.; Planken, A.; Leskelä, O.; Myöhänen, T.T.; Hänninen, A.-L.; Auvinen, P.; Alitalo, K.; Andressoo, J.-O.; Saarma, M.; Männistö, P.T. Vascular endothelial growth factor C acts as a neurotrophic factor for dopamine neurons in vitro and in vivo. Neuroscience 2011, 192, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Izumi, Y.; Wakita, S.; Kanbara, C.; Nakai, T.; Akaike, A.; Kume, T. Integrin α5β1 expression on dopaminergic neurons is involved in dopaminergic neurite outgrowth on striatal neurons. Sci. Rep. 2017, 7, 42111. [Google Scholar] [CrossRef] [Green Version]
- Kuric, E.; Ruscher, K. Reversal of stroke induced lymphocytopenia by levodopa/benserazide treatment. J. Neuroimmunol. 2014, 269, 94–97. [Google Scholar] [CrossRef]
- Ohlin, K.E.; Francardo, V.; Lindgren, H.S.; Sillivan, S.E.; O’Sullivan, S.S.; Luksik, A.S.; Vassoler, F.M.; Lees, A.J.; Konradi, C.; Cenci, M.A. Vascular endothelial growth factor is upregulated by L-dopa in the parkinsonian brain: Implications for the development of dyskinesia. Brain 2011, 134, 2339–2357. [Google Scholar] [CrossRef] [Green Version]
- Lindau, N.T.; Banninger, B.J.; Gullo, M.; Good, N.A.; Bachmann, L.C.; Starkey, M.L.; Schwab, M.E. Rewiring of the corticospinal tract in the adult rat after unilateral stroke and anti-Nogo-A therapy. Brain 2014, 137, 739–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemmar, A.; Chen, C.-C.; Weinmann, O.; Kast, B.; Vajda, F.; Bozeman, J.; Isaad, N.; Zuo, Y.; Schwab, M.E. Oligodendrocyte- and Neuron-Specific Nogo-A Restrict Dendritic Branching and Spine Density in the Adult Mouse Motor Cortex. Cereb. Cortex 2018, 28, 2109–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbik, F.; Cafferty, W.B.J.; Strittmatter, S.M. Myelin associated inhibitors: A link between injury-induced and experience-dependent plasticity. Exp. Neurol. 2012, 235, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, D.; Coffin, R.S.; Anderson, P.N. The Nogo receptor, its ligands and axonal regeneration in the spinal cord; a review. J. Neurocytol. 2002, 31, 93–120. [Google Scholar] [CrossRef]
- Gopalakrishna, R.; Mades, A.; Oh, A.; Zhu, A.; Nguyen, J.; Lin, C.; Kindy, M.S.; Mack, W.J. Cyclic-AMP induces Nogo-A receptor NgR1 internalization and inhibits Nogo-A-mediated collapse of growth cone. Biochem. Biophys. Res. Commun. 2020, 523, 678–684. [Google Scholar] [CrossRef]
- Chung, D.; Shum, A.; Caraveo, G. GAP-43 and BASP1 in Axon Regeneration: Implications for the Treatment of Neurodegenerative Diseases. Front. Cell Dev. Biol. 2020, 8, 567537. [Google Scholar] [CrossRef]
- Sist, B.; Fouad, K.; Winship, I.R. Plasticity beyond peri-infarct cortex: Spinal up regulation of structural plasticity, neurotrophins, and inflammatory cytokines during recovery from cortical stroke. Exp. Neurol. 2014, 252, 47–56. [Google Scholar] [CrossRef]
- Gorup, D.; Bohaček, I.; Miličević, T.; Pochet, R.; Mitrečić, D.; Križ, J.; Gajović, S. Increased expression and colocalization of GAP43 and CASP3 after brain ischemic lesion in mouse. Neurosci. Lett. 2015, 597, 176–182. [Google Scholar] [CrossRef]
- Shah, A.M.; Ishizaka, S.; Cheng, M.Y.; Wang, E.H.; Bautista, A.R.; Levy, S.; Smerin, D.; Sun, G.; Steinberg, G.K. Optogenetic neuronal stimulation of the lateral cerebellar nucleus promotes persistent functional recovery after stroke. Sci. Rep. 2017, 7, 46612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khorchid, A.; Fragoso, G.; Shore, G.; Almazan, G. Catecholamine-induced oligodendrocyte cell death in culture is developmentally regulated and involves free radical generation and differential activation of caspase-3. Glia 2002, 40, 283–299. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.W.; Althomsons, S.P.; Hyrc, K.L.; Choi, D.W.; Goldberg, M.P. Oligodendrocytes from forebrain are highly vulnerable to AMPA/kainate receptor-mediated excitotoxicity. Nat. Med. 1998, 4, 291–297. [Google Scholar] [CrossRef]
- Ersland, K.M.; Skrede, S.; Stansberg, C.; Steen, V.M. Subchronic olanzapine exposure leads to increased expression of myelination-related genes in rat fronto-medial cortex. Transl. Psychiatry 2017, 7, 1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, F.; Konkalmatt, P.; Mokashi, C.; Kumar, M.; Zhang, Y.; Ko, A.; Farino, Z.J.; Asico, L.D.; Xu, G.; Gildea, J.; et al. Dopamine D(2) receptor modulates Wnt expression and control of cell proliferation. Sci. Rep. 2019, 9, 16861. [Google Scholar] [CrossRef] [PubMed]
- Talhada, D.; Feiteiro, J.; Costa, A.R.A.R.; Talhada, T.; Cairraõ, E.; Wieloch, T.; Englund, E.; Santos, C.R.C.R.; Goncąlves, I.; Ruscher, K.; et al. Triiodothyronine modulates neuronal plasticity mechanisms to enhance functional outcome after stroke. Acta Neuropathol. Commun. 2019, 7, 216. [Google Scholar] [CrossRef] [Green Version]
- Maten, G.; Henck, V.; Wieloch, T.; Ruscher, K. CX3C chemokine receptor 1 deficiency modulates microglia morphology but does not affect lesion size and short-term deficits after experimental stroke. BMC Neurosci. 2017, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, T.; Erickson, A.; Kuric, E.; Shamloo, M.; Hara-Nishimura, I.; Inácio, A.R.L.; Wieloch, T.; Ruscher, K. The asparaginyl endopeptidase legumain after experimental stroke. J. Cereb. Blood Flow Metab. 2010, 30, 1756–1766. [Google Scholar] [CrossRef]
- Madinier, A.; Wieloch, T.; Olsson, R.; Ruscher, K. Impact of estrogen receptor beta activation on functional recovery after experimental stroke. Behav. Brain Res. 2014, 261, 282–288. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Supplier/Cat# | Application/Dilution | Secondary Antibody/Dilution |
|---|---|---|---|
| Anti-Nogo A | R&D Systems, cat# AF3098 | IF, 1:100 | Anti-goat Cy3, 1:400 |
| WB, 1:500 | Anti-goat biotinylated, 1:4000 ; Biotin-HRP 1:3000 | ||
| Anti-Nogo Receptor | Millipore, cat# AB15138 | WB, 1:2000 | Anti-rabbit HRP, 1:15,000 |
| glutathione-S-transferase π | BD Transduction Laboratories, cat# 610719 | IF, 1:200 | Anti-mouse biotinylated 1:400; streptavidin AF488, 1:400 |
| (GST π) | |||
| Growth Associated Protein | Millipore, cat# MAB347 | WB, 1:1500 | Anti-mouse HRP, 1:10,000 |
| (GAP 43) | |||
| OLIG-2 | R&D Systems, cat# AF2418 | IHC, 1:200 | Anti-rabbit biotinylated, 1:400 |
| Tyrosine hydroxylase (TH) | Millipore, cat# AB152 | IHC, 1:1000 | Anti-rabbit biotinylated, 1:400 |
| WB, 1:5000 | Anti-rabbit HRP, 1:15,000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Talhada, D.; Marklund, N.; Wieloch, T.; Kuric, E.; Ruscher, K. Plasticity-Enhancing Effects of Levodopa Treatment after Stroke. Int. J. Mol. Sci. 2021, 22, 10226. https://doi.org/10.3390/ijms221910226
Talhada D, Marklund N, Wieloch T, Kuric E, Ruscher K. Plasticity-Enhancing Effects of Levodopa Treatment after Stroke. International Journal of Molecular Sciences. 2021; 22(19):10226. https://doi.org/10.3390/ijms221910226
Chicago/Turabian StyleTalhada, Daniela, Niklas Marklund, Tadeusz Wieloch, Enida Kuric, and Karsten Ruscher. 2021. "Plasticity-Enhancing Effects of Levodopa Treatment after Stroke" International Journal of Molecular Sciences 22, no. 19: 10226. https://doi.org/10.3390/ijms221910226
APA StyleTalhada, D., Marklund, N., Wieloch, T., Kuric, E., & Ruscher, K. (2021). Plasticity-Enhancing Effects of Levodopa Treatment after Stroke. International Journal of Molecular Sciences, 22(19), 10226. https://doi.org/10.3390/ijms221910226